纤维素的结晶结构
纤维素的结晶结构

Meyer-Misch 模型 ① 晶胞参数: a=8.35Å,b=10.3Å ,c=7.9Å
β=84º纤维素分子链只占据结晶单元 的4个角和中轴,而每个角上的链为4 个相邻单位晶胞所共有,即每个晶胞 只含2个(4*1/4+1)链单位。 ② 中间链和位于角上的链走向相反,轴 向高度差半个葡萄糖基。b轴的长度 是纤维二糖的长度,这些链围绕着纵 轴扭转180°。
3.折叠链结晶结构理论 (fringed-micelle with chain-folding)
纤维素大分子有呈折叠链状的可能,这些折 叠链状的线型纤维素大分子仍然能形成结晶 结构,即所谓折叠链结晶,或所谓片晶。人 们认为片晶就如同缨状微胞结构中的微胞; 伸出的分子就像缨状分子。
片的厚度即相当于沿纤维轴向出现的100 ~200 Å的自同周期的大小(X射线衍射发现 纤维在纵向上存在周期)。
➢ 褐腐材的结晶度较正常材低
① 重氢取代法(氘化法)
对纤维素进行重氢化反应,控制一定条件,使重氢只取代无定形区中 的OH,使OH变成OD,而结晶区中的OH不参加反应,3500cm-1 谱带(OH)下降,而在2530 cm-1(OD)处出现OD吸收谱带, a3500cm-1/a2530 cm-1称为结晶度指数。
相 对 强 度
X射线管结构
X-射线衍射的基本原理
晶体是由原子或原子基团等按照一定规律在空间内有 规则排列而构成的固体。当它被X射线照射后,各个原 子散射X射线。这些散射线符合相干波的条件,因而产 生干涉现象。所谓X射线衍射,实质上就是研究这些散 射波的干涉。衍射线就是经过相互干涉而加强的大量散 射线所组成的射线。
纤维素结构

纤维素的结构引言纤维素是地球上存在的最丰富的可再生有机资源, 在高等植物、细菌、动物、海藻等生物中广泛存在, 每年总量有几百亿吨, 具有巨大的经济开发价值[1]。
五十年代至六十年代,由于合成高分子材料的兴起,纤维素资源的开发研究受到极大的影响。
七十年代初期,由于国际上出现了石油危机,这种曾被忽视的可更新资源又再次被重视起来.能否利用这些丰富的可再生资源是解决未来能源问题的关键因素。
因此,世界各国都很重视纤维素的研究与开发[2]。
纤维素结构是纤维素性能研究及应用的基础,本文就纤维素的化学剂物理结构进行了概述。
1纤维素的化学结构纤维素的元素组成为:C=44.44%,H=6.17%,O=49.39%, 其化学实验式(C 6H 10O 5)n (n 为聚合度,一般高等植物纤维素的聚合度为7000—150000)[3]纤维素大分子的基环是脱水葡萄糖,其分子式为(C 6H 10O 5)。
纤维素的化学结构是由D-吡喃葡萄糖环彼此以β- 1, 4-糖苷键以C1椅式构象联结而成的线形高分子化合物[4],其结构表达式如图1所示。
非还原端 纤维二糖 还原端图1 纤维素链结构除两端的葡萄糖基外,每个葡萄糖基上都有三个游离羟基,分别位于C 2、C 3和C 6位上,所以纤维素的分子可以表示为[[C 6H 7O 2(OH)3]n,其中C 2和C 3位上为仲醇羟基,C 6位上为伯醇羟基,他们的反应能力不同,对纤维素的性质具有重要影响,如纤维素的酯化、醚化、氧化和接枝共聚,以及纤维素之间的分子间氢键作用,纤维素的溶胀与水解都与纤维素的羟基有关。
纤维素大分子两端的葡萄糖末端基,其结构和性质不同,一端的葡萄糖末端基在C4上存在一个苷羟基,此羟基的氢原子易转移,与基环上的氧原子结合,使氧环结构转变为开链式结构,在C1处形成醛基,具有潜在还原性,固有隐形醛基之称。
左端的葡萄糖末端为非还原性的,由于纤维素的每一个分子链一端是还原性,另一端是非还原性,所以纤维素分子具有极性和方向性。
纤维素(一) - 纤维素(一)

5.2 纤维素
(1)纤维素的化学结构
5.2 纤维素
(1)纤维素的化学结构
两种类型的氢键:
➢ 分子内氢键,能赋予单 一分子链一定的刚度;
➢ 分子间氢键,对纤维素 超分子结构的形成有重 要作用。
5.2 纤维素
(1)纤维素的化学结构 羟基→氢键: ➢范德华力 < 氢键 < 共价键
氢键的键能: 20~35千焦/摩尔 范德华力键能:10~20千焦/摩尔 C-O-C主键力: 300~400千焦/摩尔
的距离缩短,形成氢键和范德华力。
5.2 纤维素
(2)纤维素的物理结构—微细结构
➢ 基原纤(elemental fibril):一般由几根至几十根长
链分子,互相平行或螺旋状地按一定距离、相位 稳定地结合在一起的大分子束,直径1~3 nm, 具有一定的柔曲性。
5.2 纤维素
(2)纤维素的物理结构—微细结构
附加知识——纳米纤维素应用
➢被广泛应用于基于纳米纤维素的增强聚合物复合材料、 药物载体、食品、化工产品、生物支架材料、贵重金属 纳米材料合成模板等研究。
5.2 纤维素
(2)纤维素的物理结构—结晶结构
纤维素超分子结构的二相体系理论 纤维素的结晶体聚集态结构 纤维素的结晶度计算 纤维素的可及度
Relative Crystallinity / %
5
10
15
20
25
30
35
40
2 Theta / degree
➢ 原纤(fibril)、微原纤 (microfibril):由若干
根基原纤平行排列在一 起的较粗的大分子束, 直径为12~30nm。
5.2 纤维素
(2)纤维素的物理结构
纤维素概况简介

纤维素相关的专利数量也很多,涉及纤维素的制备、改性、应用等方面。
相关行业报告与统计数据
行业报告
一些权威机构和行业协会发布了一系列 关于纤维素及其相关领域的行业报告和 统计数据,涉及市场规模、发展趋势、 竞争格局等方面。
VS
统计数据
一些政府部门和权威机构发布了一系列关 于纤维素及其相关领域的统计数据,涉及 产量、消费量、进出口等方面。
纤维素可以作为食品添加剂,增加食品的口感、 营养价值和饱腹感。
保健食品
某些特殊纤维素的提取物,如菊粉、葡聚糖等, 具有改善肠道健康、降低血糖等保健功能。
替代脂肪
某些高纤维食品可以作为脂肪的替代品,有助于 控制热量摄入和预防肥胖。
纤维素在医药工业中的应用
药物载体
纤维素可以作为药物载体,用于药物缓释和靶向给药系统。
• 纤维素具有高度的吸水性,可以吸收大量的水分并形成凝胶状物质,这 使得它在食品加工和药物制造中具有一定的应用价值。
• 纤维素具有很好的透气性和透湿性,可以作为纺织品和纸张的原料,也 可以用于制造过滤材料和防水材料等。
02
纤维素来源与分布
天然纤维素来源
植物细胞壁
纤维素是植物细胞壁的主要成 分,占植物体干重的比例高达
纤维素在纸张制造中的应用
增强纸张强度
纤维素能够提高纸张的抗 张强度,使纸张更加耐折 、耐磨,延长使用寿命。
提高纸张吸墨性
纤维素具有亲水性,能够 提高纸张的吸墨性能,使 印刷更加清晰、流畅。
降低生产成本
纤维素来源于天然植物, 相比合成材料,可以降低 纸张制造的成本。
纤维素在食品工业中的应用
食品添加剂
纤维素纳米纤维是一种新型纳米 材料,具有优异的力学性能、高 比表面积和良好的生物相容性, 广泛用于复合材料、生物医学、 环境治理等领域。
纤维素(三) - 纤维素(三)
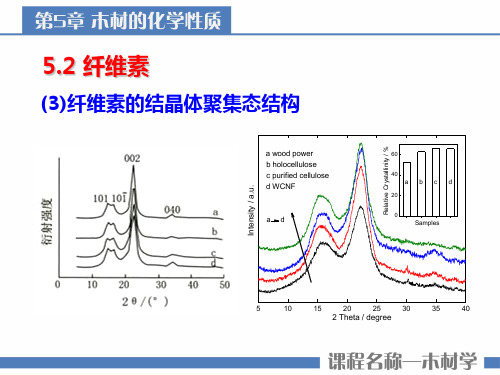
(3)纤维素的结晶体聚集态结构
a wood power b holocellulose c purified cellulose d WCNF
ad
60
40 abcd
20
0 Samples
Intensity / a.u.
Relative Crystallinity / %
5
10
15
20
25
5.2 纤维素
(4)纤维素的物理性质
表面电化学性质:
①羟基和醛基可使纤维素表 面在水溶剂中带负电;
②正电子由于热运动结果在 纤维表面由近而远有一浓 度分布。
(a)离子层 (b)吸附层 (d) 扩散层
5.2 纤维素
(4)纤维素的物理性质
表面电化学性质:
在湿法纤维板制造工艺中,为了减少纤维的吸湿,一 般在纤维板的浆料中施加石蜡乳液进行阻水处理。但 石蜡微粒带有负电荷,于浆料中加入硫酸铝作沉淀剂 ,使石蜡留存在纤维表面上,经处理的浆料制成的纤 维板,防水性能和体积稳定性均有所改善,提高产品 质量。
5.2 纤维素
(5)纤维素的化学性质
纤维素的降解 氧化降解:
例:2,2,6,6-四甲基哌啶氧自由基(TEMPO)的选择性催化氧化
纤维素经氧化剂作用后,羟基氧化成醛基、酮基或 羧基,形成氧化纤维素。随着官能团的变化,纤维素 的聚合度也同时下降,发生氧化降解后,纤维素的机 械强度降低。
5.2 纤维素
5.2 纤维素
(5)纤维素的化学性质
纤维素的酯化
如醋酸酐与纤维素的-OH基作用
[C6H7O2(OH)3]n+3n(CH3CO)2O→ [C6H7O2(OCOCH3)3]n + 3nCH3COOH
生物质结构化学——第三章 纤维素(1)

(3) 结晶度和可及度的关系:
A = a + ( 100 – a )
A—可及度
—结晶区表面的纤维素分子数
a—结晶度
(4) 测定方法
物理法
结晶度:X-射线衍射法、红外光谱法、密度法
可及度:水解法、重水交换法、甲酰化法
化学法
4、纤维素的细纤维结构
纤维素织态结构模型
结晶区
非结晶区:无定形区
纤维素大分子是一种结晶区和无 定形区交错结合的体系,从结晶区到 无定形区是逐步过渡的,无明显界限, 一个纤维素分子链可以经过若干结晶 区和无定形区。每一个结晶区称之为 微晶体,也称之为胶束或微胞。
OH
OH
HO HO
α -型
O
OH OH
HO HO
在直立键上
β -型
O
OH OH
在平伏键上
为什么在溶液中达到平衡时,β型占64%,而α型仅占36%?为什么β 型在酸中水解速度比α型小的多??
(8)纤维素二糖是支配纤维素分子构象的基本单 元,它的长度约为100nm,即单位为100nm的小分 子重复聚合而成为大分子。
4.1 纤维素织态结构模型
结晶区
1条分子穿过 若干结晶区 和非结晶区
无定形区
(1)结晶区的特点 :
纤维素分子链取向良好,密度较大,结晶区纤维素的 密度为1.588g/cm3,分子间的结合力强,故结晶区对强度的 贡献大。
(2)非结晶区的特点:
纤维素分子链取向较差,分子排列秩序性差,分子间
距离较大,密度小,无定形区纤维素密度为1.50g/cm3。且
分子间氢键结合数量少,故无定形区对强度的贡献小。
四、纤维素大分子间的氢键及其影响
1.氢键的形成及其性质
纤维素的结晶结构

:s与a之夹角
点阵基矢(原子间距)a
入射线单位矢量s0
0:s0与a之夹角
一维劳埃方程的导出
原子列中任意两相邻原子(A与B)散射线间光程差()为: =AM-BN=acos-acos0 散射线干涉一致加强的条件为=H,即: a(cos-cos0)=H H——任意整数
纤维素X与纤维素IV晶胞参数大致相等。
纤维素结晶变体的相互转化
+液体氨
蒸发
纤维素I
+H2O 100º C +NaOH 100º C +NaOH 20º C
NH3-纤维素I
≥200º C
纤维素ⅢI
Na-纤维素II
+NaOH
Na-纤维素III
+H2O +液体氨 +H2O 蒸发
纤维素IVI
纤维素II
NH3-纤维素II
CHINESE ACADEMY OF FORESTRY
纤维素结晶结构
组员:郭 飞 张毛毛 周贤武 温留来 张龙飞
2012-12-04
1
3.纤维素的物理结构
3.1 纤维素的结晶结构 晶体的基本概念
晶体:物质内部质点(原子、分子、离子)呈规律排列的固体, 为“空间格子”状。 晶胞:晶体的最小重复单位。可以用3个晶轴的长度a、b、c 及其夹角α 、β 、γ 6个晶胞参数来描述。
或
a· 0)=H (s-s b· 0)=K (s-s c· 0)=L (s-s 三维晶体若要产生衍射,必须同时满足上述三个方程
Scherrer公式
Scherrer公式用于样品晶粒尺寸的计算
D=K·λ /(Bhkl ·cosθ)
其中,D为沿垂直于晶面(hkl)方向的晶粒直径,
纤维素水解

纤维素水解
纤维素水解是一个广泛应用于工业和生物科学领域的过程。
纤维素是一种多糖
类聚合物,主要存在于植物细胞壁中,包括木质素和纤维素。
纤维素水解是将纤维素分解为更简单的单糖,如葡萄糖,以便更好地利用其作为生物质资源。
纤维素的结构
纤维素是由葡萄糖分子通过β-1,4-糖苷键连接而成的线性多糖,具有高度的结
晶性和稳定性。
这种结构赋予了纤维素出色的机械强度和耐久性,同时也增加了其降解的难度。
纤维素水解的方法
纤维素水解通常采用酶解法和酸解法两种主要方法。
酶解法
酶解法是目前应用最为广泛的纤维素水解方法之一。
在酶解过程中,纤维素酶
通过降解纤维素的β-1,4-糖苷键来将纤维素水解为葡萄糖。
常用的纤维素酶包括纤
维素酶、β-葡聚糖酶等。
酶解法具有选择性高、反应条件温和等优点,但同时也存在酶的稳定性、成本等方面的挑战。
酸解法
酸解法是另一种纤维素水解的方法,通过在酸性条件下将纤维素水解成葡萄糖。
常用的酸包括硫酸、盐酸等。
酸解法具有操作简单、反应速度快等优点,但会产生大量的废弃物,并对环境造成污染。
纤维素水解的应用
纤维素水解是生物质能源利用的重要途径之一。
通过将纤维素水解成葡萄糖,
可以进一步转化为乙醇、生物柴油等可再生燃料。
同时,纤维素水解产生的糖类还可以用于生物化学品和生物材料的生产,促进生物经济的发展。
纤维素水解技术的不断发展将为可再生能源和生物资源开发提供更多可能性,
促进绿色和可持续发展的实现。
天然纤维素结晶变体I结晶变体II简谈

天然纤维素结晶变体I结晶变体II简谈摘要:纤维素(celluloe)是由葡萄糖组成的大分子多糖。
不溶于水及一般有机溶剂。
是植物细胞壁的主要成分。
在自然。
具有一定构象的纤维素高分子链按一定的秩序堆砌,便成为纤维素的微晶体,微晶体的组成单元称为晶胞。
在纤维素中存在着化学组成相同,而单元晶胞不同的同质多晶体(结晶变体),常见的结晶变体有四种,即纤维素Ⅰ、Ⅱ、Ⅲ、Ⅳ。
本文将这种介绍纤维素Ⅰ、纤维素Ⅱ之间的转化。
关键字:纤维素结晶变体转化结构1、简述纤维素纤维素是由D-吡喃型葡萄糖基(失水葡萄糖)组成。
简单分子式为(C6H10O5)n;化学结构式可用下二式表示:霍沃思式是由许多D-葡萄糖基(1-5结环),藉1-4,β-型联结连接起来的,而且连接在环上碳原子两端的OH和H位置不相同,所以具有不同的性质。
式中n为聚合度。
在天然纤维素中,聚合度可达10000左右;再生纤维素的聚合度通常为200~800。
在一个样品中,各个高分子的聚合度可以不同,具有多分散性。
椅式由于内旋转作用,使分子中原子的几何排列不断发生变化,产生了各种内旋转异构体,称为分子链的构象。
纤维素高分子中,6位上的碳-氧键绕5和6位之间的碳-碳键旋转时,相对于5位上的碳-氧键和5位与4位之间的碳-氧键可以有三种不同的构象。
如以g表示旁式,t表示反式,则三种构象为gt、tg、和gg。
多数人认为,天然纤维素是gt构象,再生纤维素是tg构象。
在纤维素分子链中,存在着氢键。
这种氢键把链中的O6(6位上的氧)与O2'以及O3与O5'连接起来使整个高分子链成为带状,从而使它具有较高的刚性。
在砌入晶格以后,一个高分子链的O6与相邻高分子的O3之间也能生成链间氢键。
2、纤维素结晶变体及其结构纤维素的聚集态结构是研究纤维素分子间的相互排列情况(晶区和非晶区、晶胞大小及形式、分子链在晶胞内的堆砌形式、微晶的大小)、取向结构(分子链和微晶的取向)等。
天然纤维素和再生纤维素纤维都存在结晶的原纤结构,由原先结构及其特性可部分地推知纤维的性质,所以为了解释以纤维素为基质的材料的结构与性能关系,寻找制备纤维素衍生物的更有效方法,则研究纤维素合成的机理、了解纤维素的聚集态结构,在理论研究和实际应用方面都有重要的意义。
纤维素知识 1

互穿聚合物网络(IPN)天然纤维素包括细菌纤维素、海藻和高等植物(如棉花、苎麻、木材等)均属于纤维素I型。
纤维素I分子链在晶胞内是平行堆砌的,纤维素II是纤维素I经由溶液中再生(regeneration)或经丝光处理(mercerization)得到的结晶变体,是工业上使用最多的纤维素形式。
纤维素II 与纤维素I有很大的不同,它是由两条分子链组成的单斜晶胞,属于反平行链的堆砌。
纤维素是一种β-(1-4)-D-糖苷键连接的线型高聚物,由X 射线衍射发现存在四种结晶形态,即纤维素Ⅰ、Ⅱ、Ⅲ和Ⅳ,不同晶型纤维素的C1,C4 和C6 的化学位移具有明显的差别,这种差别可能是因为不同晶型纤维素的链构象转变或晶体堆砌对吡喃葡萄糖单元C4 和C6 的影响差异造成的.基于在非晶区的链段运动显示窄谱线,而晶区的刚性链以及分布在非晶区的刚性链产生宽谱线,因此可以利用宽、窄谱线的峰面积(S b 和S n)求取结晶度χ c纤维素I和纤维素II在C6上的差别就是因为吡喃葡萄糖单元C6位羟基的构象不同,纤维素I为t - g构象,纤维素II和无定型纤维素则为g - t构象。
天然纤维素I也存在两种不同的晶体结构,即纤维素Iα和Iβ。
13C NMR谱指出它们之间最大的差别在C1的化学位移上,Iα 为单峰,Iβ 为双峰。
高分子链构象参数:无扰尺寸(A):A 值愈小,高分子链愈柔顺。
A 值只取决于高分子的近程结构,与高聚物的分子量无关。
空间位阻参数(σ):空间位阻参数是指由于高分子链的内旋转受阻而导致分子尺寸增大程度的量度,其值愈小,高分子链愈柔顺Flory 极限特征比(C∞ ):Flory 极限特征比是指高分子链由于键角限制和空间位阻造成分子链伸展的程度,。
一般来说,C∞ 愈小,链愈柔顺。
合成的柔顺性高聚物的C 值在5~7 范围内,而天然高分子多数高于此范围。
持续长度(q):持续长度q 广义为高分子链在第一个键方向上的投影,它表征分子链的支撑能力。
晶纤维名词解释-概述说明以及解释

晶纤维名词解释-概述说明以及解释1.引言1.1 概述晶纤维是一种具有晶体结构的纤维材料,其特点是具有高强度、高模量、低比重和优异的耐热性能。
它由纳米级晶体构成,这些晶体在纤维中具有有序排列的特点,使得晶纤维具有出色的力学性能和优异的热传导性能。
晶纤维的制备方法多样,可以通过溶剂法、湿法纺丝法、干法纺丝法等不同工艺得到。
晶纤维在各个领域都有广泛的应用。
在纺织领域,晶纤维可以用于制作高强度的纺织品,如防弹衣、护甲等。
此外,晶纤维还可以用于制作高性能的绳索和缆绳,被广泛应用于登山、航海等领域。
在航空航天领域,晶纤维则可以用于制作航空器的结构材料,因其轻便和高强度的特点而备受青睐。
同时,晶纤维的耐高温性能也使其成为航空发动机喷嘴等高温部件的理想材料。
可以预见,随着科技的不断进步和新材料的开发,晶纤维在未来将有更广阔的应用前景。
其出色的力学性能和耐热性能使其在新能源、新材料、生物医药等领域的应用成为可能。
与此同时,随着制备工艺的不断改进和成本的降低,晶纤维的商业化生产将更加普及,为各行各业带来更多的机遇和挑战。
综上所述,本文将对晶纤维的定义、制备方法以及应用领域进行详细的介绍和解释。
通过深入了解晶纤维的特点和优势,我们能够更好地掌握和应用这一材料,为各行业的发展和创新做出更大的贡献。
文章结构部分内容的编写如下:1.2 文章结构本文将按照以下结构展开论述晶纤维的相关内容:2.1 晶纤维定义:首先,我们将详细解释晶纤维的定义,并介绍其在纤维材料中的特殊性质和结构。
2.2 晶纤维制备方法:接下来,我们将探讨晶纤维的制备方法,包括熔融法、湿法和干法等不同的制备技术,并对各种方法的优缺点进行比较和分析。
2.3 晶纤维应用领域:然后,我们将介绍晶纤维在各个领域的广泛应用,包括纺织、建筑、医疗等方面,以及其在未来发展中的潜在应用领域。
3.结论3.1 对晶纤维的重要性进行总结:在本部分,我们将总结晶纤维在现代社会中的重要性,强调其在工业、科学研究和生活中的关键作用。
天然纤维素结晶变体I、结晶变体II简谈

天然纤维素结晶变体I、结晶变体II简谈(作业1)摘要:纤维素(cellulose)是由葡萄糖组成的大分子多糖。
不溶于水及一般有机溶剂。
是植物细胞壁的主要成分。
在自然。
具有一定构象的纤维素高分子链按一定的秩序堆砌,便成为纤维素的微晶体,微晶体的组成单元称为晶胞。
在纤维素中存在着化学组成相同,而单元晶胞不同的同质多晶体(结晶变体),常见的结晶变体有四种,即纤维素Ⅰ、Ⅱ、Ⅲ、Ⅳ。
本文将这种介绍纤维素Ⅰ、纤维素Ⅱ之间的转化。
关键字:纤维素结晶变体转化结构1、简述纤维素纤维素是由D-吡喃型葡萄糖基(失水葡萄糖)组成。
简单分子式为(C6H10O5)n;化学结构式可用下二式表示:霍沃思式是由许多D-葡萄糖基(1-5结环),藉1-4,β-型联结连接起来的,而且连接在环上碳原子两端的OH和H位置不相同,所以具有不同的性质。
式中n为聚合度。
在天然纤维素中,聚合度可达10000左右;再生纤维素的聚合度通常为200~800。
在一个样品中,各个高分子的聚合度可以不同,具有多分散性。
椅式由于内旋转作用,使分子中原子的几何排列不断发生变化,产生了各种内旋转异构体,称为分子链的构象。
纤维素高分子中,6位上的碳-氧键绕5和6位之间的碳-碳键旋转时,相对于5位上的碳-氧键和5位与4位之间的碳-氧键可以有三种不同的构象。
如以g表示旁式,t表示反式,则三种构象为gt、tg、和gg。
多数人认为,天然纤维素是gt构象,再生纤维素是tg构象。
在纤维素分子链中,存在着氢键。
这种氢键把链中的O6(6位上的氧)与O2'以及O3与O5'连接起来使整个高分子链成为带状,从而使它具有较高的刚性。
在砌入晶格以后, 一个高分子链的O6与相邻高分子的O3之间也能生成链间氢键。
2、纤维素结晶变体及其结构纤维素的聚集态结构是研究纤维素分子间的相互排列情况(晶区和非晶区、晶胞大小及形式、分子链在晶胞内的堆砌形式、微晶的大小)、取向结构(分子链和微晶的取向)等。
纤维素的大分子结构

纤维素的大分子结构纤维素是一种由β-葡萄糖单体组成的天然聚合物大分子。
它是地球上最常见的有机化合物之一,在植物细胞壁中起着关键的结构和功能作用。
纤维素的大分子结构决定了它的物理性质和化学性质,对于理解纤维素的特性和应用至关重要。
纤维素的大分子结构是由若干个β-葡萄糖单体通过β-1,4-连接键连接而成的线性聚合物。
β-葡萄糖单体有两个C1和C4碳原子,它们通过氧原子形成1,4-葡萄糖醚键。
这种键的特殊性决定了纤维素的特殊性质,如生物降解性和高强度。
纤维素的结构中的OH基团没有被化学修饰,因此纤维素是一种天然的、无毒的高分子化合物。
纤维素在自然界中主要存在于植物细胞壁中。
在植物细胞中,纤维素通常以微纤的形式存在,形成了复杂的网状结构。
纤维素的微纤具有一定的直径和长度,纤维素纤维在纳米尺度上呈平行排列,形成了纤维素纤维束和纤维。
纤维素的大分子结构非常有序,这种有序结构使纤维素具有很高的拉伸强度和模量。
纤维素纤维的强度和模量远远超过钢铁,因此纤维素具有很高的生物力学性能。
纤维素还具有超强的吸水能力,纤维素纤维能够吸收大量的水分,使其体积增大,并形成高度结晶的纤维素水胶体。
在纤维素纤维中,纤维素链之间通过氢键和范德华力相互作用。
这种相互作用使纤维素具有相对稳定的二级结构。
纤维素链通常以平行排列的方式组织在一起,形成纤维素纤维束和纤维。
纤维素的线性结构和氢键相互作用决定了纤维素的高度结晶性和热稳定性。
纤维素还具有很高的生物降解性和可再生性。
纤维素是植物细胞壁中的主要组分,它在自然界中被微生物和酶降解。
纤维素的降解产物是水和二氧化碳,没有任何有害的副产物。
这种生物降解性使纤维素成为一个非常重要的可再生材料,可以广泛应用于纺织、造纸、食品、医药等领域。
总结来说,纤维素的大分子结构是由若干个β-葡萄糖单体通过β-1,4-连接键连接而成的线性聚合物。
纤维素以微纤的形式存在于植物细胞壁中,并且形成了复杂的网状结构。
纤维素的有序结构使其具有很高的拉伸强度和模量,而其生物降解性和可再生性使其成为一个重要的可持续发展材料。
纳米纤维素的结晶度

纳米纤维素的结晶度1纳米纤维素的简介纳米纤维素是一种从植物中提取的纳米级纤维素,其直径约为10-90纳米。
与传统的微米级纤维素相比,纳米纤维素具有更高的比表面积和更好的可溶性,在材料科学、生物医学、环境保护等领域具有广泛的应用前景。
2纳米纤维素的结晶度概念纳米纤维素的结晶度是指其纤维素晶体的数量和排列程度。
纤维素是构成植物细胞壁的一种主要成分,其晶体结构由许多葡萄糖分子构成。
随着纤维素晶体的增长,其晶体结构会逐渐排列成纤维素纤维,形成杆状结构。
3结晶度的影响因素纳米纤维素的结晶度受到多种因素的影响,包括原料来源、制备方法、溶液浓度、加热温度、离子强度等。
其中,原料来源和制备方法是影响纤维素晶体结构的主要因素。
不同的原料来源和制备方法会产生不同的晶体形态和结晶度,从而影响纳米纤维素的性质和应用。
4结晶度的测定方法目前,常用的纳米纤维素结晶度测定方法主要包括X射线衍射、红外光谱、偏振显微镜和热分析等。
其中,X射线衍射是最常用的方法之一。
X射线衍射可以通过测量衍射峰的位置和强度来确定纤维素晶体的结构和结晶度,但需要很高的仪器精度和样品制备技术。
5结晶度对纳米纤维素性质的影响结晶度是影响纳米纤维素结构和性质的重要参数之一。
纤维素晶体的结晶度越高,纳米纤维素的机械强度、热稳定性和抗水解性等性质就越好。
同时,高结晶度的纳米纤维素还具有更好的光学性能和可逆吸附性能,可用于光学材料和环境治理等领域。
6结晶度的调控随着纳米纤维素在各个领域中的应用越来越广泛,对其结晶度的调控也越来越重要。
目前,影响纳米纤维素结晶度的方法主要包括化学处理、物理处理和结构设计等。
化学处理是通过改变溶液成分和pH 值等来调控纤维素晶体结构和结晶度;物理处理是通过热处理、机械剪切等方式来促进晶体生长和调控结晶度;结构设计是通过构建有序和多级结构来调控纤维素纤维的结晶度和组织结构。
7结论纳米纤维素的结晶度是影响其性质和应用的重要参数之一。
随着技术的不断改进和深入研究,我们相信纳米纤维素的结晶度调控会带来更多的技术创新和应用突破。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
精品课件
纤维素 II的晶胞特征
① 属于单斜晶系,晶胞参数平均值: a=7.93Å,b=9.18Å , c=10.34Å ,γ=117.31º
② 相邻分子链是反向平行。 ③ 角链上的伯羟基为gt位,链的方向向上,中心链上的伯羟
基为tg位,链的方向向下。 ④ 中心链相对于角链在纤维轴c的方向上相互错开0.216c。 ⑤ 纤维素 II中形成的氢键网较纤维素 I中复杂。前者比后者
X-射线衍射法测定纤维素相对结晶度案例-1
➢ 球磨时间增加,结晶度下降
X-射线衍射法测定纤维素相对结晶度案例-1
➢ 球磨时间增加,结晶度下降
X-射线衍射法测定纤维素相对结晶度案例-2
➢水对纤维素大分子排列影响较大
① 木粉沸水煮2h,结晶度 明显提高。这意味着水 分子使无定形区大分子 链定向排列,重新产生 新的结晶区。
四、劳埃方程
由于晶体中原子呈周期性排列,劳埃设想 晶体为光栅(点阵常数为光栅常数),晶 体中原子受X射线照射产生球面散射波并 在一定方向上相互干涉,形成衍射光束。
1. 一维劳埃方程
任意方向上原子散射线单位矢量s
:s与a之夹角
点阵基矢(原子间距)a
入射线单位矢量s0
0:s0与a之夹角
一维劳埃方程的导出
精品课件
晶面和晶面指数
精品课件
纤维素单元细胞的结晶变体
固态下的纤维素存在5种结晶变体,即天然纤维素I、人造
纤维素II、纤维素III、纤维素IV和纤维素X。5种结晶变体
各有不同的晶胞结构,晶胞参数如下:
a
b
c
β
纤维素I
8.35Å
10.3Å
7.9Å
84º
纤维素II
8.10Å
10.3Å
9.1Å
62º
原子列中任意两相邻原子(A与B)散射线间光程差()为: =AM-BN=acos-acos0
散射线干涉一致加强的条件为=H,即:
a(cos-cos0)=H
H——任意整数
a(cos-cos0)=H表达了单一原子列衍射线方向()与入 射线波长()及方向(0)和点阵常数的相互关系,称为
一维劳埃方程。
亦可写为 :
cosβ0)=kλ
c(cosγ - cosγ0) = lλ ➢ 布拉格方程 2dsinθ= nλ ➢ Scherrer公式 D=K· λ /(Bhkl · cosθ)
背景
1912年劳埃(M. Van. Laue)用X射线照射五水硫酸铜( CuSO4·5H2O)获得世界上第一张X射线衍射照片,并由光 的干涉条件出发导出描述衍射线空间方位与晶体结构关 系的公式(称劳埃方程)。
(1)纤维素大分子链不在一个微胞内终 止,而是贯穿了一个以上的微胞(晶区) 和微胞间物质(非晶区),分子长度与 微胞长度无一定关系,晶区和非晶区无 明显的界面。 (2)纤维素大分子从晶区逸出以缨状的形 式进入非晶区,由高度结晶的有序区至 完全无结晶的无序区是连续过渡的。
精品课件
2.缨状基微纤维理论(fringed-fibril theory)
堆砌较为紧密,在热力学上更稳定。
精品课件
纤维素结晶区与非结晶区的关系
纤维素纤维中纤维素的聚集态是很复杂的。 一相结构理论没有得到公认。 现在较普遍承认的是两相结构理论,即纤维素纤维中的 纤维素是以结晶相和无定形相共存的。
精品课件
纤维素结晶区与非结晶区的关系 1.缨状微胞理论
长链分子间的规整排列构成结晶微胞。而伸出的无 规则排列的分子成为缨状须从。故称为缨状微胞理 论(fringed micelle theory)。
如:以Cu K射线照射
NaCl表面,当=15和 =32时记录到反射线。
设入射线与反射面之夹角为,称掠射角或布拉格角,则按反射定律, 反射线与反射面之夹角也应为。
2.布拉格方程的导出
考虑到: ①晶体结构的周期性,可将晶体视为由许多相互平行且晶
面间距(d)相等的原子面组成;
②X射线具有穿透性,可照射到晶体的各个原子面上;
0、0及0——s0与a、b及c的夹角
或
、及——s与a、b及c的夹角
a·(s-s0)=H b·(s-s0)=K c·(s-s0)=L
三维晶体若要产生衍射,必须同时满足上述三个方程
Scherrer公式
Scherrer公式用于样品晶粒尺寸的计算 D=K· λ /(Bhkl · cosθ) 其中,D为沿垂直于晶面(hkl)方向的晶粒直径,
任选两相邻面(A1与A2),反射线光程差=ML+LN=2dsin ;干涉一致加强的条件为=n,即 2dsin=n 式中:n——任意整数,称反射级数,d为(hkl)晶面间 距,即dhkl。
衍射产生的必要条件: “选择反射”即反射定律+布拉格方程。
即当满足此条件时有可能产生衍射;若不满 足此条件,则不可能产生衍射。
a·(s-s0)=H
2. 二维劳埃方程
a(cos-cos0)=H b(cos-cos0)=K
或
0及0——s0与a及b的夹角 及——s与a及b的夹角
a·(s-s0)=H
b·(s-s0)=K
单一原子平面受X射线照射必须同时满足两个方程,才 可能产生衍射。
3. 三维劳埃方程
a(cos-cos0)=H b(cos-cos0)=K c(cos-cos0)=L
片晶的分子链是垂直于薄片的平面的。
一个片晶到相邻的片晶之间有很多起联接作
用的大分子贯穿着。
精品课件
纤维素结晶结构的研究方法
X-射线衍射法 红外光谱法 交叉极化魔角旋转核磁共振法 拉曼光谱
精品课件
X-射线衍射法
X-射线的产生与X射线
当高速运动的电子撞击到一个金属靶上时,靶面上被电 子撞击的部位就产生电磁波辐射,其中一部分为X射线。在 管电压没超过某一数值Vk(激发电压)时,只有连续射线 谱产生。超过Vk时,若干强度很高的特征谱线叠加在连续 谱线上,为特征X射线谱。
CHINESE ACADEMY OF FORESTRY
纤维素结晶结构
组员:郭 飞 张毛毛 温留来 张龙飞 周贤武
2012-12-04
精品课件
1
3.纤维素的物理结构
3.1 纤维素的结晶结构 晶体的基本概念
晶体:物质内部质点(原子、分子、离子)呈规律排列的固体, 为“空间格子”状。
晶胞:晶体的最小重复单位。可以用3个晶轴的长度a、b、c 及其夹角α、β、γ6个晶胞参数来描述。
相 对 强 度
X射线管结构
X-射线衍射法
X-射线衍射的基本原理
晶体是由原子或原子基团等按照一定规律在空间内有 规则排列而构成的固体。当它被X射线照射后,各个原 子散射X射线。这些散射线符合相干波的条件,因而产 生干涉现象。所谓X射线衍射,实质上就是研究这些散 射波的干涉。衍射线就是经过相互干涉而加强的大量散 射线所组成的射线。
k为Scherrer常数(通常为0.89), λ为入射X 射线波长,θ为布拉格衍射角, Bhkl为衍射峰的 半高峰宽(rad)。
纤维素相对结晶度
纤维素的结晶度是指纤维素构成的结晶区占纤维素整体的 百分数,它反映纤维素聚集时形成结晶的程度。 CrI=结晶区样品含量/(结晶区样品含量+非结晶区样 品含量)
精品课件
Blackwell模型
① 晶胞参数: a=16.34Å ,b=15.72Å ,c=10.38Å ,β=97.0º
② 纤维素分子链占据结晶单元的 4个角和中轴。(这种晶胞含8 条分子链横截面)
③ 中间链和位于角上的链是沿同 一方向的平行链,中间链在高 度上与位于角上的链半个葡萄 糖基。
④ 链分子的薄片平行于ac面,所 有的伯羟基均为tg构象(见第 38页)。
随后,布拉格父子(W.H.Bragg与W.L.Bragg)类比 可见光镜面反射安排实验,用X射线照射岩盐(NaCl), 并依据实验结果导出布拉格方程。
一、布拉格方程
1.布拉格实验
散射角2 :入射线方向与散 射线方向之间的夹角。
选择反射:当X射线以某 些角度入射时,记录到 反射线,其它角度入射, 则无反射。
精品课件
3.折叠链结晶结构理论 (fringed-micelle with chain 纤维素大分子有呈折叠链fo状ld的in可g能),这些折
叠链状的线型纤维素大分子仍然能形成结晶
结构,即所谓折叠链结晶,或所谓片晶。人
们认为片晶就如同缨状微胞结构中的微胞;
伸出的分子就像缨状分子。
片的厚度即相当于沿纤维轴向出现的100 ~200 Å的自同周期的大小(X射线衍射发现 纤维在纵向上存在周期)。
精品课件
七个晶系及其晶胞参数
精品课件
晶面和晶面指数
晶面:结晶格子内所有的格子点全部集中在相互平行的等间距 的平面群上,这些平面叫做晶面,晶面的间距为d。用
晶 面指数(Miller指数)来标记。(通过晶体中原子中心
的平面叫作晶面) 三个晶轴截距的倒数,通分,分子作为晶面指数。
如截距分别为2a、3b、 c,倒数为1/2、1/3、1/1,通分为 3/6、2/6、6/6,则(326)就是晶面指数。
•I002 —(002)面的衍射强度 纤维素II结晶度计算
CrI%=(I101-Iam)/ I101 × 100 Iam — 2θ为15º附近无定形区散 射强度
•I101—(101)面的衍射强度
Turley法
相对结晶度衍射示意图(王欣 香根草构造与基本特性研究)
将样品木粉在室温下 压成薄片,然后做成 2θ强度曲线,样品扫 描范围为3°~ 40°( 2θ)角, 在扫 描曲线上2θ = 22° 附近(002)衍射的极 大峰值( I002 ) , 2θ= 18°附近有一 极小值( Iam ) 。
X-射线衍射法