zql-第五届全国动物生物技术分会学术年会参考word
第五届全国动物生物技术分会学术年会

Prof.George E.Seidel,Jr
Department of Biomedical Sciences,Animal Reproduction and Biotechnology Laboratory,Colorado State University,USA
Title:
吴珍芳 教授
华南农业大学
题目:木聚糖酶基因克隆及其在转基因育种上的研究
15:00—15:30
李子义 教授
吉林大学农学部畜牧兽医学院
题目:转基因克隆雪貂的研究进展
15:30—16:00
曾文先 教授
西北农林科技大学
题目:
16:00—16:10
茶歇
16:10—16:40
杨利国 教授
华中农业大学
题目:激素基因免疫研究进展
题目:禽类及中华鳖胚胎原始生殖细胞的迁移以及异种嵌合体的制备
16:00—16:10
茶歇
16:10—16:40
连正兴 教授
中国农业大学
题目:
16:40—17:10
欧阳红生
吉林大学
题目:抗猪瘟转基因猪研究
17:10—17:40
周欢敏 教授
内蒙古农业大学生物工程学院
题目:
17:40—18:10
学生
题目:
大会分组报告:功能基因与分子育种及重大疾病控制
The Division of Structural Biology,The Wellcome Trust Centre for Human Genetics,OxfordUniversity.
题目:Manipulation of Viral microRNAs for Novel Vaccines
第五届全国生物多样性保护与持续利用研讨会论文摘要

其他生物多样性保护与可持续利用从两难到双赢及其现实途径吕一河 傅伯杰 陈利顶中科院生态环境研究中心系统生态重点实验室北京 100085生物多样性保护与可持续利用是当今生物多样性科学研究的五大核心领域之一它与社会人文因素紧密相关保护区已经成为生物多样性保护与可持续利用的主导模式和试验场所本文以保护区为背景分析实践中存在的种种两难境地及其原因综述和评价了生物多样性保护与可持续利用协调统一实现双赢的可能途径包括生态旅游基于社区的保护CBC/基于社区的自然资源管理CBNRM综合保护与开发项目ICDP内部驱动项目IMP和生物多样性资源管理诸模式这些模式的成功运作将会对人类观念行为的改变产生积极的推动反过来又会促进生物多样性资源保护和可持续利用实践的顺利发展构成良性循环生物多样性资源保护成本和利用收益在不同利益集团和时空尺度上能否实现公平分配是影响各种模式成功与否的关键因素生物多样性理论在工业生态学中的应用陆宏芳 彭少麟(中国科学院华南植物研究所广州 510650)生物多样性理论作为经典生态学的关键性理论之一在工业生态学中亦有着广阔的应用空间目前生物多样性理论在工业生态学中的应用主要集中于物种多样性研究与生态系统多样性研究两个层次由于污染废弃物种类及各种类的内部组分均具有多样性的特征其净化回收并非任何单一物种的加环所能完成在工业生态系统废弃物回收再利用中的作用方面用物种多样性理论增加配搭由不同生物净化种类组成的生产增益环即可有效实现原有废弃物的有效回收再利用又可实现直接的经济价值同时由于污染物的多源性其净化回收实难在单一的工业生态系统中得以实现因而需要运用生态系统多样性理论从生态系统层面考虑从资源到生产到产品到流通消费到废弃物净化再生的全过程中的生态系统多样性调整与整合随着工业转型与工业生态学研究的不断深入生物多样性理论在工业生态中的应用领域必将同时向宏观的景观尺度和微观的基因尺度全面推进全面系统地推动工业生态学的发展四川都江堰地区生物多样性概况及其特点与意义庄 平 高贤明 冯正波 靳昌伟中科院植物所华西亚高山植物园 四川都江堰 611830本文将都江堰境内的生态系统分为5个类型即城镇生态系统农业生态系统森林生态系统高寒生态系统和河流生态系统区内拥有6个植被型24个群系当地植被垂直带谱由亚热带常绿阔叶林高山流石滩稀疏植被等7个植被垂直带构成并有河谷沼泽湖及冰斗湖苔藓园等特殊隐域生境和一些特有植被群落类型都江堰拥有高等植物3284种占我国高等植物种类的11.3%,展现了温性植物区系为主的基本特征同时特少单型属分别占我国相应属的16.5%13.6%和10.2%,国家与四川省级珍稀濒危保护植物种类繁多境内脊椎动物种类434种占我国脊椎动物种类的8.2%,其中中国脊椎动特有种及其各类群的相对比率均超过了我国相应特有动物种类的综合水平而且珍稀濒危动物种类极其丰富都江堰农牧资源遗传多样性也非常突出其中部分品种是不可多得的宝贵遗传资源都江堰生态系统的特点在于类型多样生境脆弱结构复杂互补依赖区位独特过渡性强条件优越地位重要物种多样性特点在于种类丰富成份交融特有性强珍稀种多显示度高遗传多样性特点在于品种繁多资源独特价值高作者认为都江堰及其临近地区丰富独特的生物多样性不但对于我国而且对于世界都具有难以替代的生态意义与价值当地生物多样性是都江堰古水利工程得以持续发挥效益的根本支撑并为道家天人合一的伦理思想的传播提供了天然的讲台因而对于我国的社会发展和世界文明具有深远影响生物多样性为我们提供了400亿元/年左右的经济价值为当地同期GDP的7.4倍尤其是间接经济价值的受益面已远远超过了都江堰行政区域范围对周边尤其是成都平原的经济发展具有不容忽视的意义和价值同时本文映证了都江堰所在的华西雨屏带是值得关注的生物多样性关键区域生物多样性的保护将始终是当地最根本和重要的工作任务天目山生物多样性可持续发展管理研究杨淑贞¹ 程爱兴²1. 天目山国家级自然保护区管理局浙江临安311311(2. 临安市林业局浙江临安311300要使天目山保护区的生物多样性真正走上可持续发展的道路就应针对目前保护区面临的主要问题和困难着重加强对原生性森林生态系统的保护控制人工毛竹林对天然林的蚕食趋势提高生物多样性的全面监测水平探索合理利用自然资源发展区内和周边社区经济等方面的管理工作确保天目山保护区的健康发展和生态经济社会效益的全面发挥民间植物分类系统及其用来进行区域性物种多样性快速评估的探讨王锦秀1 刘宏茂1 许再富1 傅德志21. 中国科学院西双版纳热带植物园昆明 6663032. 中国科学院植物研究所北京 100093本研究分别在西双版纳景洪市附近勐腊县的勐仑镇和象明乡选择了三个傣族村寨采用生物多样性快速编目Rapid Biodiversity Inventory的方法展开调查调查分普遍调查与样方调查两种4名科技人员和4名傣族村民分2组在相同的区域和样方内对植物进行鉴定比较同时采取访谈interview的方法分别在三个寨子的各种生境中随机抽取80种植物包括草本灌木乔木及藤本请不同年龄的村民进行辨认以了解不同年龄的傣族村民对植物认知知识的掌握程度研究结果显示依赖民间植物分类系统的原居民识别当地物种的数量和鉴定速度都明显高于受过专业训练的植物分类学家并且民间植物命名直接告诉我们物种的生态地理分布特征甚至作用等信息在某种程度上说明民间分类系统的信息存储量高于现代分类系统原居民识别当地物种的速度胜过受过专业训练的植物分类学家说明局部地区民间分类系统作为信息存取系统具有存取速率更快更有效的特点民间分类是建立在对自然界中明显的间断和相似性的识别以及对当地动植物物种形态和行为特征观察的基础上它有阶层结构其命名法就反映了这种分类系统的阶层结构而且很大程度上与西方生物分类系统相关因为民间命名的分类群是对类群的形态生态和行为特征的普遍描述所以是最不主观的因此很有可能从民间分类体系中寻求用于热带生物多样性快速评估的好方法西双版纳傣族也有自己的植物命名和分类系统他们对植物的定义为hong ebin同时把植物分为栽培和野生两大类他们对植物的命名也采用双名法前边一词代表植物的生活型或者经济型按生活型傣族把植物分为Mai木本Ya (草本)Hei (藤本)Gu (蕨类)和Man (块根)等类型按经济型分把植物分为Ma (果树)Pa (蔬菜)Kao(五谷)Tu豆类和Luo(花卉)等第二个词则进一步阐明植物的特征如形态颜色和气味等特性如生长特点和特殊性状用途生境和种源等本文通过调查傣族原居民对植物的识别程度以及与分类学家对植物鉴定的物种数量多少相比较探讨民间分类系统能否在地区性物种多样性快速评估中运用傣族民间分类系统用于物种多样性评估有如下优点识别率高识别速度快信息量大调查迅速评估结果可以直接反馈到原居民中能调动原居民的积极性参与到保护中来等民间分类系统当然也存在缺点如对文化价值大的物种分得很详细而对文化价值少的物种则分得粗一些其次少数分布较广的植物在不同地方会有不同地方名local name这使得物种多样性快速评估工作只能限于局部区域热带是世界上生物多样性最丰富的地区近年来热带森林面积锐减导致大量物种流失因此急需对生物多样性快速分析评估的有效方法物种多样性评估是生物多样性保护的基础民间植物分类和系统学知识在物种多样性保护中的作用应引起人们足够认识可以考虑将民间植物分类系统作为当前分类系统的辅助在热带地区的局部或社区水平上进行物种多样性快速评估利用世界银行贷款促进我国生物多样性研究的发展侯淑琴 田文忱中国科学院生物多样性委员会办公室北京 100864我国的生物多样性研究工作自1985年开始立项项目的目的是支持85期间中国科学院汪 松研究员陈灵芝研究员施立明院士张新时院士钱迎倩研究员洪德元院士分别主持的3个生物多样性方面的重大研究项目在生物多样性研究的三个层次上对中国的生物多样性开展全面而深入的研究工作以了解中国生物多样性的现状和濒危机制确定和保护持续利用的优先重点并提出有效的策略与措施建立适合中国国情的全国的生物多样性保护与持续利用的研究和信息管理体系加强公众的生物多样性保护意识提高政府在生物多样性保护与持续利用方面的决策能力中国科学院1989年开始酝酿成立生物多样性研究组1990年召开第一次生物多样性研讨会1992年正式成立中国科学院生物多样性委员会1994年中国科学院生物多样性委员会与国家环保局国家林业局教育部国家海洋局和农业部共同主办召开第一次全国生物多样性学术研讨会进行学术交流每二年召开一次至今研讨会已召开了四届为促进我国生物多样性研究的发展中国科学院生物多样性委员会争取并负责实施世界银行贷款环境技术援助项目生物多样性研究与信息管理子项目该项目自1993年启动在12个子项目研究所和有关管理部门的共同努力下在中国普及生物多样性概念为建立较强实力的研究队伍进行国内外培训提高了研究水平并利用世界银行贷款改善了部分研究条件和设施加强了研究的支撑力度科研工作取得令人瞩目的研究成果包括正在建设的中国生物多样正式由国家专款资助开展的研究项目课题已覆盖全国各研究单位和大学尤其在生物多样性研究与信息管理上做出了领先的作用第一个建立了信息共享的网站通过这些项目的实施不仅极大地提高了公众的生物多样性保护意识提高了政府在生物多样性保护与持续利用方面的决策能力也使中国在国际生物多样性研究领域具有一定的影响项目执行以来各子项目研究所的有关研究人员积极努力共发表研究论文 1600多篇其中SCI索引刊物论文达数百篇出版专著 128多部一批学术论文已在国际上产生良好的影响燕山地区表土孢粉植物群初步研究杨振京1, 2 许清海3 刘志明2 阳小兰3 王开发41. 中国科学院植物研究所植被数量生态学重点实验室北京 100093(2. 中国地质科学院水文地质环境地质研究所河北正定 050803)(3. 河北省科学院地理研究所石家庄 050011)(4. 同济大学海洋地质与地球物理系上海 200092燕山地区地处华北平原的北缘燕山山脉横亘于京津冀的北部滦河纵贯全区流入渤海本区地质历史悠久山区地貌峰谷参差山势高峻平原沃野千里自然环境垂直分异明显其气候植被随海拔高程的变化呈带状展布自然景观从暖温带特点经中温带最后变成具有寒温带的特点表土花粉分析结果表明燕山地区随海拔高程的降低可划分为5个孢粉组合带及相应的5个表土孢粉植物群从山顶至海滨可依次分成山地草甸针阔叶混交林或以针叶树为主的针阔叶混交林山地灌草丛滨海平原草甸和滨海草甸或盐生草甸它们基本上反映了该区现代植被的整体特征表土孢粉植物群与当地现代植被带在组成上与空间排列方面基本相同与当地现代植被的建群种属优势种属基本一致但在分布高度和组成成分等方面稍有差异本文的研究成果可为燕山地区自然生态环境保护区建设及今后的发展方向合理利用自然资源提供科学依据为首都北京周围的生态环境建设提供理论指导棉花对昆虫的化学防御研究进展周秋菊1,3 魏伟1* 钱迎倩1 马克平1 张永军2 潘贤丽3(1. 中国科学院植物研究所植被数量生态学重点实验室北京 100093)2. 中国农业科学院植物保护研究所北京 1000943. 华南热带农业大学海口分校海口 571101目前对棉花抗虫性的研究和利用主要有两个方面的内容一是研究利用棉花自身的抗虫防御体系二是应用现代生物技术培育转基因抗虫棉化学防御是棉花自身的抗虫防御机制中最主要的一种防御方式棉花中已明确具有抗虫作用的生化物质主要有三大类即单宁类黄酮类和萜烯类化合物现阶段棉花抗虫工程中常用外源基因表达产物多是单基因编码的蛋白质如Bt蛋白蛋白酶抑制剂和外源凝集素等这些外源基因表达产物对棉铃虫等棉花害虫形成的防御从某种意义上讲亦属于化学防御的范畴在外源基因导入棉花之前各棉花品种在漫长的进化过程中体内均形成了一套有利于自身发育的防御基因体系而当外源抗虫基因被导入后必将打破其自身固有的连锁群对其各种性状和生理代谢特别是作为多种病虫害抗性基础的他感化合物会产生各种各样的影响从而可能会进一步影响植物与有害生物之间的关系植物抗虫性和生物防治是目前IPM所提倡的两种最有效也是最安全的方法但是从众多研究结果来看植物抗虫性与天敌的关系并不是简单的一种形式从二者在田间的的配伍性来看表现为拮抗性加和性和增效性等多种形式本文综述了转基因棉花有抗虫功效的外源基因和他感化合物之间的交互作用同时从三营养层的角度对他感化合物和外源基因对棉花害虫主要天敌的相互关系进行探讨希望对棉花的IPM以及可持续性发展有所启发* 通讯作者weiwei@草酒的演变兼论文化多样性的保护王锦秀1 高天刚 傅德志21. 中国科学院西双版纳热带植物园, 昆明 666303(2. 中国科学院植物所北京 100093本文研究了我国草酿酒的历史并调查了西双版纳地区物种多样性丰富程度明显不同的三个傣族村寨利用草酿酒的情况以判明在不同物种多样性水平下草这一独特文化形态演变的方式由此推演相关生物多样性的历史演变提供生物多样性和地区文化多样性的保护提供新的思路和方法晋朝304年嵇含所著南方草木状被誉为世界上最早的区域性植物志共记载了南方两广及云南的重要植物80余种其中还有对草一种酒的记载南海多美酒不用檗单杵米粉杂以众草叶冶葛汁朽溲之大如卵置蓬蒿中隐蔽之.但对草中所利用的物种没有详细列出也没有说明它的利用部位17世纪屈大均在广东新语中证实了嵇含所描述的草当时在广东的一些地方也使用由豆类与米结合象山桔辣蓼马蓼等植物酿成作者在云南南部西双版纳考察时发现当地的傣族基诺族爱伲族都有利用草及稻米包谷酿酒的传统习惯其草的制作过程也与南方草木状中描述的极为相似又傣族是百越后裔所以他们利用的草可能与一千七百多年前南方草木状中描述的南越民族所利用的草系出同源那么这种文化传统的现状如何草中利用了哪些物种在历史上其物种多样水平又发生了哪些变化这些问题没有人详细研究利用民间访谈的方法结合证据标本的采集和古文献研究等方法对西双版纳地区三个傣族村寨利用草的物种多样性加以考察结果发现以下村寨按照它们周围相同面积内的物种多样性水平由高到低排列小勐养自然保护区附近的曼伞寨至今仍利用草酿酒利用物种多达92个配方中物种数可根据需要而调整勐仑的曼俄寨5年前已经不再利用草酿酒从前酿酒的老人波为还记得利用的物种数有65个而景洪市附近的曼龙匡寨十几年前就不酿酒了更不再利用草酿酒几位老人零星记得草中17种植物的名称但在他们村寨周围只找到其中的12种调查中我们发现草的利用已经存在严重问题首先是一些野生物种数量减少很难满足他们酿酒所需这些植物的利用方法逐渐失传了在曼伞的访谈中村民告诉我们祖先利用的物种数有一百多种到他们这一代好多植物没有了如他们酿制一次酒大约需要10千克红花丹但现在在曼伞寨周围已经找不到红花丹只好弃而不用其次制作草这种传统已经处于濒危状况物种多样性保护良好的曼伞仍保存这一传统而物种多样性水平低的曼俄不再酿酒只有70岁的老人波为知道草的配方因为找植物太难5年前已经不再利用草酿酒一旦老人去世这种传统也将随之消亡曼龙匡寨草已经失传上述情况显然与西双版纳近年来热带森林片段化加剧物种流失有很大的关系据估测40年来西双版纳已有数百个物种流失或濒危一种文化传统之所以消亡大多归因于另一种文化的冲击但草这种至少存在了1700年的传统文化形态面临流失原因却不仅仅在于此从以上的分析不难看出土著居民居住地周围物种多样性水平的下降是草这一传统处于濒危状况的一个重要原因这个案例也折射出在历史上随着人类利用自然程度的加剧草这一文化形态逐渐丢失的过程它足以警示我们今天应如何保护传统文化尤其是热带民族森林文化物种多样性是热带森林文化多样性赖以生存的最重要的物质基础没有这个基础一切文化形态都无从存在所以保护文化多样性不应局限于挖掘整理还应重视保护它赖以存在的最重要的物质基础物种多样性而近年来人们只注意到文化多样性对物种多样性保护所做的贡献而忽视物种多样性对文化多样性存在的重要作用本文强调物种多样性与文化多样性联系密切二者不可分割开来尤其要重视物种多样性对文化多样性保护的贡献大熊猫适宜栖息地恢复指标研究申国珍1 谢宗强1 李俊清21. 中国科学院植物研究所植被数量生态学重点实验室北京 1000932. 北京林业大学资源与环境学院北京 100083该文从地形因子森林群落因子和主食竹因子三方面建立了大熊猫栖息地退化与恢复的指标体系地形因子指标主要包括海拔坡度坡向等森林群落指标主要包括林木高度灌木高度灌木株数林木株数灌木种类云冷杉林木组成比例灌木盖度和森林郁闭度竹子密度竹子基径幼竹比例和竹子高度则是衡量主食竹质和量的主要指标研究结果表明,王朗自然保护区大熊猫栖息地恢复的标准是生长在海拔2 600~3 000 m之间坡度为25-45°的西坡西南坡和西北坡的中坡部位的森林这种森林的林木高度<15m灌木高度<2m灌木株数>5000株/hm2林木株数<300株/hm2灌木种类>5种以上云冷杉林木组成>50%以上灌木盖度80~100%森林郁闭度<50%竹子密度为 35~102株/m2竹干基径>4.4 mm幼竹比例>25%竹子高度为1.8~2.8 m的森林环境在此基础上提出了保护恢复和修复三种恢复大熊猫栖息地途径生物多样性数据集成模式初探乔慧捷 赵海军 纪力强中国科学院动物研究所北京 100080为了能够为生物多样性保护和持续利用的决策与实践提供科学准确的信息方便生态学家和分类学家在海量的信息中有效寻找到所需的资源生物多样性信息的有效集成是亟待解决的问题国外大的博物馆或博物馆联盟和一些政府或非政府的组织机构已经在这方面作了很多工作并且提供了一些接口供我们使用国内也已经建立了中国生物多样性信息系统(CBIS)和中国物种信息系统CSIS等类似的集成系统但是在数据集成方面还有很多工作要做从生态学家和分类学家需求的角度出发探讨建立一个操作简单实用广泛的数据库平台的模式根据生物多样性源数据的特点以及现有的其它领域数据集成的经验尝试采取数据格式转换模式与数据互操作模式相结合的方法现在国内生物多样性数据的情况是现已数据化的资源大部分集中在几个数据源点大都在Windows平台下实现的格式比较统一进行格式转换和相互访问比较方便没有上网的还都以文本的方式记录数据格式转换模式和数据互操作模式的结合是比较适合现状的一种集成方式在这种结合的模式下可将现存多种数据格式经专门的数据转换程序按照规定好的格式在各个数据源点进行格式转换并分别存放在源点的数据库中每个源点的数据库格式是相同的发布系统在一个单独的节点它不隶属于任何一个数据源点但含有所有数据源点的配置信息可以通过统一的接口最大限度的访问各个数据源点的所有数据资源这样发布系统对每一个数据源点的访问方式是相同的访问简单也可以很方便的从其它数据源点通过统一的接口获得所需要的信息集成度高每一个源点自己管理自己的数据更新之后的数据在发布系统处可以同时得到体现继承了数据互操作模式的数据实时更新的优点通过这种分布式组织结构可以最大限度的做到统一格式的高集成性和分布存储的高度灵活性集中控制分散管理具体的发布方法根据应用对象和使用条件的不同可采用硬件介质光盘方式或者网站的形式在完成国内数据的集成以后利用国外现有的数据库提供的检索接口将多个国外数据库集成在一起进行数据发掘并将发掘到的信息与国内数据充分结合使提供的信息更加完整系统和全面。
第五届中国博鳌高端猪业科技论坛暨第三届中国生猪产业职业教育高峰论坛圆满召开

12640卷第11热点聚焦HEAT FOCUSING第五届中国博鳌高端猪业科技论坛暨第三届中国生猪产业职业教育高峰论坛圆满召开本刊编辑/甄梦莹2023年11月10—12日,由中国博鳌高端猪业科技论坛组委会和国家现代猪业产教融合共同体共同主办的“第五届中国博鳌高端猪业科技论坛暨第三届中国生猪产业职业教育高峰论坛”在海南文昌希尔顿酒店正式开幕(见图1)。
近年来,我国生猪产业出现了资本推动、成本血拼的局面,尤其产业自2019年开始恢复,至2021年达到正常水平,直到当前产能出现相对过剩,行业严重内卷。
未来已来,生猪产业谁主沉浮本届论坛的召开旨在探讨生猪产业热点、难点问题,助力生猪产业高质量发展。
图1 论坛会场全景1 开幕式嘉宾汇聚中国博鳌高端猪业科技论坛秘书长、中国种猪信息网总编辑孙德林教授,中国农业科学院哈尔滨兽医研究所仇华吉研究员,华南农业大学邓近平研究员,中国农业科学院北京畜牧兽医研究所熊本海研究员,华南农业大学张守全教授,湖南农业大学文利新教授,重庆三峡职业学院校长、国家现代猪业产教融合共同体理事长李炯光教授,湖北生物科技职业学院校长龚伏廷教授,广东科贸职业学院校长王海林教授,山东畜牧兽医职业学院校长李学太教授,广西畜牧兽医学会养猪分会理事长许典新研究员,安徽省猪业协会副会长/秘书长、安徽省农业农村厅李东风研究员,山东省畜牧协会猪业分会秘书长、山东省畜牧总站周开锋正高级畜牧师,天津市畜牧兽医学会理事长、《猪业科学》杂志编委会主任王红军研究员,天津市农业发展服务中心副主任、《猪业科学》杂志主编郭云峰推广研究员,天津市农业科学院党支部书记兼所长、《猪业科学》杂志编委会副主任鄢明华研究员,天津市动物疫病预防控制中心主任、《猪业科学》杂志副主编梁智选研究员等专家、职业院校校长、生猪产业协会及学会嘉宾出席大会开幕式。
大北农2亿头猪工程副总裁、兆丰华集团CEO 范伟,上海卡苏生物科技有限公司副总经理兼市场总监周立臣,宁波三生生物科技股份有限公司执行总裁李琴,天津现代天骄农业科技股份有限公司董事长孙超,长沙绿叶生物科技有限公司第一副总裁何兴国,威隆·亚士可国际制药集团市场部经理陈国保,河北真信粘土科技发展有限公司技术总监戴进松,宁波第二激素厂技术总监韩玉帅,福建光华百斯特生态农牧发展有限公司董事长林长光,美加宝畜牧有限公司总经理马建升,昆明正大有限公司产业化总监兰志刚,成都英孚克斯科技有限公司大客户经理司明涛,等企业嘉宾出席大会开幕式。
中国农业生物技术学会—动物生物技术分会
中国农业生物技术学会—动物生物技术分会
致赞助单位函
尊敬的贵公司有关领导:
中国农业生物技术学会--动物生物技术分会理事会研究决定在2011年7月26日至27日在内蒙古呼和浩特召开第六届动物生物技术分会学术年会。
本次年会将邀请国内外动物生物技术领域著名专家做高级别的学术报告。
组委会希望社会各界能够给予本次会议强有力的资金支持,促进人员与技术的交流,为我国动物生物技术发展与进步做出重要贡献。
本次会议为赞助企业提供了高级别的交流平台,这也必将成为各赞助企业展示公司风貌的最佳场所。
赞助单位赞助金额与相关待遇
您的支持,将使本次年会更加精彩;您的帮助,将使数百人受益!
汇款地址:
户名:内蒙古大学
开户行:中国建设银行内蒙古分行营业部呼伦南路支行
账号:15001706667050001905
备注应写清:2011ACAB 赞助
日。
第五届 生物化学实验技能大赛实验报告

第五届生物化学实验技能大赛实验报告制备蝇蛹壳聚糖工艺改良及其理化性质研究Improved Technology of Extract Chitosan from the Pupa and Study the Physical Chemistry Character制备蝇蛹壳聚糖工艺改良及其理化性质研究郭诗静,金昂丹,梁剑云华南农业大学 生命科学学院(注:按姓名字母排序,不分先后)摘要:利用蝇蛹作为实验原料,通过改良后的酸法脱灰分,碱法脱蛋白质和脂肪制成甲壳素后,通过热浓碱法脱乙酰基处理提取壳聚糖。
改良的工艺缩短了加工的时间,但是也保证了质量。
提取过程中分别使用室温、加热、超声波三种不同的方法脱蛋白,脱色方法也一改以往用有机溶剂的方法,改用双氧水;干燥恒重法测定其水分,旋转黏度计测定其黏度,通过对成品进行这些理化性质的检测。
实验得沸水的处理效果最好,超声细胞粉碎机对提取有一定点作用,双氧水的脱色效果不错,可以进行进一步研究和推广。
关键词:蝇蛹 甲壳素 壳聚糖 水分 黏度 脱乙酰度目录1 前言 (04)1.1 甲壳素与壳聚糖及其研究状况 (04)1.2 家蝇与提取工艺 (05)1.3 超声波 (05)2 可行性分析 (06)3 实验目的 (06)4 实验原理 (06)5 实验设备 (07)5.1 仪器 (07)5.2 玻璃器具 (07)6 实验材料及试剂 (07)6.1 原材料 (07)6.2 试剂 (08)6.3 试剂的配制 (08)6.4 材料的处理 (08)7 本实验的制作流程 (11)8 制作工艺比较 (15)9 各种理化性质方法比较 (15)9.1 用干燥恒重法测定其水分 (15)9.2 用旋转黏度计测定其黏度 (16)9.3 用酸碱滴定法测定游离氨基和脱乙酰度 (17)10 结论 (18)11 注意事项 (18)12 优点 (18)参考文献 (19)英文摘要 (20)附录 (21)感想 (22)1前言1.1 甲壳素(chitin)与壳聚糖(chitosan) 及其研究状况壳聚糖(chitosan)在自然界中通常以甲壳素(chitin)的形式存在。
科苑荟萃

后引发机体免疫抑制 , 可继发多种病原混合感染。 场在仔猪断奶后有意识地对仔猪实行短期限饲以控 经
证实 ,C 2 P V 感染对胸腺 淋巴细胞的损伤在 P WS 制其采食最 防止因过度采食而引起的消化不良和 M 致病机理中发挥主导作用,巨噬细胞和树突状细胞 腹泻。 减少仔猪断奶后的营养成激。 就当前 国内外的
与 不 同季节 的环境 温度 、光 照周期 以及 动物 的内分
泌特点有关。子宫内生长阻滞 ( R) I G 的新生仔猪 U
Hale Waihona Puke ( 《 . )证明母猪妊娠后期的热应激会严重阻碍 P 05 , 0
2 0 02 0 6 0 4合 理 限饲 可减 少早 期 断奶 对仔 猪 的
的比例在夏季为 1. %。显著嵩于秋季的 48 % 影 响( 宗俊 , 01 2 . 3 殷 卞圄保 . 中才 ) 王 为了提高母猪的年生产力和栏舍利用率 ,现代
仔猪( 自然分娩 ) 为对照作相关分子微生物生态学分 析。 方法 : 利用剖腹取胎的方式将仔猪从母体子宫中
2 00 2 0 6 0 1仔 猪 初 生 重 与 产 仔 季 节 的 相 关 性 研 取 出放 于屏 障系 统 中饲养 ,于 7l 时转 移至 消毒 l E龄
究( 陈才 勇 。 王恬 。 永久 ) 霍
囊蝎 综台痖致 病机 理 ( 贾贝 贝 。 刘兴友 , 刘长 明 )
和生活环境的突变 , 会加剧其断奶后的腹激反应 , 具
断奶仔猪多系统衰竭综合症于 19 年首次在 体表现为断奶后食欲下降、 97 消化不 良、 下痢、 生长停 加拿大西部发现,随后许多网家报道了本病的存在 滞、 饲料利用率降低、 抗病力下降、 失重甚至死亡O 使 和发生 , 相继证实该病是由圆环病毒科 ( io i— 生产成本增加 , C czr r ri 严重影响猪场的经济效益。 如何减轻 de 猪圆环病毒 2型(o i d vu2P V 2 引 和克服早期断奶仔猪的应激反应 ,是当代猪生产中 a) Pr n r oi s ,C 一 ) ce c r 起的o M 典型特征是全身性淋巴肿大。 P WS 病毒感染 最关注的技术之一。 并提 出了许多有效措施。 有的猪
动物营养学报最终格式要求

亲爱的作者:您好!12祝贺您的论文终审通过被本刊录用,请抓紧时间按终审专家意见并对照以下论文模板3(修订版)认真修改论文。
4论文模板涵盖了本刊研究性论文须要遵循的各项标准和规则,如格式、单位、数字、统5计、图、表、标点符号、正斜体、名词术语以及参考文献的著录规则等。
如您认真对照本模6板修改论文,不但将提高论文的质量和被引率,而且将大大缩短论文在本刊网刊和纸刊上的发表时间。
78您的论文可能还要进行多次修改,请大力支持和配合本刊编辑的工作。
9谢谢!《动物营养学报》编辑部1011通则:121.字体字号:为了便于编辑和专家审读你的稿件,请全文用word的五号字(除表格中内1314容外)、Times New Roman字体、不加粗编写,加行号,全文通栏和自动换行。
表格中15内容用小五号字。
162.标点符号:建议中文段落用中文标点符号及其规则著录,英文摘要和中英文混排文字(参17考文献、表注)用英文标点符号及其规则著录。
参考文献、表注及名词术语后附英文与18缩写中所有标点符号后均不空格。
3.中文摘要:要求1)简洁:排除常识内容;2)独立:不得引用文中参考文献号、图号和1920公式;3)具体:尽量用具体数字说明研究取得的进展;4)便文献于收录:尽量避免包21含公式、上下标等(研究模型等特殊情况除外),以便国内外数据库收录文本数据。
关键词要选准,选准才能有利于同行在数据库中检索到您的论文和提高您论文的被引率。
22234.结果:尽量用表格将试验数据列出,尽量不用柱形图或折线图,因为表格数据最能准确反映试验的可信度和测量数据的精度。
24255.英文摘要:要请英文水平高的老师帮助你把关和修改。
266.名词术语:全文中多次出现的名词术语,后括弧标注英文名(全部小写)及其缩写,后文直接用英文缩写以节省篇幅,如平均日增重(average daily gain,ADG)。
但全文中仅出2728现1次的中文名词术语,一般不标注英文名及其缩写。
第五届(2018)规模化养猪新技术国际研讨会胜利召开

| 2018年第38卷第03期 总254期 | 872018年3月17-18日,由上海市农业科学院畜牧兽医研究所、上海生猪业行业协会、上海种猪工程技术研究中心、上海市饲料兽药行业协会以及中国品牌猪肉大联盟论坛委员会主办的“第五届(2018)规模化养猪新技术国际研讨会”在杭州盛泰开元名都大酒店胜利召开。
来自全国各地的业内嘉宾400余人参加了本次盛会,同时华中农业大学陈焕春院士受邀出席本次论坛并做报告。
本次大会开幕式由上海市农业科学院畜牧兽医研究所副所长谈永松主持,上海市农业科学院党委副书记、纪委书记徐伟林在开幕式上致辞(图1)。
此外,大会还邀请了杨亮董事长(上海富朗特)、Allan Schinckel 教授(美国普渡大学)、管武太教授(华南农业大学)、黄红梅总经理(上海新邦)、孙安权董事长(上海奥格)、章亭洲博士(浙江科峰)、穆玉云博士(上海新农)、车炼强副研究员(四川农业大学)、Michel Guillaume 教授(法国欧密斯)、夏良宙教授(台湾屏东科技大学)、朱志平研究员(中国农业科学院)、徐军军副总经理(上海洁斐然)、刘学陶研究员(台湾农业科技研究院,宁波第二激素厂资助)、施增斌经理(海博莱)、刘纪玉经理(环山集团)、吴灿文总经理(播恩集团)、万鸽总监(郑州新航道)、程建博士(美国普渡大学)、赵长生总经理(武汉华科智农)、唐罗恩博士(加裕公司)、刘俊经理(浙江沃德威先)、Jovanka Krnjaic 资深咨询师(卡美农业咨询)、王彦君博士(北京挺好)、周晓艳编辑(中国农业出版社)、林育圣博士(上海派斯德)、刘昊副教授(厦门中鲨)、朱琳娜博士(上海欧耐施)、余冰教授(四川农业大学)、王建霞博士(上海杰隆)、冯定远教授(华南农业大学,上海百立资助)、吴国忠博士(嘉兴科瑞)、高佃平博士(中牧股份)、刘惠莉研究员(上海市农业科学院畜牧兽医研究所)、张桂红教授(华南农业大学)、曾容愚博士(天康生物)、何启盖教授(华中农业大学)、常天明博士(武汉科前资助)等,共计37位资深专家(包括4位国外嘉宾)作了38场精彩绝伦的报告,为与会者分享了他们在猪群健康及其综合调控(管理、营养)技术的研究进展和成果。
猪催乳素的真核表达与生物活性验证

华南农业大学学报 Journal of South China Agricultural University 2024, 45(2): 179-189DOI: 10.7671/j.issn.1001-411X.202302022谢社风, 韩贝贝, 高凤磊, 等. 猪催乳素的真核表达与生物活性验证[J]. 华南农业大学学报, 2024, 45(2): 179-189.XIE Shefeng, HAN Beibei, GAO Fenglei, et al. Eukaryotic expression and bioactivity verification of porcine prolactin[J]. Journal of South China Agricultural University, 2024, 45(2): 179-189.猪催乳素的真核表达与生物活性验证谢社风1,韩贝贝1,高凤磊2,马 莹1,李 莉1,张守全1,邹 娴3,卫恒习1,4(1 国家生猪种业工程技术研究中心/广东省农业动物基因组学与分子育种重点实验室/华南农业大学 动物科学学院,广东 广州 510642; 2 广东农工商职业技术学院 热带农林学院, 广东 广州 510507; 3 广东省农业科学院动物科学研究所, 广东 广州 510640; 4 岭南现代农业科学与技术广东省实验室 茂名分中心, 广东 茂名 525000)摘要: 【目的】催乳素(Prolactin,PRL)具有广泛的生理调节作用,但其多效性机制仍不清楚。
为了更好地研究猪PRL的多效性,本研究制备猪源PRL真核重组蛋白并验证其生物活性。
【方法】利用分子克隆技术将猪PRL基因克隆到慢病毒表达载体pCDH-CMV-MCS-EF1-GFP+Puro中,经慢病毒包装获得携带猪PRL基因的PRL−慢病毒;用浓缩的PRL−慢病毒感染CHO-K1细胞,经嘌呤霉素筛选后,获得能够分泌PRL重组蛋白的阳性细胞系CHO-K1-PRL;利用镍柱亲和层析法对重组蛋白进行纯化并进行LC-MS/MS质谱鉴定,利用HC11细胞体外培养体系验证PRL重组蛋白的生物活性。
第五届中国畜牧科技论坛在重庆开幕

来贺信 ,向论坛的召开和重庆市畜牧科技 院建院
6 0周年表示热烈祝贺 ,向与会代表和全国畜牧科
究所所长蔡辉益研究员、 中国农业科学院饲料研究
所副所长齐广海研究员 、 北京市饲料工业协会 书长
技工作者致 以诚挚的问候 。农业部副部长、 九三学
社 中央副主席 、 中国农学会会长张桃林 , 农业部原 副部长洪绂 曾, 农业部原副部长 、 中国奶业协会理 事长刘成果 , 中国农业科学院副院长 、 中国工程 院
重大科技项 目研究 ,突破一系列重大技术瓶颈 , 为
现代畜牧业发展提供雅有 的科技支撑。 . 力 张桃林说, 畜
牧业的发展 , 直接关系到老百姓餐桌上的 肉蛋奶 , 以 可
说—头连着产业, —头连着民生。 畜产品的 质量安全, 与
人民群众的身体缝康息 息相关, 是广大人民
关
也涵盖了各级经销商和终端养猪户 , 宣传点面达数 百万次 ,并持续扩大与大众媒体和地方媒体的合 作; 观众组织方面 , 除加强 呼叫中心的一对一邀请 外 ,还将扩大和优化专业千人买家团的组织工作 ;
下一步工作要求 : 各地要切实加强安全用药监
管工作 , 加大违禁药物查处力度。要高度重视兽药 残 留监控工作 ,认真组织实施 2 1 年下半年兽药 0 1 残留监控计划 , 按时上报检测结果 。检测出阳性样 品的省份要严格执行阳性样品跟踪检测和追溯制 度, 并上报跟踪检测结果 。要积极参与残留检测方
mM^ l , d . .D c m 7 9
色、 ” 环保 为主题 , 国内外 多年从事微生态领域 邀请
研究的专家学者, 针对微生态发酵技术以及微生态在动
中 圈
I 动保资讯
观众和展商的在线注册和分类管理 ;媒体宣传方 面, 成立展会新闻中心 , 组织 6 4家平 面 、 网络媒体 和 2 家知名企业 内刊共 同合作 ,宣传面不仅涵盖 2 从畜牧业生产到终端畜产品的各个环节大小企业 ,
第五届饲料新技术国际论坛暨北京市生物饲料技术研讨会在北京召开
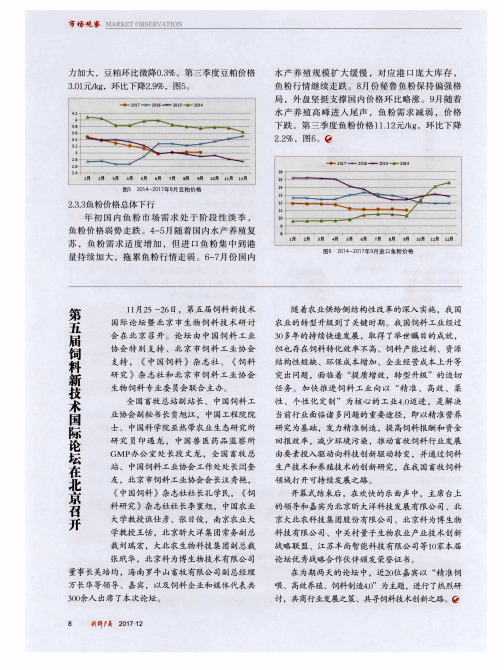
2 . 2 % ,图6 。
・ .-2 0 1 1- _ - 2 0 1 6 — ÷ -2 0 1 5・ ..2 0 1 4
生产技 术 和养殖技术 的创新研 究 ,在我 国畜牧 饲料
领域打 开可持 续发展之路 。 开幕 式结束后 ,在 欢快 的 乐曲声 中,主席 台上
《 中国饲料》杂志社社长孔学民, 《 饲
丑 I - - I JI l 料 研 究 》杂志社社 长李 寰旭 ,中国农 业 大 学教授 谯仕彦 、张 日俊 ,南京农 业大 学教授 王恬 北 京昕大 洋集 团常务 副总
GM P办公 室 处 长 段 文 龙 , 全 国 畜 牧 总
3前行业面临诸多问题的重要途径,即以精准营养 研 究为基础 ,发 力精 准制造 ,提 高饲料报 酬和资金
回报 效率 ,减 少环境 污染 ,推动 畜牧 饲料行 业发展
由要 素投入 驱动 向科技 创新驱动 转 变,并通过 饲料
站 、中国饲料 工业 协会 工作处处 长闫奎 友 ,北京市饲料 工业协会会 长 汪秀艳 ,
图6 2 0 1 4 - 2 0 1 7 年9 月进E l 鱼粉价格
随 着农 业供给侧 结梅 性改革的深入 实施 ,我 国
霎 晋
国 际 论 坛 暨 北 京 市 生 物 饲 料 技 术 研 讨
会 在 北 京 召 开 。论 坛 由 中 国饲 料 工 业
农业的转 型升级到 了关键 时期 。我 国饲料 工业经过
猪水肿病菌毒素a亚单位(slt-ⅱea)单抗的制备与slt-ⅱea与f18ab菌毛a亚单..

扬州大学硕士学位论文组菌正确表达了F18ab菌毛A亚单位和SLT-IIeA亚单位的联合表达产物。
为了研究联合表达产物的免疫学特性,我们用ICR小鼠进行了免疫原性试验。
己诱导表达后的重组菌菌体裂解物乳化后两次免疫小鼠,二次免疫后8天眶静脉采血做ELISA,检测抗体效价,同时设立pSLT一[1ea、pFl07重组菌诱导表达产物的对照组。
结果表明:本重组菌能有效对小鼠进行保护,保护率达到7/8。
关键词:猪水肿病F18ab菌毛志贺样毒素II型变异体联合表达亚单位融合蛋白免疫印迹刖罱猪水肿病(SwineEdemadisease,ED)是由几种特定O抗原(主要为0139,其次为0138、0141)的产veto细胞毒素大肠杆菌(VTEC)弓I起的致死性传染病,多发生于断奶后不久的仔猪,病猪常出现一系列神经症状如共济失调,惊厥和后肢瘫痪,死亡剖检可见皮下组织、肠系膜、冒大弯粘膜下层水肿等11J。
该病发病率低,但死亡率可高达90%以上,给养猪业造成了相当大的经济损失{341。
随着近几十年来深入研究,对水肿病的发病机理有了较为明确的认识:仔猪断奶1周后,由于断奶应激,母源抗体的逐渐丧失,消化道内环境的改变,饲养管理,营养条件,环境因素的忽然改变,导致机体的抵抗力下降,当易感仔猪感染某些可产生志贺样毒素的大肠杆菌时,细菌以自身的F18ab菌毛黏附于小肠绒毛上,从而在小肠内定居和繁殖,在这些过程中产生SLT.1ie毒素并被吸收。
毒素被吸收后,毒素的B亚单位与细胞上球丁糖基神经酰胺(Gb4)发生特异性结合,随后A亚单位进入细胞内,通过其KNA.N.糖苷酶(KNA-N—Glgcosidase)活性,可脱去真核细胞rRNA的28s亚基上的一个特定的腺嘌呤残基,使氨酰基tRNA不能与核糖体结合,从而阻断细胞内蛋白质的合成,导致细胞死亡。
由于SLT—lie是一种血管毒索,因此当其被肠道吸收后,可在不同部位引起血管内皮细胞损伤,改变血管通透性,导致病猪出现水肿,当脑部水肿后,可压迫神经组织,从而出现神经症状【4】。
NR2F2基因调控猪PK15细胞增殖和凋亡的研究

畜牧兽医学报 2023,54(8):3242-3251A c t a V e t e r i n a r i a e t Z o o t e c h n i c a S i n i c ad o i :10.11843/j.i s s n .0366-6964.2023.08.011开放科学(资源服务)标识码(O S I D ):N R 2F 2基因调控猪P K 15细胞增殖和凋亡的研究张万锋,赵天枝,李 娇,尤紫薇,杨 阳,蔡春波,高鹏飞,曹果清,郭晓红*,李步高*(山西农业大学动物科学学院,太谷030801)摘 要:旨在探究N R 2F 2基因对猪P K 15细胞增殖和凋亡的影响㊂本研究在猪P K 15细胞中过表达N R 2F 2基因,通过q R T -P C R ㊁W e s t e r n b l o t ㊁C C K -8㊁E d U ㊁流式细胞术㊁划痕试验等方法检测了过表达N R 2F 2基因对细胞增殖的影响;采用C O -I P 技术验证N R 2F 2与S m a d 4蛋白结合能力;用q R T -P C R 方法检测了N R 2F 2基因对S m a d 4基因表达以及下游基因C y c l i n B ㊁C D K 1㊁C D K 4㊁C yc l i n D 1表达的影响;挽救试验过表达N R 2F 2基因且干扰S m ad 4基因检测下游基因及增殖基因K i 67㊁P C N A 的表达㊂结果发现,过表达N R 2F 2基因极显著上调了K i 67基因的表达(P <0.01),显著上调了P C N A 的表达(P <0.05),促进了细胞周期的进程,抑制了细胞凋亡,促进了细胞的增殖㊂C O -I P 结果证明了N R 2F 2与S m a d 4蛋白物理性结合,且发现过表达N R 2F 2基因后,S m a d 4基因表达显著上升(P <0.01),S m a d 4基因调控的下游基因C y c l i n B ㊁C D K 1㊁C D K 4㊁C yc l i n D 1表达显著升高(P <0.01);干扰N R 2F 2基因后,S m ad 4基因表达显著下降(P <0.01),S m a d 4基因调控的下游基因表达显著降低(P <0.01)㊂挽救试验发现,过表达N R 2F 2基因后,干扰S m a d 4基因的表达,能显著降低C y c l i n D 1㊁C yc l i n B ㊁C D K 1以及K i 67㊁P C N A 的表达(P <0.01,P <0.05)㊂本研究结果表明,过表达N R 2F 2基因后促进了细胞的增殖,抑制凋亡,且N R 2F 2蛋白能与S m ad 4蛋白结合并影响S m a d 4基因的表达,进而影响下游基因的表达㊂关键词:N R 2F 2基因;S m a d 4基因;细胞增殖;P K 15细胞中图分类号:S 828.2 文献标志码:A 文章编号:0366-6964(2023)08-3242-10收稿日期:2022-12-26基金项目:国家自然科学基金面上项目(32272846);山西省重点研发计划项目(202102140601005)作者简介:张万锋(1995-),男,山西朔州人,博士生,主要从事动物遗传育种与繁殖研究,E -m a i l :z h a n g w a n f e n g123@126.c o m *通信作者:郭晓红,主要从事动物遗传育种与繁殖研究,E -m a i l :g _x i a o h o n g@126.c o m ;李步高,主要从事动物遗传育种与繁殖研究,E -m a i l :ji n r e n n @163.c o m S t u d y o n N R 2F 2G e n e R e g u l a t i n g P r o l i f e r a t i o n a n d A p o pt o s i s o f P o r c i n e P K 15C e l l s Z H A N G W a n f e n g ,Z H A O T i a n z h i ,L I J i a o ,Y O U Z i w e i ,Y A N G Y a n g,C A I C h u n b o ,G A O P e n g f e i ,C A O G u o q i n g ,G U O X i a o h o n g *,L I B u ga o *(C o l l e g e o f A n i m a l S c i e n c e ,S h a n x i A g r i c u l t u r a l U n i v e r s i t y ,T a i gu 030801,C h i n a )A b s t r a c t :T h i s s t u d y a i m e d t o e x p l o r e t h e e f f e c t o f N R 2F 2g e n e o n t h e p r o l i f e r a t i o n a n d a p o pt o s i s o f p o r c i n e P K 15c e l l s .I n t h i s s t u d y ,t h e e f f e c t o f o v e r e x pr e s s i o n o f N R 2F 2g e n e o n c e l l p r o l i f e r a t i o n w a s d e t e c t e d b y q R T -P C R ,W e s t e r n b l o t ,C C K -8,E d U ,f l o w c y t o m e t r y,s c r a t c h a s s a y a n d o t h e r m e t h o d s i n p o r c i n e P K 15c e l l s .C O -I P t e c h n o l o g y w a s u s e d t o v e r i f y t h e b i n d i n ga b i l i t y o f N R 2F 2t o S m a d 4p r o t e i n .T h e e f f e c t o f N R 2F 2g e n e o n S m a d 4g e n e e x pr e s s i o n a n d t h e e x p r e s s i o n o f d o w n s t r e a m g e n e C y c l i n B ,C D K 1,C D K 4,C y c l i n D 1w e r e d e t e c t e d b y q R T -P C R.T h e e x p r e s s i o n o f d o w n s t r e a m g e n e s a n d p r o l i f e r a t i n g ge n e s K i 67,P C N A w e r e d e t e c t e d b y r e s c u e e x p e r i m e n t o v e r e x pr e s s e d N R 2F 2g e n e a n d i n t e r f e r e d w i t h S m a d 4g e n e .T h e r e s u l t s s h o w e d t h a t o v e r e x p r e s s i o n o f N R 2F 2g e n e s i g n i f i c a n t l y u p r e g u l a t e d t h e e x pr e s s i o n o f8期张万锋等:N R2F2基因调控猪P K15细胞增殖和凋亡的研究K i67(P<0.01),a n d s i g n i f i c a n t l y u p r e g u l a t e d t h e e x p r e s s i o n o f P C N A(P<0.05),p r o m o t e d c e l l c y c l e p r o g r e s s i o n,i n h i b i t e d c e l l a p o p t o s i s,a n d p r o m o t e d c e l l p r o l i f e r a t i o n.T h e C O-I P r e s u l t s p r o v e d t h a t N R2F2a n d S m a d4p r o t e i n s w e r e p h y s i c a l l y b o u n d.A f t e r o v e r e x p r e s s i o n o f N R2F2g e n e,t h e e x p r e s s i o n o f S m a d4g e n e i n c r e a s e d s i g n i f i c a n t l y(P<0.01),a n d t h e e x p r e s s i o n o f d o w n s t r e a m r e g u l a t e d g e n e s b y S m a d4g e n e C y c l i n B,C D K1,C D K4a n d C y c l i n D1s i g n i f i c a n t l y i n c r e a s e d(P<0.01).A f t e r i n t e r f e r i n g w i t h N R2F2g e n e,t h e e x p r e s s i o n o f S m a d4g e n e s i g n i f i c a n t l y d e c r e a s e d(P<0.01),a n d t h e e x p r e s s i o n o f d o w n s t r e a m g e n e s r e g u l a t e d b y S m a d4g e n e w a s s i g n i f i c a n t l y d e c r e a s e d(P<0.01).T h e r e s c u e e x p e r i m e n t s s h o w e d t h a t a f t e r o v e r e x p r e s s i o n o f N R2F2g e n e,i n t e r f e r i n g w i t h t h e e x p r e s s i o n o f S m a d4g e n e c o u l d s i g n i f i c a n t l y r e d u c e t h e e x p r e s s i o n o f C y c l i n D1,C y c l i n B,C D K1,K i67a n d P C N A (P<0.01,P<0.05).I n s u mm a r y,o v e r e x p r e s s i o n o f N R2F2g e n e p r o m o t e d c e l l p r o l i f e r a t i o n a n d i n h i b i t e d a p o p t o s i s,a n d N R2F2p r o t e i n c o u l d b i n d t o S m a d4p r o t e i n a n d a f f e c t t h e e x p r e s s i o n o f S m a d4,w h i c h i n t u r n a f f e c t e d t h e e x p r e s s i o n o f d o w n s t r e a m g e n e s.K e y w o r d s:N R2F2g e n e;S m a d4g e n e;c e l l p r o l i f e r a t i o n;P K15c e l l*C o r r e s p o n d i n g a u t h o r s:G U O X i a o h o n g,E-m a i l:g_x i a o h o n g@126.c o m;L I B u g a o,E-m a i l: j i n r e n n@163.c o m细胞增殖是细胞重要的生命活动,也是机体生长㊁发育㊁繁殖和遗传的基础[1-3]㊂细胞增殖是一个复杂而有序的过程,主要受细胞周期因子和周期蛋白依赖激酶调控,这些基因由很多信号通路所调控,如W n t信号通路㊁N o t c h信号通路㊁T G F-β信号通路,而这些通路又受到许多转录因子的调控[4-11]㊂鸡卵清蛋白上游启动子转录因子2(C O U P-T F I I,也叫N R2F2)是核受体家族超家族成员,在1986年因直接结合于卵清蛋白的启动子区调节其转录被发现[12]㊂在脊椎动物中,鸡卵清蛋白上游启动子转录因子家族由3个成员组成,核受体亚家族2组F成员1(N R2F1)㊁N R2F2和核受体亚家族2组F成员6(N R2F6)㊂虽然它们在不同的染色体上,但氨基酸序列是高度同源的[13]㊂N R2F2基因在组织中广泛表达,在骨骼㊁肌肉㊁脂肪形成和代谢平衡中起重要调节作用[14-16]㊂N R2F2作为一种转录因子,通过多种机制调控下游靶基因的转录㊂N R2F2可以通过与D N A结合元件结合,直接或间接激活基因表达㊂N R2F2可以与W n t家族成员10b(W n t10b)启动子区域结合,抑制W n t10b的表达[17]㊂N R2F2可以与C C A A T增强子结合蛋白(C/E B Pα)启动子结合,抑制C/E B Pα的表达[18]㊂N R2F2可以与维甲酸X受体(R X R)的D N A结合位点竞争性结合[19]㊂N R2F2还作为一种转录抑制剂,可与R X R和甲状腺激素受体相互作用,抑制P P A R的表达[20]㊂此外,N R2F2还可以通过直接结合R X R的L B D结构域来抑制核受体的转录[21]㊂为了确定N R2F2基因对细胞增殖的作用机制,M a等[22]研究发现,在W J-M S C s细胞中敲除N R2F2基因后导致C y c l i n D1和C D K4表达降低,细胞增殖减慢㊂Q i n等[23]对敲除N R2F2基因的P C3细胞与正常细胞进行了转录组比较,通过基因富集发现,在N R2F2基因表达低时,T G F-β信号通路下游调控的基因全部富集,说明N R2F2能调控T G F-β信号通路㊂敲除N R2F2基因后,显著改变了T G F-β信号通路下游基因p21㊁p15和C y c l i n D1的表达㊂W a n g等[24]研究发现,N R2F2基因在直肠癌细胞中与P T E N㊁S m a d4基因的表达相关,而S m a d4基因是T G F-β信号通路的关键基因㊂S m a d4基因主要功能是参与T G F-β信号通路的信号传导㊂进一步研究发现,N R2F2蛋白能够与S m a d4蛋白结合,与突变型的S m a d4不能结合,直接证明了N R2F2与S m a d4之间的结合作用㊂表明在癌细胞中,N R2F2基因可以通过与S m a d4基因相互作用,进而影响T G F-β信号通路传导㊂目前,围绕N R2F2基因的研究主要集中在其对人类癌症等疾病㊁小鼠肌肉与脂肪等组织的作用机理,针对N R2F2基因调控猪源细胞增殖的机制还未见报道㊂本试验旨在探究N R2F2基因对细胞增殖㊁凋亡的影响,为进一步研究猪生长发育的分子机制提供理论基础㊂3423畜牧兽医学报54卷1材料与方法1.1材料1.1.1细胞系和质粒本研究中使用的P K15细胞和293T细胞来自山西农业大学动物遗传育种与繁殖博士点实验室㊂p L e n t i-C M V-G F P-S V-p u r o㊁p L e n t i-C M V-N R2F2-G F P-S V-p u r o㊁p C M V-d R8.91㊁p C M V-V S V-G㊁p C D-N A3.1-F l a g㊁p C D N A3.1-F l a g-N R2F2㊁p H S-C R054㊁p H S-C R054-s g1㊁p H S-C R054-s g2质粒均为山西农业大学动物遗传育种实验室保存㊂1.1.2 主要试剂 P r i m e S c r i p t T M R T M a s t e r M i x㊁R N A i s o P l u s r e g e n t和S Y B R P r e m i x E x T a q I I购于T a K a R a公司;P B S粉末㊁青链霉素混合液㊁胰蛋白酶购于中国索莱宝公司;F B S和D M E M购自G i b c o公司;兔二抗㊁鼠二抗购于L I-C O R公司; L i p o f e c t a m i n e3000购自I n v i t r o g e n公司;C C K-8试剂盒购自D o j i n d o公司;E d U试剂盒㊁细胞周期试剂盒㊁细胞凋亡试剂盒购自凯基生物公司;R I P A裂解液(强)㊁蛋白酶抑制剂㊁蛋白凝胶试剂盒㊁N C膜㊁脱脂奶粉购自博士德公司;β-a c t i n抗体购自博奥森公司;N R2F2抗体购自A b c a m公司;P C N A抗体购自C S T公司;s i R N A购自吉玛公司,s i R N A序列见表1㊂表1s i R N A序列T a b l e1S e q u e n c e s o f s i R N A名称N a m e序列S e q u e n c eS m a d4C A C C A G G A A U U G A U C U C U C A G G A U U S m a d4A A U C C U G A G A G A U C A A U U C C U G G U G s i R N A-N C-F U U C U C C G A A C G U G U C A C G U T Ts i R N A-N C-R A C G U G A C A C G U U C G G A G A A T T1.2试验方法1.2.1慢病毒包装及转染细胞转染前1d,胰酶消化293T细胞,接种细胞于10c m细胞培养皿,培养1d,当细胞浓度达到60%~80%时进行转染㊂用L i p o f e c t a m i n e3000将10μg p L e n t i-C MV-G F P-S V-p u r o或p L e n t i-C MV-N R2F2-G F P-S V-p u r o质粒与7.5μg p C MV-d R8.91㊁5μg p C MV-V S V-G包装质粒共转染293T细胞中,转染6h后,更换为完全培养基,继续培养48㊁72h后,收集293T细胞培养液,4ħ㊁1000r㊃m i n-1离心5m i n,去除细胞碎片,0.45μm针式滤器过滤,收集病毒液置于-80ħ冰箱,用于感染P K15细胞㊂将P K15细胞接种于6孔板中,待细胞汇合度达到50%~60%,每孔加入1m L病毒上清液,感染6h后,更换为完全培养基培养,感染72h后,荧光显微后荧光显微镜检测E G F P的表达,形成稳定转染N R2F2基因的细胞㊂1.2.2质粒和s i R N A转染 P K15细胞接种至10c m培养皿中,使用含10%F B S和1%双抗的低糖D M E M培养基在37ħ,5%C O2条件下进行培养㊂将细胞进行消化计数,接种至6孔板中,细胞生长汇合至40%~50%即可进行转染㊂利用L i p o-f e c t a m i n e3000脂质体分别将p H S-C R054㊁p H S-C R054-s g1㊁p H S-C R054-s g2质粒㊁s i R N A-N C及s i R N A-S m a d4转染至P K15细胞中,每组3个重复㊂按转染试剂操作说明,取两个1.5m L离心管,分别加入125μL D M E M培养液,然后于其中一管加入100p m o l s i R N A或4μg质粒吹打混匀;另一管加入10μL转染试剂吹打混匀,室温静置5m i n 后,将含有s i R N A或质粒的培养液加入含转染试剂的培养液中吹打混匀,室温静置20m i n㊂将制备好的转染溶液加至6孔板,并用D M E M加至每孔2m L,于37ħ,5%C O2中培养,6h后更换为含10%胎牛血清的培养液㊂转染后观察结果并收取细胞进行后续试验㊂1.2.3 R N A提取及实时荧光定量P C R 使用R N A i s o P l u s提取总R N A,提取方法参照试剂盒说明书㊂将提取的总R N A用核酸蛋白测定仪测定其纯度及浓度,O D260n m/O D280n m为1.8~2.0的总R N A用于后续研究,采用P r i m e S c r i p t T M R T M a s t e r M i x试剂盒将R N A反转录成c D N A㊂以18S r R N A为内参基因,采用q R T-P C R对基因的表达水平进行检测㊂q R T-P C R反应体系: c D N A2μL,2ˑS Y B R P r e m i x E x T a qⅡ10μL,上㊁下游引物(表2)各0.5μL,R N A a s e F r e e d d H2O补至20μL㊂反应程序:95ħ30s;95ħ5s,60ħ30s,35个循环;熔解曲线程序为95ħ15s,60ħ35s,95ħ㊂结果采用2-ΔΔC t法进行分析,每个样本做3次重复㊂1.2.4总蛋白提取和W e s t e r n b l o t检测提取细胞总蛋白,利用核酸蛋白浓度测定仪测定蛋白浓度㊂取200n g蛋白上样,进行聚丙烯酰胺凝胶电泳㊂电泳结束后,采用湿转法进行转膜,5%封闭蛋白粉室温封闭1h,一抗采用1ʒ1000稀释,4ħ孵44238期张万锋等:N R 2F 2基因调控猪P K 15细胞增殖和凋亡的研究表2 q R T -P C R 引物序列T a b l e 2 T h e p r i m e r s s e qu e n c e o f q R T -P C R 基因G e n e引物序列(5'ң3')P r i m e r s e qu e n c e 产物长度/b p P r o d u c t s l e n gt h N R 2F 2F :G A A C A T T T G C G A A C T G G C C GR :G C A C T G T G C T G C G T T C A G151P C N A F :G T G A T T C C A C C A C C A T G T T C R :T G A G A C G A G T C C A T G C T C G145K i 67F :A G C C C G T A T C G T G T G C A A A A R :C C T G C A T C T G T G T A A G G G C A122S m a d 4F :T T TG C G T C A G T G T C A T C G R :T G C T C T G C C T T G G G T A A T236C yc l i n D 1F :G C G A G G A A C A G A A G T G C G R :T G G A G T T G T C G G T G T A G A T G C192C yc l i n B F :T G G C T A G T G C A G G T T C A G R :C A G T C A C A A A G G C A A A G T199C D K 1F :C C C T C C T G G T C A G T T C A T R :T A G G C T T C C T G G T T T C C466C D K 4F :G C A T C C C A A T G T T G T C C G R :G G G G T G C C T T G T C C A G A T A12518SF :C C C A CG G A A T C G A G A A A G A G R :T T G A C G G A A G G G C A C C A122育过夜㊂隔天回收一抗,P B S T 洗膜3次,加入荧光二抗,避光室温孵育1h ,洗膜后使用L I C O R 仪器曝光,使用仪器自带软件I m a ge S t u d i o 计算分析条带光密度值㊂1.2.5 C C K -8㊁E d U 试验 将细胞接种至96孔细胞板中,每孔细胞约1000个,在铺板后0㊁1㊁2㊁3㊁4㊁5d 分别进行检测,检测前每孔加入10μL 的C C K -8试剂,在细胞培养箱孵育3h 后,使用酶标仪检测450n m 吸光度㊂取处于对数生长期的细胞,以每孔4000~10000个细胞接种于96孔板中,每孔加入100μLE d U 染液和100μL H o e c h s t 33342溶液,染色完毕后使用荧光显微镜成像㊂1.2.6 流式细胞术检测 将细胞接种至6c m 细胞培养皿,培养至细胞浓度80%~90%时,收集的细胞在70%冷乙醇中重悬,4ħ固定过夜,复温后加入400μL P I 染色,4ħ避光孵育30m i n,流式细胞仪检测㊂将细胞接种至6c m 细胞培养皿,培养至细胞浓度80%~90%时,P B S 洗2次,收集细胞;加入500μL 的B i n d i n g Bu f f e r 悬浮细胞,加入5μL A n -n e x i n V -A P C 混匀后,加入5μL 7-A A D 染液,混匀,室温㊁避光㊁孵育15m i n;流式细胞仪检测㊂1.2.7 伤口愈合试验 将细胞接种至6孔板中,待其长满后,用1m L 蓝枪头在六孔板内划线,P B S清洗2次,加培养基继续培养,观察0㊁12㊁24㊁36㊁48h 时划痕处细胞的生长状态㊂1.2.8 C O -I P 试验 收集转染了过表达N R 2F 2且携带F l a g 标签蛋白载体的P K 15细胞,加入40μL F l a g 树脂的裂解液在4ħ下轻轻摇晃孵育过夜,然后用500μL 细胞裂解缓冲液漂洗树脂,混匀仪上室温漂洗5m i n ,4ħ500r ㊃m i n -1离心5m i n ,重复3次,加入50μL 洗脱缓冲液,漩涡震荡20s ,放摇匀仪上,室温洗脱10~15m i n,漩涡震荡20s ,4ħ12000r ㊃m i n -1离心5m i n,取上清到新的离心管中,用于W e s t e r n b l o t㊂1.2.9 数据分析 应用S P S S 22.0进行统计分析㊂数据采用单因素方差分析和独立样本t 检验进行比较,P <0.05为差异显著,P <0.01为差异极显著㊂2 结 果2.1 慢病毒包装及N R 2F 2基因过表达效率病毒液感染P K 15细胞48h 后,荧光显微镜观察发现O E -N C 组和O E 组都表达了绿色荧光蛋白(图1A )㊂q R T -P C R 和W B 检测过表达N R 2F 2基因的效率,结果发现,慢病毒液感染P K 15细胞后,5423畜牧兽医学报54卷N R2F2基因m R N A水平和蛋白水平极显著增加(P<0.01,图1B~D)㊂A.绿色荧光蛋白E G F P表达(标尺为100μm);B~D.过表达N R2F2基因m R N A和蛋白水平表达㊂ ** 表示差异极显著(P<0.01), * 表示差异显著(P<0.05)㊂下同A.E x p r e s s i o n o f g r e e n f l u o r e s c e n t p r o t e i n E G F P(b a r=100μm);B-D.E x p r e s s i o n o f N R2F2g e n e a t m R N A a n d p r o t e i n l e v e l s. ** i n d i c a t e e x t r e m e l y s i g n i f i c a n t d i f f e r e n c e(P<0.01), * i n d i c a t e s i g n i f i c a n t d i f f e r e n c e(P<0.05).T h e s a m e a s b e l o w图1过表达N R2F2基因效率F i g.1T h e e f f i c i e n c y o f o v e r e x p r e s s i o n o f N R2F2g e n e2.2过表达N R2F2基因对细胞增殖、凋亡的影响慢病毒感染P K15细胞后,结果显示K i67基因m R N A表达极显著增加(P<0.01,图2A); q R T-P C R㊁W B检测结果显示,P C N A表达量显著增加(P<0.05,图2A㊁2B㊁2C)㊂C C K-8结果显示,过表达N R2F2基因后细胞的吸光度值随着时间的增加极显著上升(P<0.01),差异具有统计学意义,表明促进了细胞的增殖(图2D)㊂E d U结果显示,过表达N R2F2基因后S期细胞数量显著增多(P< 0.01),表明促进了细胞的增殖(图2E㊁F)㊂流式细胞仪检测过表达N R2F2基因后细胞周期变化,发现过表达N R2F2基因后,处于G1期细胞的数量显著低于对照组(P<0.01),S期细胞的数量显著高于对照组(P<0.01),表明N R2F2基因可以改变细胞周期,促进G1期向S期转变,从而促进细胞增殖(图2G㊁H)㊂通过流式细胞仪检测过表达N R2F2基因对细胞凋亡的影响,结果发现,过表达N R2F2基因后,早凋细胞占比增多,晚凋细胞占比减少,表明N R2F2基因可以抑制细胞的凋亡(图2I)㊂通过划痕试验检测N R2F2基因对细胞迁移的影响,结果表明,过表达N R2F2基因后,细胞迁移速度加快,在48h后基本汇合,而对照组细胞还未完全融合(图2J)㊂2.3N R2F2与S m a d4物理结合并调控其表达进而影响下游基因的表达为了确定N R2F2基因调控细胞增殖的具体机制,本研究通过蛋白相互作用软件预测了一些与N R2F2相互作用的蛋白㊂发现能与N R2F2互作的蛋白有15个,其中我们将关注点聚焦在S m a d4蛋白上(图3A)㊂进行了C o-I P分析来检测N R2F2和S m a d4之间的相互作用㊂用F l a g标签蛋白调取与它结合的复合物,能检测到S m a d4蛋白,说明N R2F2蛋白能与S m a d4蛋白相互作用行使功能(图3B)㊂为了探究N R2F2基因是否调控S m a d4基因的表达,检测了过表达和敲除N R2F2基因对S m a d4基因表达的影响,发现过表达N R2F2基因后,S m a d4基因表达量升高(P<0.01,图4A),下游基因C y c l i n B㊁C D K1㊁C D K4㊁C y c l i n D1表达量均显著上升(P<0.01,P<0.05,图4C);而敲除N R2F2基因后,S m a d4基因表达量降低(P<0.01,图4B),下游基因C y c l i n B㊁C D K1㊁C D K4㊁C y c l i n D1表达量均显著下降(P<0.01,图4D)㊂64238期张万锋等:N R 2F 2基因调控猪P K 15细胞增殖和凋亡的研究A~C .过表达N R 2F 2基因后增殖相关基因表达变化;D.C C K -8检测细胞增殖;E ㊁F .E d U 检测细胞增殖;G ㊁H.流式细胞术检测细胞周期;I .流式细胞术检测细胞凋亡;J .划痕试验检测细胞增殖A -C .T h e e x p r e s s i o n c h a n g e o f p r o l i f e r a t i o n r e l a t e d g e n e s a f t e r o v e r e x p r e s s i o n o f N R 2F 2g e n e ;D.C e l l pr o l i f e r a t i o n w a s d e -t e r m i n e d b y C C K -8;E ,F .C e l l p r o l i f e r a t i o n w a s d e t e r m i n e d b y E d U ;G ,H.C e l l c y c l e w a s d e t e r m i n e d b y f l o w c y t o m e t r y;I .A p o p t o s i s w a s d e t e c t e d b y f l o w c y t o m e t r y ;J .C e l l p r o l i f e r a t i o n w a s d e t e c t e d b y s c r a t c h a s s a y图2 N R 2F 2基因对细胞增殖、凋亡的影响F i g .2 T h e e f f e c t s o f N R 2F 2g e n e o n c e l l p r o l i f e r a t i o n a n d a p o pt o s i s 挽救试验中,在过表达N R 2F 2基因后,干扰S m a d 4基因表达,S m a d 4表达量显著降低(P <0.01,图5A ),且能够显著缓解过表达N R 2F 2基因引起的S m a d 4下游基因C y c l i n D 1㊁C yc l i n B ㊁C D K 1的表达的升高,降低了它们的表达量(P <0.01,P <0.05,图5B )㊂通过检测增殖相关基因的表达,结果发现,干扰S m a d 4基因表达,能缓解过表达N R 2F 2基因对细胞增殖的影响,P C N A 和K i 67的表达量显著降低(P <0.01,P <0.05,图5C )㊂7423畜 牧 兽 医 学 报54卷A.N R 2F 2结合蛋白预测;B .C O -I P 结果A.P r e d i c t i o n o f N R 2F 2-b i n d i n g pr o t e i n s ;B .T h e r e s u l t o f C O -I P 图3 N R 2F 2结合蛋白预测及C O -I P 结果F i g .3 T h e r e s u l t s o f N R 2F 2b i n d i n g pr o t e i n p r e d i c t i o n a n d C O -IP A.过表达N R 2F 2后S m a d 4的表达变化;B .敲除N R 2F 2后S m a d 4的表达变化;C .过表达N R 2F 2后S m a d 4调控的下游基因的表达变化;D.敲除N R 2F 2后S m a d 4调控的下游基因的表达变化㊂不同大写字母表示差异极显著(P <0.01);不同小写字母表示差异显著(P <0.05),下同A.T h e e x p r e s s i o n c h a n g e o f S m a d 4a f t e r o v e r e x p r e s s i o n o f N R 2F 2;B .T h e e x p r e s s i o n c h a n g e o f S m a d 4a f t e r k n o c k o u t o f N R 2F 2;C .T h e e x p r e s s i o n c h a n g e o f S m a d 4-r e g u l a t e d d o w n s t r e a m g e n e s a f t e r o v e r e x p r e s s i o n o f N R 2F 2;D.T h e e x p r e s -s i o n c h a n g e o f S m a d 4-r e g u l a t e d d o w n s t r e a m g e n e s a f t e r k n o c k o u t o f N R 2F 2.D i f f e r e n t c a p i t a l l e t t e r s i n d i c a t e e x t r e m e l y s i g-n i f i c a n t d i f f e r e n c e s (P <0.01);D i f f e r e n t l o w e r c a s e l e t t e r s i n d i c a t e s i gn i f i c a n t d i f f e r e n c e s (P <0.05).T h e s a m e a s b e l o w 图4 N R 2F 2基因对S m a d 4及下游基因表达的影响F i g .4 T h e e f f e c t s o f N R 2F 2g e n e o n t h e e x pr e s s i o n o f S m a d 4g e n e a n d d o w n s t r e a m g e n e s 3 讨 论N R 2F 2作为一种转录因子,在细胞的生命过程中起着重要的作用㊂啮齿类动物的遗传模型揭示了N R 2F 2基因在脂肪形成㊁脂质代谢㊁肝脏糖异生㊁胰岛素分泌和血压调节等生物过程中的重要作用[25-27]㊂本研究发现,在P K 15细胞中,N R 2F 2基因促进细胞增殖㊂N R 2F 2基因对P K 15细胞的生长促进作用是通过加速细胞周期从G 1期到S 期的进程来介导的㊂这些结果表明,N R 2F 2基因能促进细胞增殖㊂在之前的研究中,N R 2F 2已被证明在细胞的增殖中发挥促进作用㊂N R 2F 2在多种肿瘤细胞中高表达,促进细胞增殖,抑制细胞凋亡[28]㊂在肾细胞癌细胞中,下调N R 2F 2可通过上调B R C A 1来抑制细胞增殖并诱导细胞凋亡[29]㊂N R 2F 2通过调节I G F -1表达来调节小鼠出生后小脑的生长和84238期张万锋等:N R 2F 2基因调控猪P K 15细胞增殖和凋亡的研究A.干扰S m a d 4的效率;B .过表达N R 2F 2且干扰S m a d 4后下游基因表达的变化;C .过表达N R 2F 2且干扰S m a d 4后增殖基因表达的变化A.I n t e r f e r e n c e e f f i c i e n c y o f S m a d 4;B .T h e e x p r e s s i o n c h a n g e o f S m a d 4-r e g u l a t e d d o w n s t r e a m g e n e s a f t e r o v e r e x p r e s s i o n o f N R 2F 2a n d i n t e r f e r i n g w i t h S m a d 4;C .T h e e x p r e s s i o n c h a n g e o f p r o l i f e r a t i o n r e l a t e d g e n e s a f t e r o v e r e x p r e s s i o n o f N R 2F 2a n d i n t e r f e r i n g w i t h S m a d 4图5 挽救试验结果F i g .5 T h e r e s u l t o f r e s c u e e x pe r i m e n t 成熟[30]㊂过表达N R 2F 2基因可改变细胞粘附蛋白和细胞增殖的表达[31]㊂敲除N R 2F 2后,K i 67基因表达减少,细胞增殖被抑制,凋亡细胞增加[32]㊂N R 2F 2沉默后,细胞增殖和侵袭减少,细胞被阻断在G 1期[33]㊂可见,N R 2F 2基因在细胞增殖中起着重要作用,但其具体的作用机制还需进一步研究㊂N R 2F 2基因属于核受体转录因子家族㊂作为一种转录因子,N R 2F 2不直接调节激酶活性㊂N R 2F 2可能通过调节其他基因的表达来调节激酶活性㊂在不同的细胞模型中,E R α㊁C E B P /α㊁W n t 10b 已被证明受N R 2F 2的调控㊂在本研究中,通过蛋白质相互作用软件预测了一些与N R 2F 2相互作用的蛋白质,发现有15个蛋白可以与N R 2F 2相互作用,其中,S m a d 4比较值得注意㊂本研究采用C o -I P 直接验证了P K 15细胞中N R 2F 2蛋白与S m a d 4蛋白的结合㊂而过表达N R 2F 2可提高S m a d 4的水平㊂N R 2F 2的沉默可降低S m a d 4的水平㊂这说明N R 2F 2可能改变了S m a d 4基因的表达㊂S m a d 4为抑癌基因,属于S m a d 家族㊂转化生长因子-β(T G F -β)信号可以通过以下反应激活:1)T G F -β与受体结合,然后诱导S m a d 2/3磷酸化;2)共同介质S m a d 4与磷酸化的S m a d s 结合,然后迁移到细胞核;3)S m a d 复合物与各种转录因子相互作用,然后调节细胞的增殖和迁移㊂先前的研究表明,S m a d 4的缺失会影响细胞的增殖和迁移㊂敲除S m a d 4的细胞增殖较慢,细胞迁移减少[34-35]㊂m i R -144可以靶向S m a d 4基因,降低S m a d 4基因的表达,从而抑制细胞增殖和迁移[36]㊂S m a d 4基因表达水平的降低增加了对D N A 拓扑异构酶抑制剂的敏感性,促进细胞凋亡,从而抑制细胞增殖[37]㊂这些研究表明S m a d 4可以调节细胞周期㊂T G F -β信号通路在组织发育㊁稳态和修复中起着重要作用,并调控细胞增殖㊁分化㊁凋亡和迁移等细胞过程[38-39]㊂S m a d 4作为唯一的通用受体和T G F -β信号通路的中介,在将受体信号转导到核中的靶基因中起核心作用[40]㊂S m a d 4的异常表达可引起细胞中T G F -β超家族成员的异常变化,影响下游靶基因的表达,并影响细胞的生长发育㊂细胞增殖周期相关因子是T G F -β信号通路的下游效应因9423畜牧兽医学报54卷子[41]㊂研究发现,敲除N R2F2可以显著下调细胞周期相关基因的表达;而过表达N R2F2可显著上调细胞增殖周期相关基因的表达㊂挽救试验中过表达N R2F2和干扰S m a d4后,可以有效缓解过表达N R2F2基因的影响㊂这说明N R2F2通过与S m a d4相互作用,影响T G F-β信号通路,从而影响细胞增殖周期相关因子的表达㊂因此,发现N R2F2通过影响S m a d4基因的表达来影响细胞增殖㊂4结论过表达N R2F2基因后促进了细胞的增殖,抑制了凋亡,且N R2F2蛋白能与S m a d4蛋白结合并影响S m a d4基因的表达,进而影响下游基因的表达㊂参考文献(R e f e r e n c e s):[1] T A K E U C H I T,N A K AMU R A H.C e l l p r o l i f e r a t i o na n d d e v e l o p m e n t[J].D e v G r o w t h D i f f e r,2014,56(5):323.[2]J I A N G M Z,X U S,B A I M,e t a l.T h e e m e r g i n g r o l eo f M E I S1i n c e l l p r o l i f e r a t i o n a n d d i f f e r e n t i a t i o n[J].A m J P h y s i o l C e l l P h y s i o l,2021,320(3):C264-C269.[3] S U N Y,L I U Y,MA X L,e t a l.T h e i n f l u e n c e o f c e l lc y c l e r e g u l a t i o n o n c h e m o t h e r a p y[J].I n t J M o l S c i,2021,22(13):6923.[4] G U R S K A L M,AM E S K,G R I T S MA N K.S i g n a l i n gp a t h w a y s i n l e u k e m i c s t e m c e l l s[J].A d v E x p M e dB i o l,2019,1143:1-39.[5] N U S S E R,C L E V E R S H.W n t/β-C a t e n i n S i g n a l i n g,D i s e a s e,a n dE m e r g i n g T h e r a p e u t i c M o d a l i t i e s[J].C e l l,2017,169(6):985-999.[6] Z HA N G Y,WA N G X.T a r g e t i n g t h e W n t/β-c a t e n i ns i g n a l i n g p a t h w a y i n c a n c e r[J].J H e m a t o l O n c o l,2020,13(1):165.[7] L I U J Q,X I A O Q,X I A O J N,e t a l.W n t/β-c a t e n i ns i g n a l l i n g:f u n c t i o n,b i o l o g i c a l m e c h a n i s m s,a n dt h e r a p e u t i c o p p o r t u n i t i e s[J].S i g n a l T r a n s d u c tT a r g e t T h e r,2022,7(1):3.[8] S P R I N Z A K D,B L A C K L OW S C.B i o p h y s i c s o fN o t c h s i g n a l i n g[J].A n n u R e v B i o p h y s,2021,50:157-189.[9] WA N G H F,Z A N G C Z,L I U X S,e t a l.T h e r o l e o fN o t c h r e c e p t o r s i n t r a n s c r i p t i o n a l r e g u l a t i o n[J].JC e l l P h y s i o l,2015,230(5):982-988.[10] T Z A V L A K I K,MO U S T A K A S A.T G F-βs i g n a l i n g[J].B i o m o l e c u l e s,2020,10(3):487.[11] P E N G D D,F U M Y,WA N G M N,e t a l.T a r g e t i n gT G F-βs i g n a l t r a n s d u c t i o n f o r f i b r o s i s a n d c a n c e rt h e r a p y[J].M o l C a n c e r,2022,21(1):104.[12] C O O N E Y A J,T S A I S Y,O'MA L L E Y B W,e t a l.C h i c k e n o v a l b u m i n u p s t r e a m p r o m o t e r t r a n s c r i p t i o nf a c t o r(C O U P-T F)d i m e r s b i n d t o d i f f e r e n t G G T C Ar e s p o n s e e l e m e n t s,a l l o w i n g C O U P-T F t o r e p r e s sh o r m o n a l i n d u c t i o n o f t h e v i t a m i n D3,t h y r o i dh o r m o n e,a n d r e t i n o i c a c i d r e c e p t o r s[J].M o l C e l lB i o l,1992,12(9):4153-4163.[13] K L E P S C H V,H E R MA N N-K L E I T E R N,B A I E R G.B e y o n dC T L A-4a n d P D-1:o r p h a n n u c l e a r r e c e p t o rN R2F6a s T c e l l s i g n a l i n g s w i t c h a n d e m e r g i n g t a r g e ti n c a n c e r i mm u n o t h e r a p y[J].I m m u n o l L e t t,2016,178:31-36.[14] P O L V A N I S,P E P E S,M I L A N I S,e t a l.C O U P-T F I Ii n h e a l t h a n d d i s e a s e[J].C e l l s,2020,9(1):101.[15] MA U R I F,S C H E P K E N S C,L A P O U G E G,e t a l.N R2F2c o n t r o l s m a l i g n a n t s q u a m o u s c e l l c a r c i n o m as t a t e b y p r o m o t i n g s t e m n e s s a n d i n v a s i o n a n dr e p r e s s i n g d i f f e r e n t i a t i o n[J].N a t C a n c e r,2021,2(11):1152-1169.[16] X I A B L,HO U L J,K A N G H,e t a l.N R2F2p l a y s am a j o r r o l e i n i n s u l i n-i n d u c e d e p i t h e l i a l-m e s e n c h y m a lt r a n s i t i o n i n b r e a s t c a n c e r c e l l s[J].B M C C a n c e r,2020,20(1):626.[17] L I L P,X I E X,Q I N J,e t a l.T h e n u c l e a r o r p h a nr e c e p t o r C O U P-T F I I p l a y s a n e s s e n t i a l r o l e i na d i p o g e n e s i s,g l u c o s e h o m e o s t a s i s,a n d e n e r g ym e t a b o l i s m[J].C e l l M e t a b,2009,9(1):77-87. [18] X U Z,Y U S T,H S U C H,e t a l.T h e o r p h a n n u c l e a rr e c e p t o r c h i c k e n o v a l b u m i n u p s t r e a m p r o m o t e r-t r a n s c r i p t i o n f a c t o r I I i s a c r i t i c a l r e g u l a t o r o fa d i p o g e n e s i s[J].P r o c N a t l A c a d S c i U S A,2008,105(7):2421-2426.[19] C H E N G X.R o l e s o f v i t a m i n A m e t a b o l i s m i n t h ed e v e l o p m e n t o f h e p a t i c i n s u l i n r e s i s t a n c e[J].I S R NH e p a t o l,2013,2013:534972.[20] O K AMU R A M,K U D O H,WA K A B A Y A S H I K I,e ta l.C O U P-T F I I a c t s d o w n s t r e a m o f W n t/β-c a t e n i ns i g n a l t o s i l e n c e P P A Rγg e n e e x p r e s s i o n a n d r e p r e s sa d i p o g e n e s i s[J].P r o c N a t l A c a d S c i U S A,2009,106(14):5819-5824.[21] L E N G X,C O O N E Y A J,T S A I S Y,e t a l.M o l e c u l a rm e c h a n i s m s o f C O U P-T F-m e d i a t e d t r a n s c r i p t i o n a l05238期张万锋等:N R2F2基因调控猪P K15细胞增殖和凋亡的研究r e p r e s s i o n:e v i d e n c e f o r t r a n s r e p r e s s i o n a n d a c t i v er e p r e s s i o n[J].M o l C e l l B i o l,1996,16(5):2332-2340.[22] MA L,HU A N G M,L I A O X H,e t a l.N R2F2r e g u l a t e s c e l l p r o l i f e r a t i o n a n d i mm u n o m o d u l a t i o n i nW h a r t o n s'j e l l y s t e m c e l l s[J].G e n e s(B a s e l),2022,13(8):1458.[23] Q I N J,WU S P,C R E I G H T O N C J,e t a l.C O U P-T F I Ii n h i b i t s T G F-β-i n d u c e d g r o w t h b a r r i e r t o p r o m o t ep r o s t a t e t u m o r i g e n e s i s[J].N a t u r e,2013,493(7431):236-240.[24] WA N G C Q,Z HO U Y,R U A N R Y,e t a l.H i g he x p r e s s i o n of C O U P-T F I I c o o p e r a t e d w i t h n eg a t i v eS m a d4e x p r e s s i o n p r e d i c t s p o o r p r o g n o s i s i n p a t i e n t sw i t h c o l o r e c t a l c a n c e r[J].I n t J C l i n E x p P a t h o l,2015,8(6):7112-7121.[25] WA N G H,N I E L,WU L,e t a l.N R2F2i n h i b i t sS m a d7e x p r e s s i o n a n d p r o m o t e s T G F-β-d e p e n d e n te p i t h e l i a l-m e s e n c h y m a l t r a n s i t i o n of C R C v i at r a n s a c t i v a t i o n o f m i R-21[J].B i o c h e n B i o p h y s R e sC o mm u n,2017,485(1):181-188.[26] X U Q J,J I A N G S J.m i R-194-5p s e r v e s a s u p p r e s s i v er o l e i n h u m a n k e l o i d f i b r o b l a s t s v i a t a r g e t i n g N R2F2[J].M o l M e d R e p,2021,23(1):57.[27] S A J I N O V I C T,B A I E R G.N e w i n s i g h t s i n t o t h ed i ve r s ef u n c t i o n s o f t h e N R2F n u c l e a r o r p h a nr e c e p t o r f a m i l y[J].F r o n t B i o s c i(L a n d m a r k E d),2023,28(1):13.[28] Q I N J,T S A I S Y,T S A I M J.T h e c r i t i c a l r o l e s o fC O U P-T F I I i n t u m o r p r o g r e s s i o n a n d m e t a s t a s i s[J].C e l l B i o s c i,2014,4(1):58.[29] Z H E N G J,Q I N W J,J I A O D,e t a l.K n o c k d o w n o fC O U P-T F I I i n h i b i t s c e l l p r o l i f e r a t i o n a n d i n d u c e sa p o p t o s i s t h r o u g h u p r e g u l a t i n g B R C A1i n r e n a l c e l lc a r c i n o m a c e l l s[J].I n t J C a n c e r,2016,139(7):1574-1585.[30] K I M B J,T A K AMO T O N,Y A N J,e t a l.C h i c k e nO v a l b u m i n U p s t r e a m P r o m o t e r-T r a n s c r i p t i o n F a c t o rⅡ(C O U P-T F I I)r e g u l a t e s g r o w t h a n d p a t t e r n i n g o ft h e p o s t n a t a l m o u s e c e r e b e l l u m[J].D e v B i o l,2009,326(2):378-391.[31] B O U D O T A,L E D I L Y F,P A K D E L F.I n v o l v e m e n to f C O U P-T F s i n c a n c e r p r o g r e s s i o n[J].C a n c e r s(B a s e l),2011,3(1):700-715.[32] Y U C T,T A N G K,S UH J M,e t a l.C O U P-T F I I i se s s e n t i a lf o r m e t a n e p h r i c m e s e n c h y m e f o r m a t i o n a n dk i d n e y p r e c u r s o r c e l l s u r v i v a l[J].D e v e l o p m e n t,2012,139(13):2330-2339.[33] P O L V A N I S,T A R O C C H I M,T E M P E S T I S,e t a l.N u c l e a r r e c e p t o r s a n d p a t h o g e n e s i s o f p a n c r e a t i cc a n c e r[J].W o r ld J G a s t r oe n t e r o l,2014,20(34):12062-12081.[34] S Y E D V.T G F-βs i g n a l i n g i n c a n c e r[J].J C e l lB i o c h e m,2016,117(6):1279-1287.[35] H E R N A N D A P Y,C H E N K,D A S A M,e t a l.S MA D4e x e r t s a t u m o r-p r o m o t i n g r o l e i nh e p a t o c e l l u l a r c a r c i n o m a[J].O n c o g e n e,2015,34(39):5055-5068.[36] S H E N G S H,X I E L,WU Y Y,e t a l.M i R-144i n h i b i t s g r o w t h a n d m e t a s t a s i s i n c o l o n c a n c e r b yd o w n-re g u l a t i n g S MA D4[J].B i o s c i R e p,2019,39(3):B S R20181895.[37] HA E G E R S M,T HOM P S O N J J,K A L R A S,e t a l.S m a d4l o s s p r o m o t e s l u n g c a n c e r f o r m a t i o n b u ti n c r e a s e s s e n s i t i v i t y t o D N A t o p o i s o m e r a s e i n h i b i t o r s[J].O n c o g e n e,2016,35(5):577-586. [38] Z H A O H D,W E I J,S U N J.R o l e s o f T G F-βs i g n a l i n gp a t h w a y i n t u m o r m i c r o e n v i r i o n m e n t a n d c a n c e r t h e r a p y[J].I n t I mm u n o p h a r m a c o l,2020,89:107101.[39]J A HN S C,L AW M E,C O R S I N O P E,e t a l.T G F-b e t a a n t i p r o l i f e r a t i v e e f f ec t s i n t u m o r s u p p r e s s i o n[J].F r o n t B i o s c i(S c h o l E d),2012,4(2):749-766.[40] L I U J Q,Y U A N B,C A O J,e t a l.AM B R A1p r o m o t e sT G Fβs i g n a l i n g v i a n o n p r o t e o l y t i c p o l y u b i q u i t y l a t i o no f S m a d4[J].C a n c e r R e s,2021,81(19):5007-5020.[41] L I U L,L I Q Q,Y A N G L,e t a l.S m a d4f e e d b a c ka c t i v a t e s t h e c a n o n i c a l T G F-βf a m i l y s i g n a l i n gp a t h w a y s[J].I n t J M o l S c i,2021,22(18):10024.(编辑郭云雁)1523。
鹿场环境中细菌的分布及耐药性调查

d546 =G9#PQ546 M7-I!-9-4KCJ$->QC4 49$!W7-JKN _$- P5Q46 L$QCc$9-!@$-LK4 ;#-I!GF9-"
落 计 数 以 菌 落 形 成 单 位 ",#D#-ER#U:$-I F-$HV! =ON#计 数 ' %'0'*!菌落形态观 察 及 分 离 纯 化 ! ! 将 上 述 培 养 的细菌进行菌落形 态 观 察!用 接 菌 环 挑 出 形 态 差 异
较大的菌落!接种于含 48 培养基的试 管中!放入 恒 温振荡器培养?G!温度 为 */ ^' 培 养 后 进 行 革 兰 氏染色!镜检出现不 同 细 菌 形 态 后 继 续 划 线 分 离 纯
J7$,9公司&数 码 凝 胶 图 像 处 理 系 统 T9-#-!%(&& 购 自上海将来实业有限公司'
#I!! 方 法 %'0'%!样品采集! ! 样 品 采 集 于 辽 宁 铁 岭 西 丰 县 某 鹿 场 ' 无 菌 采 集 鹿 场 精 饲 料 (粗 饲 料 (粪 便 及 水 分
别置于已灭 菌 的 %& :J 离 心 管 中!做 好 标 记 备 用' 将已灭菌的)个 J8 培养基平 皿分 别放 置 在 公 鹿 圈 角(公 鹿 圈 中(公 鹿 食 槽 及 母 鹿 圈 中' 开 盖 放 置
据报 道!养 殖 场 的 饲 料 及 饮 水 最 易 受 到 一 些 病 原菌的污染 ' *0!)+ 近年来!国内外学者对猪 舍(牛 舍( 鸡鸭舍及兔舍内病原微生物的污染情况进行了检测 及研究*2!B+!而对 鹿 舍 内 微 生 物 的 分 布 还 鲜 有 报 道' 本 研 究 主 要 对 鹿 场 中 精 饲 料 (粗 饲 料 (水 及 空 气 中 细 菌的分布情况进行 了 检 测!同 时 对 鉴 定 出 的 致 病 菌 和条件致病菌进行 了 药 敏 试 验!以 期 为 鹿 场 细 菌 性 疾病的防制提供有利依据'
举办全国第五届动物生物技术学术年会

举办全国第五届动物生物技术学术年会
经费申请
尊敬的孙琪信校长、科研院有关领导
由农业生物技术国家重点实验室主办的全国动物生物技术学会换届与第五次学术年会定于2010年10月17-20日在我校召开。
会议邀请了国内外动物生物技术领域资深、著名专家与学者,所邀请的大部分报告人近期均在国际著名杂志《SCIENCE》或《NATURE》发表过重要学术论文;恰逢转基因重大专项启动2周年之际,本次年会将成为展示现阶段的重要成果与成就,推动生物技术产业的进步与发展的一次重要、世界级学术会议,对提高学会以及我校知名度将起到重要作用。
会议期间将邀请国家科技部主要领导到会,并作大会报告,介绍我国动物生物技术的现状与发展趋势。
会议规模初步确定为300人,注册费900元/人,学生600元/人。
参会人员住宿费自理,大会提供餐饮、会议室租赁、论文集印刷、突出贡献科学家奖、青年科学家奖、优秀论文与优秀墙报奖励、邀请专家等其他费用。
共需经费77万元,会议收入22.5万元,缺口55万元。
本次学术年会得到了农业生物技术国家重点实验室、动物科技学院、北京市奶牛育种中心、广州温氏集团、青岛森淼集团等单位大力支持,达成意向的资助金额为40万元,特申请资助金额15万元,望予以批准。
礼
第五次动物生物技术学术年会筹委会
农业生物技术国家重点实验室
2010.4.3。
雅士勇资料

一、提高种猪受胎率
种公猪
1、提高公猪活力及精子的直线运动能力 2、加快精子成熟 3、增强种公猪的射精力度及数量 4、提高受胎率
种母猪
1、促进卵泡发育、成熟,排卵数量,降低返情率 2、保持母猪高昂代谢率,提高卵子与精子的结合
能力,提高着床能力、产仔数,降低弱仔数
雅士勇作用原理
脂肪酸
左旋肉碱
(精子、卵子的能量源)
发情 5 6 6 6 7 6 7 7 6 5 6 6 5 5 6 5 7 6 6 5 5.9 间隔 期(天)
返情 √ √ √ √ √ √ √ √ √ √ х √ √ √ √ √ √ √ √ √ 5%
实验组1
编号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 平均
短链、中长链
脂肪
左 旋
赖氨酸甲基化合成
肉
碱
进入细胞线粒体进行氧化能量释放
初生、高产动物需求量远高出自身合成量
雅士勇主要成分
每升含:
左旋肉碱 5,000mg
VB1
2,000mg
VB6
1,000mg
VB12 15mg
Vpp
10,000mg
泛酸
2,000mg
天门冬氨酸 麦氨酸 甘氨酸 蛋氨酸 山梨聚醣醇
雅士勇主要成分
每升含:
左旋肉碱 5,000mg 天门冬氨酸 10,000mg
VB1
2,000mg 麦氨酸 10,000mg
VB6
1,000mg 甘氨酸 10 ,000mg
VB12 15mg
蛋氨酸 20,000mg
Vpp
10,000mg 山梨聚醣醇 100,000mg
关于举办第五次动物生物技术学术研讨会会议通知

关于举办第五次动物生物技术学术研讨会会议通知
佚名
【期刊名称】《中国农业科技导报》
【年(卷),期】2010(0)4
【总页数】1页(P120-120)
【关键词】动物生物技术;学术研讨会;农业生物技术;生物技术领域;生物技术产业;学术交流;技术人员;转基因
【正文语种】中文
【中图分类】S188
【相关文献】
1.中国社会科学情报学会第五次全国会员代表大会暨"信息资源的利用与管理"学术研讨会会议通知 [J],
2.中国农业生物技术学会动物生物技术分会第二届全国动物生物技术学术研讨会筹委会 [J],
3.关于召开第五次蜜源植物与蜜蜂授粉专业委员会工作会议暨学术研讨会的通知[J], 中国养蜂学会
4.“全国首届动物生物技术学术研讨会暨中国农业生物技术学会动物生物技术分会成立大会”的通知 [J],
5.第六届胃肠肿瘤学术研讨会暨胃癌高级培训班广东省医学会胃肠外科学分会第五次学术会议通知 [J],
因版权原因,仅展示原文概要,查看原文内容请购买。
动物分子生物学结课论文

动物分子生物学结课论文院系:生命科学学院专业:动物学学号:0000000姓名:0000002015年6月20日目录摘要 (1)关键词 (1)Abstract (1)Keywords (1)一绿色荧光蛋白及其发光机理 (1)二GFP基因标记的方式 (2)三GFP的功能 (2)四GFP基因在分子生物学上的应用 (3)五表达绿色荧光蛋白的重组减毒鼠伤寒沙门氏菌的构建与鉴定 (3)六GFP的应用前景 (4)参考文献 (5)绿色荧光蛋白基因的发现与发展摘要来源于海洋多管水母属的绿色荧光蛋白(GFP)基因是一种能够在细胞内稳定表达,在蓝光或长紫外光的激发下,不需要任何反应底物及其他辅助因子就能发出绿色荧光的新型报告基因,无种属、组织和位置特异性,且能监测基因表达、信号转导、共转染、蛋白运输与定位,以及细胞系谱分类等。
GFP对细胞无毒性,且检测方法简单,结果真实可靠,目前在多种原核和真核生物研究中得到广泛的应用。
本文就GFP的生化特性、GFP的改进及其在分子生物学研究中的应用潜力进行简要阐述。
关键词:绿色荧光蛋白;功能;应用Discovery and development of green fluorescent protein geneAbstractThe green fluorescent protein (GFP) from the sea genus Aequorea can stably expressed in cells and can send the new green fluorescent reporter gene without any reaction substrates and other cofactors under ultraviolet excitation blue or long, it no species, organization and location-specific, and can monitor gene expression, signal transduction, co-transfection, protein transport and localization, and cell lineage classification. GFP not toxic to cells, and the methods is simple to detect the GFP, the dectecttion results are true and reliable, now, it has been widely used in a variety of prokaryotic and eukaryotic studies. Article on the biochemical characteristics, improvement of GFP, and its potential in molecular biology briefly described. Keywords: green fluorescent protein; function; application一绿色荧光蛋白及其发光机理绿色荧光蛋白(Green Fluoeescent Prootein,简称GFP)是一种在美国西北海岸所盛产的水母中所发现的一种蛋白质。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
10:10-10:50
Prof.Bing Lim
Harvard Medical School, Genome Institute ofSingapore,USA
Title:T-cell factor 3 regulates embryonic stem cell pluripotency and self-renewal by the transcriptional control of multiple lineage pathways.
17:40ห้องสมุดไป่ตู้18:10
学生
题目:
10月19日
大会主题报告:动物重大疫病分子生物学诊断与防治
主持人:待定
08:30-9:10
Prof.Peter Kaiser
IAH,Institute for Animal Health,United Kingdom, UK
Title:Immunological regulation of disease resistance in chicken
姜运良 教授
山东农业大学
题目:猪肉用性状基因启动子的多态性及功能
16:00—16:10
茶歇
16:10—16:40
才学鹏研究员
中国农业科学院兰州兽医研究所
题目:生物技术—兽用生物制品行业腾飞的翅膀
16:40—17:10
赵要风 教授
中国农业大学生物学院
题目:待定
17:10—17:40
Yuguang Zhao
题目:禽类及中华鳖胚胎原始生殖细胞的迁移以及异种嵌合体的制备
16:00—16:10
茶歇
16:10—16:40
连正兴 教授
中国农业大学
题目:
16:40—17:10
欧阳红生
吉林大学
题目:抗猪瘟转基因猪研究
17:10—17:40
周欢敏 教授
内蒙古农业大学生物工程学院
题目:
17:40—18:10
学生
题目:
大会分组报告:功能基因与分子育种及重大疾病控制
11:30-12:10
曾溢滔院士
上海交通大学医学院
题目:动物克隆与胚胎发育
大会分组报告:动物胚胎与干细胞工程
主持人:待定
13:30—14:00
谭景和 教授
山东农大动科学院
题目:待定
14:00—14:30
孙青原 研究员
中国科学动物研究所
题目:哺乳动物卵子减数分裂染色体分离调节机制
14:30—15:00
9:10-9:50
Prof.Gustavo Aguirre
School of Veterinary Medicine,University of Pennsylvania,USA
Title:Inherited Retinal Degenerations in the Dog: From Gene Discovery to Gene Therapy
第五届全国动物生物技术分会学术年会
大会报告日程
日期
时间
题目及报告人
报告内容
10月18日
大会主题报告:动物转基因与干细胞工程
主持人:李宁 院士
9:30-10:10
Prof.James M. Robl
Dept. of Veterinary & Animal Sciences,UniversityofMassachusetts,Amherst,MA,USA
16:40—17:10
潘登科研究员
中国农业科学院北京畜牧兽医研究所
题目:体细胞克隆研制敲除MSTN基因和GGTA1基因猪
17:10—17:40
雷安民 副教授
西北农林科技大学
题目:哺乳动物卵母细胞成熟调控的研究进展
17:40—18:10
学生
题目:
大会分组报告:功能基因与分子育种及重大疾病控制
李奎 教授
中国农业科学院北京畜牧兽医研究所
题目:Detection, tracing and environmental risk assessment of transgenic pig
15:00—15:30
陈静波
新疆畜牧科学院
题目:马繁殖技术初步研究
15:30—16:00
李赞东 教授
中国农业大学
The Division of Structural Biology,The Wellcome Trust Centre for Human Genetics,OxfordUniversity.
题目:Manipulation of Viral microRNAs for Novel Vaccines
吴珍芳 教授
华南农业大学
题目:木聚糖酶基因克隆及其在转基因育种上的研究
15:00—15:30
李子义 教授
吉林大学农学部畜牧兽医学院
题目:转基因克隆雪貂的研究进展
15:30—16:00
曾文先 教授
西北农林科技大学
题目:
16:00—16:10
茶歇
16:10—16:40
杨利国 教授
华中农业大学
题目:激素基因免疫研究进展
10:50-11:30
Prof.George E.Seidel,Jr
Department of Biomedical Sciences,Animal Reproduction and Biotechnology Laboratory,Colorado State University,USA
Title:
主持人:待定
13:30—14:00
方美英老师
题目:
14:00—14:30
孙效文研究员
中国水产科学研究院黑龙江水产研究所
题目:鲤鱼多个家系的QTL结果与杂种优势分析
14:30—15:00
张胜利研究员
北京奶牛中心
题目:奶牛遗传改良技术进展
15:00—15:30
赵志辉 教授
吉林大学
题目:
15:30—16:00
题目:免疫耐受疫苗开发和潜在的临床应用
大会分组报告:动物胚胎与干细胞工程
主持人:待定
13:30—14:00
李光鹏 教授
内蒙古大学实验动物研究中心 呼和浩特 010021
题目:体细胞克隆牛存在的问题及可能采取的对策
14:00—14:30
石德顺 教授
广西大学
题目:水牛干细胞的研究进展
14:30—15:00
9:50-10:30
陈化兰 研究员
中国农科院哈尔滨兽医研究所国家禽流感参考实验室
题目:H5N1亚型禽流感病毒的进化
10:30-10:40
茶歇
10:40-11:20
张改平院士
河南省农科院动物免疫学重点实验室
题目:快速检测试纸与动物疫病防控
11:20-12:00
王宾 教授
中国农业大学农业生物技术国家重点实验室