4.多细胞动物的起源
第4章_多细胞动物的起源0
两侧对称动物:其他所有门类
后生动物在胚胎发育过程中有胚层的分化, 多孔动物门只有内胚层和外胚层的初步分化, 腔肠动物门在内外胚层间又有中胶层。 自扁形动物门以后的门类都是三胚层动物。
根据体腔的有无和结构可将后生动物分为 无体腔动物:包括多孔动物门、腔肠动物门和 扁形动物门; 假体腔动物:包括线形动物门、腹毛动物门等; 体腔动物:包括环节动物以后的所有动物门类。
4.1 从单细胞动物到多细胞动物
单细胞动 物(原生 动物)
中生动物
多细胞动 物(后生 动物)
侧生动物 真后生动物
4.1 从单细胞动物到多细胞动物
进化顺序
原生动物→中生动物→后生动物
中生动物(双胚虫、异胚虫)
小型内寄生动物 结构简单 约50种 细胞数目恒定 其分类位置尚无定论
后生动物是除原生动物外所有其他动物的 总称(后生动物亚界)。动物界除原生 动物门以外的所有多细胞动物门类的总 称。
真后生动物
动物界除原生动物门以外的所有多细胞 动物门类的总称。 其特征是体躯由大量形态有分化、机能有分 工的细胞构成; 与群体原生动物的兼有营养和生殖功能的细 胞不同,其生殖细胞和营养细胞有明显的分 化。
真后生动物
依体制形态的对称情况,后生动物: 不对称动物:多孔动物门
辐射对称动物:腔肠动物门、栉水母动物门、 棘皮动物门
4.2 多细胞动物起源于单细胞动物的证据
一、古生物学 地层中动物化石
如:最古老的地层化石最简单,晚近的地层动物化 石种类多且复杂。
二、形态学 单细胞→多细胞,简单→复杂,低等→高等
如:原生动物的团藻等群体形态与多细胞动物相 似,可能为中间类型,以群体的形式过渡。
三、胚胎学 多细胞动物从受精卵开始,经卵裂、囊胚、原肠
3 多细胞动物的起源-讲课

外胚层:分化为皮肤上皮(包括皮肤腺和其他皮肤 衍生物)、神经组织、感觉器官和消化管的两端。
三. 生物发生律
德国学者赫克尔(Haeckel, E. 1866)
生物发生律(biogenetic law ): 生物发展史分为个体发育和系统发育,即个体的发 育历史和由同一起源所产生生物群的发展历史。 个体发育史是系统发育史的简单而迅速的重演。 即生物在个体发育过程中,按顺序重演其祖先的主 要发育阶段。
不同地层的化石特点: 早期地层:古生物化石越简单,越低等, 晚期地层:古生物化石越复杂,越高等。 最明显、最主要的证据是化石。
最早的化石:澳州西海岸35亿年前的兰细菌化石
(二) 形态学证据(Morphological evidence)
现存的动物种类形成了一个由简单到复杂、由低等 到高等的序列。
辐射卵裂:棘皮、两栖类
第3次卵裂以后,上层的分裂球很整齐地排列在下 层之上,呈辐射排列。
不完全卵裂:多见于多黄卵,卵黄多,细胞分裂 受阻,卵裂只在不含卵黄的部位进行。
盘裂:分裂局限于胚盘处,如乌贼、鸡. 囊胚的形成(blastulation)
5. 中胚层及体腔的形成
中胚层的形成方式: 裂体腔法(端细胞法):原口动物、高等脊索;
体腔囊法(肠体腔法):棘皮动物、毛颚动物、半 索动物、原索动物。
体腔的类型:三胚层无体腔、假体腔和真体腔
6. 胚层分化
内胚层最简单,中胚层最复杂,外胚层最特异。
内胚层:分化为消化管的大部分上皮,肝、胰、呼 吸器官,排泄器官和生殖器官的一部分; 中胚层:分化为肌肉,结缔组织、生殖和排泄器官 的大部分;
4. 原肠胚的形成(Gastrulation)
第3、4章 多细胞动物的起源多孔动物门

第三章 多细胞动物的起源
从单细胞到多细胞
多细胞动物起源于单细胞动物的证据
关于多细胞动物起源的学说 胚胎发育的重要阶段 生物发生律
第四章 多孔动物门(Porifera)
多孔动物门的主要特征
分类及分类地位 经济意义
从单细胞到多细胞?
人的胚胎发育过程
动物的发育
个体发育 系统发育
• 一.个体发育:有机体从受精卵 发育成成体的过程。包括 胚前期, 胚胎期和胚后期。 (一)胚前发育:性细胞的产生和成熟的过程。
精母细胞 4个精子 3个极体
卵母细胞
1个卵细胞
少黄卵:卵黄少,均匀
中黄卵:卵黄多,分布于中央 多黄卵:卵黄多,分布于一侧
(二)胚胎发育
受精(fertilization)与受精卵 卵裂(cleavage) 囊胚的形成(blastulation) 原肠胚的形成(gastrulation) 中胚层及体腔的形成
活习性方面无区别,如低等昆虫。 变态发育(间接发育):幼体和成体在形态、生活 习性方面的区别显著,如蛙、昆虫等。
二、系统发育
个体发育和系统发育
个体发育(ontogeny): 是指生物个体从受精卵开始到个体性成熟 或死亡为止的整个过程。在个体发育过程中,个体的生理功能、
组织结构和器官形态都发生一系列变化。
生物发生律(biogenetic law, 重演律recapitulation law):
由德国科学家赫克尔(E.Haeckel)于1866年提出。是指个体发 育史是系统发展史的简单而迅速的重演。二者的对应关系如 下:
青 蛙
个 体 发 育
受 精 卵
囊 胚
多细胞生物的起源

多细胞动物的起源姓名:王园学号:20117319专业:生物科学学院:农生院时间:2012.3.3随着科学的进步和人类的进一步探索,越来越多的例子和研究证明多细胞动物起源于单细胞动物。
在未来的日子里,揭开多细胞的起源将不会成为难题。
如:“多细胞生物起源或可追溯至寒武纪前”的发表证明人类的研究已经取得一定的成就,谜底解开之日将不会远。
一、多细胞动物起源于单细胞动物动物由单细胞演变为多细胞是动物发展史的一个重要阶段。
一切高等生物,包括动物、植物,都是多细胞的。
但多细胞动物的进化发展远较植物的快。
这是因为多细胞动物在进化过程中发展了两侧对称的体型,进而身体各部分明显分工,出现了头部,使得神经、感官等大大发展,而这些发展都是由于多细胞动物长期适应于活跃的、主动的生活方式而形成的。
原生动物——单细胞动物虽然也能完成起生命的各种活动,并有些单细胞动物结构上有一定程度的复杂化,但由各种细胞器来完成的各种不同的功能,这仅仅是一个细胞内的分化。
原生动物也有一些多细胞群体,它们只是以群体的方式存在,一般仍是以一个个细胞为独立的生活单位的,彼此之间并不发生密切联系。
多细胞动物我们称之为Metozoa,即后生动物,这是相对于原生动物(prot-ozoa)而言的。
后生动物包括除原生动物以外的绝大多数的多细胞动物。
也有学者认为在原生动物和后生动物之间存在着一个小类群--中生动物Mesozoa,这类动物寄生在海洋无脊椎动物的体内,个体细胞数目20-30个。
二、由单细胞动物发展到多细胞动物的证据现在公认多细胞动物起源与单细胞动物,证据主要如下三个方面:1、古生物学方面从不同地层中的化石种类来判断。
化石---古代动、植物的遗体或遗迹。
化石研究发现,越是古老的地层,化石种类越简单。
在太古代(地质史最古老的年代),距今32亿到距今18亿年的中生代的地层中有大量有孔虫的化石。
而晚近地层中的化石种类则较复杂,并且动物杂交的程度是一个渐进的变化过程。
普通动物学—多细胞动物的起源和多孔动物(上课版)

注解: G:胚囊小细胞向 囊腔生出鞭毛。 H、I:大细胞一端 形成一个大孔。并 向外包,里面的变 成外面。 J、K:两囊幼虫 L:小细胞内陷 M:固着生长
(三)海绵动物的分类 ①钙质海绵纲:钙质骨针,简单的水沟系,较小的体形,多浅海生 活,如白枝海绵和毛壶等。 ②六放海绵纲:六放形硅质骨针,复沟型,鞭毛室大,体形较大, 深海生活,如偕老同穴,拂子介等。 ③ 寻常海绵纲:硅质骨针(非六放)或海绵质纤维,复沟型,鞭毛室 小,不规则体型,海产或淡水产,如浴海绵,针海绵等。
(二)多孔动物的生殖和发育 ①无性生殖分出芽和形成芽球。 芽球由中胶层中的若 干原细胞聚成堆,外 包几丁质膜或骨针形 成。 所有的淡水海绵和部 分海产种类能形成芽 球。
②有性生殖:雌雄同体或异体,异体受精,精子和卵是由原细胞或 领细胞发育,受精过程很特殊,领细胞将精子带入卵进行受精。胚 胎发育中有特殊的胚层逆转现象。 幼虫:两囊幼虫。
原口动物
后口动物
扁形动物(涡虫、吸虫、绦虫)
线形动物(线虫、轮虫…) 环节动物(沙蚕、蚯蚓、蛭)
棘皮动物门(海星)
半索动物门(柱头虫) 脊索动物门 尾索动物(海鞘) 头索动物(文昌鱼) 脊椎动物(鱼类、两栖动物 、爬行动物、鸟类、哺乳动 物)
软体动物(螺类、贝类…)
节肢动物(甲壳类、蜘蛛、昆 虫…)
红毛壶
偕老同穴
拂子介
浴海绵
1、海绵动物也称侧生动物,其处于系统发生侧枝的原因 A无神经系统 B 无明显组织分化 C具有两囊幼虫 D 胚胎发育中具有胚层逆转 2、从下列陈述找出对海绵动物的不正确的叙述是 A它们的骨骼由钙质、二氧化硅或海绵质的针 B它们用吞噬方式获取食物 C所有海绵动物生活在海洋中 D海绵动物能与许多共生动物共生 3、海绵动物特有的结构 A、水沟系 B、 固着生活 C、领细胞 D、骨针
普通动物学 第四章 多细胞动物的起源

二.胚胎发育
生物发生律(重演律) • 个体发育史是系统发展史的简单而迅速的重演。
冯·贝尔法则 • 所有脊椎动物具有的共同结构总是优先发生,而不同类别动物 的特征结构则后发生。
胚胎发育的重要阶段
• 受精 → 卵裂 → 囊胚 → 原肠胚 → 中胚层及体腔形成(无脊椎动物) 神经胚(脊椎动物)
1.受精与受精卵
卵的类型(据卵黄多少分):少黄卵 中黄卵 多黄卵
卵细胞:动物极(卵黄相对较少的一端) 植物极(卵黄相对较多的一端)
2.卵裂
每次分裂后,新细胞未长大,又继续分裂,因此分裂成的细 胞越来越小。形成的新细胞称分裂球。
• 卵黄使分裂受阻。 • 植物极卵黄相对多,细胞分裂较少,细胞较大。
卵裂方式
卵的类型与卵裂
辐射卵裂
螺旋卵裂
3.囊胚形成
囊胚:分裂球形成的中空的球状胚。 囊胚腔:囊胚中间的腔。 囊胚层:囊胚壁的细胞层。
4.原肠胚形成
形成原肠腔,分化出内、外胚层。 方式:
内陷:植物极细胞向内陷入 形成胚孔(原口)
内移、外包、分层、内转(盘状囊胚)
• 内陷与外包、分层与内移常综合出现。 • 植物极形成内胚层,动物极形成外胚层。
5.中胚层和体腔形成
胚层(germ layer): 胚胎期具有共同来源、 相同形态、相同演化 方向的细胞层。
真体腔:中胚层之间的腔(从环节动物开始有真体腔)。 • 端细胞法(裂体腔法)
• 除触手冠动物以外的原口动物、高等脊椎动物。 • 体腔囊法(肠体腔法)
• 后口动物的棘皮、毛颚、半索、低等脊索动物。
第4章 多细胞动物的起源
一.从单细胞到多细胞
单细胞动物:原生动物。 多细胞动物:中生动物、侧生动物、后生动物。
多细胞动物起源于单细胞动物的证据

一、论点:
一般公认多细胞动物起源于单细胞动物。
二、证据:
1. 古生物学方面: 2. 形态学方面: 3. 胚胎学方面:
(一) 古生物学证据
古代动、植物的遗体或遗迹,经过千百万年地壳的变迁 或造山运动等,被埋在地层中形成了化石. 已经发现在最 古老的地层中化石种类是最简单的. 在太古代的地层中有 大量有孔虫壳化石,而在晚近的地层中动物的化石种类 也较复杂,并且能看出生物由低等向高等发展的顺序. 这 说明最初出现单细胞动物,后来才发展出多细胞动物.
5. 外包:动物极细胞分裂快,植物极细胞分裂慢,结果
动物极细胞逐渐向下包围植物极细胞,形成外胚层, 植物极细胞为内胚层。
原肠胚的这几种分裂形成方式常常综合出现,最常见的是内陷与外 包同时出现,分层与内移相伴而行。
原肠胚的形成
囊胚进一步 发育形成原 肠胚,此时 胚胎分化出 内、外两胚 层和原肠腔.
(三) 胚胎学证据
在胚胎发育中多细胞动物是由受精卵开始,经过卵裂、 囊胚、原肠胚等一系列过程,逐渐发育成成体.多细胞 动物的早期胚胎发育基本上是相似的.
胚胎发育的重要阶段
一、受精与受精卵
雄性个体产生雄性生殖细胞--精子 雌性个体产生雌性生殖细胞-- 卵
受精卵
少黄卵 中黄卵 多黄卵 卵黄多的一端称为植物极,另一端称为动物极.
五、中胚层及体腔的形成
绝大多数多细胞动物在形成内、外胚层后还进一步 发育形成中胚层,在中胚层间形成的腔称为真体腔。
形成方式:
1. 端细胞法:在卵裂形成囊胚过程中,植物极的一个细 胞--端细胞分裂成两个原始的中胚层细胞,以后移入 内外胚层间对称排列。这两个细胞分裂成很多细胞,形 成索状,伸入内外胚层间形成中胚层;中胚层之间裂开 形成真体腔。(如原口动物)
多细胞生物的起源
多细胞动物的起源姓名:王园学号:20117319专业:生物科学学院:农生院时间:2012.3.3随着科学的进步和人类的进一步探索,越来越多的例子和研究证明多细胞动物起源于单细胞动物。
在未来的日子里,揭开多细胞的起源将不会成为难题。
如:“多细胞生物起源或可追溯至寒武纪前”的发表证明人类的研究已经取得一定的成就,谜底解开之日将不会远。
一、多细胞动物起源于单细胞动物动物由单细胞演变为多细胞是动物发展史的一个重要阶段。
一切高等生物,包括动物、植物,都是多细胞的。
但多细胞动物的进化发展远较植物的快。
这是因为多细胞动物在进化过程中发展了两侧对称的体型,进而身体各部分明显分工,出现了头部,使得神经、感官等大大发展,而这些发展都是由于多细胞动物长期适应于活跃的、主动的生活方式而形成的。
原生动物——单细胞动物虽然也能完成起生命的各种活动,并有些单细胞动物结构上有一定程度的复杂化,但由各种细胞器来完成的各种不同的功能,这仅仅是一个细胞内的分化。
原生动物也有一些多细胞群体,它们只是以群体的方式存在,一般仍是以一个个细胞为独立的生活单位的,彼此之间并不发生密切联系。
多细胞动物我们称之为Metozoa,即后生动物,这是相对于原生动物(prot-ozoa)而言的。
后生动物包括除原生动物以外的绝大多数的多细胞动物。
也有学者认为在原生动物和后生动物之间存在着一个小类群--中生动物Mesozoa,这类动物寄生在海洋无脊椎动物的体内,个体细胞数目20-30个。
二、由单细胞动物发展到多细胞动物的证据现在公认多细胞动物起源与单细胞动物,证据主要如下三个方面:1、古生物学方面从不同地层中的化石种类来判断。
化石---古代动、植物的遗体或遗迹。
化石研究发现,越是古老的地层,化石种类越简单。
在太古代(地质史最古老的年代),距今32亿到距今18亿年的中生代的地层中有大量有孔虫的化石。
而晚近地层中的化石种类则较复杂,并且动物杂交的程度是一个渐进的变化过程。
普通动物学练习题及参考答案

普通动物学练习题及参考答案学习要点:1. 各动物门的主要特征2. 各动物门主要的纲及其重要代表动物。
3. 最早出现某种器官或组织结构的动物类群。
4. 回答简答题和论述题要有必要的连接语言,使答案显得通顺流畅,前后连贯,有头有尾。
第1章绪论一、填空题1. 现在所用的动物分类系统是以动物形态或解剖的和的总和为基础,根据古生物学、比较胚胎学和比较解剖学上的许多证据建立起来的,基本上能反映动物界的自然亲缘关系,称为。
2. 动物分类系统由大而小有、、、、、、等几个重要的分类阶元(分类等级, category),任何一个已知的动物均可无例外地归属于这几个阶元之中。
在上述分类阶元中,只有是客观存在的,其它较高的阶元都同时具有客观性和主观性。
3. 物种是生物界发展的连续性与间断性统一的基本间断形式;在有性生物,物种呈现为统一的,由占有一定空间、具有实际或潜在的种群所组成,而与其它物种这样的群体在生殖上是的。
4. 国际上目前统一采用的命名法是。
二、名词概念1. 物种;2. 双名法;第2章动物体的基本结构与机能一、填空题1. 是生物体结构与机能的基本单位。
2. 细胞是一团,由它分化出、、和等。
3. 细胞分裂可分为、和等三种类型。
4. 细胞由一次分裂结束到下一次分裂结束之间的期限称为,它包括和。
二、名词概念1. 组织;2. 器官;3. 系统。
第3章原生动物门一、填空题1. 原生动物是动物界里最原始、最低等的动物,身体由细胞构成,因此也称为。
2. 眼虫是原生动物门(填写纲的名称)的动物,大变形虫是(填写纲的名称)的动物,间日疟原虫属于原生动物门(填写纲的名称),草履虫属于原生动物门(填写纲的名称)。
3. 绿眼虫的运动器官是,大变形虫的运动器官是。
草履虫的运动器官是。
4. 眼虫在运动中具有趋光性,眼虫与趋光性调节有关的结构是和。
5. 眼虫在有光的条件下通过叶绿素利用光能进行光合作用,把二氧化碳和水合成糖类,这种营养方式称为。
02 多细胞动物的起源

一、 多细胞动物起源于单细胞动物
动物由单细胞演变为多细胞是动物发展史的一个重要阶段
一切高等生物包括动物、植物,都是多细胞的。但多细胞动物的 发展远较植物的快。这是因为多细胞动物在进化过程中发展了两侧对 称的体型,进而身体各部分明显分工,出现了头部,使得神经、感官 等大大发展,而这些发展都是由于多细胞动物长期适应于活跃的、主 动的生活方式而形成的。 原生动物------单细胞动物虽然也能完成生命的各种活动,有些单细 胞动物结构上有一定程度上的复杂化,但由各种细胞器来完成的各种 不同的功能,这仅仅是一个细胞内的分化。原生动物也有一些多细胞 群体,它们只是以群体的方式存在,一般仍是以一个个细胞为独立的 生活单位的,彼此之间并不发生密切联系。
囊胚腔的出现使胚体细胞的活动有了充分的空间。
3、囊胚:分中空有腔囊胚(海绵动物、蛙、蚯蚓)和无腔 的实囊胚(水螅)。
海星和蛙的囊胚
4、原肠胚:囊胚进一步发育进入原肠胚阶段, 此时,胚胎开 始分化出两个胚层即内、外胚层, 凹陷形成的腔叫原肠腔, 其与外界相通的开口叫原口(胚孔),
形成的方法有内陷法(纽虫)、移入法(白枝海绵)、分 层法(水螅)、内转法(乌贼)、外包法(某些软体动 物)。方法常综合应用。 外胚层 内胚层
拟配子和胚胎
直游虫的多核变形体
中生动物的系统发育
存在争议: 1)退化的扁形动物:寄生,生活史复杂,结构简单, 与扁形动物有相似之处。
中生动物的系统发育
存在争议:
2)早期后生动物的一个分支:有体细胞和生殖细胞
的分化,体表具纤毛,寄生历史长,是由最原始的 多细胞动物进化而来。
中生动物的系统发育
存在争议: 3)真正原始的多细胞动物:DNA成分的含量与原生 动物纤毛虫类的含量接近,而低于其它多细胞动物 包括扁形动物。
多细胞动物起源的学说

(三) 共生学说
认为不同种的原生生物共生在一起,最终发展成为多细 胞动物. 这一学说存在一系列的遗传学问题,因为不同 遗传基础的单细胞生物如何聚在一起形成能繁殖的多细 胞动物,这在遗传学上是难以解释的.
多细胞动物起源的学说
(一) 群体学说
1.赫克尔的原肠虫学说
其认为多细胞动物最早的 祖先是由类似团藻的球形 群体,一面内陷形成多细 胞动物的祖先. 这样的祖 先,因和原肠胚很相似, 有两胚层和原口,所以赫 克尔称之为原肠虫.
2.梅契尼柯夫的吞噬虫学说(实球 虫或无腔胚虫学说)
他认为多细胞动物的祖先 是由一层细胞构成的单细 胞动物的群体,后来个别 细胞摄取食物后进入群体 之内形成内胚层,结果就 形成为二胚层的动物,起 初为实心的, 后来才逐渐 地形成消化腔,所以梅契 尼柯夫便把这种假想的多 细胞动物的祖先叫做吞噬 虫.
4.Otto Butshli(1883)扁 囊胚虫学说
这一学说认为原始的后生动 物是两侧对称的有两胚层的 扁的动物,,扁囊胚虫通过 腹面细胞层的蠕动、爬行、 摄食,最后该动物背腹细胞 层分开成为中空的,这样逐 渐地腹面的营养细胞内陷形 成消化腔. 同时产生了内外 胚层,形成了两胚层动物. 有些学者认为丝盘虫是扁囊 胚虫现存种类的证据.
(二) 合胞体学说
这一学说主要由Hadzi(1953)和Harsan(1977)提出,认 为多细胞动物来源于多核纤毛虫的原始类群.后生动物的祖 先开始是合胞体结构,即多核的细胞,后来每个核获得一部 分细胞质和细胞膜形成了多细胞结构. 由于有些纤毛虫倾向 于两侧对称,所以合胞体学说主张后生动物的祖先是两侧对 称的,并由其发展为无肠类扁虫,认为无肠类扁虫是现在生 存的最原始的后生动物. 对该学说,持反对意见者较多,因 为任何动物类群的胚胎发育都未出现过多核体分化成多细胞 的现象,实际上无肠类合胞体是在典型的胚胎细胞分裂之后 出现的次生现象. 最主要的反对意见是不同意将无肠类扁虫 视为最原始的后生动物. 体型的进化是从辐射对称到两侧对 称,如果认为无肠类扁虫两侧对称是原始的,那么腔肠动物 的辐射对称倒成为次生的,这显然与已揭明的进化过程是相 违背的.
动物学判断题

原生动物门1.原生动物的单个细胞与高等动物体上一个细胞的不同在于它能完成一个动物体所能完成的各种生理机能。
2.绿眼虫只能进行光合营养。
()3.草履虫在有光情况下进行光合营养,无光情况下进行吞噬营养。
()4.鞭毛和纤毛结构相似,主要是由许多纵行微管构成。
()5.大变形虫的伪足既是运动细胞器又是摄食的细胞器。
()6.形成包囊是原生动物度过不良环境的一种方式。
()7.草履虫的呼吸和排泄都是通过体表进行的。
()8.草履虫体内营养物质的运输是靠细胞质的流动。
()9.接合生殖是纤毛虫特有的一种有性生殖方式。
()多细胞动物的起源1.卵的类型划分主要根据卵内卵黄的多少及分布情况。
()2.世代交替具有重要的生物学意义,无性繁殖可以保证后代的数量,有性生殖可以保证后代的质量。
()3.动物的胚胎发育阶段都是指从受精卵开始到冲破卵壳出来的整个过程。
(4.各种动物在胚胎发育过程中,形成原肠胚时,都具有5种方式。
()5.鸡卵发育过程中,形成原肠胚时主要是以内转法。
(海绵动物门1.海绵动物的体壁由皮层、中胶层、胃层三层细胞构成。
(2. 海绵动物发育过程中存在胚层逆转现象是人们将其从进化主支上排出去的主要原因。
(3.在海绵动物中,水沟系具有单沟系、双沟系和三沟系三种类型。
(4.海绵动物的胃层能分泌消化酶,能在中央腔内进行细胞外消化。
(5.海绵动物的水沟系统是对固着生活的良好适应。
()6.海绵动物的体型都是辐射对称的。
()腔肠动物门1.腔肠动物的体壁,是由外胚层、中胶层、内胚层三层细胞构成的。
()2.水螅在胚胎发育过程中经过浮浪幼虫阶段。
()3.水螅通过体壁与外界进行气体交换。
()4.我们常见的珊瑚就是珊瑚虫。
()5.水螅身体中央的空腔既有消化作用又具循环作用。
()6.海葵的生殖腺在隔膜上,故起源于中胚层。
()7海蜇能哲人,原因是其身体上有刺细胞,能放出毒液。
()8.腔肠动物在动物进化过程中占有重要地位,是真正后生动物的开始。
()扁形动物门1.扁形动物是最原始的三胚层动物。
多细胞动物的起源(1)

2021/4/4
13
2、体腔囊法(肠体腔法):
原肠背部两侧,内胚层向外产生成对的囊状突起,称体腔囊。 体腔囊脱离内胚层在内外胚层之间扩展成中胚层,其内腔即为 体腔。由于体腔囊来源于原肠内胚层故又称肠体腔(enterocoel)。 形成方式称为肠体腔法(enterocoelous method)。见于后口动物 (棘皮动物,毛颚动物,半索动物,脊索动物)。高等脊索动 物由裂体腔法形成体腔。但复杂有差别。
2021/4/4
4
第三节 胚胎发育的重要阶段
动物早期胚胎发育的几个主 要阶段是相同的。
(一)受精与受精卵:
1、雌体产生雌性生殖细胞, 称 为 卵 (egg) 。 依 卵 黄 多 少 , 分为少黄卵、中黄卵、多黄 卵。卵黄多的一端为植物极 (vegetal pole),另一端称为动 物极(animal pole)。
2021/4/4
2
(1)菱形虫纲(Rhombozoa):20-40个细胞,同种数目固定。 寄生在头足类软体动物的肾内,如双胚虫(dicyemida),异胚虫 (heterocyemida)。
(2)直泳虫纲(Orthonecta):雌大,雄小,寄生在无脊椎动物 体内,如直泳虫(orthonectid)。
2、内移(ingression):由囊胚一部分细胞移入内部形成内胚 层。没有胚孔,以后形成胚层。
2021/4/4
10
1.什么是传统机械按键设计?
传统的机械按键设计是需要手动按压按键触动PCBA上的 开关按键来实现功能的一种设计方式。
传统机械按键结构层图:
按
PCBA
键
开关 键
传统机械按键设计要点: 1.合理的选择按键的类型, 尽量选择平头类的按键,以 防按键下陷。 2.开关按键和塑胶按键设计 间隙建议留0.05~0.1mm,以 防按键死键。 3.要考虑成型工艺,合理计 算累积公差,以防按键手感 不良。
多细胞动物的起源

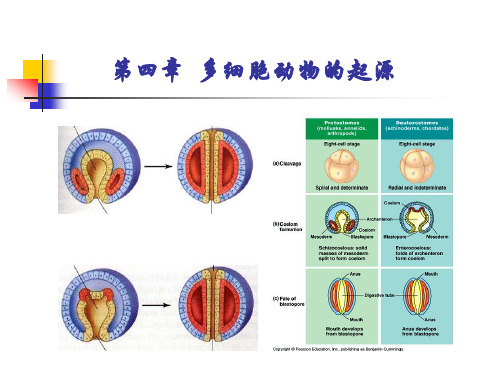
原口动物
后口动物
裂体腔法形成体腔
体腔囊发形成体腔
原口动物Protostomia——动物成体的口来源于 胚胎时期的原口(即胚孔)的一类三胚层动物 (包括扁形动物、假体腔动物、环节动物、软体动物和 节肢动物等).
2、卵裂
受精卵发生 分裂,细胞数 目在增加,形 成桑椹胚。
(一)受精和受精卵
精子Biblioteka 1. 生殖细胞少黄卵(如文昌鱼卵、海胆卵)
卵 中黄卵(如昆虫卵)
多黄卵(如鸡、鸟卵)
2. 受精:精子和卵结合成为受精卵的过程称为受精。
(二)卵裂
1. 卵裂的特点:不等分裂球长大,又进行下一次分裂,结
果越分越小。
等裂(如海胆的卵裂)
一、动物界的进化概况
大约12亿年前,动物和植物开始分化,经过漫长的岁月, 进化出地球上最庞大最繁荣的生物类群。
1、动物进化的基本线索:
1)动物是从单细胞到多细胞的,是由原始鞭毛生物进化而 来,一般认为,动物和植物有共同的祖先。
2)动物的体型是由辐射对称向两侧对称发展。
3)身体结构是由二胚层向三胚层,由无体腔向原体腔直至 真体腔发展, 由原口动物分化出后口动物,并成为动物 界进化的主干。
4)脊索和脊椎的出现是后口动物进化过程中的重要事件; 5)由水登陆、羊膜卵和恒温的出现是脊椎动物进化中的里
程碑.
一、动物界的进化概况
2、动物进化的三大趋势: 从简单到复杂 从低等到高等 从水生到陆生
3、动物界
原生动物(亚界):单细胞动物 中生动物(亚界) (少数小型内寄生动物) 后生动物(亚界) :绝大多数多细胞动物
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
成中胚层和体腔。
中胚层形成示意图
3.6 神经胚
原肠胚形成后,胚胎背部沿中线的外胚层 细胞下陷形成神经板。其两侧的外胚层细胞形 成一对纵褶,两边的纵褶靠拢并在背面愈合,
形成中空的神经管。神经管逐渐进入胚胎内部
并与表面分离。这时的发育阶段称为神经胚。 以后神经管扩展,前端形成脑;后端延伸形成 脊髓
一、概念: 德国人赫克尔用生物进化论的观点总结了胚胎
学研究成果提出的。生物发展史分为个体发育和
系统发展史2个相互密切联系的部分,即个体发育 历史和由同一起源所产生的生物类群的历史密切 联系,个体发育史是系统发展史的简单而迅速地 重演.也叫生物重演律★
举例
如青蛙的个体发育由受精卵→囊胚→原肠 胚→中胚层→无腿蝌蚪→有腿蝌蚪→成体青蛙.
第4章 多细胞动物的起源
4.1 从单细胞到多细胞 4.2 多细胞动物起源于单细胞的 证据 4.3 胚胎发育的重要阶段 4.4 生物发生律 4.5 关于多细胞动物起源的学说
4.1 从单细胞到多细胞
原生动物—单细胞或群体 后生动物—多个多种细胞, 40--60万亿 中生动物★——1毫米至数毫米, 不 到100个细胞, 两层, 体细胞和 轴细胞,蠕虫状,寄生于浅海海 底无脊椎动物体内, 50种
4.3.4 原肠胚的形成
囊胚进一步发育形成原肠胚,出现了内、 外两胚层和原肠腔. 方式: 内陷, 内移, 分层, 内转, 外 包
(1)内 陷
由囊胚的植物极细胞向内陷入,最后形 成二层细胞,如海星原肠的形成。
(2)内 移
囊胚一部分细胞移入内部形成内胚层。 开始移入的细胞充满了囊胚腔,以后逐 渐形成内胚层和出现了原肠腔,以后在 胚的一端开一胚孔。某些腔肠动物的原 肠的形成.
4.3.1 受精与受精卵
由雌、雄个体产生雌雄生殖细胞,
雌性生殖细胞称为卵;雄性生殖细胞 称为精子。精子与卵结合为一个细胞
称为受精卵。
受精
受精是新生命的起点: 指精子、卵子各自 的单倍体基因组相融合形成二倍体合子 的事件。 在通常的情况下, 受精过程包括精卵相 遇,精子穿入卵子,引发卵子发生一系 列变化, 最终是二者原核的融合,形成 二倍体的合子。
太阳能,保证了胚胎发育时所许的温度条件。
4.3.2 卵裂
受精卵的分裂称为卵裂,它是一种特殊的 分裂方式,即在每次分裂之后,未到新细胞长 大就继续不断下去;因此分裂成的细胞就越 来越小,这些细胞叫做分裂球。
卵裂的方式
由于不同类动物卵细胞内卵黄多少和 卵黄在卵内分布情况的不同,卵裂的方 式也不同。 完全卵裂 不完全卵裂
4.2 多细胞起源单细胞的证据
4.2.1 古生物学证据 大量的古生物化石(依照地质年代证明 了单细胞动物化石→多细胞动物化石的 事实). 反映了生物由低等向高等发展的 顺序。
一、细胞起源单细胞的证据
4.2.2 形态学证据 现有的动物中有单细胞动物、群体单细
胞动物、多细胞动物, 形成了由简单到复
分 层 内 转 外 包
4.3.5 中胚层及体腔的形成 大多数动物除了内、外胚层之外, 还进一步发育, 在内外胚层之间形成 了中胚层。中胚层形成的方式主要
有裂体腔法★ (端细胞法)和体腔囊
法★(肠腔法)2种。
(1) 裂体腔法
裂体腔法(端细胞法):植物极有一个细胞叫
中胚层端细胞,经一次分裂成2个原始中胚层
4.5 多细胞动物起源的学说 (自学)
(一) 群体学说(colonial theory)
赫克尔的原肠虫学说 梅契尼柯夫的吞噬虫学说 海曼的扁囊胚虫学说 (二)合胞体学说 (三 )共生学说
受精过程示意图
卵的极性
卵子的细胞核位置和细胞质成分的分布 上是非均向的,即不对称的。 根据卵黄多少将卵分为少黄卵、中黄卵 和多黄卵。卵黄相对多的一端为植物极; 另一端为动物极。
动 物 极 —— 植 物 —— 极
卵黄多少 颜色深浅 细胞大小
少 深 小
多
浅
大
优势: 色素较多的动物极向上,可以吸收到大量的
等裂
不等裂
1、完全卵裂:在分裂时,受精
卵分裂为完全分离的单个细胞. ①如果分裂的子细胞形状大小 相同的卵裂叫完全均等卵裂 (等裂)。如海胆,文昌鱼等
的卵.
②如果分裂的子细胞大小不等,
则称为完全不等卵裂(不等
裂) 。如海绵动物,蛙类等的 卵。
2、不完全卵裂:受精卵分裂
盘 不彻底,仅一部分发生分裂。 裂 ①若卵黄多,细胞质和细胞 核集中于卵一端,分裂仅限在 不含卵黄一端的叫盘裂,如乌
神经胚的形成过程
4.3.7 胚层的分化和器官形成
外胚层分化成皮肤上皮及其各种衍生物、神 经组织、主要感觉器官、消化管的两端。 中胚层分化成肌肉、结缔组织、循环系统,生
殖系统与排泄系统的大部分器官。
内胚层分化成消化管大部分的上皮、肝、胰、
呼吸器官、生殖与排泄器官的小部分.
4.4 生物发生律(biogenetic law)
细胞,对称排列在胚孔两侧,以后各自分裂形
成中胚层。在中胚层之间形成的空腔即为体腔。 如环节、软体、节肢等原口动物都以此法形成 中胚层和体腔
(2) 体腔囊法
体腔囊法(肠腔法):内胚层两侧细胞外突形成 一对体腔囊,然后脱离内胚层,在内外胚层之
间形成中胚层,由中胚层包围的空腔为体腔。
后口动物如:棘皮、半索、脊索动物以此法形
这也反映了它在系统发展过程中经历了像单细 胞动物→单细胞球状群体→腔肠动物→原始三 胚层动物→鱼类动物→有尾两栖动物→无尾两 栖动物的基本过程. 即青蛙个体发育重演了其祖先的进化过程,也 就是个体发育简短重演了它的系统发展。
生物发生律对了解动物的演化与亲缘 关系的意义
个体发育不是简单地机械的重复,个体发 育中也会有变异出现,它又不断地补充系 统发展。 个体发育与系统发展相互联系、相互制约, 系统发展通过遗传决定个体发育,个体发 育不仅简短重演系统发展,而且又补充和 丰富系统发展,即其种族发展史.
杂、由低等向高等的序列。
一、细胞起源单细胞的证据
胚胎学证据:
多细胞动物的胚胎发育中,由受精卵开 始,经过卵裂→囊胚→原肠胚→中胚层 →神经胚等一系列过程发育为成体.根据 生物发生律可以说明多细胞起源于单细 胞动物.
4. 3 胚胎发育的重要阶段
胚胎发育 胚后发育
受精卵
胚后发育
幼体
成体
卵生动物: 幼体从卵膜内孵化出来→成体 胎生或卵胎生动物: 幼体从母体内生出来→成体
表 裂
贼,鸟,鱼的卵。
②如果大量卵黄集中在卵 的中央,分裂只限于卵表面的 叫表裂。如昆虫的卵。
4.3.3 囊胚的形成
卵裂后分裂形成中空的球状胚, 称为囊胚。囊胚中间的空腔叫囊 胚腔。囊胚壁的细胞层叫囊胚层。
海胆胚胎发育模式
前两次卵裂通过 动植物轴,沿经线进 行——经裂; 第3次卵裂沿赤道方 向进行,垂直于动植 物轴——纬裂