小麦籽粒淀粉生物合成及淀粉分支酶基因(SBE-IIb)的克隆与表达
小麦淀粉合酶基因Ⅱ克隆及反义和RNAi载体构建

酶 【 。S s 主 要 参 与 了 支 链 淀 粉 中分 支 链 的 合 3 S则 】 成, 它有许 多同工酶 。如玉 米 的 S s 为 zS 、S Ⅱ S分 S IzS f l 和 zS b 马铃 薯 的 S s 至少 有 S 、 S 和 S SⅡ , S也 IS Ⅱ SⅢ 3
种[ 引。每 一种 可溶 性 S s 支链淀 粉 的合成 中也发 S在
s nh s ,G S ) 可 溶 性 淀 粉 合 酶 ( ou l sac y tae B S 与 S lbe tr h
( DP—guoe A lcs ,AD P—G c 和 A P, 中 AD l) D 其 P—
3 5的长 链 合 成 , S 在 中等 长 度 的 支链 淀 粉 合 而 SI I 成 中起 着其 它 s S所不 能替代 的作用 [l 5。 但 有关 小麦 中 3种 类 型 S s 自的功 能研 究 尚 S各
一
挥 着 不 同作 用 。玉米 的 S I DP P lmei t n S与 ( oy r ai , z o 葡 糖 聚体 ) 6—1 5的短链 合成有 关 ,SⅡf 与 了 D S l 参 P
>2 4的长链 合成 ; 马铃 薯 的 s Ⅲ催 化 了与 D 2 s P 5—
磷 酸 ( 一1一P) A P 转 变 成 AD 葡 萄 糖 G 和 T P
茎 的 主要 组成 部分 。作物 中淀粉 由直链 淀 粉 ( Amy — le和支链淀 粉 ( o) s Amy p ci) 大 类 组 成 。其 合 l et 两 o n
成过 程 主 要 在 A G 焦 磷 酸 化 酶 ( eo ie5 DP Adn s n
dp op aeguoep rp op oyae AG ae 、 i sh t lcs yo h sh rl , P s ) 淀 h s
小麦淀粉合成酶研究进展

关 键 词 : 小麦 淀粉 ; 关 键酶 ; 淀 粉合 成酶 ; 表 达调 控 中图分类号 :
S51211
文献标识码 :
A
小麦籽粒主要由淀粉 ( 约占干重的 65 %) 、蛋 白质 ( 10 % ~ 15 %) 、水 分 ( 13 % ~ 15 %) 、脂 类 (2 %) 、粗纤 维 ( 3 %) 和 灰分 ( 1 %) 等组 成 . 小 麦淀粉的用途非常广泛 , 不仅可以制作各种各样的 面包 、馒头 、饼干 、蛋糕、通心粉、面条 , 还可用 于造纸 和纺织 工业 , 作 为原 料生产 葡萄糖 浆 、胶 带 , 经过修饰以后 , 其用途更加广泛 . 淀粉的合成 机理 、合成途径 、合成过程中的酶及其作用机制比 较复杂 , 目前尚不完全清楚 . 一般认为 , 小麦籽粒 淀粉由胚乳中的双层膜结构的淀粉质体合成 , 需要 以下几种关键酶 : ADP - 葡萄糖焦磷酸化酶 , 淀粉 合成酶 、淀粉分支酶和淀粉脱分支酶 . 从合成途径 看 , 所有 参与淀 粉合成的 酶都会 影响淀 粉粒的 结 构 . 对国内外小麦淀粉合成酶研究方面的进展进行 了综述 , 希望能对该方面的研究有所启发 .
(山东省农业科学院作物研究所 , 山东 济南 250100)
摘 要 : 针 对目 前有 关淀 粉理 化特 性方 面的 研究 相对 落后 的状 况 , 对相 关文 献进 行分 析 。首先分 析小麦籽粒 淀
粉需要的几种关键酶 : ADP - 葡萄糖 、淀粉合成酶 、淀粉 分支酶 和淀粉脱 分支酶 , 其次对国 内外小 麦淀粉 合成 酶研究方面的进展进行了综述 。
E2mail : so ngjmsd @ho tmail , co m.
8
78
湖南农业大学学报 (自然科学版)
2007 年 8 月
小麦粒重形成的分子调控机制研究综述

小麦粒重形成的分子调控机制研究综述魏玮;郭嘉莲;万琳涛;徐林峰;丁明全;周伟【摘要】粒重是小麦Triticum aestivum产量构成的三大要素之一,是由多基因控制的数量性状,极易受环境因素的影响。
国内外学者围绕粒重形成的遗传特征和分子调控机制进行了大量研究,也取得了一些研究进展。
如何高效地利用前人的研究成果进行不断创新以提高小麦单产是育种工作者重要的研究课题。
围绕小麦粒重形成的构成要素、遗传特征、QTLs(quantitative trait loci,QTLs)遗传定位、籽粒质量形成候选基因的克隆与分子调控机制解析等方面的最新研究展开综述;同时总结了以往研究过程中存在的问题,并结合自身研究对研究前景进行分析后指出:为了从分子水平上全面阐明小麦粒重形成的调控机制,后续研究首先应在小麦粒重形成的关键时期对籽粒激素的变化特征进行全面分析;其次要利用全基因组关联分析和高通量测序技术进一步开发与性状密切相关联的SNP标记(single nucleotide polymorphism, SNP);最后结合作图群体进行表型与SNP分析技术为基础的基因型关联性分析,对粒重形成候选主效基因进行精细定位、图位克隆和功能解析。
表1参52%Grain weight (GW), a quantitative trait determined by several genes, is one of three key wheat yield components and is sensitive to environmental variation. Researchers at home and abroad have done many stud-ies on the genetic traits and molecular regulation mechanism of GW formation, and have made some research progress. How to make innovations to improve unit yield of wheat basedon the existing research is an important work for the researchers. In this study, the latest research progresses on components, genetic traits, genetic mapping of QTLs (quantitative trait loci, QTLs), candidate genecloning and molecular regulation of GW in wheat were reviewed, and then the problems of existing research and the future research direction involved in our recent research were also sorted out and discussed. In order to get a comprehensive knowledge about the molecular mechanism on the production of GW, researchers should pay more attention to three key items. First-ly, the variation of hormone in the development of GW should be concerned and measured. Secondly, GWAS (genome-wide association study, GWAS) together with high throughput sequencing should be adopted to exploit SNP (single nucleotide polymorphism, SNP ) markets conferring to some special agricultural trait. Finally, the candidate gene should be mapped based on SNP marker-assisted selection in the mapping population, and then the candidate genes should be cloned and their function in the regulatory network involved in the formation of GW should be covered. This work will provide a good reference for further research.【期刊名称】《浙江农林大学学报》【年(卷),期】2016(033)002【总页数】9页(P348-356)【关键词】作物遗传育种;小麦;粒重;产量;分子调控;综述【作者】魏玮;郭嘉莲;万琳涛;徐林峰;丁明全;周伟【作者单位】浙江农林大学农业与食品科学学院/浙江省农产品品质改良技术研究重点实验室,浙江临安311300;浙江农林大学农业与食品科学学院/浙江省农产品品质改良技术研究重点实验室,浙江临安311300;浙江省开化县农作物技术推广站,浙江开化 324300;浙江勿忘农种业科学研究院有限公司,浙江杭州310020;浙江农林大学农业与食品科学学院/浙江省农产品品质改良技术研究重点实验室,浙江临安311300;浙江农林大学农业与食品科学学院/浙江省农产品品质改良技术研究重点实验室,浙江临安311300【正文语种】中文【中图分类】S512.1小麦Triticum aestivum是重要的粮食作物。
“七大农作物育种”重点专项成果推荐

“七大农作物育种”重点专项成果推荐编者按:“七大农作物育种”重点专项(以下简称“育种专项”)按照“加强基础研究、突破前沿技术、创制重大品种、引领现代种业”的总体思路,依据总体目标部署了优异种质资源鉴定与利用、主要农作物基因组学研究、育种技术与材料创新、重大品种选育、良种繁育与种子加工五大任务,贯通基础研究、共性关键技术研究、集成示范3个层面。
项目实施5年来,育种专项在基础研究、新材料创制及新品种选育等研究成果的转化应用、人才培养及平台建设等方面成效显著,在生物技术平台开发、种质资源创制与鉴定、优良基因挖掘和产量性状形成的分子基础等领域均取得了一系列突破性研究进展,形成了一批有重要应用前景的突破性研究成果。
单倍体育种高效技术体系构建与应用单倍体育种技术体系为国内首创。
玉米方面,率先精细定位并独立克隆了两个玉米单倍体诱导关键基因,发明了诱导系分子育种方法,育成系列高频单倍体诱导系;首次提出基于籽粒油分鉴别单倍体的技术原理,实现了单倍体鉴别的自动化,开辟了鉴别技术新途径;建立了工程化纯系生产技术体系,突破单倍体加倍技术瓶颈,打破传统选系模式,并在全国实现规模化应用。
油菜方面,国际上首次创制出具有双单倍体诱导特性八倍体油菜诱导系,表现出替代油菜传统的小抱子培养技术的前景,在油菜及十字花科蔬菜亲本创制、品种选育、远缘杂交、基因编辑中广泛应用。
小麦方面,建立了基于小麦玉米杂交的小麦双单倍体高效生产技术规程,年产小麦双单倍体不少于2万个,突破了从小麦玉米杂交“单倍体产生”到“单倍体生产”的系列关键技术瓶颈,成功应用于小麦温光敏核不育系、恢复系、常规品种的高效创制及双单倍体群体构建等。
蔬菜方面,建立了大白菜游离小抱子规模化培养技术,创制了高维生素C、高p胡萝卜素的桔红心和高花青苷的紫色叶球双单倍体系300余份,获得抗性等综合性状优良的优势材料8份。
玉米单倍体育种技术体系是我国具有完全自主知识产权的研究成果,该技术的应用激发了我国种业研发的活力,所选育的杂交种得到大面积推广,创造了巨大的经济社会利益。
EMS 诱变技术在小麦上的应用

EMS诱变技术在小麦上的应用曹亚萍∗ꎬ武银玉ꎬ范绍强ꎬ张凤琴ꎬ连㊀晋ꎬ高㊀炜(山西省农业科学院小麦研究所ꎬ临汾041000)摘㊀要:小麦遗传基础日趋狭窄成为小麦遗传改良的瓶颈ꎮ甲基磺酸乙酯(EMS)是一种高效稳定的烷化类诱变剂ꎬ能诱发产生高密度系列等位基因点突变ꎬ可在创造作物新品种㊁新种质㊁遗传材料以及解决育种工作中某些特殊问题等方面取得突破ꎮ本文从EMS诱变的原理和特点着手ꎬ简述了小麦EMS诱变及突变体鉴选方法ꎬ绘制了EMS诱变研究备选方案图ꎬ为小麦科研工作者提供研究思路和参考依据ꎮ同时对EMS诱变技术在抗病基因克隆㊁品质性状改良㊁叶片持绿机制研究㊁农艺性状解析㊁突变体库构建等方面的研究进展进行了前沿报道ꎬ分析了EMS诱变在小麦研究中存在的问题及应对方法ꎬ并在此基础上对EMS诱变技术在小麦上的发展前景进行展望ꎮ这对于丰富小麦遗传资源㊁加快育种进程和开展基因功能研究具有重要意义ꎮ关键词:小麦ꎻ甲基磺酸乙酯ꎻ化学诱变ꎻ突变体中图分类号:S512ꎻQ813.5文献标志码:ADOI:10.3969/j.issn.1007 ̄7146.2019.05.002ApplicationofInductionTechnologywithEMSonWheatCAOYaping∗ꎬWUYinyuꎬFANShaoqiangꎬZHANGFengqinꎬLIANJinꎬGAOWei(InstituteofWheatResearchꎬShanxiAcademyofAgriculturalScienceꎬLinfen041000ꎬChina)Abstract:Oneofthebottlenecksforwheatimprovementisthenarrowingofwheatgeneticbasis.Ethylmethylsulfonate(EMS)isanefficientandstablealkylationmutagensꎬwhichcaninduceaseriesofallelemutationswithhighdensityꎬandmakebreakthroughsincreatingofnewvarietiesꎬnewgermplasmꎬgeneticmaterialsandsolvingsomespecialprob ̄lemsinbreeding.BasedontheprinciplesandcharacteristicsofEMSmutagenesisꎬthispaperbrieflydescribesthemeth ̄odofEMSinductionandmutagenesisselectionforwheatꎬanddrawsanalternativeschemeofEMSmutagenesisre ̄searchꎬwhichprovidesresearchideasandreferenceforwheatresearchers.AtthesametimeꎬtheresearchprogressofEMSmutagenesistechnologyindiseaseresistancegenecloningꎬqualitytraitimprovementꎬleafgreeningmechanismre ̄searchꎬagronomictraitanalysisꎬandmutantlibraryconstructionwasreported.TheproblemsandcountermeasuresofEMSmutagenesisinwheatresearchwereanalyzed.ThedevelopmentprospectofEMSmutagenesistechnologyinwheatwasprospected.Thisisofgreatsignificanceforenrichingwheatgeneticresourcesꎬacceleratingbreedingprocessesandstudyinggenefunction.Keywords:wheatꎻethylmethylsulfonateꎻchemicalmutagenesisꎻmutant第28卷第5期2019年10月激㊀光㊀生㊀物㊀学㊀报ACTA㊀LASER㊀BIOLOGY㊀SINICAVol.28No.5Oct.2019收稿日期:2019 ̄04 ̄24ꎻ修回日期:2019 ̄05 ̄28ꎮ基金项目:山西省农业科学院农业科技创新研究课题项目(YCX2018412)ꎻ山西省重点研发计划项目(201703D211007 ̄4)ꎮ∗通讯作者:曹亚萍ꎬ研究员ꎬ主要从事小麦种质创新与遗传育种研究ꎮE ̄mail:cyping180@163.comꎮ㊀㊀小麦在长期演变过程中ꎬ由于人工选择和自然进化ꎬ导致遗传基础日趋狭窄ꎬ遗传脆弱性逐渐增加ꎮ绿色革命期间ꎬ半矮秆表型的选择减少了小麦遗传多样性[1]ꎻ近年来ꎬ随着小麦集约化生产和商品性经营ꎬ小麦育种以市场需求为导向ꎬ比较集中地利用少数遗传资源ꎬ许多抗逆㊁抗病虫㊁优质等优异基因逐渐丢失ꎬ育成品种的遗传基础更加狭窄ꎬ对于生物和环境胁迫愈加脆弱ꎬ致使育种进程缓慢ꎬ难以适应农业生产快速发展的需要ꎬ成为小麦遗传改良的瓶颈ꎮ丰富的遗传性状和基因资源是达到品种选育目标的重要基础ꎬ由于小麦属内遗传基因有限ꎬ而常规杂交育种所依据的主要遗传学原理是基因自由组合ꎬ只能利用已有基因进行重组ꎬ不能产生新的基因ꎬ难以解决小麦基因资源狭窄问题ꎮ刘志勇等[2]分析小麦育种现状ꎬ提出未来需大力加强种质资源的原始创新ꎮ长期实践证明ꎬ改变这种现状最基本㊁快速㊁有效的途径之一是诱变育种方法ꎬ诱发突变技术是创造作物新种质㊁丰富遗传多样性和培育优良新品种的一种重要技术手段ꎮ小麦诱变技术是人为利用物理诱变因素(如紫外线㊁X射线㊁γ射线㊁β射线㊁快中子㊁激光㊁离子束等)和化学诱变剂(如烷化剂㊁叠氮化物㊁碱基类似物㊁亚硝基化合物㊁抗生素等)诱发小麦基因组产生变异ꎬ从而创制出自然界原来没有的或一般常规方法难以获得的新类型㊁新性状㊁新基因ꎮ物理诱变因高能射线引起ꎬ染色体畸变率高㊁结构变异广泛ꎬ染色体组紊乱ꎬ后代不育率高ꎬ分离类型广ꎬ纯合世代长ꎻ化学诱变是化学药剂与遗传物质发生生化反应ꎬ结果多是基因的点突变ꎬ纯合世代较短ꎮ甲基磺酸乙酯(ethylmethanesulfonateꎬEMS)是一种高效稳定的烷化类化学诱变剂ꎬ能诱发产生高密度系列等位基因点突变ꎬ获得丰富的遗传材料ꎬ解决小麦育种中种质资源匮乏的问题ꎬ也为相关基因的精细定位㊁克隆及功能分析等提供了研究平台ꎬ在作物诱变技术中应用最广泛㊁效果最好ꎮ1㊀EMS诱变原理及特点EMS属于烷化剂ꎬ线性分子式为CH3SO2OC2H5ꎬ分子量为124.16ꎬ能与醇混溶ꎬ微溶于水ꎮEMS诱发的突变主要通过两个步骤来完成ꎬ首先鸟嘌呤(G)的N ̄7位置被烷基化ꎬ成为一个带正电荷的季铵基团ꎬ从而发生两种遗传效应:一是转换型突变ꎬ烷化的鸟嘌呤(G)不再与胞嘧啶(C)配对ꎬ从而造成GʉC碱基对变成T=A碱基对ꎻ二是颠换型突变ꎬ鸟嘌呤的N ̄7位置烷基化后ꎬ糖苷键断裂ꎬ造成脱嘌呤ꎬ该位置缺失ꎬ在随后的DNA分子复制过程中ꎬ4种碱基都有可能进入到其互补位置ꎬ发生置换现象ꎮ如碱基置换发生于编码多肽区域ꎬ则因可影响密码子而使转录㊁翻译遗传信息发生变化ꎬ以一种氨基酸取代原有的另一种氨基酸ꎻ也可能出现终止密码使多肽链合成中断ꎬ不能形成原有蛋白质而完全失去某种生物学活性ꎮ此外ꎬ诱变剂也可与核苷结构的磷酸反应ꎬ形成酯类而将核苷酸从磷酸与糖分子之间切断ꎬ产生染色体缺失ꎮSidhu等[3]采用生物信息学分析方法ꎬ在小麦品种IndianEMS诱变群体中ꎬ共检测到14130个点突变ꎬ突变频率为每5kb一个ꎬ其中70%转换㊁30%颠换ꎬ并发现存在于染色体远端区域的基因与近端区域中存在的基因相比更容易发生突变ꎮ这些DNA结构上的变化一方面可能改变遗传信息ꎬ引起基因功能丧失ꎬ如抗锈病基因成为敏感型基因ꎻ另一方面可能促使不表达的基因或区段被激活ꎬ而表现出被掩盖的性状ꎮEMS作为化学诱变剂能够引起单一碱基对改变而形成点突变ꎬ染色体畸变相对较少ꎬ不需要进行遗传转化ꎬ可以在短时间内获得大量功能基因的点突变ꎬ引起不同基因的等位变异ꎮ与其它诱变剂相比ꎬEMS诱变后产生的突变频率高ꎬ且多为显性突变体ꎬ易于突变体的筛选ꎻ与常规杂交育种相比ꎬEMS诱变具有随机性ꎬ诱变后可获得丰富的种质遗传材料ꎬ具有种质创新频率高㊁遗传变异谱宽㊁基因纯合周期短等特性ꎬ可解决小麦遗传基础狭窄问题ꎬ有效弥补小麦常规育种方法短时间难以获得新性状和新基因的不足ꎮ2㊀小麦EMS诱变研究现状近年来ꎬEMS诱变技术在国内外得到大规模研究与应用ꎬ在水稻[4ꎬ5]㊁玉米[6]㊁大豆[7ꎬ8]㊁高梁[9]㊁烟草[10]㊁苜蓿[11]㊁蓖麻[12]㊁谷类[13ꎬ14]㊁油料作物[15 ̄17]㊁果蔬[18 ̄23]㊁木本植物[24 ̄26]㊁小麦近缘物种[27ꎬ28]等植物上均取得显著成就ꎮ由于普通小麦是异源六倍体ꎬ基因组庞大ꎬ高达17Gb[29]ꎬ应用EMS进行诱变育种的研究远远落后于模式植物拟南芥[30 ̄32]和水稻[33ꎬ34]等ꎬ但也在诸多方面取得一定进展ꎮ2.1㊀在抗病基因研究方面利用EMS诱变获得抗病基因突变体在研究抗病593第5期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀曹亚萍等:EMS诱变技术在小麦上的应用㊀㊀㊀基因结构㊁功能等方面具有独特优势ꎬ寻找抗病基因区段序列发生变异的突变体是阐释基因功能最直接最有效的方法ꎮ锈病是小麦生产上危害较重也是研究较多的病害之一ꎬ张维宏等[35]用EMS处理小麦抗叶锈病近等基因系TcLr19的种子ꎬ于诱变3代(M3)获得6个感病突变材料ꎬ遗传稳定率达70%以上ꎻHussain等[36]用EMS诱导中抗叶锈小麦品种NN ̄Gandum ̄1ꎬ得到9个高抗和2个高感突变体ꎬSNP分析将抗性突变体变异位点定位于1B染色体短臂(1BS)ꎬ该位点的谷氨酸被丙氨酸取代ꎬ导致蛋白质结构改变ꎻMago等[37]报道ꎬ利用EMS诱导获得抗锈病基因敏感型突变体ꎬ已有几个抗锈病基因从小麦中克隆ꎮ此外ꎬ李韬等[38]用EMS诱变抗赤霉病小麦地方品种黄方柱和海盐种ꎬ得到病小穗率均显著高于相应野生型的6个突变系ꎬ是研究赤霉病扩展抗性的理想材料ꎮ陈洋等[39]用EMS处理抗黄矮病小麦-中间偃麦草易位系YW642的种子ꎬ筛选出18个黄矮病抗性丧失程度不等的突变体ꎬ分子标记检测结果表明这些突变体分别在1~4个分子标记位点上发生变异ꎬ说明这些突变体中抗黄矮病基因Bdv2及其附近区域有不同碱基位点发生突变ꎬ为小麦抗黄矮病基因克隆和功能基因组学研究奠定了坚实的材料基础ꎮ耿皆飞[40]以EMS诱变花培品系H261ꎬ获得小麦类病斑突变体LF2010ꎬ并将其突变基因lm3定位到小麦6BL染色体上6B03和6B40之间2.36M物理距离之中ꎬ同时找到58个候选基因ꎮ2.2㊀在品质性状改良方面随着人民生活水平的提高ꎬ小麦多样化食材成为适应市场经济的必然ꎬ因而对小麦籽粒最终用途要求也不尽相同ꎬ小麦品质改良成为科研工作者关注的焦点ꎮ研究较多的是糯性小麦ꎬSlade等[41]创建了普通小麦和硬粒小麦突变体库ꎬ将EMS化学诱变技术与定向诱导基因组局部突变技术(targetingin ̄ducedlocallesionsINgenomesꎬTILLING)相结合ꎬ筛选到246个等位变异位点ꎬ获得丰富的遗传信息和糯性小麦突变体ꎬ并育成糯性较好的小麦新品种ꎻ李晓等[42]用EMS诱变京411ꎬ以Wx ̄A1为候选基因ꎬ用TILLING技术检测所创建的突变群体ꎬ获得Wx ̄A1基因的7个点突变ꎬ突变密度为1/67kbꎬ其中有功能变异的4个错义突变系均可稳定遗传至下一代ꎬ其直链淀粉含量降低2.8%~7.4%ꎮ张纪元等[43]利用EMS诱变创制软质小麦宁麦9号高分子量谷蛋白亚基突变体ꎬ获得Ax1㊁Dx2㊁Bx7㊁By8㊁Dy12㊁Ax1+By8缺失突变系ꎬ其谷蛋白大聚体和谷蛋白/醇溶蛋白比值均有不同程度降低ꎬ为小麦品质研究奠定了良好的材料基础ꎮ淀粉占小麦籽粒胚乳的70%左右ꎬ是决定小麦磨粉㊁加工品质的重要因素ꎬ高直链淀粉被认为是抗性淀粉(resistantstarchꎬ简称RS)ꎬ又称抗酶解淀粉和难消化淀粉ꎬ其性质类似溶解性纤维ꎬ对于维持肠道健康具有良好作用ꎬ同时具有一定的瘦身效果和保健意义ꎮ薛芳等[44]用EMS处理新春11小麦种子ꎬ筛选出7个抗性淀粉含量高且综合性状优良的M2突变家系ꎮ张贞彩等[45]用EMS处理济麦20和济麦22ꎬ分别得到糊化粘度变异程度不同的突变体ꎬ可形成不同品质㊁不同功能的淀粉材料ꎮMishra等[46]鉴定了一组包含101个EMS诱导的突变系(M4)群体ꎬ分别在约89%和38%的突变体系中观察到直链淀粉和抗性淀粉含量明显区别于野生型ꎬ群体中直链淀粉含量变化范围为3%~76%ꎬ抗性淀粉含量的变化范围为1%~41%ꎻ并用两种不同的直链淀粉含量突变体系研究了20种淀粉代谢途径基因的定量表达模式ꎬ鉴定出直链淀粉生物合成候选基因ꎮ2.3㊀在叶片持绿机制研究方面EMS诱变技术为小麦植株及叶片功能期研究提供了便利ꎮDerkx等[47]采用EMS诱变方法得到小麦扬花期相同而后期冠层快速和慢速衰老突变体ꎬ通过研究突变体对产量和氮分配的影响ꎬ发现延迟衰老仅在较高氮供应时才显现ꎬ低氮供应增加了所有品系的衰老速率ꎬ并用田间试验证实了两种衰老模式ꎮ此外ꎬ通过对小麦叶片早熟突变体m68研究发现ꎬ叶片衰老表型受单个隐性核基因控制ꎬ转录因子和蛋白质转运基因在叶片衰老开始时起作用ꎬ尤其是WRKY家族和锌指转录因子ꎬ叶绿素和碳代谢相关的基因在后期发挥作用[48]ꎮ2.4㊀在农艺性状解析方面株高和分蘖是影响小麦产量的两个主要农艺性状ꎬXu等[49]用EMS处理普通小麦望水白ꎬ获得一个高分蘖矮秆突变体NAUH167ꎬ随后用NAUH167/Su ̄mai3的RIL2:6群体构建了基于分子标记的遗传图谱ꎬ并将控制两种性状的主效QTL(QHt.nau ̄2D)定位于2DSꎬ其侧翼标记为Xcfd11和Xgpw361ꎬ最后用2011I ̄78/NAUH167群体对QHt.nau ̄2D进行了物理定位ꎮ赵天祥等[50]采用EMS突变技术构建了小麦693㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀激㊀光㊀生㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第28卷品种偃展4110的突变体库ꎬ并且对其进行形态学分析和鉴定ꎬ得到株高在10~15cm左右的特矮变异类型ꎮW98是用EMS处理小麦品种偃展1号获得的突变体ꎬ其矮秆与圆粒性状呈显著相关ꎬ用W98与高秆长粒的墨西哥品种10th12配制杂交组合ꎬ对F2ʒ3分离群体进行遗传分析发现ꎬ圆粒性状由1对不完全显性基因控制ꎻ激素敏感性试验表明ꎬ该突变体与野生型都对赤霉素处理不敏感ꎬ但野生型对油菜素内酯不敏感而突变体W98则表现敏感[51]ꎮMo等[52]利用外显子捕捉技术ꎬ结合一个EMS分离群体ꎬ对小麦4BS染色体上的一个矮秆基因区域进行了鉴定ꎬ发现在高秆突变体后代中大约存在一个1.9Mb的缺失ꎬ该缺失区间包含9个基因ꎬ其中之一为Rht ̄B1基因ꎮ为了研究分蘖的潜在遗传变异ꎬKuraparthy等[53]用EMS诱变二倍体小麦ꎬ得到分蘖能力受到损害的突变体ꎬ将分蘖抑制基因tin3定位在染色体3AL远端10%位置ꎬ并开发出一个与tin3共分离的RFLP标记Xpsr1205ꎬ促进了小麦有效分蘖的改良ꎮ抽穗期作为普通小麦重要农艺性状之一ꎬ对适应不同生态环境条件具有至关重要的作用ꎬ通过调节小麦抽穗期ꎬ使其与光㊁温等环境因子变化密切协调ꎬ从而提高小麦适应性和稳产性ꎮ刘国祥[54]对偃展4110EMS突变体库60份抽穗期突变体研究发现ꎬ光周期对突变体抽穗期的影响极为显著ꎬ通过对光周期基因Ppd ̄Dl的克隆测序与比对ꎬ发现突变体有单碱基突变㊁C缺失㊁C/T转换㊁G插入和T插入多种类型ꎬ这些点突变造成启动子区碱基转换㊁氨基酸改变以及内含子调控序列变化ꎬ从而导致抽穗期发生变异ꎮZhang等[55]用EMS处理YZ4110ꎬ获得晚抽穗期突变体m605ꎬ这种晚期抽穗性状由一个名为TaHdm605的隐性基因控制ꎬ采用遗传作图方法将TaHdm605基因定位在3DL分子标记cfd152和barc42之间ꎬ而后进一步将该基因座定位到包含26个预测基因的1.86Mb物理基因组区域ꎬ为TaHdm605克隆以及改变小麦抽穗期奠定了遗传基础ꎬ并且该突变体可在秋季播种时至少延迟7天ꎮWu等[56]从EMS处理普通小麦品种望水白的突变体文库中获得突变体Meh0239ꎬ通过对其进行形态学㊁生理学㊁解剖学和遗传学的研究ꎬ鉴定出一个与产量性状相关的多效性基因Yt1ꎬ该突变体为小麦染色体7DS上Xwmc506远端约3.1cM处单个隐性突变ꎮ2.5㊀在突变体库构建方面据Krasileva等[57]报道ꎬ用EMS处理四倍体小麦Kronos和六倍体小麦Cadenz种子ꎬ提取M2植株DNA进行外显子捕捉测序ꎬ在1535份Kronos和1200份Cadenza的EMS群体中ꎬ在基因水平上鉴定出超过一千万个突变位点ꎬ平均每个单株存在2705(四倍体)和5351(六倍体)个位点ꎬ突变频率大约在35~40个SNP/kbꎮ对于单个基因来说ꎬ大约有23~24个突变位点造成了错义或提前终止ꎮ由于单个突变单株在整个基因组水平上均含有大量突变位点ꎬ将突变体与野生型材料进行杂交并连续回交ꎬ可以纯化遗传背景以消除其它突变位点对目标性状造成的影响ꎮ该突变体库包含四倍体小麦Kronos和六倍体小麦Cadenzꎬ可以用作改善小麦营养品质㊁籽粒大小㊁鉴定等位基因等方面的研究ꎬ也是小麦功能基因组学研究的宝贵遗传资源ꎬ同时也为解析在人工或自然选择中被忽略掉的隐性突变提供帮助ꎮ在鉴定突变位点的基础上ꎬ作者又对突变位点进行了注释ꎬ相关突变信息可以通过www.dubcovskylab.uc ̄davis.edu/wheat ̄tilling网站进行查询ꎬ山东农业大学付道林组有四倍体Kronos的突变体库ꎬ国内感兴趣的研究人员可以向他们申请(信息来源于小麦研究联盟)ꎮ3㊀小麦EMS诱变突变体选择3.1㊀小麦EMS诱变及突变体表型选择诱变材料需选用待处理品种(系)的纯系ꎮ育种研究需要根据育种目标选用具有较好综合性状㊁只需进行少数性状改进的当地推广品种或高代品系ꎻ遗传研究需要选择目标性状突出的亲本材料ꎮ小麦EMS诱变处理宜采用种子处理方式ꎬ具体操作如下:首先ꎬ将待处理种子在室温下用蒸馏水浸种10h左右ꎬ使种子充分膨胀或萌动ꎬ随后放在吸水纸上晾干ꎬ将种子含水量控制在20%以下ꎻ其次ꎬ以磷酸缓冲液(pH=7)为溶剂ꎬ配制0.5%~0.8%的EMS化学诱变剂ꎬ将晾干的种子在室温下浸种8~10hꎻ最后ꎬ将诱变后的种子装入小网袋中ꎬ在自来水下反复冲洗1hꎬ除去种子胚上残留的EMSꎬ风干备用ꎬ5天内播种ꎮ值得注意的是ꎬ不同基因型材料对EMS敏感程度不同ꎬ不同处理时间㊁不同EMS浓度及不同浓度与时间组合对突变频率影响也具有较大差异ꎮ首次应用时需谨慎对待ꎬ最好设不同剂量EMS浓度来处理种子ꎬ以达到理想效果ꎻ另外ꎬEMS具有强烈的致癌性和挥发性ꎬ常用5%硫代硫酸钠作为解毒剂ꎬ因此在操作过程中要注意安全防护ꎬ严格遵守试验793第5期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀曹亚萍等:EMS诱变技术在小麦上的应用㊀㊀㊀规则ꎮ诱变M1表型不分离ꎬ一般按单株或单穗(每株1穗)收获ꎻM2是变异最大的世代ꎬ也是选择的关键时期ꎬ可根据育种目标及性状遗传特点选择各种表型突变体(图1)ꎬ如株高㊁株型㊁穗型㊁叶色㊁分蘖㊁生育期㊁育性等性状ꎻM3代及以后ꎬ随着世代的增加ꎬ性状分离减少ꎬ有些性状一经获得即可迅速稳定ꎮ图2和图3为笔者采用EMS诱变得到的表型稳定突变体ꎬ目前已用于突变基因遗传和功能研究ꎮ图1㊀EMS诱变M2代表型突变体Fig.1㊀PhenotypicmutantsofEMSinductioninM2generation(a)良星99色泽突变ꎻ(b)济麦22分蘖力突变ꎻ(c)晋麦47号生育期突变ꎻ(d)冀麦325株型突变(a)MutationofcolorofLiangxing99ꎻ(b)MutationoftilleringabilityofJimai22ꎻ(c)MutationofgrowthstageofJinmai47ꎻ(d)MutationofplanttypeofJimai325图2㊀晋麦47号(a)及其抗白粉病突变体(b)Fig.2㊀Jinmai47(a)anditspowderymildewresistantmutant(b)3.2㊀利用TILLING技术定向筛选突变体尽管EMS在表型选择方面依旧被广泛利用ꎬ但存在EMS诱发产生的点突变难以鉴定的问题ꎮTILLING是由美国FredHutchinson癌症研究中心StevenHenikoff领导的研究小组发展建立的一种反向遗传学研究方法ꎬ它将诱发产生高频率点突变的化学诱变方法与PCR筛选技术和高通量检测方法有效结合ꎬ以发现分析目标区域点突变ꎬ是一种全新的高通量㊁低成本的反向遗传学研究方法ꎮTILLING作为一种定向点突变筛选技术ꎬ对目标突变体的筛选不受遗传背景㊁基因互作㊁表型特征㊁生长环境等因素影响ꎬ鉴定准确性高ꎬ能够实现高通量㊁大群体㊁多基因㊁多性状的快速高效鉴定ꎬ提高突变体鉴选效率ꎮTILLING提供了从分子水平上定向规模化筛选突变体的技术平台ꎬ尤其对品质和营养成分等无法从植株表型上加以选择的性状筛选尤为有利ꎮ其技术原理是将传统的酶切技术与PCR技术相结合后采用红外双色荧光系统进行结果鉴定ꎬ从而筛选出相应的突变体ꎮ首先ꎬ提取具有性状分离的M2植株的基因组DNAꎬ将多个不同样品的DNA进行等量混合ꎬ构建DNA池ꎻ其次ꎬ以此DNA池为模板进行PCR扩增ꎬ并将扩增片段进行退火形成DNA片段的异源双链分子ꎬ采用CELI进行酶切后ꎬ利用红外双色荧893㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀激㊀光㊀生㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第28卷图3㊀长6878及其黄叶突变体Fig.3㊀Chang6878anditsyellowleafmutant(a)苗期ꎻ(b)抽穗期ꎻ(c)灌浆期ꎻ(d)子粒(a)Germinationstageꎻ(b)Headingstageꎻ(c)Fillingstageꎻ(d)Grain注:每张图片左或上为野生型(长6878)ꎬ右或下为突变体Note:Theleftorupperpartofeachpictureiswildtype(Chang6878)ꎬwhiletherightorlowerpartismutant光电泳分析技术进行电泳ꎻ再次ꎬ对检测到突变的DNA池中每个单株的DNA样品进行筛选ꎬ找出相应的突变体ꎻ最后ꎬ从对应的突变体植株中筛选出有变化的表型ꎮ详细操作方法见参考文献[58]和[59]ꎮ值得注意的是ꎬ按目标基因序列设计引物ꎬ是TILL ̄ING技术的一个重要步骤ꎬ设计的好坏直接影响TILLING的筛选效果ꎮ3.3㊀突变体真实性检测虽然普通小麦为自花授粉植物ꎬ但也存在1%~4%的天然异交率ꎬ如果诱变后代不能严格套袋自交ꎬ必须确认突变体的变异来源ꎬ才能对诱变后代进行遗传变异评价和基因功能研究ꎮ利用表型鉴定通常难以鉴定诱变后代突变体的真实性ꎬ而利用分子标记分析可以有效排除诱发突变体中的假突变体ꎮ耿皆飞等[60]在小麦EMS突变体真实性检测方面进行了首次报道:LF2010㊁LF2099和LF2100是小麦品系H261经EMS诱变后遗传稳定的突变体ꎬ用分别位于小麦21条染色体上特异性和稳定性均好的21对SSR引物对突变体及其亲本进行检测ꎬH261与LF2010和LF2099的差异SSR标记为0个ꎬ但与LF2100的差异SSR标记为10个ꎻSNP芯片分析结果表明ꎬH261与LF2010和LF2099之间的差异位点分别为66和12个ꎬ与LF2100之间的差异位点为2846个ꎮ证明LF2010和LF2099突变体与亲本H261的遗传背景高度一致ꎬ是H261经过EMS诱变的后代ꎬ而LF2100是天然异交或机械混杂产生的假突变体ꎮSSR标记和SNP芯片2种方法均可有效鉴定EMS突变体的真实性ꎬ由于SNP芯片可以进行高通量和全基因组水平分析ꎬ在小麦突变体真实性鉴定方面具有更大应用潜力ꎮ4㊀EMS诱变存在问题及解决方案EMS诱变原理是进行DNA碱基配对的干扰ꎬ使其发生碱基置换从而形成点突变ꎮ这种方法以诱发基因突变为目的ꎬ实质上主要依赖于物种在诱变条件下所发生的基因随机突变ꎬ其突变机理是在诱变条件下DNA复制过程中所产生的一个或几个碱基的变化ꎬ这一过程是随机突变的过程ꎬ具有突变位点不确定性㊁突变方向偶然性的特点ꎮ由于碱基变化是随机的ꎬ整个变异过程无目的性ꎬ即使是用EMS处理相同品种并且重复同样诱变条件ꎬ也无法预知必然可以出现某一特殊性状尤其是目标性状改良的突变ꎮ由表1可知ꎬ不同材料EMS诱变结果具有较大差异ꎬ同一部位突变率差异较大且具随机性ꎬ多项报道也进一步证实了这一点ꎮDhaliwal等[1]报道了一个用EMS处理春小麦品种Indian生成的突变体群体ꎬ在M4代观察到的表型稳定群体中ꎬ植物高度的变化最常见ꎬ其次是叶形态ꎮ许云峰等[61]用EMS对小麦品种烟农15进行诱变处理ꎬ在M3代得到11个农艺性状发生明显变异的突变系ꎬ以籽粒大小和株高2个性状的变异幅度最大ꎬ并且均有复合性状突变出现ꎮGuo等[62]采用0.5%㊁1.0%和1.5%三种EMS浓度处理京411种子ꎬ表型鉴定结果表明ꎬ除生育期外ꎬ其余性状的突变频率均随着EMS浓度的增加而增加ꎮ993第5期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀曹亚萍等:EMS诱变技术在小麦上的应用㊀㊀㊀表1㊀小麦EMS诱变群体中不同性状类型突变率(%)Tab.1㊀MutationratesofdifferenttraittypesinwheatEMSinducedpopulation(%)WildtypeEMSConcentration+TimeMutationtypeSpikeLeafStemFertilityMaturityOthersTotalmutationrateReferencesShengnong10.4%+16h0.710.890.490.220.041.513.86[63]Yunong2010.8%+12h0.602.203.495.1511.44[64]Yanzhan41101.2%0.260.631.063.341.591.228.10[50]Yanzhan41100.7%0.200.930.802.221.120.435.70[50]Xinong991.0%+10h5.122.093.960.082.140.2013.59[65]Xinong9791.0%+8h1.600.644.000.000.160.006.40[65]Xinong9771.0%+12h3.241.393.700.000.690.239.25[65]Xiaoyan221.0%+12h3.601.275.080.001.690.2111.86[65]㊀㊀应用小麦EMS诱变技术ꎬ除构建突变体库外ꎬ还需要针对性地创制新基因和新材料ꎬ尤其是应用于育种研究时需要对少数不利性状进行改良ꎬ实现定向诱变ꎮ为了解决EMS诱变的随机性和研究目标的定向性这一矛盾ꎬ需要在诱变群体中有效添加选择压ꎬ以提高目标突变体选择准确性和利用效率ꎬ进而实现对小麦某一特殊性状进行遗传改良ꎮ如对诱变群体M2进行干旱㊁低温㊁高温㊁盐碱等环境胁迫ꎬ可以鉴定出各种抗逆性强的突变体ꎻ对M2进行病㊁虫㊁菌接种鉴定或土壤带菌操作等试验ꎬ可鉴定出抗各种病害突变体ꎻ对M2籽粒进行蛋白电泳分析ꎬ可检测品质性状相关基因突变体ꎻ提取M2植株基因组DNAꎬ按目标基因序列设计引物ꎬ用TILLING技术定向筛选ꎬ可鉴定更多相关基因突变ꎮ图4给出了小麦EMS诱变研究备选方案ꎬ旨在为科研工作者提供一种研究方法和策略ꎬ以便根据研究目标选择性借鉴ꎮ5㊀小麦EMS诱变技术发展前景5.1㊀培育小麦新品种近年来ꎬ我国小麦生产上大面积种植品种的产量㊁品质㊁抗性大多取得了明显改进ꎬ但由于生态环境等因素影响ꎬ对于各地育种家来说ꎬ如能改进某个品种的某1~2个性状ꎬ即可产生良好的社会和经济效益ꎮ小麦EMS诱变育种的主要特点是对少数不利性状进行改良ꎬ在小麦育种实践中发挥不可替代的作用ꎮ基于TILLING的诱变技术ꎬ将是一种高效定向性育种技术ꎬ它不但继承了诱变育种稳定快㊁只改变原亲本单个目标性状的传统优点ꎬ而且无须耗时的转基因和连续的杂交㊁回交过程ꎮ由TILLING分析所鉴定的大量目标突变体分别含有不同的或新的等位变异ꎬ通过与常规育种技术有效结合ꎬ能够实现目标性状优良等位变异的基因聚合ꎬ从而创制综合性状优良的育种新材料ꎬ促进小麦耐逆㊁高产㊁优质新品种的培育ꎮEMS诱变创造出有利性状的变异ꎬ可以作为优良育种亲本或自交纯合作为品种推广应用ꎬ也可从育种材料转为基因组学研究的重要基础材料ꎮ5.2㊀诱生小麦新基因在当前小麦种质资源库新基因极度缺乏㊁遗传资源日益枯竭的状况下ꎬEMS高效诱导点突变和不易造成染色体畸变的优势ꎬ被广泛应用于小麦研究中ꎬ能够在短时间获得新性状和新基因ꎬ极大程度丰富了小麦种质资源ꎮ这些资源源于人工诱变ꎬ控制这些性状的基因或等位基因与来自自然变异的基因或等位基因通常是不相同的ꎬ是一种新的基因资源ꎮ这些表型性状变异是由人工诱变获得的新型等位基因变异ꎬ可以与传统品种进行杂交ꎬ增加小麦育种的创新性ꎬ丰富自然界种质资源ꎻ部分优异资源将成为分子设计育种的理想材料ꎮ5.3㊀图位克隆基因选择具特定变异的稳定突变株ꎬ与其野生型或同种性状中表型差别较大的品种配制杂交组合ꎬ培育大分离群体ꎬ可以对控制该性状的基因进行精细定位㊁图位克隆ꎬ并为了解变异产生的分子遗传机制提供基础材料ꎮEMS诱变丰富了图位克隆的数量与资源ꎬ随着分子生物学研究的深入和技术的更新ꎬ这种方法思路在小麦功能基因的发掘与利用方面会越来越深入ꎮ004㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀激㊀光㊀生㊀物㊀学㊀报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第28卷。
公示-石河子大学研究生招生信息
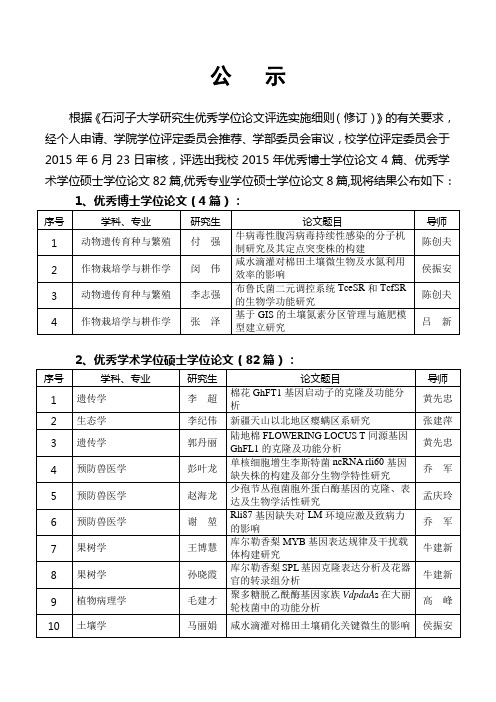
公示根据《石河子大学研究生优秀学位论文评选实施细则(修订)》的有关要求,经个人申请、学院学位评定委员会推荐、学部委员会审议,校学位评定委员会于2015年6月23日审核,评选出我校2015年优秀博士学位论文4篇、优秀学术学位硕士学位论文82篇,优秀专业学位硕士学位论文8篇,现将结果公布如下:1、优秀博士学位论文(4篇):序号学科、专业研究生论文题目导师1 动物遗传育种与繁殖付强牛病毒性腹泻病毒持续性感染的分子机制研究及其定点突变株的构建陈创夫2 作物栽培学与耕作学闵伟咸水滴灌对棉田土壤微生物及水氮利用效率的影响侯振安3 动物遗传育种与繁殖李志强布鲁氏菌二元调控系统TceSR和TcfSR的生物学功能研究陈创夫4 作物栽培学与耕作学张泽基于GIS的土壤氮素分区管理与施肥模型建立研究吕新2、优秀学术学位硕士学位论文(82篇):序号学科、专业研究生论文题目导师1 遗传学李超棉花GhFT1基因启动子的克隆及功能分析黄先忠2 生态学李纪伟新疆天山以北地区瘿螨区系研究张建萍3 遗传学郭丹丽陆地棉FLOWERING LOCUS T同源基因GhFL1的克隆及功能分析黄先忠4 预防兽医学彭叶龙单核细胞增生李斯特菌ncRNA rli60基因缺失株的构建及部分生物学特性研究乔军5 预防兽医学赵海龙少孢节丛孢菌胞外蛋白酶基因的克隆、表达及生物学活性研究孟庆玲6 预防兽医学谢堃Rli87基因缺失对LM环境应激及致病力的影响乔军7 果树学王博慧库尔勒香梨MYB基因表达规律及干扰载体构建研究牛建新8 果树学孙晓霞库尔勒香梨SPL基因克隆表达分析及花器官的转录组分析牛建新9 植物病理学毛建才聚多糖脱乙酰酶基因家族VdpdaA s在大丽轮枝菌中的功能分析高峰10 土壤学马丽娟咸水滴灌对棉田土壤硝化关键微生的影响侯振安11 作物遗传育种韩泽刚枯萎病菌诱导棉花幼苗根部基因表达谱的分析及GhERFB101基因的克隆张薇12 生态学王静娅干旱区典型盐生植被群落土壤团聚体及其生物学特性的分析张凤华13 植物营养学陶瑞化肥减量有机替代下滴灌农田氮素转化与N2O排放褚贵新14 作物栽培学与耕作学刘军长期连作及秸秆还田棉田土壤有机碳组分变化研究刘建国15 生物化学与分子生物学刘逸泠杂交狼尾草再生体系的建立及转sikPIP基因的研究祝建波16 生物化学与分子生物学史梦婷miR-193a影响BVDV在MDBK细胞内致病机制的研究陈创夫17 作物遗传育种王昊龙小麦淀粉分支酶SBEIIa、SBEIIb基因序列多态性分析及功能标记的开发李卫华18 应用化学盛文波基于阿维菌素控制释放功能性聚合物的制备及其性能研究贾鑫19 农业水土工程李文昊新疆长期膜下滴灌棉田土壤盐分离子分布特征与时空迁移规律研究王振华20 农业电气化与自动化丁竹青基于LabVIEW Vision的加工番茄外部缺陷在线检测系统设计与研究坎杂21 应用化学郭豪氮杂环卡宾催化C-X键形成研究何林22 工业催化黄超锋基于“界面效应”设计高稳定性乙炔氢氯化反应Au基催化剂朱明远23 水利水电工程刘毅伊犁黄土滑坡特征、成因及稳定性分析程建军24 农产品加工及贮藏工程高聪一株产几丁质酶细菌的筛选鉴定、酶学性质及降解几丁质的研究童军茂25 工业催化陈昆乙炔氢氯化反应碳载体孔径调控及改性研究代斌26 机械设计及理论冉兵加工番茄酱后皮渣双作用分离试验研究坎杂27 应用化学赵赫基于咔唑的有机小分子光电材料的合成及其性能研究刘志勇28 生物化工乔醴峰基于核酸适配体的β-葡萄糖醛酸苷酶一步纯化固定化研究及其应用李春29 工业催化王洋甲醇定向转化制苯乙烯反应机理的理论研究康丽华30 工业催化周昌健大黄酸-氨基酸偶联物的合成及抗肿瘤活性初步研究代斌31 农业机械化工程冯哲苹果不同受载时接触应力测量分析及损伤预测吴杰32 生物化工汤恒基于β-葡萄糖醛酸苷酶结构解析的耐热性研究李春33 应用化学张晶2-三氟乙酰基苯酚与活泼碳碳叁键串联反应研究代斌34 机械制造及其自动化徐虎博不同内部品质库尔勒香梨振动频谱特性分析研究吴杰35 机械电子工程高国刚基于DSP数字图像处理的哈密瓜品质检测研究马本学36 机械设计及理论王梦籽棉回潮率自动测量装备的研究坎杂37 区域经济学王淑娟向西开放背景下新疆优势产业选择及发展研究李豫新38 产业经济学张震新疆生产性服务业对经济增长的影响研究程广斌39 农业经济管理王洁菲基于产业安全的新疆棉花产销体系优化研究王力40 企业管理陈艳小微企业创业领导力开发研究————以石河子市科技型小微企业为例刘追41 企业管理徐兵变革型领导、心理授权与知识型员工离职倾向关系研究石冠峰42 金融学姜勇新疆农村金融普惠的空间差异及集聚效应研究徐敏43 产业经济学帅林遥中国新疆边境贸易便利化影响因素研究李豫新44 马克思主义民族理论与政策吴良平一个净土宗寺院再造研究:以玛纳斯大佛寺为例龙开义45 企业管理韩宏稳新生代员工组织公平感与离职倾向关系研究石冠峰46 马克思主义中国化研究田海峰唐安西四镇屯城布局研究张安福47 金融学蔡玉婧兵团小微企业的融资困境及对策研究何剑48 区域经济学张盼盼天山北坡经济带城市综合承载力评价研究程广斌49 会计学齐云飞行业成长性与我国上市公司现金持有杨兴全50 区域经济学张静新疆外贸结构转型升级路径研究龚新蜀51 农业经济管理胡世勇合肥市经济技术开发区被征地农民消费行为研究胡宜挺52 金融学蒋世辉新疆地方农户与兵团职工金融需求差异研究马乃毅53 区域经济学李崇基于包容性增长的兵团居民幸福感提升研究朱金鹤54 马克思主义中国化研究豆孝蕊兵团组织文化研究闫卫华55 药物分析学刘媛几种纳米复合材料的制备及其在电化学传感器中的应用研究唐辉56 肿瘤学李一鑫胃癌中VEGF、VEGFR2表达的临床病理学意义及其对预后的影响张文杰57 病理学与病理生理学李文琴Notch1信号通路相关基因mRNA表达与乳腺癌变、进展相关性研究曹玉文58 病理学与病理生理学谢玉汶GEFT基因对横纹肌肉瘤细胞生物学行为的影响李锋59 外科学王圣超MHCI-VEGFR2抗原肽单链三聚体DNA疫苗的抗肿瘤转移作用及其机制牛建华60 药剂学侯雪娇玫瑰花总黄酮的提取纯化及其抗离体心肌缺血再灌注损伤研究陈文61 流行病与卫生统计学庞鸿瑞新疆维吾尔族、哈萨克族农村居民高同型半胱氨酸血症流行特点及与相关疾病的关系郭淑霞62 护理学陈玲创伤后应激障碍症状学特征与眼动脱敏再加工疗效关系探讨张桂青63 肿瘤学谷倬宇COX-2、MMP-14和胰腺癌细胞增殖、侵袭、转移的关系研究李军64 肿瘤学李焕焕塞来昔布联合吉非替尼在非小细胞肺癌中的实验研究巩平65 外科学卞正君KLD-12多肽/TGF-β1诱导MSCs向髓核样细胞分化的实验研究孙建华66 病理学与病理生理学李肃PLCE1在食管鳞癌组织中的表达及其对肿瘤细胞增殖和侵袭的影响研究陈云昭67 生物化学与分子生物学王旭晖Neuritin通过Notch信号通路在血管生成中的作用黄瑾68 生药学戴沩特克斯橐吾根及根茎的化学成分及抗补体活性研究王琪69 病理学与病理生理学唐娜人巨细胞病毒和环境危险因素互作对新疆汉族原发性高血压易感性的影响何芳70 流行病与卫生统计学陈娟PPARγ基因单核苷酸多态性与维吾尔族代谢综合征的相关研究郭淑霞71 流行病与卫生统计学崔俊营基于多水平模型的兵团居民自评健康趋势和影响因素研究唐景霞72 流行病与卫生统计学何菲新疆农村地区维吾尔族和哈萨克族人群糖尿病和空腹血糖受损患病率及其影响因素郭淑霞73 内科学马莉活性维生素D3对人肾小球系膜细胞中Akt级mTOR表达的影响杨晓萍74 病原生物学王婵靶向Mcl-1的shRNA对结核杆菌感染巨噬细胞的凋亡影响章乐75 生理学张文雯新疆哈、汉族人群原发性高血压与缝隙连接基因多态性的相关性研究司军强76 肿瘤学冯文磊血管内皮前体细胞对间充质干细胞增殖能力的影响吴向未77 病理学与病理生理学何丽MTB PPS对MTB耐药性产生的作用及其机制研究张万江78 药理学龚福恺治疗白血病中药的活性分子及靶点挖掘——α-Viniferin 抑制K562 细胞生长的机制研究张波79 社会医学与卫生事业管理盖莉莉新疆农村妇女宫颈癌筛查项目组织现状研究井眀霞80 社会医学与卫生事业管理黄蓉哈密地区卫生人力现状及配置研究秦江梅81 护理学吴复琛新疆维吾尔族、哈萨克族失能老年人居家主要照护者照护负担及影响因素研究王玉环82 内科学王婉秋辛伐他汀对2型糖尿病动脉硬化大鼠血浆VEGF、TGF-β1及CTRP3水平的影响研究孙侃3、优秀专业学位硕士学位论文(8篇):序号学科、专业研究生论文题目导师1 农业硕士养殖领域刘文骁彩鲫与建鲤杂交F1代生物学特性研究陈宏2 兽医硕士胡政香绵羊肺炎支原体新疆塔城流行株的分离鉴定及油佐剂灭活苗免疫原性初步研究孟庆玲3 农业硕士作物领域康亚龙连作对加工番茄土壤微生物群落及多样性的影响蒋桂英4工程硕士化学工程领域吴丰田铜催化C-N/C-O偶联合成N-芳基咪唑和苯并噁唑的催化体系研究谢建伟5工程硕士食品工程领域张继贤基于鹰嘴豆蛋白美拉德肽的制备技术研究与产品开发程卫东6 临床医学硕士肿瘤学王宁滑膜肉瘤上皮间叶不同组织成分中EMT相关基因的表达与意义李锋7 临床医学硕士肿瘤学吴何兴VEGF、CD34在肝泡状棘球蚴浸润性生长中的作用张示杰8临床医学硕士妇产科学纪莎HPVL1基因甲基化在维汉两组宫颈癌中的相关性分析玛依努尔·尼牙孜以上名单从今日起在校内公示7天,公示期内校学位评定委员会按程序受理个人或集体提出的异议;无异议者,即可作为我校2015年优秀博士、硕士学位论文。
植物基因克隆技术研究进展

植物基因克隆技术研究进展植物基因克隆技术是近年来生物科学领域的研究热点之一。
这项技术的目的是通过克隆植物的基因片段,进而研究植物的基因组及其功能,以推动植物育种和农业生产的发展。
本文将综述植物基因克隆技术的研究现状、克隆位点选择、克隆片段制备以及应用等方面的进展。
植物基因克隆技术发展迅速,其研究范围已经涉及到了许多方面。
例如,研究人员利用该技术克隆了抗逆、抗病、高抗虫等具有重要应用价值的基因,并在转基因植物研究中广泛应用。
植物基因克隆技术还被应用于植物进化和系统生物学研究,为揭示植物物种演化和遗传多样性提供了有力支持。
克隆位点选择是植物基因克隆技术的关键步骤之一。
选择合适的克隆位点,可以大大提高基因克隆的效率和成功率。
根据文献报道,植物基因克隆中常用的克隆位点包括:质粒、酵母人工染色体(YAC)、细菌人工染色体(BAC)、P1人工染色体(PAC)和 cosmids等。
这些克隆位点的选择应根据具体的研究目标和植物基因的特点来决定。
例如,对于需要长期稳定遗传的克隆,质粒可能不是最佳选择,因为其在细胞内的拷贝数会随着代数的增加而逐渐减少。
而对于需要大规模克隆和组装复杂基因组的植物,则可以选择使用酵母人工染色体或细菌人工染色体等大容量克隆载体。
克隆片段制备是植物基因克隆技术的另一个关键步骤。
根据文献报道,克隆片段制备的主要方法包括:鸟枪法、定向克隆、适应性扩增和基于连接酶的扩增等。
这些方法的优缺点各不相同。
例如,鸟枪法可以快速制备大量文库,但需要使用大量的起始DNA样品。
定向克隆可以确保克隆片段的方向正确,但需要设计特定的引物和模板。
适应性扩增可以在一定程度上降低成本,但需要使用特殊的引物和反应条件。
基于连接酶的扩增方法可以在一定程度上保证扩增的特异性,但需要使用高纯度的DNA样品和特殊的连接酶。
植物基因克隆技术的应用范围非常广泛。
例如,研究人员利用该技术成功克隆了多个抗逆、抗病和高抗虫的基因,并通过转基因技术将这些基因导入到农作物中,显著提高了农作物的抗性和产量。
小麦胚乳14-3-3蛋白的表达及其淀粉体淀粉合成酶的互作

作物学报ACTA AGRONOMICA SINICA 2009, 35(8): 1445−1450/zwxb/ ISSN 0496-3490; CODEN TSHPA9E-mail: xbzw@DOI: 10.3724/SP.J.1006.2009.01445小麦胚乳14-3-3蛋白的表达及其与淀粉体淀粉合成酶的互作宋健民1戴双1李豪圣1刘爱峰1程敦公1楚秀生1 Ian J TETLOW2 Michael J EMES21山东省农业科学院作物研究所, 山东济南 250100; 2 University of Guelph, Guelph, N1G 2W1, Canada摘要: 从小麦胚乳中克隆了14-3-3基因, 并将其分别插入pET29c和pET41c质粒, 用热激法转化大肠杆菌BL21-CodonPlus (DE3)-RP, 得到高效表达的蛋白, 但融合蛋白主要以包涵体的形式存在。
可溶性的融合蛋白可直接通过S-蛋白琼脂糖树脂纯化。
包涵体经8 mol L−1尿素溶解变性, 稀释复性后, 结合到S-蛋白琼脂糖树脂上, 也得到纯化的融合蛋白。
复性后的融合蛋白对蔗糖合酶活性表现抑制作用, 说明包涵体14-3-3融合蛋白恢复活性。
将结合14-3-3融合蛋白的S-蛋白琼脂糖树脂作为诱饵与小麦胚乳淀粉体提取液进行亲和杂交, 与14-3-3蛋白特异互作的淀粉合成酶结合到S-蛋白琼脂糖树脂上, Western 检测结果表明, 淀粉体淀粉合酶I (SSI)、淀粉合酶II (SSII)、淀粉分支酶Iia (SBEIIa)、淀粉分支酶Iib (SBEIIb)和ADP焦磷酸化酶大亚基 (SH2)与14-3-3蛋白存在互作, 而淀粉分支酶I (SBEI)、淀粉磷酸化酶(SP)、D-酶(DE)和ADP焦磷酸化酶小亚基(BT2)不能与14-3-3蛋白结合, 说明小麦胚乳14-3-3蛋白对淀粉体淀粉合成具有一定的调控作用。
关键词: 小麦14-3-3蛋白; 表达; 淀粉合成酶; 蛋白互作Expression of a Wheat Endosperm 14-3-3 Protein and Its Interactions with Starch Biosynthetic Enzymes in AmyloplastsSONG Jian-Min1, DAI Shuang1, LI Hao-Sheng1, LIU Ai-Feng1, CHENG Dun-Gong1, CHU Xiu-Sheng1, Ian J TETLOW2, and Michael J EMES21 Crop Research Institute, Shandong Academy of Agricultural Sciences, Jinan 250100, China;2 University of Guelph, Guelph, N1G 2W1, CanadaAbstract: Wheat endosperm starch is the major determinant of grain yield and processing quality. The quality and quantity of starch are controlled by a number of starch biosynthetic enzymes. 14-3-3 proteins, involved in many biological processes, are ubiquitous and important regulators in all eukaryotic cells from yeast to mammals and plants. Protein-protein interactions betweena wheat endosperm 14-3-3 protein and starch biosynthetic enzymes from amyloplast were investigated in this study. A 14-3-3 gene was cloned from developing wheat endosperm and inserted into plasmid vectors pET29c and pET41c. The recombinant vectors were transformed into Escherichia coli strain BL21-CodonPlus (DE3)-RP and expressed at very high level. The fusion protein existed mainly as an insoluble inclusion body after extraction by BugBuster Protein Extraction Reagent. The soluble fusion pro-tein was purified by bounding to S-protein agarose, while the inclusion body should be dissolved in 8 mol L−1 urea and refolded firstly. Sucrose synthase activity was observed to be inhibited by exogenous recombinant 14-3-3 protein in a dosage-dependent manner, which suggested the refolded protein was successfully activated and could be used in the following research. The purified recombinant 14-3-3 protein was bound to S-protein agarose as a biochemical bait, and then incubated with wheat amyloplast ex-tract. Proteins interacting specifically with the 14-3-3 protein and remaining on the resin were analyzed by SDS-PAGE and West-ern blot. These assays showed that starch synthase I (SSI), starch synthase II (SSII), starch branching enzyme IIa (SBEIIa), starch branching enzyme IIb (SBEIIb), and ADP glucose pyrophosphorylase large subunit (SH2) interacted with 14-3-3 protein, whereas本研究由国家自然科学基金项目(30700492), 国家高技术研究发展计划(863计划)项目(2006AA100102), 国家科技支撑计划项目(2006BAD01A02), 引进国际先进农业科学技术计划(948计划)项目[2006-G2(B)], 公益性行业科研专项(NYHYZX07-002), “泰山学者”建设工程, 山东省农业良种工程项目资助。
小麦淀粉分支酶基因SBE-Ⅱb植物表达载体的构建及表达鉴定

小 麦淀 粉 分 支酶 基 因 S B E — I I b植 物 表 达 载 体 的构 建 及 表 达 鉴 定
王 自布 , 齐军仓 , 李卫华 , 曹连莆 , 石培春
( 1 . 贵州省生物资源开发利用特色重点实验室 , 贵州 贵 阳 5 5 0 0 1 8 ; 2 . 石河子大学 农学 院, 新疆 石河子 8 3 2 0 0 3 ) 摘要: 从花后 1 5 d的小麦 ( T r i t i c u m a e s t i v u m L . ) 胚乳中提取 R N A, 根据 G e n e B a n k上已经公 布的小麦淀粉分 支 酶基因( S B E . 1 I b ) 序列 ( A Y 7 4 0 4 0 1 . 1 ) 设计引物 , 经 R T - P C R扩增得到一条 2 5 1 1 b p的条带 , 测序结果 与公布的 序列一致。将克 隆得到 的基 因构建到了植物表达载体 p P Z P 3 5 S上 , 通过农杆菌叶盘转化法转化烟草叶片 , 经卡 那霉素筛选 、 P C R扩增鉴定证明 , S B E — I I b基 因已成功转入烟草 细胞 中, 获得 4个转基 因株系 。转基 因烟草叶
Ab s t r a c t : T h e t o t a l R N A w a s i s o l a t e d f r o m w h e a t ( T r i t i c u n a e s t i v u m L . )e n d o s p e r m 1 5 d a y s a f t e r p o l l i .
Co n s t r u c t i o n o f Pl a n t Tr a n s f o r ma t i o n Ve c t o r o f Wh e a t S t a r c h
210976091_《高抗性淀粉小麦籽粒抗性淀粉含量指标和检测方法》标准解读

标准评析《高抗性淀粉小麦籽粒抗性淀粉含量指标和检测方法》标准解读■ 邓志英1 迟松岐1 于海霞1 胥 倩1 彭 莉2 田纪春1,2*(1.山东农业大学农学院 小麦品质育种研究室;2.山东华田农业科技有限公司)摘 要:《高抗性淀粉小麦籽粒抗性淀粉含量指标和检测方法》(T/Cl 005-2022)是中国国际科技促进会于2022年2月8日发布的团体标准,标准明确规定了高抗性淀粉小麦籽粒中抗性淀粉含量指标及其测定方法,旨在指导育种家培育更好的高抗性淀粉小麦新品种,满足人民群众对生活质量和健康水平的新需求。
文章重点解读《高抗性淀粉小麦籽粒抗性淀粉含量指标和检测方法》标准制定的背景、含量指标制定的依据及其测定方法的核心技术内容,对《高抗性淀粉小麦籽粒抗性淀粉含量指标和检测方法》标准的应用及推广具有重要的指导意义。
关键词:高抗性淀粉小麦,抗性淀粉含量,检测方法,标准DOI编码:10.3969/j.issn.1002-5944.2023.04.025Interpretation of Association Standard on the Index and DetectionMethod of Resistant Starch Content in High Resistant Starch WheatGrainDENG Zhi-ying1 CHI Song-qi1 YU Hai-xia1 XU Qian1PENG Li2 TIAN Ji-chun1, 2*(1. Wheat Quality Breeding Research Group, Agronomy College of Shandong Agricultural University;2. Shandong Huatian Agricultural Technology Co., Ltd.)Abstract: In order to help wheat breeders cultivate better new varieties with high resistance starch content, China International Association for Promotion of Science and Technology issued the association standard on the index and detection method of resistant starch content in high resistant starch wheat grain (T/Cl 005-2022) on February 8, 2022. The standard clearly stipulates the index and determination method of resistant starch content in high resistant starch wheat grain, which will meet the new needs of the people for life quality and health level. This article mainly explains the background of the standard, the basis for the content index and the core technical content of the determination method, which has important guiding signifi cance for the application and promotion of the standard.Keywords: high resistant starch wheat, content of high resistant starch, detection method, standard基金项目:本文受泰安市科技特派员项目(项目编号:2021PTY022)和国家自然科学基金项目(项目编号:31871613)资助。
小麦SBEⅡa、SSⅡa基因片段的克隆及VIGS重组载体的构建

小麦SBEⅡa、SSⅡa基因片段的克隆及VIGS重组载体的构建李淼淼;南富波;刘伟;李卫华【摘要】[目的]克隆小麦SBEⅡa、SSⅡa基因部分序列并构建二者的VIGS重组载体,为在小麦上利用VIGS技术研究SBEⅡa、SSⅡa基因与小麦淀粉合成的关系奠定基础.[方法]根据GenBank中公布的小麦SBEⅡa(AF286319.1)、SSⅡa(AF155217.2)基因序列分别设计特异引物,利用RT-PCR技术克隆SBEⅡa、SSⅡa基因的特异片段.然后以BSMV:γ-PDS为原载体,通过酶切连接用SBEⅡa、SSⅡa基因片段分别将原载体中的PDS基因进行替换,从而构建BSMV:γ-SBEⅡa 和BSMV:γ-SSⅡa重组载体.[结果]克隆的SBEⅡa和SSⅡa基因片段长分别是178 bp和171 bp.序列分析表明:SBEⅡa基因片段与小麦SBEⅡa(AF286319.1)序列同源性为100%;、SSⅡa基因片段与SSⅡa(AF155217.2)序列同源性也为100%;酶切鉴定结果表明成功构建BSMV:γ-SBEⅡa和BSMV:γ-SSⅡa重组载体.[结论]构建的BSMV:γ-SBEⅡa和BSMV:γ-SSⅡa重组载体将为下一步利用VIGS技术在小麦上研究SBEⅡa、SSⅡa基因与小麦淀粉合成的关系奠定基础.【期刊名称】《新疆农业科学》【年(卷),期】2013(050)011【总页数】6页(P1961-1966)【关键词】小麦;SBEⅡa;SSⅡa;VIGS载体【作者】李淼淼;南富波;刘伟;李卫华【作者单位】石河子大学农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子832003;石河子大学农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子832003;石河子大学农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子832003;石河子大学农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子832003【正文语种】中文【中图分类】S512.1;S1880 引言【研究意义】病毒诱导基因沉默(Virus induced gene silencing,VIGS)是近年来发展起来的一种快速、高效应用于研究植物基因功能的反向遗传学新技术[1 ~2]。
基因工程在食品行业中的应用

作业11:RNA干涉(RMAi)基因沉默(gene silencing) 是指转基因植物中特定基因由于种种原因不表达或表达量很低的遗传现象,是近十多年来在转基因植物中发现的一种后生遗传现象.基因沉默大体可以分为两类:位置效应引起的基因沉默和同源依赖的基因沉默。
其中,同源依赖的基因沉默又可以分为转录水平的基因沉默(transeript ional genesilencing, TCS)和转录后水平的基因沉默(post transcriptional gene silencing,PTCS). 前者通常与DN甲基化有关,表现为anRVI不能正常合成,造成基因失活。
后者虽能合成aRV,但随后被降解而不能积累,并同时诱导与外源基因同源的内源性基因沉默。
许多研究表明,转录后水平的基因沉默是引起基因沉默的主要方式。
近年来,随着转录后基因沉默机制的深入探讨,人们能够利用它有目的使特定基因降低表达或不表达。
马铃薯(Solanum tuberosum L. )淀粉是一种重要的食品和工业原料。
尤其是马铃薯直链淀粉,因其特殊的理化性质而被广泛应用。
然而,目前生产上推广的马铃薯栽培品种的直链淀粉含量仅为总淀粉含量的17%,因此,培育高直链淀粉品种对于扩大马铃薯的应用范围和提高其经济价值将具有重要的意义。
用RNA干扰技术(RNA interfence, RMAI) ,以淀粉分支酶(StarchBranching Enzyme, SBE) 基因sbe A和sbe B为靶标进行同时干扰,以期待选有出马铃薯高直链淀粉的新品系,在生产上有其积极的生态意义和经济意义。
2:实验技术路线1:克隆到淀粉分支酶(SBE) 基因: sbe A和sbe B的部分片段。
测序结果表明克隆的sbe A 序列大小为1510该序列与GnBank中已公布的序列的同源性为99%.克隆的she B序列大小为3020,该序列与GnBank中已公布的序列的同源性为9%。
湖北武汉市2023年高考冲刺生物模拟试题含解析

2023学年高考生物模拟试卷请考生注意:1.请用2B铅笔将选择题答案涂填在答题纸相应位置上,请用0.5毫米及以上黑色字迹的钢笔或签字笔将主观题的答案写在答题纸相应的答题区内。
写在试题卷、草稿纸上均无效。
2.答题前,认真阅读答题纸上的《注意事项》,按规定答题。
一、选择题:(共6小题,每小题6分,共36分。
每小题只有一个选项符合题目要求)1.可遗传变异是生物的遗传物质发生改变而导致的变异,但是科学家却发现一些特别的变异:虽然DNA的序列没有改变,但是变异却可以遗传给后代,把这种现象称为表观遗传。
下列关于基因和性状的关系说法错误的是()A.基因可以通过控制蛋白质的结构直接控制生物体的性状,也可以通过控制酶的合成来控制代谢过程进而控制生物体的性状B.基因与基因,基因与基因产物,基因和环境之间相互作用,共同调控生物的性状C.表观遗传中,核内遗传物质在亲子代之间传递不再遵循孟德尔遗传规律D.表观遗传的一种解释:基因在转录和翻译过程中发生了一些稳定性的改变2.如图为神经调节中两个重要的结构模式图,下列选项中错误的是()A.甲图中③内的物质为神经递质,它可以和⑥上的特异性受体结合B.图甲为图乙中结构c的局部放大图像C.图乙中b受损的情况下,刺激a仍有感觉,但是肌肉不能收缩D.图乙中的X所示神经纤维为传入神经3.下图表示蝴蝶兰在正常条件下和长期干旱条件下CO2吸收速率的日变化,据图分析,下列叙述正确的是A.长期干旱条件下,蝴蝶兰可通过夜间吸收CO2以适应环境B.长期干旱条件下,叶肉细胞在10~16时不能进行光反应C.正常条件下,12时CO2吸收速率最快,植株干重最大D.长期干旱条件下,叶肉细胞在0 ~4时不能产生ATP和[H]4.利用人胰岛B细胞构建cDNA文库,然后通过核酸分子杂交技术从中筛选目的基因,筛选过程如图所示。
下列说法错误的是()A.cDNA文库的构建需要用到逆转录酶B.图中的菌落是通过稀释涂布法获得的C.核酸分子杂交的原理是碱基互补配对D.从该文库中可筛选到胰高血糖素基因5.大麦种子吸水萌发时淀粉大量水解,新的蛋白质和RNA分别在吸水后15~20min和12h开始合成。
玉米淀粉分支酶基因SBEⅡb高效沉默表达载体的构建及转化表达

玉米淀粉分支酶基因SBEⅡb高效沉默表达载体的构建及转化表达李楠;赵吉元;尹悦佳;柳青;郭嘉;王云鹏;刘相国;郝东云;贺红霞【摘要】淀粉分支酶( starch branching enzyme, SBE)是淀粉合成的限速酶。
为了进一步研究SBEⅡb沉默对玉米生长及直链淀粉合成的影响,克隆了玉米( Zea mays)淀粉分支酶SBEⅡb基因片段,构建了SBEⅡb的发卡结构表达载体pTFU-SBEⅡb hairpin,用农杆菌介导法将其导入玉米HiⅡ幼胚中,经除草剂筛选获得了194株转化再生植株,其中4株结实,获得转基因种子35粒。
T1代植株经PCR及试纸条检测获得3株阳性材料,半定量RT-PCR结果得出SBEⅡb的表达量降低,推断出基因表达水平降低对直链淀粉的合成具有正效应。
%Starch branching enzyme ( SBE) is a rate-limiting enzyme in the synthesis of starch.To declare the effect of SBEⅡb silencing on maize growth and amylase synthesis, SBEⅡb gene from Zea mays was cloned, and its expression vector pTFU-SBEⅡb containing hairpin structure was constructed.The construct was transformed into maize HiII immature embryos by Agrobacterium-mediated method.194 regenerated plants were obtained through herbicide screening, 4 of them produced 35 seeds. Three positive plants were detected by PCR and BAR strip from T1 generation plants.Semi-quantitative RT-PCR results showed that the expression of SBEⅡb was reduced.And then, we deduced that the reducing level of SBEⅡb has positive effect on amylose synthesis.【期刊名称】《生物技术进展》【年(卷),期】2015(000)002【总页数】7页(P120-126)【关键词】玉米;淀粉分支酶SBEⅡb;ihpRNA载体;直链淀粉【作者】李楠;赵吉元;尹悦佳;柳青;郭嘉;王云鹏;刘相国;郝东云;贺红霞【作者单位】吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033; 吉林农业大学生命科学学院,长春130118;吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033;吉林省农业科学院农业生物技术研究所,长春130033【正文语种】中文玉米(Zea mays L.)是世界栽培广泛的三大作物之一,其产量高于水稻和小麦。
籽粒淀粉合成酶与淀粉合成关系的研究进展

籽粒淀粉合成酶与淀粉合成关系的研究进展王自布;黄燕芬;吴坤;任翠娟;姜金仲【摘要】Starch biosynthesis is a complex biochemical process that involves in a series of enzymes reaction. This paper reviews the function of starch synthase ( SSS ) , starch branching enzymes ( SBE ) , starch debranching enzymes ( DBE ) and their isoenzymes in the process of starch synthesis, and discusses the relationship between starch synthase and starch synthesis pathway in the grain, then prospects the research focus on the studies of complex biochemical reactions in starch synthesis pathway.%淀粉的生物合成是一个复杂的生化过程,涉及一系列酶参与其中。
本文综述了淀粉合成过程中淀粉合成酶( SSS)、淀粉分支酶( SBE)、淀粉去分支酶( DBE)及其同工酶在淀粉合成过程的功能,及与淀粉合成关系的研究进展,并针对淀粉合成途径研究中复杂的生化反应和研究热点进行了展望。
【期刊名称】《生物技术进展》【年(卷),期】2013(000)005【总页数】6页(P336-341)【关键词】小麦籽粒;淀粉;淀粉合成酶【作者】王自布;黄燕芬;吴坤;任翠娟;姜金仲【作者单位】贵州师范学院,贵州省生物资源开发利用特色重点实验室,贵阳550018;贵州师范学院,贵州省生物资源开发利用特色重点实验室,贵阳550018;贵州师范学院,贵州省生物资源开发利用特色重点实验室,贵阳550018;贵州师范学院,贵州省生物资源开发利用特色重点实验室,贵阳550018;贵州师范学院,贵州省生物资源开发利用特色重点实验室,贵阳550018【正文语种】中文植物的籽粒是储藏器官,由于没有叶绿体存在,不能进行光合作用,因此其能量需要通过叶片进行光合作用来提供,叶片通过卡尔文循环固定CO2,在造粉体中合成淀粉,并通过一系列运输途径到达储藏器官,以淀粉形式储存能量。
反义Trxs基因导入对弱筋小麦豫麦18淀粉积累及淀粉合成关键酶表达的影响

作物学报ACTA AGRONOMICA SINICA 2013, 39(10): 1856−1863 / ISSN 0496-3490; CODEN TSHPA9E-mail: xbzw@DOI: 10.3724/SP.J.1006.2013.01856反义Trxs基因导入对弱筋小麦豫麦18淀粉积累及淀粉合成关键酶表达的影响任江萍王亚英王新国王娜陈新孟晓丹李永春尹钧*河南农业大学 / 国家小麦工程技术研究中心, 河南郑州 450002摘要: 以转反义硫氧还蛋白基因小麦TY18-99和TY18-100及其相应未转基因对照为材料, 于2007—2009年通过盆栽试验系统研究了反义Trxs基因(anti-Trxs)导入对弱筋小麦豫麦18籽粒灌浆过程中淀粉积累、淀粉合成关键酶活性以及淀粉合成酶基因表达的影响。
结果表明, 反义Trxs基因对小麦籽粒淀粉积累有一定的正效应。
2个转基因株系的总淀粉和支链淀粉的积累速率较对照显著提高; 在整个籽粒形成过程中, 反义Trxs基因导入显著提高了淀粉分支酶(SBE)活性。
在籽粒灌浆中期和后期, 反义Trxs基因导入显著提高了腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)和可溶性淀粉合酶(SSS)活性, 降低了颗粒束缚型淀粉合酶(GBSS)活性。
实时荧光定量RT-PCR分析表明, 反义Trxs基因促进了AGPase、SBE I和SSS (SSS I、SSS II和SSS III)基因的表达, 抑制了GBSS I基因的表达。
上述结果说明, 反义Trxs基因导入后促进了AGPase、SBE I和SSS基因的转录, 从而使籽粒灌浆期间的淀粉积累速率发生了改变, 最终显著提高了总淀粉和支链淀粉的含量, 降低了直链淀粉含量, 进而提高了淀粉的支/直比, 这可能是转基因小麦淀粉品质改善和产量提高的主要原因。
关键词:小麦; 反义Trxs基因; 淀粉合成酶Effects of Antisense Thioredoxin s on Starch Accumulation and Expressions of Enzymes Related to Starch Synthesis in Weak-gluten Wheat Cultivar Yumai 18 REN Jiang-Ping, WANG Ya-Ying, WANG Xin-Guo, WANG Na, CHEN Xin, MENG Xiao-Dan, LI Yong-Chun, and YIN Jun*Henan Agricultural University / National Engineering Research Centre for Wheat, Zhengzhou 450002, ChinaAbstract: To clarify the functional mechanisms of starch accumulation by the antisense-thioredoxin s (anti-Trxs) gene into wheat, we conducted a pot experiment using two transgenic wheat lines, TY18-99 and TY18-100, and their wild type “Yumai 18” from October 2007 to June 2009. The effects of the anti-Trxs on starch accumulation and key enzymes involved in starch synthesis were compared between the transgenic lines and the wild type. Results indicated that the rates of starch and amylopectin accumulationsin grain were significantly higher in the transgenic lines than in the wild type. During the whole grain-filling period, activity of starch-branching enzyme (SBE) in the transgenic lines increased significantly by 49.2% compared with that in the wild type. At late grain-filling stage, activities of ADP-glucose pyrophosphorylase (AGPase) and soluble starch synthase (SSS) in the transgenic lines increased by 45.8% and 16.0%, respectively. However, activity of granule-bound starch synthase (GBSS) in the transgenic lines had no significant variation. The real-time RT-PCR assay indicated that the introduction of anti-Trxs resulted in increased expressions of SSS (SSS I, SSS II, and SSS III), SBE I, and AGPase genes and reduction of GBSS I transcription. Therefore, the enhanced expressions of AGPase, SBE I, and SSS may be the main reason for increase in starch synthesis in anti-Trxs transgenic lines.Keywords: Wheat; Anti-sense thioredoxin s gene; Starch synthesis enzymes本研究由国家自然科学基金项目(30771332, 30971771)资助。
人工合成淀粉原理

人工合成淀粉原理淀粉是一种广泛存在于植物体内的多糖,是植物体内最主要的储能物质。
淀粉由数千个葡萄糖分子组成,主要分为两种:支链淀粉和直链淀粉。
支链淀粉分支较多,分子量较大,可在植物体内形成颗粒状结构,直链淀粉分子量较小,不容易形成颗粒状结构。
淀粉的结构和性质决定了它在食品、化工、医药等领域的广泛应用。
但是,天然淀粉的生产成本较高,加工难度大,因此人工合成淀粉成为了一个备受关注的研究领域。
人工合成淀粉原理是通过模拟植物体内淀粉合成过程,利用生物技术手段,将淀粉的合成过程转移到微生物体内,从而实现淀粉的人工合成。
淀粉合成过程是一个复杂的生物合成过程,需要多种酶的参与,包括淀粉合成酶、淀粉合成酶辅助蛋白等。
淀粉合成酶是淀粉合成的关键酶,可以将葡萄糖分子逐一连接成淀粉分子。
而淀粉合成酶辅助蛋白则可以调节淀粉合成酶的活性,从而控制淀粉分子的合成速度和分支程度。
在人工合成淀粉的过程中,首先需要筛选出一种合适的微生物,使其具有较高的淀粉合成能力。
目前,已经发现了多种具有淀粉合成能力的微生物,包括大肠杆菌、酵母菌、蓝藻等。
其中,大肠杆菌是最常用的淀粉合成微生物之一,因为其具有较高的淀粉合成能力和易于培养的特点。
接下来,需要在微生物体内引入淀粉合成酶和淀粉合成酶辅助蛋白基因,使其能够进行淀粉合成。
这一步需要利用分子生物学技术,将目标基因克隆到适当的表达载体中,并将其导入到微生物中。
在微生物体内,目标基因会被转录成mRNA,再经过翻译和折叠,最终形成具有活性的淀粉合成酶和淀粉合成酶辅助蛋白。
最后,需要通过调节微生物的培养条件,如温度、pH值、营养物质等,以及添加适当的诱导剂,如异丙基β-D-硫代半乳糖苷(IPTG),来促进淀粉的合成。
在这一过程中,可以利用不同的微生物菌株、不同的淀粉合成酶和淀粉合成酶辅助蛋白组合,来实现不同类型、不同性质的人工合成淀粉。
总之,人工合成淀粉是一项复杂而又具有广泛应用前景的生物技术,其原理是通过模拟植物体内淀粉合成过程,利用微生物进行淀粉合成。
反义RNA

反义RNA技术在食品工业中的应用学院:食品科学技术学院专业:食品质量与安全:学号:2012312674 姓名:祖云鸿摘要:随着基因工程技术的蓬勃发展和代谢调控研究的深入, 反义技术作为一种温和调控的基因工程技术, 开始向世人展示其无穷的魅力。
反义基因的概念、分子生物学基础以及作用原理, 对反义基因技术及其在现代植物研究中的应用进行了概述。
随着现代生物技术的蓬勃发展,极大的推动了食品工业朝着高技术方向房展,主要体现在以下四个方面:首先是以基因工程技术为核心,利用基因工程、细胞工程技术对食品进行改造与改良;二是利用发酵工程、酶工程技术将农副业原材料加工成商品,如酒类、调味品、酸奶类等发酵制品;三是利用生物技术产品进行二次开发,形成新的产品,如功能性低聚糖、食品添加剂等;四是利用酶工艺、发酵技术、生物反应器等对传统食品加工工艺进行改造,降低能耗,提高产出率,改善食品品质【1】。
总之,在以基因工程为核心内容的生物技术领域中,正逐渐形成一门崭新的食品生物技术重要分支学科,并已在食品工业领域中取得丰硕的成果。
Abstract:With the vigorous development of genetic engineering techniques in depth and metabolic regulation study, antisense technology as a mild-control genetic engineering technology, began to show the world its charm.The concept of antisense gene-based molecular biology and the role of principle, the antisense gene technology and its application in modern plant research are outlined. With the rapid development of modern biotechnology, which greatly promoted the food industry moving in the direction of high-tech exhibitions, mainly in the following four areas:First, based on genetic engineering technology as the core, the use of genetic engineering, cell engineering technology to transform and improve food;Second, the use of fermentation engineering, enzyme engineering technology sales of agricultural rawmaterials into goods, such as wine, spices, yogurt and other fermented products category;Third, the use of biotechnology products for secondary development, the formation of new products, such as functional oligosaccharide, food additives, etc.;Fourth, the use of enzyme technology, fermentation technology, biological reactors for the transformation of traditional food processing technology, reduce energy consumption, increase productivity, improve food quality。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
分类号:密级:学号:2009207009 单位代码:10759石河子大学博士学位论文小麦籽粒淀粉生物合成及淀粉分支酶基因(SBE-IIb)的克隆与表达学位申请人王自布齐军仓教授指导教师李卫华教授申请学位门类级别农学博士学科、专业名称作物遗传育种研究方向荒漠绿洲区作物高产优质性状的遗传育种所在学院农学院中国〃新疆〃石河子2012年03月Starch Biosynthesis and Cloning and Expression of Starch Brangching enzymeⅡb (SBE-Ⅱb) in Wheat EndospermA Dissertation Submitted toShihezi UniversityIn Partial Fulfillment of the Requirementsfor the Degree ofDoctor of AgricultureByWang Zi-bu(Crop Breeding and Genetics)Dissertation Supervisor: Prof. Qi Jun-cangProf. Li Wei-huaMarch, 2012石河子大学学位论文独创性声明及使用授权声明学位论文独创性声明本人所呈交的学位论文是在我导师的指导下进行的研究工作及取得的研究成果。
据我所知,除文中已经注明引用的内容外,本论文不包含其他个人已经发表或撰写过的研究成果。
对本文的研究做出重要贡献的个人和集体,均已在文中作了明确的说明并表示谢意。
研究生签名:时间:年月日使用授权声明本人完全了解石河子大学有关保留、使用学位论文的规定,学校有权保留学位论文并向国家主管部门或指定机构送交论文的电子版和纸质版。
有权将学位论文在学校图书馆保存并允许被查阅。
有权自行或许可他人将学位论文编入有关数据库提供检索服务。
有权将学位论文的标题和摘要汇编出版。
保密的学位论文在解密后适用本规定。
研究生签名:时间:年月日导师签名:时间:年月日摘要小麦在我国是仅次于水稻的第二大重要的粮食作物,其产量与品质是直接关系着人类食物的供应程度和营养水平。
小麦籽粒的主要成分是淀粉,淀粉是以碳水化合物的形式储存的,人类80 %左右的能量都是由淀粉提供;淀粉除作为食品外,在工业方面作为原材料的应用更加广泛:造纸、纺织、能源、化工、制药、机械、建筑等行业对淀粉的需求潜力很大,若小麦育种能够向专业化发展,那么不同的行业对淀粉的需求必将会再上一个台阶。
因此深入研究小麦籽粒淀粉的生物合成,为以后利用生物技术来提高淀粉的产量及改良其品质奠定基础,对选育淀粉以及品质专用型的小麦具有重要意义。
为此,本研究选用不同基因型的两个高淀粉和两个低淀粉含量的六倍体小麦为材料,对高、低淀粉含量的小麦品种籽粒灌浆过程中淀粉含量积累的动态变化、淀粉合成关键酶活性的动态变化、淀粉合成关键酶基因表达差异、同时对酶活性的动态变化与淀粉积累动态以及酶基因表达差异之间存在的关系进行了相关性分析。
并且通过反转录的方法克隆得到了高淀粉小麦籽粒胚乳的SBEⅡb 的cDNA,并构建了蛋白表达载体,在大肠杆菌中诱导进行过量表达并对其表达条件进行了优化,还分别构建了SBEⅡb 基因的超量表达载体和SBEⅡb 基因的RNAi 干扰表达载体,并在模式植物中系统的分析了它们的分子特征、表达特性和功能活性等。
主要结果如下:1、小麦籽粒灌浆过程中总淀粉及淀粉组分的积累动态变化,结果表明高淀粉含量的品种和低淀粉含量的品种在籽粒淀粉积累量和积累速率上均存在明显的基因型差异。
Logistic 方程拟合淀粉积累过程发现,支、直链淀粉最终积累量取决于积累启动时间的早晚和积累速率的大小,而最大积累速率出现时间的早晚对其调节作用较小。
2、小麦籽粒灌浆过程中不同类型籽粒中的GBSS、SSS、SBE 和DBE 酶活性变化均呈现单峰曲线,峰值出现在12 -24 d不等。
4 个品种的GBSS 酶活性均与直链淀粉的积累速率呈极显著正相关,与总淀粉的积累速率达到显著或极显著正相关,但与支链淀粉的积累速率的相关性不显著。
SBE 和SSS 酶活性与总淀粉的积累速率和支链淀粉的积累速率呈显著或极显著正相关,而与直链淀粉积累速率的相关不显著。
DBE 活性与淀粉积累速率的相关性不显著,淀粉合成关键酶活性间的相关性分析表明高淀粉含量的品种DBE 活性与SSS、GBSS 和SBE 达到显著性或极显著性水平,它们之间可能存在着协同作用。
3、采用实时荧光定量RT-PCR 方法,对灌浆期籽粒淀粉合成酶相关基因的转录水平进行了检测和比较,并对其表达量与相应酶活性的相关性做了分析。
结果表明,束缚态淀粉合成酶基因(GBSS)、可溶性淀粉合成酶基因(SSS)、淀粉分支酶基因(SBE)、淀粉去分支酶(DBE)酶活性均呈单峰曲线变化,利用实时荧光定量PCR 技术检测9 个基因在不同淀粉含量的4 个供试品种花后不同时期的相对表达量,结果表明,在花后6 d这些基因开始表达,在灌浆的中期花后12 -18 d不等有表达的小高峰,两个高淀粉含量的品种在不同时期的酶活性及其酶基因的相对表达量均比两个低淀粉含量的品种相对较高;这些基因表达谱与酶活性相关分析显示,除GBSS 外其它几种淀粉合成酶基因均与相应酶活性呈显著或极显著正相关,而且GBSS酶活性到达峰值时间稍迟于DBE、SSS、SBE 等酶,说明DBE、SSS、SBE 基因可能主要通过转录水平来控制籽粒淀粉的合成,而GBSS 基因可能主要通过转录后水平来控制籽粒淀粉的合成。
I4、采用RT-PCR 技术克隆了小麦胚乳基因SBEⅡb 全长cDNA 序列,同时构建了原核生物表达载体pET28(a+)-SBEⅡb,对不同的IPTG 诱导浓度和诱导时间等条件的优化结果表明,37 ℃,8 h 能诱导SBEⅡb 融合蛋白的最大量表达,在0.5 mmol/L IPTG 浓度下可以成功表达出了SBEⅡb 蛋白。
为进一步体外研究SBEⅡb的物理化学性质及催化机理奠定基础。
5、构建SBEⅡb 的RNAi 表达载体,并对其转化烟草,检测其对直链淀粉合成的影响。
以植物表达载体PZP35S 为基础,构建了由组成型启动子35S调控的SBEⅡb 基因的RNAi 植物表达载体。
实时荧光定量PCR (qRT-PCR)检测其在转基因烟草中的相对表达量,并测定转基因植株中直链淀粉的含量和支链淀粉含量的变化。
结果表明:经qRT-PCR 证实SBEⅡb 基因的表达量较对照下降,而且支链淀粉含量变化显著,直链淀粉较对照有上升但变化没有达到显著水平。
6、构建SBEⅡb 基因的过量表达载体,将克隆得到的基因SBEⅡb 构建到了植物表达载体pPZP35S 上,通过农杆菌叶盘转化法转化烟草叶片,经卡那霉素筛选、PCR 扩增鉴定证明,SBE-Ⅱb 基因已成功转入烟草细胞中,获得4 个转基因植株。
RT-PCR 半定量和实时荧光定量RT-PCR 分析表明,转基因烟草株系的叶片中有SBEⅡb 基因的表达,与对照植株相比,转SBEⅡb 基因的植株中的转录本达到了显著性的变化,但是其淀粉分支酶活性没有达到显著性变化。
关键词:小麦;淀粉;淀粉合成酶;实时荧光定量PCR;基因表达IIAbstractWheat is the second most important food crop next to rice in china, and its output and quality are directly related to the human food supply degree and nutrition level. The major components of the wheat grain is starch, which exists in the form of carbohydrate, and about 80 % of all human energy is provided by the starch. Starch out as food outside, in the industrial sector as the raw material of the application more widely: paper, textile, energy, chemical, pharmaceutical, machinery, building and so on. So many industries to the needs of the starch is a potential, especially after the professional development of wheat breeding, different industries to the needs of the starch will again on a higher level. So further research of wheat kernels starch biosynthesis for later use biological technology to improve the production and improving their starch quality lays the foundation, and the special quality of breeding starch of wheat to have the important meaning. Therefore, this research choose different genotypes of two high starch and two low starch content of hexaploid wheat for material, and the first objective of this study was to analyse the dynamic accumulation of starch, amylose and amylopectin and to determine the patterns of the activities of key enzymes involved in starch synthesis and the ralationship to the accumulation speed of starch, amylose and amylopectin during grain filling period. The second objective was to analyze expression profiles of starch synthesis genes by qRT-PCR during endosperm development in wheat. The third objective was to clone the cDNA of SBEⅡb, and then construct the original nuclear expression vector and optimize expression condition of Prokaryotic Expression in E.coil. Constructing the interference of expression vector and overexpression vector, and then in the mode of plants to analyze and their molecular features, expression characteristic and function activity, etc.1. Correlation analysis indicated that there were obvious genotype difference between lower starch content and higher starch content starch about starch accumulating rate and Kernel filling rate. The result showed that the accumulation courses of amylopectin, amylose and total starch were well fitted to the logistic equation by relating amylopectin, amylose and total starch contents against days after pollination (DAP). The simulation parameters revealed that the higher contents of amylopectin and amylose resulted from earlier initiating accumulation time and greater accumulation rate.2. The results indicated that the activities of soluble starch synthase (SSS), granule bound starch synthase (GBSS), starch branching enzyme (SBE) and starch debranching enzyme (DBE) changed in the pattern of a single peak curve during grain filling, peak value is about between 12 d and 24 d; The accumulation rate of amylose was highly significantly correlated with the activities of GBSS, and the accumulation rate of amylopectin was significantly or sharp significantly correlated with the activities of SSS and SBE; and the accumulation rate of starch was not correlated with the activities of starch debranching enzyme (DBE), among activities of the starch synthesis enzymes correlation analysis indicated that starch debranching enzyme (DBE) of higher starch content was significantly or sharp significantly correlated with the activity of SSS、SBE and GBSS, which were interacted in starch biosynthesis.III3. The expression levels of genes were detected by real-time PCR. The results indicated that the starch enzymes activity of granule bound starch synthase ( GBSS) , soluble starch synthase gene ( SSS) , starch branching enzyme gene ( SBE), starch debranching enzyme(DBE)demonstrated a single peak curve during grain filling. To test the relative expression values of the genes in nine different grain filling stages, 6 day after pollination (DAP), 9d, 12d, 15d, 18d, 21d, 27d, 24d, 30d, by real-time quantitative RT-PCR(qRT-PCR) , and the data of starch enzymes activity from 4 wheat varieties, during grain filling middle stage, 12 -18 DAP reach reached their peaks and began to drop. The expression profile showed that starch biosynthesis enzymes genes all had high expression level in earlier stage of endosperm development. The relative expression levels of GBSS, SSS, SBE and DBE in high starch wheat varieties were higher than those in low starch wheat varieties. The correlate analysis indicated that in addition to GBSS, the relative expression level s of SBE, SSS and DBE were significantly or highly correlated with the activities of SBE, SSS and DBE at 15 DAP.The time of GBSS reached peak values were later than others. These suggested that SBE, SSS, DBE may control starch synthesis at the transcriptional level, and GBSS may control starch synthesis at the post transcriptional level.4. The cDNA of SBEⅡb cDNA from wheat endosperm was cloned by RT-PCR. The SBEⅡb cDNA were inserted pET28a (+) vector by molecular biological technologies, including restriction endonuclease digestion, electrophoresis, gel reclamation DNA ligation and so on, and then transformed to E. coil BL21 (DE3). The SBEⅡb fusion protein was expressed and the expression conditions were optimized. Through the research on optimization of expression induction time and the concentration of IPTG and so on, and results showed the expression level of SBEⅡb fusion protein reached the highest for 8 hours cultured at 37℃and 0.5 mmol/L IPTG can effectively induced the expression of SBEⅡb in prokaryotic expression system. The experimental result lay a foundation for further study on the physical and chemical properties of SBEⅡb in vitro.5. To obstain RNAi expressive vector of SBEⅡb gene, this integrated in tobacco genome and studies its effect on amylase content. RNAi vector was constructed with pPZP35S and pUCm-T, and Expression vector RNAi containing 35S promoter to regulate gene expression. qRT-PCR analysis results indicated that the SBEⅡb gene expression value is lower than CK (check plants) in the leaves of all transformed plants. Comparing with the CK about starch content, the amylopectin content of the transformed plants reached significantly changes, however, the amylose showed no significant difference.6. The plant expression vector for pPZP35S was constructed. The plasmid was transformed into tobacco by leaf dish transformation, and four transgenic plants are obtained by kanamycin selecting and PCR analysis, and the results demonstrated that SBEIIb gene was transformed into tobacco cells, and gained four transformed plants, successfully. SqRT-PCR and qRT-PCR analysis results indicated that the SBEIIb gene expressed in the leaves of all transformed plants. Comparing with the check plants, the transcriptional level of the transformed plants reached significantly changes, however, the Starch branching enzyme activity showed no difference between them.Key words:Wheat (Triticum aestivum L.); Starch; Starch Synthase; Qualitative Real-Time PCR; Gene ExpressionIV目录摘要 (I)Abstract (III)目录 (V)缩略词 (VII)引言 (IX)第一章文献综述 (1)1.1 小麦籽粒的来源、结构及其组分 (1)1.2 籽粒淀粉的组成、结构及性质 (2)1.2.1 直链淀粉 (3)1.2.2 支链淀粉 (3)1.3 籽粒淀粉的生物合成 (5)1.3.1 淀粉合成酶(SS) (6)1.3.2 淀粉分支酶(SBE) (11)1.3.3 淀粉去分支酶(DBE) (13)1.4 淀粉合成同工酶及其与淀粉的关系 (15)1.4.1 SS同工酶及其与淀粉的合成 (15)1.4.2 SBE同工酶及其与支链淀粉的合成 (19)1.4.3 DBE同工酶及其与淀粉的合成 (22)1.5 籽粒淀粉合成中关键酶基因的表达 (24)1.6 淀粉品质的遗传改良 (26)1.7 本研究的目的和意义 (27)第二章含量不同的两类小麦淀粉积累及关键酶活性变化的研究 (28)2.1 材料与方法 (29)2.1.1 材料与种植 (29)2.1.2 田间取样 (29)2.1.3 测定项目、方法 (29)2.1.4 数据统计与分析 (29)2.2 结果与分析 (29)2.2.1 籽粒淀粉含量的动态变化 (29)2.2.2 淀粉合成关键酶活性的变化 (33)2.2.3 淀粉积累速率和酶活性间的相关性分析 (36)2.3 讨论 (37)2.3.1 不同淀粉类型的小麦品种籽粒淀粉的积累特征 (37)2.3.2 淀粉合成的关键酶 (37)第三章小麦籽粒胚乳淀粉合成酶基因表达及酶活性分析 (39)3.1 材料和方法 (39)3.1.1 材料 (39)3.1.2 方法 (40)3.1.3 数据统计与分析 (40)3.2 结果与分析 (41)3.2.1 RT-PCR 结果 (41)V3.2.2 9个基因在两类不同淀粉含量的小麦品种中的表达谱 (42)3.2.3 籽粒胚乳淀粉合成酶活性的差异 (44)3.2.4 淀粉合成酶基因表达相对量与对应酶活性的相关性 (45)3.3 讨论 (45)3.3.1 实时荧光定量PCR (46)3.3.2 淀粉合成酶及其酶基因的表达 (46)3.3.3 高淀粉含量品种和低淀粉含量含量品种淀粉含量差异的机理 (47)第四章小麦胚乳基因SBEⅡb cDNA的克隆与原核表达条件优化 (48)4.1 材料和方法 (48)4.1.1 材料 (48)4.1.2 试验方法 (48)4.2 结果与分析 (50)4.2.1 SBEⅡb 基因cDNA 的扩增 (50)4.2.2 阳性克隆的菌液PCR 鉴定 (52)4.2.3 SBEⅡb cDNA 原核生物表达载体构建 (52)4.2.4 诱导表达 (52)4.3 讨论 (54)第五章小麦胚乳淀粉分支酶基因(SBEⅡb)功能初步分析 (57)5.1 材料与方法 (57)5.1.1 材料与试剂 (57)5.1.2 试验方法 (58)5.2 结果与分析 (59)5.2.1 SBEⅡb基因cDNA克隆及鉴定 (59)5.2.2 pZP35S-SBEⅡb 转化烟草 (60)5.2.3 SBEⅡb基因干扰片段及正反插入序列的酶切鉴定 (61)5.2.4 SBEⅡb基因的功能分析 (63)5.3 讨论 (67)5.3.1 淀粉合成酶基因在淀粉改良中的应用 (67)5.3.2 RNAi在淀粉品质改良中的应用 (68)第六章结论与展望 (69)6.1 结论概述 (69)6.2 主要创新点 (69)6.3 展望 (70)参考文献 (71)致谢 (84)个人简介 (85)导师评阅表 (86)VI缩略词英文缩写英文名称中文名称6-BA 6-benzyladenine 6-苄基腺嘌呤Amp Ampicillin 氨苄青霉素AGPase ADP-glucose pyrophosphorylase 腺苷二磷酸葡萄糖焦磷酸化酶ADP-G Adenoslne diphosphate glucose 腺苷二磷酸葡萄糖BSA Bovine serum albumin 牛血清白蛋白bp base pair 碱基对CB Carbenicillin 羧苄青霉素DAP Days after pollination 花后天数DBE Starch-debranching enzyme 淀粉去分支酶DEPC Diethyl Pyrocarbonate 焦炭酸二乙酯DNA Deoxyribonucleic acid 脱氧核糖核酸DMSO Dimethyl sulfoxide 二甲基亚砜dNTP deoxyribonucleoside tripospate 脱氧核苷三磷酸DTT Dithiotheitol 二硫苏糖醇EB Ethidium bromide 溴化乙锭E. coli Escericia coli 埃希氏大肠杆菌EDTA Ethylenediamine tetracetic acid 乙二胺四乙酸EDTA Ethylendiaminotetraacetic acid 乙二胺四乙酸GBSS Granule-bound starch synthase 束缚态颗粒型淀粉合成酶GL Glycerin 甘油GPM Phosphoglucomutase 磷酸葡萄糖变位酶GUS Glucuronidase β-葡糖甘酸酶G-250 Coomassie brilhant blueG-250 考马斯亮蓝Hepes N-[2-hydroxyethl]piperazine-N’[2-ethanesulfonic acid]N-2-羟乙基哌嗪N’-2-乙磺酸HK Hexokinase 己糖激酶IPTG Isopropylthio-β-D-galactoside 异丙基-β-D-硫代半乳糖Iso Isoamylase 异淀粉酶kDa kilodalton 千道尔顿Kana Kanamycin 卡那霉素LB Lyria broth LB培养基MS Murashige and Skllg medium MS培养基VIINAA Naphthalene acetic acid 萘乙酸NaAC Sodium acetate 醋酸钠ORF Open Reading Frame 开放阅读框PAGE Polymeric acrylic 聚丙烯酰胺PEP Phosphoenolpyruvate 磷酸烯醇式丙酮酸PK Pyruvate kinase 丙酮酸激酶PVP Polyvinyl lpyrrolidone 聚乙烯吡咯烷酮Pul Pullulanase 极限糊精酶qRT-PCR Quantitative real-time PCR 实时定量PCR Rif Rifampicin 利福平RT-PCR Reverse transcerption PCR 反转录PCR SBE Starch-branching enzyme 淀粉分支酶SDS Sodium dodecyl sulfate 十二烷基硫酸钠SS Starch synthase 淀粉合成酶SQ-PCR Semi-quantitative polymerase chainreaction 半定量PCRTE Tris-CL,EDTA buffer Tris-CL,EDTA 缓冲液TEMED Tetramethylethylenediamine N,N,N',N'-四甲基乙二胺X-gal 5-Bromo-4-chloro-3-indolyl-β-D-galactoside5-溴-4氯-3-吲哚-β-D-半乳糖苷引言淀粉是高等植物碳水化合物的主要储存形式。