中国农业科学院植物保护研究所揭示水稻程序性细胞死亡新机制
2023年植保题库

植保题库:填空题:1.水稻上常发生旳病害有:稻瘟病、水稻条纹叶枯病、稻曲病、水稻纹枯病。
2.农民常说旳水稻钻心虫,实际上就是二化螟、三化螟。
3.水稻卷叶虫实际上就是:稻纵卷叶螟4.水稻稻曲病对产量影响较大,为害稻粒呈霉团,俗称:灰苞。
5.二化螟和三化螟,成虫不为害水稻,幼虫钻蛀水稻茎秆,卵重要产在叶片上,在苗期和分蘖期导致枯心苗;在孕穗初期侵入,导致枯孕穗,在孕穗末期和抽穗初期侵入,咬断穗颈,导致白穗或虫伤株。
6.二化螟初孵幼虫还会在叶鞘取食导致枯鞘。
7.水稻最轻易受二、三化螟为害旳是:分蘖期和孕穗期。
8.稻纵卷叶螟是一种迁飞性害虫。
原是局部发生,间歇为害,现已成为常发性害虫。
9.稻飞虱重要群集在稻株旳下部取食,用刺吸式口器刺进稻株组织吸食汁液。
虫量大时引起稻株下部变黑,瘫痪倒伏,俗称“冒顶,透天”导致严重减产或失收。
10.我企业防治稻飞虱旳药剂重要有旋扫、锐虱、拂净、飞悬、捕飞。
11.稻瘟病在水稻整个生育期中都可以发生,为害秧苗,称苗瘟,为害叶片称叶瘟,为害穗称穗瘟,为害节称节瘟。
12.蔬菜上常发生旳吸汁类害虫是:粉虱、蓟马、蚜虫。
13.蔬菜上常发生旳食叶害虫是:小菜蛾、甜菜夜蛾、斜纹夜蛾、菜青虫和跳甲。
14.斜纹夜蛾初孵幼虫群集取食,3龄前仅食叶肉,残留上表皮及叶脉,呈白纱状后转黄,易于识别。
4龄后进入暴食期,多在傍晚出来为害。
15.棉花苗期到成株期,常见旳根部病害有:棉花立枯病和棉花枯黄萎病。
二、选择题:单项选择题5分1、在一天当中,喷洒农药旳最佳时段是:( C )A、上午或下午B、中午阳光直射时C、清晨或傍晚D、有风旳时候2、如下是细菌性病害旳是( B )。
A、柑橘炭疽病B、白菜软腐病C、水稻纹枯病D、水稻条纹叶枯病3、如下是病毒引起旳病害是( A )。
A、水稻条纹叶枯病B、水稻纹枯病C、白菜干烧心D、黄瓜炭疽病4、棉花盲蝽蟓会导致(C).A、穿孔B、烂叶C、公棉花或多头棉D、落花5、褐、白稻飞虱以( A )虫态为害水稻A成虫,若虫;B、卵;C卵、若虫;D成虫、卵三、是非判断题:5分1、稻飞虱、稻纵卷叶螟都是迁飞性旳害虫。
《盐胁迫下水稻苗期生理响应及应答机制》

《盐胁迫下水稻苗期生理响应及应答机制》一、引言随着全球气候的变化,土壤盐渍化问题日益严重,对农业生产产生了巨大的影响。
水稻作为我国最重要的粮食作物之一,其生长受到盐胁迫的威胁也愈发明显。
因此,研究盐胁迫下水稻苗期的生理响应及应答机制,对于提高水稻抗盐性、保障粮食安全具有重要意义。
本文旨在探讨盐胁迫对水稻苗期生理指标的影响,以及水稻的应答机制,以期为农业生产提供理论依据。
二、材料与方法1. 材料选取当地常见的水稻品种作为试验材料,培育至苗期阶段。
2. 方法(1)盐胁迫处理将水稻苗期植株置于含有不同浓度盐溶液的培养环境中,模拟盐胁迫条件。
设置不同浓度梯度,如0(对照组)、50、100、150mM NaCl等。
(2)生理指标测定测定不同盐浓度处理下的水稻叶片的叶绿素含量、光合作用速率、气孔导度等生理指标。
(3)应答机制分析通过转录组测序、蛋白质组学等方法,分析盐胁迫下水稻的基因表达、蛋白质变化等应答机制。
三、盐胁迫下水稻苗期的生理响应1. 叶绿素含量变化随着盐浓度的增加,水稻叶片的叶绿素含量逐渐降低。
高盐环境下,叶绿体的结构受到破坏,导致叶绿素合成受阻。
2. 光合作用速率变化盐胁迫下,水稻的光合作用速率降低。
这可能是由于气孔导度降低、光合酶活性受抑等因素所致。
3. 渗透调节物质变化在盐胁迫下,水稻体内脯氨酸、可溶性糖等渗透调节物质含量升高,以维持细胞内外的渗透平衡。
四、水稻的应答机制1. 基因表达变化转录组测序结果显示,盐胁迫下水稻的基因表达发生显著变化,涉及光合作用、渗透调节、抗氧化等途径的相关基因表达上调或下调。
2. 蛋白质组学分析蛋白质组学分析表明,盐胁迫下水稻的蛋白质表达也发生改变,如与渗透调节、抗氧化相关的蛋白质含量升高,参与光合作用的酶类活性受到调控等。
3. 抗逆性物质合成与积累在盐胁迫下,水稻体内合成并积累了一系列抗逆性物质,如抗氧化酶类、渗透调节物质等,以应对盐胁迫带来的不利影响。
五、结论本文通过研究盐胁迫下水稻苗期的生理响应及应答机制,发现盐胁迫对水稻的生长产生不利影响,导致叶绿素含量降低、光合作用速率下降等生理指标的变化。
水稻测试题库

水稻测试题库您的姓名: [填空题] *_________________________________1. 稻瘟病菌主要以分生孢子和菌丝在病稻或病谷上越冬,成为次年发病的主要侵染源。
做好化学药剂的预防非常重要,常见的化学药剂有( )。
[单选题] *三环唑稻瘟灵春雷霉素以上均正确(正确答案)2. 水稻发生纹枯病后,在叶鞘、叶片和穗部都表现为不同的症状,由( )病原菌引起的。
[单选题] *立枯丝核菌(正确答案)稻梨孢串珠镰孢黄单胞菌3. 水稻纹枯病病斑在叶鞘和叶片上较为常见,病斑具有( )典型特征。
[单选题] *云纹状(正确答案)黄褐色“三部一线”条形斑4. “健苗防病种子处理+浸种催芽技术”是MAP稻区苗期主推的核心技术,通过药剂浸种可有效预防恶苗病。
[单选题] *咪鲜胺(正确答案)三环唑苯醚甲环唑戊唑醇5. 水稻常见穗部病害有( ) [单选题] *绵腐病恶苗病纹枯病稻曲病(正确答案)细菌性条斑病6. 水稻秧苗期间,若管理不当,易发生立枯病、绵腐等病害,为做好此类病害预防,苗期一般常用以下( )药剂进行喷雾处理。
[单选题] *甲霜灵·噁霉灵(正确答案)咪鲜胺丁草胺氰氟草酯7. 稻曲病主要集中在水稻生育时期最易感病。
[单选题] *分蘖期幼穗形成至抽穗扬花(正确答案)齐穗期灌浆期8. 稻曲病是危害穗部的典型病害,不仅影响产量和品质,而且病菌中含有对人、畜、禽有毒物质。
此病害主要采取喷施戊唑醇、苯醚甲环唑等药剂进行预防,施药时期极其重要,主要在水稻时期进行喷施。
[单选题] *分蘖期孕穗期(正确答案)齐穗期灌浆期9. 细菌性条斑病溢出的菌脓,可借风雨、露滴、水流及叶片之间的接触等途径传播,进行再侵染。
若田间发现有发病地块,用化学药剂防治时应先喷施然后喷施。
[单选题] *未发病地块、发病地块(正确答案)发病地块、未发病地块只喷发病地块以上说法均不对10. 二化螟以虫态在稻草、稻桩及茭白、玉米、高粱、甘蔗、油菜、蚕豆、麦类以及芦苇、稗、李氏禾等杂草根茎、茎杆中越冬。
中美科学家合作研究揭示植物脱落酸的一种信号通路运作新机制

解 了两种重要嗜热型真菌的基因组 , 为未来高效制造生 物燃 料的研究打下 了坚实的基础 。这两种真菌分别名为“ 瑞斯 太 梭孢壳霉 ” 耐热 性毁 丝霉 ” 其所 产生 的酶 的活跃 温度在 和“ , 7 氏度至 8 氏度之 间。基 因测序结果 表明 , O摄 0摄 这两种 真 菌分别含有 37 80万和 39 6 0万个碱基对。 使用生物燃料替代石油和煤炭等化石燃 料 , 以减少碳排 放、 保护生态环境 的想 法落 实起来 困难 很 多 , 战之一 是至 挑 今没有找到经济 的生物燃料制造 方法。最初 , 物燃料生产 生
日发现甘草药用成分 遗传 基因
nL )一P 2  ̄e s P C复合物结构 , 中发 现 S R 2和 A A受 体对 从 nK B P2 P C识别 中的惊人 相似 性。SR 2是广泛 存在 于植物 中的 nK
一
类 SrT r e/ h 类蛋 白激酶 , 与植 物体 内多种 信号 途径 的转 参 据 21 0 1年 1 1月 2 9日《 科技 日报》 报道 , 日本大 阪大 学 的一个研究小组发 现 , 有两个遗传基 因参与 了合 成甘草 的医 [ 神经科 学]
・
7 ・ 0
生物学教学 2 2 第3卷) 期 0 年( 7 第5 1
公 司科学家兰迪 ・ 尔卡领 导的 一个 国际科研团 队近 日破 贝
抚顺 西露天煤矿是我国最大的琥 珀产地 , 也是我 国唯一
的含虫琥珀产地 。抚 顺琥 珀的时代 为早 始新世 。其 昆虫 类
群多样性较高 , 中国科学 院南京地质古生物研究 所馆藏标 本
口罩都无法 防范 的微小颗粒。 人类 至今仍 然没有完全 弄清楚什 么样 的颗粒物才 能长 成雾滴 , 它里面 各种 各样 的颗 粒物 在雾 水 里面 是 怎么 分布 的 。而这篇题为 ” 一雨事件 中多环芳烃 的污染特 征 ” 雾 的文 章 , 次在雾化 学及有机 污染物分布机 理方面取得 了实质性 首
重磅!近十年我国农业科技的30个标志性成就公布!

重磅!近十年我国农业科技的30个标志性成就公布!党的十八大以来,我国农业科技取得了一系列重大突破,农业科技整体研发实力进入世界前列,在生物育种、土壤改良、畜禽水产养殖、植保与疫病防控、精准栽培、设施农业、农机装备、产后加工、质量安全、绿色发展等多个领域,涌现出一大批标志性成果,创新水平进入世界第一方阵。
全国农业科技创新工作会议召开在即,微观三农从今日起,将连续刊发一组报道,为读者梳理分享十年来我国农业科技的30个标志性成就,期待更多朋友了解、关心、支持农业科技工作。
1.水稻基因组学研究及应用国际领先开创了水稻研究从传统遗传图谱向全基因组水平转变的先河,引领了水稻精准设计育种的新方向,攻克了水稻生产中产量与多个重要性状之间相互制约的世界性育种难题,突破了水稻超高产与高品质协同改良的理论和技术瓶颈,奠定了我国在水稻新品种创制理论和技术领域的国际领跑地位,是农业领域重大基础理论突破,具有世界性、革命性意义。
2.超级稻亩产突破1000公斤全国水稻各科技创新团队,选育了多个亩产超过1000公斤的超级稻新品种,年均推广面积超过1.3亿亩,2018年“超优千号”品种在云南个旧再创超级杂交稻百亩示范片亩产1152.3公斤新高。
超级稻的研发应用,成为农业科技自主创新、协同攻关的典范,为保障我国粮食安全发挥了重要作用。
3.水稻育种屡获重大新突破,育种水平又攀新高峰创新两系法杂交水稻技术,建立了光温敏不育系的两系法杂种优势有效利用的新途径,实现了超级杂交稻超高产、米质优、抗性强的有机结合。
提出超高产专用早稻育种理论与方法,并创制优质稻米新种质。
培育了中嘉早17、龙粳31、五峰优T025等优质新品种,为确保口粮绝对安全提供了坚实的科技支撑。
4.寒地早粳稻优质高产多抗龙粳系列新品种国际领先开创了寒地早粳稻育种的独特理论与技术体系,在优异种质材料创制、寒地早粳稻优质高产多抗新品种培育等方面,达到国际同类研究领先水平,在黑龙江省内外累计推广1.37亿亩。
研究的背景和目的意义-PPT精品资料

2.研究内容
不同土壤水分胁迫和氮肥水平下,稻麦籽粒灌浆和胚乳 细胞增殖与乙烯的关系。不同土壤水分胁迫和氮肥水平下, 稻麦胚乳细胞淀粉粒形成和胚乳细胞核酸代谢与乙烯的关系。 揭示乙烯对稻麦粒重形成的生理机制。
3.研究方法
3.1水分试验方案
3.2.1材料 水稻:扬稻6号(籼稻),武育粳8号(粳稻) 小麦:扬麦15,扬麦16 3.2.2栽培方法 采用盆钵栽培,按普通方法栽培。水稻每盆3穴,粳稻双本, 籼稻单本。小麦每盆播种16粒,在三叶一心时间苗至每盆8 株。
1.研究的背景和目的意义
1.1本项研究的重要性
人类正面临粮食、资源和环境等迫切问题,其中粮
食问题尤为突出。水稻和小麦是人类最主要的粮食作物,
与玉米一起被称作三大粮食作物。
据联合国粮农组织统计,2000年 世界稻麦种植面积为3.65亿hm2,占谷 物 总 面 积 的 54.18% 。 我 国 2019 年 水 稻种植面积4963万hm2,占粮食作物 面积的49.21%,稻麦总产2.71 亿吨, 占 粮 食 作 物 总 产 的 57.74% 。 稻 麦 产 量的高低关系到我国乃至世界的粮 食安全和国计民生。
水分胁迫是导致植株体内乙烯释放量增加的一个重要 因素。水分胁迫能引起1-氨基环丙烷-1-羧酸(ACC)合成酶 的增加,从而导致干旱条件下ACC的急剧积累和乙烯的大量 释放,其结果是抑制叶片生长,减少水分消耗,促进植株衰 老。Yang等和Apelbau等用离体小麦叶片为材料阐明干旱诱 导乙烯的合成和基础乙烯一样,亦遵循蛋氨酸(Met)→S腺苷蛋氨酸(SAM)→ACC→乙烯(C2H4)的代谢途径。Hoff man等的工作进一步指出,干旱时乙烯前体ACC除了可以转变 成乙烯外,还能够转变成丙二酰基-1-氨基环丙烷-1-羧酸 (MACC)从而调节乙烯的生成。
我国作物遗传育种学科的发展现状与“十三五”发展重点

作物遗传育种学科在农业科学中占有核心地位[1],其根本任务是从基因型和环境2个层面研究并形成作物持续高产、优质、高效的理论、方法和技术,是关于大田作物生产与品种遗传改良的学科。
学科的创新发展,对保障国家粮食安全具有重大的意义。
粮食产量的“十一连增”[2]离不开作物遗传育种学科的创新和进步。
近年来,随着生物技术、信息技术和新材料技术的快速发展,我国的传统作物遗传育种学科迎来了新的发展机遇,成为生命科学领域具有发展潜力的学科之一,在基础、应用基础和应用研究方面取得了突破性进展,研发了1批重大的科技成果,对国家粮食安全和农业可持续发展做出了显著贡献。
1我国作物遗传育种学科的主要研究进展1.1基因组学等新技术广泛渗透近10a ,由“Next Generation Sequencing (NGS )”技术引领的基因组学技术正在一个空前的高速度推动下迅猛发展。
目前,高通量NGS 技术已经成为生命科学领域中应用最为广泛的研究手段。
例如,完成了小麦A 、D 基因组等图谱的绘制[3,4];构建了第2代玉米单体型图谱,其中包含了5500万个SNP 标记[5];对二倍体棉花———雷蒙德氏棉全基因组进行测序,并阐述了棉花基因组的多倍化及其纤维发育[6]。
此外,基DOI :10.16318/ki.hbnykx.2015.06.018河北农业科学,2015,19(6):66-70Journal of Hebei Agricultural Sciences编辑蔡海燕我国作物遗传育种学科的发展现状与“十三五”发展重点张江丽1,董文琦2,杜晓东3*(1.中国农业科学院科技管理局,北京100081;2.河北省农林科学院,河北石家庄050051;3.河北省农林科学院农业信息与经济研究所,河北石家庄050051)摘要:作物遗传育种学科的创新发展,对促进现代农业的健康发展和保障我国粮食安全具有重大的意义。
作者总结了近年来我国在作物遗传育种学方面取得的主要进展,从基因组学、种质资源保护与利用、新基因挖掘、作物杂种优势机理及利用、分子标记育种、分子设计育种、作物细胞工程和诱变育种等方面分析了“十三五”重点发展的方向。
2023年度中国科学十大进展

中国科学十大进展Top Ten Scientific Advances in China引用格式:2023年度中国科学十大进展. 中国科学院院刊, 2024, 39(3): 582-587, doi: 10.16418/j.issn.1000-3045.20240229002.Top ten scientific advances in China, 2023. Bulletin of Chinese Academy of Sciences, 2024, 39(3): 582-587, doi: 10.16418/j.issn.1000-3045.20240229002. (in Chinese)2023年度中国科学十大进展“中国科学十大进展”遴选活动旨在宣传我国重大基础研究科学进展,激励广大科技工作者的科学热情,开展基础研究科学普及,促进公众了解、关心和支持基础研究,在全社会营造浓厚的科学氛围,已成为盘点我国基础研究领域年度重大科学成果的品牌活动。
2023年度第19届“中国科学十大进展”遴选活动由国家自然科学基金委员会主办,国家自然科学基金委员会高技术研究发展中心(基础研究管理中心)和科学传播与成果转化中心承办,《中国基础科学》《科技导报》《中国科学院院刊》《中国科学基金》《科学通报》5家编辑部协办,分为推荐、初选、终选、审议4个环节。
共推荐了2022年12月1日至2023年11月30日期间正式发表的600多项科学研究成果;由近100位相关学科领域专家从中遴选出30项成果;在此基础上邀请了包括中国科学院院士、中国工程院院士在内的2 100多位基础研究领域高水平专家对30项成果进行投票,评选出10项重大科学研究成果;经国家自然科学基金委员会咨询委员会审议,最终确定入选2023年度“中国科学十大进展”的成果名单。
1 人工智能大模型为精准天气预报带来新突破天气预报是国际科学前沿问题,具有重大的社会价值。
我国农业分子育种发展现状

2023年第1期中 国 甜 菜 糖 业2023No.1文章编号:1002-0551(2023)01-0030-03我国农业分子育种发展现状张德财(依安东方瑞雪糖业有限责任公司,齐齐哈尔161500)摘 要:近年来,全球范围内生物育种技术不断取得重大突破,现代种业已进入“常规育种+现代生物技术育种+信息化育种”的“4.0时代”,正迎来以全基因组选择、基因编辑、合成生物及人工智能等技术融合发展为标志的新一轮科技革命。
本文介绍了当前中国种业发展历程并对比了目前国际作物分子育种发展现状,分析了中国分子育种面临的挑战和机遇。
关键词:分子育种;转基因;生物安全中图分类号:F323.2 文献标识码:B doi:10.3969/j.issn.1002-0551.2023.01.0060 前言粮食安全是“国之大者”,培育优良品种是保障国家粮食安全的根本途径。
“种子是农业发展的芯片”,发展生物育种技术,是发展现代农业、解决农业安全问题的重要支撑。
在我国已经成为共识。
生物育种发展已经被列入我国科技发展的重大战略方向之一,抢占生物育种技术及其产业发展制高点,已经成为各国增强农业产业核心竞争力、保障农业安全的重大战略选择。
国际上将育种发展分为4个典型阶段(图1):育种1.0时代,人类驯化了大量野生植物进入农耕文明;育种2.0时代,育种家主要依赖经验并把统计学、数量遗传学和杂交育种策略应用到优良品种选育中;育种3.0时代,先进的生物技术包括分子标记辅助选择、基因工程在育种中广泛应用;随着人工智能、基因编辑、合成生物学等学科发展,育种进入由前沿科学技术引领的“生物技术+信息技术+人工智能”育种4.0时代。
传统常规育种大多依赖育种家经验,育种效率低、精准度差、育种周期长。
生物育种基于遗传学、分子生物学、基因组学、计算生物学和系统生物学理论,依赖先进生物技术,与生命科学基础研究密不可分,将成为支撑未来现代种业长足发展的决定力量。
深圳科学家成功破解墨兰基因

·35·种业资讯 深圳科学家成功破解墨兰基因 等基因工程技术可使作物所需水分减少25%科学家们透露,在作物中过度表达单一蛋白质的简单基因调整可能会导致植物需要的水分减少25%,但这不会影响植物的常规产量。
科学家希望这项突破性研究将带来新一代的节水农业,帮助受干旱和气候变化的困扰地区种植更多粮食。
来自伊利诺斯大学领导的一个国际科学家小组的研究发现,可以增加一种名为Photosystem II Subunit S(PsbS)的特定蛋白质,迫使植物部分关闭其气孔。
气孔是二氧化碳或氧气出入的门户,有助于调节光合作用。
科学家最初的假设是,通过限制气孔打开,植物不会通过蒸腾损失更多的水分,并且随后不需要尽可能多的水分生长。
然而研究人员怀疑植物在减少气孔打开的同时仍然可以吸收所需的二氧化碳。
这个假设在烟草作物中进行了测试,结果令人难以置信。
通过增加PsbS 表达,气孔开口减少,进入植物的二氧化碳与水逸出的比率提高了25%。
这实际上意味着植物将需要少25%的水来达到相同的光合速率。
实验还表明,采用基因工程技术改造的植物和未改造植物之间的总产量或大小没有显著差异。
该研究发表在《自然·通讯》杂志上。
(科学探索)中国农科院揭示水稻先天免疫新机制日前,中国农业科学院植物保护研究所植物抗病功能基因研究组揭示了水稻等单子叶植物特有的SD-1类型受体激酶在细胞死亡和水稻先天免疫反应中的调控机制。
该研究为水稻广谱抗病分子育种提供了新的理论切入点。
相关研究成果在《细胞—宿主与微生物(Cell Host & Microbe)》上发表。
植物类病斑突变体往往可以增强植物的广谱抗病性,对研究植物的先天免疫机制和创制新的广谱抗病材料具有重要意义。
王国梁研究员团队长期从事水稻抗病性遗传学研究,前期研究中鉴定到一个类病斑突变体spl11,该突变体显著增强了对稻瘟病和白叶枯病的抗耐性,图位克隆结果显示SPL11编码U-Box 类型泛素连接酶。
215494073_水稻病虫害绿色防控技术推广应用探究

农业灾害研究 2023,13(4)水稻病虫害绿色防控技术推广应用探究陆 勇1,谭献良2平南县植保站,广西平南 537300摘要 在我国水稻种植发展的过程中,病虫害产生的危害降低了水稻的产量和质量,为有效控制水稻中的农药残留量和污染程度、保障粮食安全及农民经济收入,需要加大水稻病虫害防治工作力度。
基于此,分析了当前水稻病虫害发生危害和防控面临的主要问题,提出了水稻病虫害绿色防控技术推广策略与措施,探究了水稻病虫害绿色防控技术推广应用模式。
关键词 水稻病虫害;绿色防控;推广应用中图分类号:S435.11 文献标识码:B 文章编号:2095–3305(2023)04–0049-03随着我国农业生产水平和人们生活质量的提高,对农产品品质提出了更高的要求。
但是由于传统种植模式下病虫害防治工作存在不足之处,导致水稻产量不高且抗逆性较差,严重制约了农民增产增收。
基于此,主要分析了水稻病虫害绿色防控技术的作用机理及其优势,以期为进一步推动我国水稻病虫害绿色防控技术发展提供参考依据。
1 水稻病虫害发生动态分析想要进一步增强水稻病虫害的综合治理效果,种植者需要充分了解水稻病虫害的种类分布、发生规律和危害特点,这样能够有针对性地选择绿色防控技术进行综合治理。
据有关部门统计,2022年平南县共种植水稻面积约为5 933.3 hm2,主要病虫害发生面积13.2万hm2。
1.1 稻飞虱稻飞虱的发生特点和危害要引起充分的重视。
在实际防治的过程中,可以通过使用化学农药和生物农药开展有效防治工作。
首先,需要根据当地的气候条件和土壤条件等因素选择适宜的农药品种,并且合理进行田间试验,从而保证所选的农药达到最佳效果。
其次,要做好相关的农药管理工作,确保用药安全可靠、无污染等。
同时,应当注意避免将杀虫剂与其他物质混合施用,这样会导致害虫产生抗药性而无法发挥出应有作用。
最后,还应加强对稻田生态环境的检查和控制,一旦发现有大量若虫或成虫,必须及时采取措施予以消灭。
水稻主要病害生物防治的研究进展

水稻主要病害生物防治的研究进展一、内容综述随着全球人口的增长和粮食需求的不断提高,水稻作为世界上最重要的粮食作物之一,其产量和质量对人类生存和发展具有重要意义。
然而水稻生产过程中病虫害的发生严重制约了水稻产量的提高和质量的保障。
为了解决这一问题,科学家们对水稻主要病害生物防治的研究取得了显著进展。
本文将综述近年来在水稻主要病害生物防治方面的主要研究进展,包括病原物鉴定、病害监测预警、生物防治技术研究等方面。
首先病原物鉴定是病害防治的基础,通过对水稻病原菌、病毒和寄生线虫等病原物的鉴定,科学家们可以明确病害的类型和来源,为制定针对性的防治措施提供依据。
近年来基因组学技术的发展为水稻病原物鉴定提供了新方法,如基于PCR技术的分子标记辅助鉴定技术、基于转录组测序技术的基因组学分析方法等,这些技术的应用大大提高了病原物鉴定的准确性和效率。
其次病害监测预警是病害防治的关键环节,通过对水稻生长过程中的各项指标进行实时监测,可以及时发现病害的发生和蔓延趋势,为采取有效的防治措施提供科学依据。
近年来随着遥感技术、无人机技术和人工智能技术的发展,水稻病害监测预警技术得到了极大提升,如基于遥感技术的多光谱影像分析方法、基于无人机技术的大范围快速监测方法等,这些技术的应用使得病害监测预警更加精确、高效和全面。
生物防治技术研究是实现水稻病害可持续控制的重要途径,通过研究和应用各种生物防治剂,如微生物制剂、昆虫防控剂和植物源性农药等,可以有效降低化学农药的使用量,减少环境污染,同时提高农业生产的经济效益。
近年来以基因工程为核心的生物防治技术研究取得了重要突破,如抗性基因的克隆和表达、新型生物防治剂的研发等,这些成果为水稻病害生物防治技术的推广应用奠定了坚实基础。
水稻主要病害生物防治的研究进展涉及病原物鉴定、病害监测预警和生物防治技术研究等多个方面,这些研究成果为实现水稻高产、优质、高效生产提供了有力支持。
然而由于水稻生产环境的复杂性和病害种类的多样性,未来仍需进一步深化研究,开发出更多高效、安全、环保的病害防治技术和方法。
核盘菌通过类似整联蛋白SSITL...

核盘菌通过类似整联蛋白(SSITL)抑制寄主的抗病反应目 录摘 要 (I)ABSTRACT (IV)缩略词表 (VIII)1. 前言综述 (1)1.1 核盘菌的危害及其防治 (1)1.1.1 核盘菌的危害及其生物学特性 (1)1.1.2 作物菌核病的防治研究 (1)1.2 植物病原菌与寄主植物的互作 (5)1.2.1 植物天然的的物理及生理生化防卫屏障 (5)1.2.2 植物的先天免疫系统 (6)1.2.3 植物的后天免疫系统 (10)1.2.4 植物的非寄主抗性 (13)1.2.5 不同类型植物病原菌的侵染策略以及互作方式 (14)1.2.6 核盘菌的侵染策略 (16)1.3基因功能研究的策略 (19)1.3.1丝状真菌的遗传转化的研究进展 (19)1.3.2基因的超标达、敲除和沉默 (20)1.3.3 蛋白质的定位 (24)1.4 Integrin以及Integrin–like基因的研究进展 (26)1.4.1 整联蛋白的结构 (27)1.4.2 整联蛋白的信号传导 (29)1.4.3整联蛋白在微生物中的生物学功能 (30)1.5 本项研究的目的和意义 (32)2. 材料与方法 (33)2.1 菌株及植物材料 (33)2.2 基因的生物信息学分析 (33)2.3 核酸的实验操作 (34)2.3.1 DNA的提取 (34)2.3.2 质粒的提取 (34)2.3.3 总RNA的提取 (35)2.3.4 RT和Real–Time PCR (35)2.3.5 Northern blot (36)2.4 蛋白质的实验操作 (37)2.4.1 SSITL的原核表达 (37)2.4.2 抗体血清的制备、效价(ELISA)以及特异性(Western blot)的检测 (37)2.4.3 SSITL的免疫胶体金亚细胞定位 (39)2.4.4 核盘菌侵染洋葱表皮过程中SSITL的免疫荧光定位 (40)2.5 相关载体的构建 (40)2.6 ATMT介导的真菌和植物转化 (41)2.7 生物学特性的实验研究 (43)2.7.1 生长速度、致病力、菌丝顶端分支以及菌落形态的观察 (43)华中农业大学2012届博士研究生学位论文2.7.2 菌核的培养、大小及重量的测定和菌核萌发的研究 (43)2.7.3 核盘菌产草酸能力的测定 (44)2.7.4 核盘菌侵染拟南芥叶片过程的观察 (45)2.8 SSITL与植物诱导抗性的关系 (45)2.8.1 核盘菌侵染拟南芥过程中SSITL基因的表达情况 (45)2.8.2 核盘菌侵侵染拟南芥过程中拟南芥局部抗性的动态变化 (45)2.8.3 核盘菌侵侵染拟南芥过程中拟南芥系统抗性的动态变化 (46)2.8.4 SSITL在植物中表达对植物的抗病性的影响 (46)3. 结果与分析 (47)3.1 SSITL的生物信息学分析 (47)3.1.1 SSITL的序列分析 (47)3.1.2 SSITL蛋白的同源比对分析及高级结构预测 (49)3.2 SSITL对核盘菌生物学特性的影响 (53)3.2.1 SSITL基因在核盘菌不同生长时期的表达 (53)3.2.2 SSITL基因沉默对核盘菌生物学特性的影响 (53)3.3 SSITL抗体的制备以及免疫胶体金亚细胞定位 (61)3.3.1 SSITL的原核诱导表达 (61)3.3.2 抗血清效价以及特异性测定 (63)3.3.3 SSITL蛋白的亚细胞定位 (63)3.4 SSITL基因在核盘菌与植物互作过程中的作用 (67)3.4.1 核盘菌侵染拟南芥时,SSITL基因的表达情况 (67)3.4.2 核盘菌SSITL对拟南芥局部防卫反应的影响 (68)3.4.3 核盘菌SSITL对拟南芥系统防卫反应的影响 (70)3.4.4 SSITL在寄主植物中瞬时表达对植物抗病性的影响 (74)3.4.5 SSITL在寄主植物中组成型表达对植物抗病性的影响 (79)3.4.6 SSITL的表达对烟草的影响 (81)4. 讨论 (83)4.1 SSITL基因生物学功能的深入探讨 (83)4.1.1 SSITL基因的序列分析 (83)4.1.2 SSITL基因的功能分析 (85)4.2 SSITL参与抑制植物诱导抗性 (87)4.2.1 SSITL基因在核盘菌侵染过程中被诱导表达 (88)4.2.2 SSITL参与抑制植物的局部抗性 (88)4.2.3 SSITL参与抑制植物的系统抗性 (89)4.2.4 SSITL基因在植物中表达后,植物的抗性受到抑制 (90)4.3 研究SSITL的互作蛋白以及作用机理 (90)4.4 结论与展望 (92)5. 参考文献 (94)附录: (116)博士期间发表的论文 (121)致 谢 (122)核盘菌通过类似整联蛋白(SSITL)抑制寄主的抗病反应摘 要核盘菌(Sclerotinia sclerotiorum)属于子囊菌门,是一种世界性分布的典型的死体营养型病原真菌。
中国植物科学与农学顶尖科学家排名

中国植物科学与农学顶尖科学家排名标题:植物科学与农学之巅:中国顶尖科学家排名导语:在中国植物科学与农学领域,一批批杰出的科学家通过不懈的努力与研究,为我们揭开了自然界的神秘面纱,并为农业的发展与改善做出了重要贡献。
本文将带您深入了解中国植物科学与农学顶尖科学家的排名,希望能为您呈现一个全面、深入的视角。
一、引言与背景植物科学与农学作为相互交织的领域,对于食品安全、生态环境和农业可持续发展等方面具有重要意义。
中国的植物科学与农学研究自改革开放以来取得了突飞猛进的发展,出现了一大批国际知名的科学家。
他们在科学创新、科研成果和植物育种方面都取得了举世瞩目的成就。
二、中国植物科学与农学顶尖科学家排名1. 中国农业科学院杂交水稻研究中心主任中国工程院院士袁隆平袁隆平作为我国稻作品种改良领域的开创者,他率先成功培育出了超级杂交水稻。
这项突破性成果为实现世界粮食安全做出了巨大贡献。
袁隆平不仅在农学领域有着杰出的成就,同时他也是一位富有爱心的科学家,多次帮助贫困地区改善水稻种植条件,为农民致富做出了积极努力。
2. 中国农业大学教授张福锁张福锁是中国植物病理学的杰出代表之一。
他在病毒病和真菌病防治方面做出了卓越的贡献。
张福锁教授领导的团队成功开发出了一种抗病性强的转基因作物,为农作物的抗病育种提供了新思路。
他的研究对于保障农作物的产量和质量具有重要的意义。
3. 中国科学院植物研究所研究员马万里马万里是我国植物生理学和植物分子生物学研究领域的知名科学家之一。
他对植物逆境胁迫响应机制的研究深入浅出,取得了一系列创新性成果。
他的研究成果对于提高作物的逆境适应性和生产力具有重要意义。
4. 中国农业科学院作物科学研究所研究员金绍阳金绍阳教授在作物遗传改良领域拥有丰富的学术经验和卓越的研究成果。
他的研究主要集中在小麦的基因克隆和利用、耐逆性与品质形成机制等方面。
他的研究不仅拓宽了我们对小麦的认识,还为小麦的品质改良和产量提高提供了有力的支持。
拮抗水稻病原菌假单胞菌AH_菌株的鉴定及全基因组序列分析

拮抗水稻病原菌假单胞菌AH菌株的鉴定及全基因组序列分析胡逸群,沈文杰,陈晴晴,张爱芳∗㊀(安徽省农业科学院植物保护与农产品质量安全研究所,安徽合肥230001)摘要㊀[目的]探究菌株AH的生防潜力和基因组序列信息,以解析其防病机制,挖掘次级代谢产物基因资源㊂[方法]采用稀释涂布平板法分离水稻的叶际微生物,筛选具有拮抗效果的菌株;根据形态学㊁16SrDNA和gyrB基因测序和系统发育分析,鉴定其种属;利用平板抑菌试验明确其抑菌谱;大田喷施菌悬液明确其防治效果㊂继而,利用第二代Illumina与第三代PacBio结合的测序方法对生防菌进行全基因组测序,对测序数据进行基因组组装㊁基因预测与功能注释㊁次级代谢产物合成基因簇预测等分析㊂[结果]AH是一株短杆状恶臭假单胞菌株,对稻瘟病菌㊁茎点霉菌㊁水稻纹枯病菌和水稻黄单胞菌具有广谱抗菌活性,且田间施用AH菌液可显著降低水稻白叶枯病和稻瘟病的病害等级㊂抗生素敏感性测试揭示菌株AH对氨基糖苷类抗生素高度敏感㊂AH基因组全长为5889125bp,编码5215个基因,GC含量为63.74%,有19个rRNA㊁77个tRNA和78个nRNA,antiSMASH预测得到8个次级代谢产物合成基因簇㊂[结论]AH菌株具有良好生防潜力,深层机制解析可为病害防控提供科学依据㊂关键词㊀恶臭假单胞AH;生物防治;稻瘟病;水稻白叶枯病中图分类号㊀S435.11㊀㊀文献标识码㊀A㊀㊀文章编号㊀0517-6611(2023)20-0138-09doi:10.3969/j.issn.0517-6611.2023.20.034㊀㊀㊀㊀㊀开放科学(资源服务)标识码(OSID):IdentificationandWholeGenomeAnalysisofPseudomonasputidaAHStrainAntagonistictoRicePathogensHUYi⁃qun,SHENWen⁃jie,CHENQing⁃qingetal㊀(InstituteofPlantProtectionandAgro⁃productsSafety,AnhuiAcademyofAgricultur⁃alSciences,Hefei,Anhui230001)Abstract㊀[Objective]BiocontrolpotentialandgenomeinformationofstrainAHwereanalyzedtorevealmechanismofdiseasepreventionandexploregeneresourcesofsecondarymetabolites.[Method]Themethodofspreadingplateswasusedtoscreenantagonisticstrainsfromriceleaves.Thespecieswasidentifiedbymorphological,16SrDNAandgyrBsequencingandphylogeneticanalysis.Theflatstandofftestandspra⁃yingbacterialsuspensioninfieldclarifieditsbiocontrolpotential.Subsequently,thegenomesequenceofAHstrainwasobtainedusingathirdgenerationPacbioSMRTsequencingplatformandsecond⁃generationIlluminasequencingtechnology.Then,genomeassembly,geneprediction,functionannotation,andsecondarymetabolitebiosyntheticgeneclusterpredictionwereperformed.[Result]AHwasashortrodstrain,belongedtoPseudomonasputida,andshowedbroad⁃spectrumantimicrobialactivityagainstMagnaportheoryzae,Phomasp.,Rhizoctoniasolani,andXan⁃thomonasoryzae.SprayingAHsuspensionsignificantlyreduceddiseasegradecausedbybacterialblightandblastdisease.Antibioticsusceptibili⁃tytestingrevealedthatAHshowedhighsensitivitytoaminoglycosideantibiotics.ThetotallengthofAHgenomewas5889125bp,encoding5215geneswith63.74%GCcontent,including19rRNA,77tRNAand78nRNA.And,eightsecondarymetabolitesynthesisgeneclusterswerepredictedbyantiSMASHplatforminAHgenome.[Conclusion]BasedongoodbiocontrolpotentialofAHstrain,theanalysisofdeepmechanismcanprovidescientificbasisfordiseasepreventionandcontrol.Keywords㊀PseudomonasputidaAH;Biocontrol;Riceblast;Ricebacterialblight基金项目㊀国家自然科学基金项目(32202273);安徽省自然科学基金项目(2108085QC117)㊂作者简介㊀胡逸群(1992 ),女,安徽灵璧人,助理研究员,博士,从事水稻病原菌致病机制和病原菌与生防菌互作研究㊂∗通信作者,研究员,从事水稻抗病性鉴定与病原菌致病机制研究㊂收稿日期㊀2023-05-12㊀㊀为缓解农药过度施用与环境保护之间的矛盾,微生物来源的生防菌剂或制品有较好的开发前景㊂假单胞菌属㊁芽孢杆菌属和木霉属来源的微生物,对多种植物或动物病原菌具有良好的抑菌活性,因此被广泛用作生物防治剂[1-4]㊂假单胞菌属具有代谢多样性的特点,可产生多种次级代谢产物,如吩嗪类物质(phenazines)㊁硝吡咯菌素(pyrrolnitrin)㊁2,4-二乙酰基间苯三酚(2,4-diacetylphloroglucinol)㊁黄绿菌素(pyocyanin)和环脂肽(cycliclipopeptides,CLPs)等㊂此外,假单胞菌属细菌也能通过诱导系统抗性的方式,抑制病原菌定殖或生长,对植物具有保护作用[5-8]㊂铜绿假单胞菌P.aeruginosaUPMP3菌株合成的吩嗪粗提物,可显著减轻狭长孢灵芝引起的油棕榈茎基底感染[9]㊂从苹果花上分离的P.orientalF9菌株表现出对火疫病菌的良好拮抗特性[10]㊂此外,大多数假单胞菌属菌株可作为植物根际促生菌,分泌生长调节剂来增强植物防御病原菌的能力㊂从印度青蒿根际分离的P.aeruginosa可以促进植物生长,且对植物病原菌链格孢㊁黄曲霉和尖孢镰刀菌表现出很好的生防活性[11]㊂恶臭假单胞P.putida与近缘物种P.fluorescens和P.bras⁃sicacearum可作为生物防治剂和植物生长促进剂[12-13]㊂P.putida与其他有益微生物复合施用时,显著增加了水果营养元素的含量[14-15]㊂P.putidaB2017菌株通过产生铁载体pyoverdine来抑制植物病原菌[16]㊂在有益于植物的同时,生防菌也需要使自己免于如活性氧爆发之类的快速且强烈的植物防御反应所导致的损伤㊂P.putidaRRF3通过改变根部基因的转录表达来刺激植物产生防御反应,并通过改变根际的化合物组分来保护自身[17]㊂一直以来,我国稻米主产区的真菌㊁细菌病害频发,稻瘟病菌可在水稻营养生长期侵染叶片导致叶瘟,在生殖生长期侵染稻穗导致穗颈瘟,严重降低稻米产量和品质[18]㊂镰孢菌Fusarium的复合侵染,如藤仓镰孢菌F.fujikuroi㊁层出镰孢菌F.proliferatum㊁拟轮枝镰孢菌F.verticillioides等,会降低种子发芽率,或引起水稻恶苗病[19]㊂此外,黄单胞菌Xan⁃thomonas是一类重要的植物病原细菌,可侵染400多种植物,包括稻㊁棉㊁豆等粮食作物和柑橘㊁香蕉和木薯等经济作物[20]㊂其中,X.oryzaepv.oryzae和X.oryzaepv.oryzicola是稻黄单胞的2个致病变种,分别引起水稻白叶枯病和细菌性条㊀㊀㊀安徽农业科学,J.AnhuiAgric.Sci.2023,51(20):138-146斑病,危害严重[21-22]㊂面对防病增产与生态保护之间日益突出的矛盾,广谱抑菌菌株的开发利用是减少化学制剂使用㊁降低环境污染的有效策略㊂叶际微生物的生境更为复杂多变,具备良好的环境适应性和多种生防机制㊂基于此,笔者通过分离水稻叶片微生物㊁平板对峙筛选拮抗菌株㊁多基因测序和系统发育分析鉴定得到一株具有良好抗菌活性的恶臭假单胞P.putidaAH菌株,田间试验明确其对水稻白叶枯病和稻瘟病具有防治效果㊂利用第二代Illumina与第三代PacBio结合的方法进行全基因组测序,并对测序数据进行基因组组装㊁基因预测与功能注释㊁次级代谢产物合成基因簇分析,为AH菌株的基因功能研究提供分子遗传信息㊂恶臭假单胞P.putidaAH菌株的研究可为水稻病害防控提供科学依据㊂1㊀材料与方法1.1㊀供试菌株和培养条件㊀稻瘟病菌Magnaportheoryzae㊁纹枯病菌Rhizoctoniasolani㊁茎点霉菌Phomasp.㊁水稻白叶枯病菌Xanthomonasoryzaepv.oryzae㊁尖孢镰孢菌Fusariumoxyspo⁃rum㊁层出镰孢菌F.proliferum㊁短小杆菌Curtobacteriumsp.,均保存于笔者所在实验室㊂细菌培养用LB培养基,白叶枯病菌培养用NA培养基,培养条件为28ħ,保存介质为50%甘油㊂真菌培养用PDA培养基,培养条件为28ħ,无菌滤纸对菌丝块进行低温保存㊂1.2㊀稻瘟拮抗细菌的分离和筛选㊀为从水稻叶际分离稻瘟病菌的拮抗细菌,用无菌剪刀将水稻叶片组织切成小块,在无菌水中浸泡30min,6000r/min离心3min获得上清液㊂将悬浮液进行梯度稀释(10-2㊁10-3㊁10-4㊁10-5),取不同浓度的稀释液50μL涂布于3个LB板上,平板于28ħ培养2d㊂挑取形态差异的单菌落进一步纯化保存,用于拮抗菌的筛选㊂稻瘟病菌孢子液稀释至2ˑ107个/mL,涂布PDA平板后,28ħ培养3d㊂取2μL上述细菌菌液滴于上述平板,28ħ培养3d,周围有抑菌圈的细菌即为目的菌,纯化并保存㊂1.3㊀拮抗细菌的形态观察㊀取20μL新鲜菌悬液滴到碳膜铜网,静置5min后用滤纸吸去多余液体㊂将2%磷钨酸滴在碳膜铜网,静置染色2min后用滤纸吸去多余液体,室温干燥㊂在透射电子显微镜下观察细菌形态,采集图像分析㊂1.4㊀细菌分类鉴定㊀利用细菌基因组DNA提取试剂盒提取基因组DNA㊂使用细菌通用引物[23]扩增16SrDNA基因(27F,5ᶄ-AGAGTTTGATCATGGCTCAG-3ᶄ;1492R,5ᶄ-ACG⁃GTTACCTTGTTACGACTT-3ᶄ)㊂使用特异性引物扩增gyrB序列(F,5ᶄ-GAGCAGACCTACGTCCACGGTGTG-3ᶄ;R,5ᶄ-GTCGATCATCTTGCCAACGACCGC-3ᶄ)㊂PCR扩增产物经过纯化后送生工生物技术公司进行测序㊂测序结果与BLAST(https://www.ncbi.nlm.nih.gov/)中核酸数据进行比对,选择同源性较高的近缘物种序列,利用MEGAX软件使用邻接法构建系统发育树㊂1.5㊀平板对峙试验㊀5种植物病原菌(M.oryzae,R.solani,F.oxysporum,F.proliferatum,X.oryzaepv.oryzae)和2种附生菌(Phomasp.,Curtobacteriumsp.)作为拮抗测试的靶标菌㊂将上述分离得到的拮抗细菌在液体LB中培养至生长对数期(OD600nm=1.0),用于平板对峙试验㊂拮抗真菌试验:将新鲜菌丝块放置在PDA平板中央,将2μL拮抗细菌菌悬液滴在平板边缘㊂平板于28ʎC培养箱培养3 7d,拍照并记录数据㊂拮抗细菌试验:将靶标菌的菌悬液(OD600nm=1.0)稀释100倍,涂布于LB平板,28ʎC培养1d后再将2μL拮抗细菌的菌悬液滴在上述平板边缘㊂平板放置在28ʎC培养箱培养3d,拍照并记录数据㊂1.6㊀田间防治试验㊀籼稻常规稻品种WH26作为参试材料,评价拮抗细菌对水稻3种病害(稻瘟病㊁水稻白叶枯病㊁细菌性条斑病)的田间防治效果㊂1月龄的水稻苗用于接种病原菌㊂病原菌接种前1和3d,分别用拮抗菌的菌悬液(OD600nm=0.5)均匀喷施水稻幼苗,以叶片布满微液滴为限㊂水稻白叶枯病菌接种:用无菌剪刀沾取X.oryzaepv.oryzae的菌悬液(OD600nm=0.5),在距离叶尖边缘约2cm处剪下叶片㊂选取10株水稻,每株取3片叶片进行接种处理㊂接种14d后进行病斑长度测量㊂水稻细菌性条斑病菌接种:用无针头的1mL注射器将X.oryzaepv.oryzicola菌悬液(OD600nm=0.5)注入水稻叶片的细胞间隙,注意避免对植物造成另外的机械损伤㊂选取10株水稻,每株取3片叶片进行接种处理㊂接种7d后进行病斑长度测量㊂稻瘟病菌接种:将M.oryzae孢子悬液浓度调整为2ˑ105个/mL,均匀喷施于水稻叶片,以叶片布满微液滴为限,接种7d后观察统计发病情况㊂1.7㊀抗生素敏感性试验㊀采用平板加药法测试拮抗细菌对庆大霉素(Gm)㊁卡那霉素(Kn)㊁氨苄霉素(Amp)和利福平(Rif)的敏感性㊂固体LB培养基中分别加入上述抗生素,配制终浓度分别为20.0㊁10.0㊁5.0㊁2.5μg/mL的抗生素平板㊂将拮抗菌的菌悬液(OD600nm=1.0)稀释100倍,均匀涂布于上述平板,28ħ培养1d,拍照并计数菌落㊂1.8㊀细菌全基因组测序㊀第二代Illumina与第三代PacBio结合的测序方法进行全基因组测序㊂离心收集新鲜的AH菌体,试剂盒法提取基因组DNA(GenomicDNAIsolationKit,TaKaRa)㊂使用G-tubes(Covaris)剪切基因组DNA,EXOVII处理DNA并进行DNA末端修复,制备SMRTbellDNA文库㊂基因组的从头组装基于三代数据,使用falcon进行原始数据的自我矫正及基因组初步组装,得到一致性序列㊂使用GenomicConsensus软件对原始数据再次矫正,使用sprai对一致性序列进行环化处理,得到环化的细菌基因组[24],Circos软件构建基因组圈图[25]㊂使用Prodigal软件预测编码基因[26],Rfam预测非编码RNA[27]㊂2㊀结果与分析2.1㊀生防细菌的分离㊀通过稻瘟孢子液平板的筛选方法,分离得到5株具有抑菌作用的细菌菌株,其中抑菌作用最显著的一株命名为AH,对其进行后续研究㊂在PDA培养基上培养2d后,AH菌落形态为黄白色,表面粗糙,边缘凹陷(图1A㊁B)㊂在LB培养基上,AH菌落形态与PDA培养基相似,菌体更为致密(图1C㊁D)㊂透射电镜观察到AH呈短杆菌(图1E㊁F)㊂93151卷20期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀胡逸群等㊀拮抗水稻病原菌假单胞菌AH菌株的鉴定及全基因组序列分析注:使用平板划线法(A,C)和菌液悬滴法(B,D),分别在PDA培养基(A,B)和LB培养基(C,D)培养菌株AH㊂透射电镜观察菌体形态(E:5.0μm;F:500nm)㊂Note:StrainAHwasculturedonPDAmedium(A,B)andLBmedium(C,D)bystreakplatemethod(A,C)anddroppingplateofliquidbacterialcul⁃ture(B,D),respectively.P.putidaAHstrainwasobservedbytransmissionelectronmicroscope(TEM),whichwastreatedwith2%phosphotungsticacidonthecoppergridtostainfor2minon5.0μm(E)and500nm(F)scale.图1㊀菌株AH的形态学特征Fig.1㊀ThemorphologicalfeaturesofstrainAH2.2㊀生防细菌的鉴定㊀AH的16SrDNA基因序列长度为1405bp,BLAST比对结果显示该序列与P.putida㊁P.wayam⁃bapalatensis㊁P.muyukensis㊁P.monteilii㊁P.plecoglossicida㊁P.shirazica和P.asiatica等菌株高度相似,相似度均高于99.87%,需进一步确定种属㊂利用特异性引物扩增得到gyrB基因序列,长度为708bp,BLAST比对结果显示该序列与P.putidaNX-1㊁P.putidaPC2同源性为97.74%,与P.wayam⁃bapalatensisRW3S1相似度为96.15%,与P.muyukensisCOW39相似度为93.04%,与P.monteilii各菌株相似度均为92%㊂利用MEGAX软件构建系统发育树,表明AH属于恶臭假单胞菌P.putida(图2)㊂图2㊀菌株AH基于gyrB基因的系统发育树Fig.2㊀PhylogenetictreebasedonthenucleotidesequencesofthecompletegyrBgene041㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀安徽农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀2023年2.3㊀菌株AH的生防潜力㊀菌株AH对参试的7种菌株均表现拮抗活性,其中对稻瘟病菌㊁水稻白叶枯病菌㊁短小杆菌和茎点霉抑制作用显著,抑菌圈直径均大于1.3cm(图3A㊁B㊁F㊁G,图4)㊂对于水稻纹枯病菌㊁尖孢镰孢菌和层出镰孢菌的抑菌圈直径小于1.0cm(图3C㊁D㊁E,图4)㊂注:平板拮抗测试AH对稻瘟病菌(A)㊁茎点霉(B)㊁纹枯病菌(C)㊁尖孢镰孢菌(D)㊁层出镰孢菌(E)㊁短小杆菌(F)㊁水稻白叶枯病菌(G)的抑菌活性㊂H.平板模式图㊂Note:TestedpathogensincludedMagnaportheoryzae(A),Phomasp.(B),Rhizoctoniasolani(C),Fusariumoxysporum(D),F.proliferatum(E),Curto⁃bacteriumsp.(F),andXanthomonasoryzaepv.oryzae(G).Experimentalmodeldiagram(H).图3㊀菌株AH对参试菌株的平板拮抗测试Fig.3㊀AntagonisticefficacyofP.putidaAHagainsttestedstrains图4㊀菌株AH的平板抑菌效果Fig.4㊀AntagonisticefficacyofP.putidaAHagainsttestedstrains㊀㊀田间防治试验中,水稻叶片喷施AH菌悬液不影响水稻生长(图5A)㊂仅接种稻瘟孢子液时,叶片出现急性型病斑㊁慢性型病斑㊁褐点和白点型病斑的混合症状(图5B㊁C);接种前用AH菌悬液处理后症状显著减轻㊂水稻白叶枯病菌接种后,叶片呈现黄白色皱缩症状,沿叶脉扩展㊂AH喷施后症状显著减轻㊂且AH处理组的叶片皱缩现象比白叶枯病菌接种组的症状延迟3d出现㊂水稻细菌性条斑病菌接种后,接种部位叶片出现黄褐色的短病斑,AH菌液处理对病害进程无明显影响(图5)㊂说明田间喷施菌株AH悬液可有效减轻水稻白叶枯病和稻瘟病的病害等级㊂2.4㊀菌株AH基因组分析㊀采用二代Illumina与第三代PacBio结合的测序技术对菌株AH进行全基因组测序,获得高质量Reads102114条,数据量1.27G㊂AH基因组长为5889125bp,GC含量为63.74%,包含5215个编码序列,其中包括19个rRNA㊁77个tRNA和78个nRNA㊂基于GC偏差㊁GC含量㊁非编码RNA(rRNA为红色表示,tRNA为蓝色表示,sRNA为绿色表示)和COG注释等信息,使用Circos软件构建的基因组圈图见图6㊂全基因组测序数据已提交至GenBank,收录号为SAMN34209943㊂利用NR数据库进行蛋白质序列相似性比对显示,共有5183个基因被注释,占基因总数的99.4%;在KEGG数据库中比对得到5150个基因,占比98.8%;在COG㊁Swiss⁃Prot和GO数据库中分别比对得到4164㊁3858㊁3715个基因,占比分别为79.8%㊁74.0%和71.2%㊂㊀㊀对菌株AH基因组数据进行KEGG和GO数据库分析,结果见图7㊂KEGG注释结果表明,菌株AH基因组中有2810个编码蛋白的基因富集到六大类42条代谢通路中㊂其中,代谢功能相关的基因占比最大,其中氨基酸代谢㊁碳水化合物代谢㊁辅酶因子和维生素代谢㊁能量代谢相关通路的14151卷20期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀胡逸群等㊀拮抗水稻病原菌假单胞菌AH菌株的鉴定及全基因组序列分析数量较高㊂表明菌株AH具有代谢多样性的特征㊂GO数据库按照生物过程㊁细胞组分和分子功能三类对菌株AH基因功能进行分类,其中细胞组分中膜组分相关基因㊁分子功能中的ATP结合和金属离子结合相关基因占比较大㊂注:稻瘟病菌(上),水稻细菌性条斑病菌(中)和白叶枯病菌(下)在感病水稻品种WH26上的发病情况㊂Note:Theseveritiesofriceblast(upper),bacterialleafstreak(middle),andbacterialblight(lower)inthesusceptiblericevarietyWH26wereexhibi⁃ted.图5㊀菌株AH对水稻病害的防治效果Fig.5㊀Thebiocontroleffectagainstthreemajorpathogensonriceplants2.5㊀次生代谢产物合成基因簇的预测㊀使用antiSMASH在线平台对菌株AH基因组中次级代谢产物合成基因簇进行预测,分析表明该基因组包含8个次级代谢产物基因簇,其中5个是非核糖体肽合成酶(NRPS)基因簇或NRPS-like基因簇(图8㊁表2)㊂根据基因簇同源性,AH可能合成2类已知的抗菌物质viscosin和putisolvin㊂分析发现,Cluster1与BGC0001100来源的Streptomycesrochei的合成基因簇相似性为13%;Cluster2和Cluster5都与BGC0000413来源的Pseudo⁃241㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀安徽农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀2023年monasprotegensPf-5的pyoverdin合成基因簇相似,相似度分别为10%和4%;Cluster3与BGC0001312来源的PseudomonasfluorescensSBW25的viscosin合成基因簇相似度为50%㊂Cluster4与Pseudomonasputida的putisolvin合成基因簇相似性为50%;Cluster5与BGC0000413来源的Pseudomonasprote⁃gensPf-5的pyoverdin合成基因簇相似性为4%;Cluster8与BGC0001692来源的XenorhabdusnematophilaATCC19061的nematophin合成基因簇相似性为12%㊂另外Cluster6和Cluster7未比对到相似基因簇,暗示菌株AH中可能存在新物质合成基因簇,具有研究和应用潜力㊂注:Circos(v0.69)软件绘制菌株AH的基因组圈图,从内到外分别代表GC偏移㊁GC含量㊁非编码RNA(红色是rRNA,蓝色是tRNA,绿色是sR⁃NA)㊁前导链的COG注释和后随链的COG注释㊂Note:ThemapwasdrawnusingCircos(v0.69).Fromthecentertotheperiphery,differentcomponentsrepresentGCskew,GCcontent,non⁃codingRNA(rRNA,red;tRNA,blue;sRNA,green),COGannotationoftheleadingstrand,andCOGannotationofthelaggingstrand.图6㊀菌株AH的基因组圈图Fig.6㊀GraphicalcircularmapofthegenomeofP.putidaAH2.6㊀菌株AH对氨基糖苷类抗生素高度敏感㊀采用平板加药法对菌株AH的抗生素敏感性进行测定发现,外源卡那霉素或庆大霉素浓度为2.5μg/mL的条件下,菌株AH生长出现明显抑制现象;利福平浓度为10μg/mL时,AH生长被抑制,20μg/mL时抑制作用明显;在所有测试浓度下,氨苄霉素均未对AH生长产生影响(图9)㊂这表明AH菌株对氨基糖苷类抗生素(庆大霉素和卡那霉素)高度敏感,且抑菌作用存在剂量依赖性㊂3㊀结论与讨论该研究通过平板抑菌测试,筛选得到5株具有生防效果的菌株,其中菌株AH效果最好,经多基因测序比对,鉴定为恶臭假单胞P.putidaAH㊂进一步测试发现AH对植物病原细菌水稻白叶枯菌和病原真菌稻瘟病菌㊁纹枯病菌都有较好的抑菌活性,其抑菌谱具有一定的广谱性,具有研究价值㊂在线平台antiSMASH预测发现P.putidaAH中有8个次级代谢产物合成基因簇,可能合成viscosin㊁putisolvin等抗菌物质㊂putisolvin对多种病原菌具有拮抗活性,一般通过破坏细胞膜和抑制DNA合成的机制发挥抗菌作用,也能作为生物表面活性剂影响细菌群体感应与生物膜形成[28]㊂但AH菌株的合成基因簇与已知基因簇比对的相似度来看,均未超过50%,仅有Cluster3与P.fluorescensSBW25的viscosin合成基因簇相似度为50%,Cluster4与P.putida的putisolvin合成基因簇相似性为50%㊂Cluster6和Cluster7未比对到相似基因簇,另外的Cluster1㊁2㊁5㊁8基因簇与已知菌株基因簇的相似度为13%㊁10%㊁4%㊁12%㊂此外,经过比对发现胺甲萘降解菌株P.putidaSimmons01㊁P.putidaNBRC14164菌株有6个次级代谢产物合成基因簇,耐低碳氧菌株P.putidaKT2440有7个合成基因簇[29]㊂这些结果暗示菌株AH的8个基因簇除合成上述已知物质,还可能合成其余未报道的物质,具有代谢多样性的特征,有较好的研究和应用潜力㊂34151卷20期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀胡逸群等㊀拮抗水稻病原菌假单胞菌AH菌株的鉴定及全基因组序列分析注:A.KEGG注释结果;B.GO注释结果㊂Note:A.KEGGannotationresults;B.GOannotationresults.图7㊀菌株AH的基因功能分析Fig.7㊀GenefunctionsintheP.putidaAHgenome441㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀安徽农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀2023年注:AntiSMASH在线平台预测菌株AH的次级代谢产物合成相关基因簇,8个基因簇的信息如图㊂不同颜色代表不同类型的基因㊂Note:SecondarymetaboliteassociatedgeneclusterswerepredictedbyantiSMASHonlineplatform.EightgeneclusterswererespectivelyconcatenatedonP.putidaAHgenomeaccordingtotheorderandlength.Differentcolorsrepresentdifferenttypesofgenesasshown.图8㊀菌株AH的8个次级代谢产物合成基因簇Fig.8㊀SecondarymetabolismgeneclustersinP.putidaAH表2㊀菌株AH的8个次级代谢产物合成基因簇信息Table2㊀DetailinformationofeightsecondarymetabolicgeneclusterspredictedbyantiSMASHonlineplatform区域Region类型Type起始位置From终止位置To已知相似基因簇Mostsimilarknowncluster相似度Similarityʊ%Region1redox⁃cofacter432762454909lankacidinC,NRP+Polyketide13Region2NRPS23660232450206lankacidinC,NRP10Region3NRPS29018672964859viscosin,NRP50Region4RiPP⁃like,NRPS30037943050744putisolvin,NRP50Region5NRPS39933334046286pyoverdine,NRP4Region6NAGGN42863814301182 Region7Hserlactone,butyrolactone52087815229401Region8NRPS-like57325305775961nematophin,NRP12图9㊀菌株AH的抗生素敏感性测试Fig.9㊀AntibioticsusceptibilityofP.putidaAH㊀㊀菌株AH的抗生素敏感性测试结果表明其对氨苄霉素的耐药性很强,而分析AH菌株的基因组数据发现,其中并不含有编码氨苄霉素抗性的基因,但鉴定得到多个resist⁃ance⁃nodulationagainsttestedstrainscelldivision(RND)类型的抗生素外排泵基因,这表明菌株AH可能是通过外排泵系统来阻止氨苄霉素的外源施加对细菌造成的损伤㊂除RND类54151卷20期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀胡逸群等㊀拮抗水稻病原菌假单胞菌AH菌株的鉴定及全基因组序列分析外排泵外,AH基因组中还鉴定到smallmultidrugresistance(SMR)类的外排泵,靶向氨基糖苷类抗生素㊂而抗生素敏感性测试表明菌株AH对氨基糖苷类抗生素高度敏感,推测可能是由于相关基因表达水平较低或发生沉默而无法发挥蛋白正常功能,导致菌株AH不足以防御氨基糖苷类抗生素㊂菌株AH的全基因组数据为后续分子操纵提供了遗传信息,也为深层的防病机制解析提供了基础㊂参考文献[1]BIESSYA,FILIONM.PhenazinesinplantagainsttestedstrainsbeneficialPseudomonasspp.:Biosynthesis,regulation,functionandgenomics[J].En⁃vironmentalmicrobiology,2018,20(11):3905-3917.[2]FIRAD,DIMKICᶄI,BERICᶄT,etal.BiologicalcontrolofplantpathogensbyBacillusspecies[J].Journalofbiotechnology,2018,285:44-55.[3]YINN,ZHAOJL,LIUR,etal.BiocontrolefficacyofBacilluscereusstrainBc⁃cm103againstMeloidogyneincognita[J].Plantdisease,2021,105(8):2061-2070.[4]BALTHAZARC,NOVINSCAKA,CANTING,etal.BiocontrolactivityofBacillusspp.andPseudomonasspp.againstBotrytiscinereaandothercan⁃nabisfungalpathogens[J].Phytopathology,2022,112(3):549-560.[5]BONNICHSENL,SVENNINGSENNB,RYBTKEM,etal.Lipopeptidebio⁃surfactantviscosinenhancesdispersalofPseudomonasfluorescensSBW25biofilms[J].Microbiology(Reading),2015,161(12):2289-2297.[6]JUNSR,WASSENAARTM,NOOKAEWI,etal.DiversityofPseudo⁃monasgenomes,includingpopulus⁃associatedisolates,asrevealedbycom⁃parativegenomeanalysis[J].Appliedandenvironmentalmicrobiology,2015,82(1):375-383.[7]NANDIM,SELINC,BRASSINGAAKC,etal.PyrrolnitrinandhydrogencyanideproductionbyPseudomonaschlororaphisstrainPA23exhibitsne⁃maticidalandrepellentactivityagainstCaenorhabditiselegans[J].PLoSOne,2015,10(4):1-19.[8]MISHRAJ,ARORANK.Secondarymetabolitesoffluorescentpseudo⁃monadsinbiocontrolofphytopathogensforsustainableagriculture[J].Ap⁃pliedsoilecology,2018,125:35-45.[9]PARVINW,GOVENDERN,OTHMANR,etal.PhenazinefromPseudo⁃monasaeruginosaUPMP3inducedthehostresistanceinoilpalm(ElaeisguineensisJacq.)⁃Ganodermaboninensepathosystem[J].Scientificreports,2020,10(1):1-12.[10]ZENGERERV,SCHMIDM,BIERIM,etal.PseudomonasorientalisF9:Apotentantagonistagainstphytopathogenswithphytotoxiceffectintheap⁃pleflower[J].Frontiersinmicrobiology,2018,9:1-13.[11]CHANDRAH,KUMARIP,BISHTR,etal.PlantgrowthpromotingPseud⁃omonasaeruginosafromValerianawallichiidisplaysantagonisticpotentialagainstthreephytopathogenicfungi[J].Molecularbiologyreports,2020,47(8):6015-6026.[12]BLAKNEYAJC,PATTENCL.Aplantgrowth⁃promotingpseudomonadiscloselyrelatedtothePseudomonassyringaecomplexofplantpathogens[J].FEMSmicrobiologyecology,2011,77(3):546-557.[13]PASTORN,MASCIARELLIO,FISCHERS,etal.PotentialofPseudo⁃monasputidaPCI2fortheprotectionoftomatoplantsagainstfungalpathogens[J].Currentmicrobiology,2016,73(3):346-353.[14]KOYJ,KIMJS,KIMKD,etal.Microscopicalobservationofinhibition⁃behaviorsagainstDiaporthecitribypre⁃treatedwithPseudomonasputidastrainTHJ609⁃3ontheleavesofcitrusplants[J].Journalofmicrobiology,2014,52(10):879-883.[15]HEY,PANTIGOSOHA,WUZ,etal.Co⁃inoculationofBacillussp.andPseudomonasputidaatdifferentdevelopmentstagesactsasabiostimulanttopromotegrowth,yieldandnutrientuptakeoftomato[J].Journalofap⁃pliedmicrobiology,2019,127(1):196-207.[16]DAURA⁃PICHO,HERNÁNDEZI,PINYOL⁃ESCALAL,etal.Noantibi⁃oticandtoxicmetabolitesproducedbythebiocontrolagentPseudomonasputidastrainB2017[J].FEMSmicrobiologyletters,2020,367(9):1-9.[17]KANDASWAMYR,RAMASAMYMK,PALANIVELR,etal.ImpactofPseudomonasputidaRRF3ontheroottranscriptomeofriceplants:In⁃sightsintodefenseresponse,secondarymetabolismandrootexudation[J].Journalofbiosciences,2019,44(4):98.[18]GUPTAL,VERMANIM,KAURAHLUWALIASK,etal.Molecularviru⁃lencedeterminantsofMagnaportheoryzae:Diseasepathogenesisandre⁃centinterventionsfordiseasemanagementinriceplant[J].Mycology,2021,12(3):174-187.[19]CENYK,LINJG,WANGYL,etal.ThegibberellinproducerFusariumfujikuroi:Methodsandtechnologiesinthecurrenttoolkit[J].Frontiersinbioengineeringandbiotechnology,2020,8:1-17.[20]NAKAYINGAR,MAKUMIA,TUMUHAISEV,etal.Xanthomonasbacte⁃riophages:Areviewoftheirbiologyandbiocontrolapplicationsinagricul⁃ture[J].BMCinmicrobiology,2021,21(1):1-20.[21]JIANGN,YANJ,LIANGY,etal.ResistancegenesandtheirInteractionswithbacterialblight/leafstreakpathogens(Xanthomonasoryzae)inrice(OryzasativaL.)⁃anupdatedreview[J].Rice,2020,13(1):1-12.[22]KUMARA,KUMARR,SENGUPTAD,etal.Deploymentofgeneticandgenomictoolstowardgainingabetterunderstandingofrice-Xanthomonasoryzaepv.oryzaeinteractionsfordevelopmentofdurablebacterialblightresistantrice[J].Frontiersinplantscience,2020,11:1-23.[23]LAMICHHANEJR,BALESTRAGM,VARVAROL.Occurrenceofpota⁃tosoftrotcausedbyErwiniacarotovora(synonymPectobacteriumcaroto⁃vorum)inNepal:Afirstreport[J].Plantdisease,2010,94(3):382.[24]HUNTM,SILVAND,OTTOTD,etal.Circlator:Automatedcirculariza⁃tionofgenomeassembliesusinglongsequencingreads[J].Genomebiolo⁃gy,2015,16(1):1-10.[25]KRZYWINSKIM,SCHEINJ,BIROLI,etal.Circos:Aninformationaes⁃theticforcomparativegenomics[J].Genomeresearch,2009,19(9):1639-1645.[26]HYATTD,CHENGL,LOCASCIOPF,etal.Prodigal:Prokaryoticgenerecognitionandtranslationinitiationsiteidentification[J].BMCbioinfor⁃matics,2010,11(1):1-11.[27]GRIFFITHS⁃JONESS,BATEMANA,MARSHALLM,etal.Rfam:AnRNAfamilydatabase[J].Nucleicacidsresearch,2003,31(1):439-441.[28]CÁRCAMO⁃OYARCEG,LUMJIAKTASEP,KÜMMERLIR,etal.QuorumsensingtriggersthestochasticescapeofindividualcellsfromPseudo⁃monasputidabiofilms[J].Naturecommunications,2015,6:1-9.[29]DEMLINGP,ANKENBAUERA,KLEINB,etal.PseudomonasputidaKT2440endurestemporaryoxygenlimitations[J].Biotechnologybioengi⁃neering,2021,118(12):4735-4750.641㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀安徽农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀2023年。
专题20实验与探究-2023年高考真题和模拟题生物分项汇编(全国通用)(学生版)

专题20 实验与探究2023年高考真题一、单选题1.(2023·北京·统考高考真题)研究者检测了长期注射吗啡的小鼠和注射生理盐水的小鼠伤口愈合情况,结果如图。
由图可以得出的结论是()A.吗啡减缓伤口愈合B.阿片受体促进伤口愈合C.生理条件下体内也有吗啡产生D.阿片受体与吗啡成瘾有关二、综合题2.(2023·北京·统考高考真题)为了研究城市人工光照对节肢动物群落的影响,研究者在城市森林边缘进行了延长光照时间的实验(此实验中人工光源对植物的影响可以忽略;实验期间,天气等环境因素基本稳定)。
实验持续15天:1~5天,无人工光照;6~10天,每日黄昏后和次日太阳升起前人为增加光照时间;11~15天,无人工光照。
在此期间,每日黄昏前特定时间段,通过多个调查点的装置捕获节肢动物,按食性将其归入三种生态功能团,即植食动物(如蛾类幼虫)、肉食动物(如蜘蛛)和腐食动物(如蚂蚁),结果如图。
(1)动物捕获量直接反映动物的活跃程度。
本研究说明人为增加光照时间会影响节肢动物的活跃程度,依据是:与1~5、11~15天相比,______________。
(2)光是生态系统中的非生物成分。
在本研究中,人工光照最可能作为___________对节肢动物产生影响,从而在生态系统中发挥作用。
(3)增加人工光照会对生物群落结构产生多方面的影响,如:肉食动物在黄昏前活动加强,有限的食物资源导致___________加剧;群落空间结构在___________两个维度发生改变。
(4)有人认为本实验只需进行10天研究即可,没有必要收集11~15天的数据。
相比于10天方案,15天方案除了增加对照组数量以降低随机因素影响外,另一个主要优点是________________。
(5)城市是人类构筑的大型聚集地,在进行城市小型绿地生态景观设计时应__________。
A.不仅满足市民的审美需求,还需考虑对其他生物的影响B.设置严密围栏,防止动物进入和植物扩散C.以整体和平衡的观点进行设计,追求生态系统的可持续发展D.选择长时间景观照明光源时,以有利于植物生长作为唯一标准3.(2023·北京·统考高考真题)自然界中不同微生物之间存在着复杂的相互作用。
世界一流科技期刊文章精选
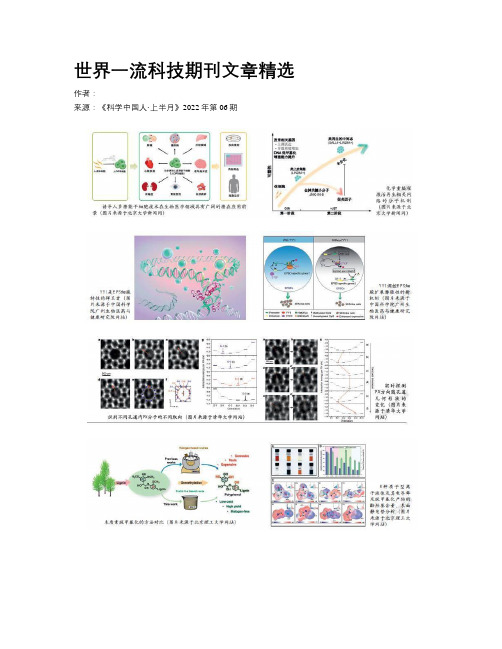
世界一流科技期刊文章精选作者:来源:《科学中国人·上半月》2022年第06期泡泡铸造软体机器人技术《自然》封面:人造肌肉和夹具。
《自然》杂志第7884期封面文章报道了泡泡铸造软体机器人新技术。
软体机器人能执行多种复杂任务,包括抓握、爬行或游泳等,近年来泡泡铸造软体机器人受到学术界的普遍关注。
但是,制造此类软体致动器一直较为烦琐。
科学家们开发了一种基于泡泡的方法,能用来制造各种基于弹性聚合物的致动器。
他们先将液态弹性体注入模具,随后注入空气,从而得到一个拉长的泡泡。
这个泡泡会在重力作用下上升,并形成致动器的内部腔室,通过改变模板和流动过程,就能得到各种各样的复杂结构。
心咽部结构拆解与原始被囊动物的固着生活方式《自然》封面:“住屋”是一类生物特有的一种胶质滤食结构。
《自然》杂志第7885期封面文章报道了被囊动物的演化研究。
被囊动物(如海鞘)与脊椎动物的亲缘关系最近。
这类海洋生物大部分在幼体时能自由游动,但在变成定栖的成体后会固着、无法移动。
然而,尾海鞘纲的被囊动物不会发生这种转变,终生都能自由游动。
研究团队聚焦尾海鞘纲的心脏发育,发现它们的心咽部基因调控网络因大规模基因丢失而“拆解”。
这种拆解导致其失去了原始被囊动物特征性的固着生活方式。
这或许与“住屋”这类帮助尾海鞘纲从固着生活方式演化适应有关。
山岳地形对北美季风的机械强迫作用《自然》封面:一场季风暴雨穿过亚利桑那州东南部时的一道闪电。
《自然》杂志第7886期封面文章报道了北美季风的差异。
在夏季,一个强降雨帶沿墨西哥西海岸延伸至美国西南部、长达一千多公里,构成了北美季风。
一般认为季风是太阳对海洋和陆地加热不均所致。
这会导致气压变化,最终形成的风会拖动凉爽潮湿的海洋空气在陆地上产生降雨。
研究表明,墨西哥的马德雷山脉令急流转向,抬升了暖湿空气,形成对流型降雨。
虽然地面加热也会发生,但其影响不足,北美季风应该是在机械强迫作用下形成的。
人类直觉与AI推动数学进步《自然》封面:人工智能引导的直觉。
毒氟磷——“植物癌症”防控良方

病毒病被称为“植物癌症”,水稻一旦感染,轻则大幅减产,重则绝收。
荣获2014年度国家科技进步二等奖的防治农作物病毒病及媒介昆虫新农药研制与应用项目,为这一顽症开出了治病良方,实现了南方水稻黑条矮缩病的有效防控。
业内专家表示,抗水稻病毒病药剂创制、水稻病毒病及其媒介昆虫全程免疫防控技术处于国际领先水平,为实现全国同步小康战略目标作出了新的贡献。
病毒病一直被称为“植物癌症”,南方水稻黑条矮缩病是其中一种。
水稻黑条矮缩病主要是由白背飞虱传毒,由于其发生、侵染及传播机制极其复杂,一直缺乏有效的防控药剂和技术。
植物病毒病给农业生产带来了极为严重且相当广泛的危害,被誉为农业生产中的世界性难题。
十几年前,化学防治是控制农作物病虫害的常用措施,但传统化学农药的长期不合理使用,会导致病虫抗性增强。
同时,传统农药使用所带来的环境污染、生态破坏以及农产品中农药残留也会对人体健康造成威胁。
因此,开展农作物主要病虫害新药剂、新剂型研究与应用技术集成,已成为当前农业及植保行业中必须解决和重点技术攻关的科学问题和现实课题。
针对粮食作物重大病毒病、病毒传媒害虫、迁徙性病害,本着以新药剂为主体,进行田间应用示范技术集成和推广,筛选出效果好、成本低、残留低,环境友好,经济、社会和生态效益显著的重大病虫害防控应用技术和规范化施用方法这一原则,以提高我国主要粮食作物重大病虫害的防治水平,保障农业生态环境安全和食品安全为目的,该团队产、学、研、推协同攻关,历时10年,终于在病毒病防控磷系药剂和防控技术两方面均取得了系统性的创新成果。
在药剂开发方面,项目组以绵羊体内活性成分α-氨基膦酸酯为先导,基于免疫诱导激活分子靶标,建立全新化学生物学筛选方法和针对多种植物病毒病的离体、活体筛选模型,以动物中发现的天然氨基膦酸为先导,设计合成了α-氨基膦酸酯等多个系列的上千个新化合物,并从中发现了高效抗植物病毒活性先导结构。
经进一步结构优化,研究人员发现了具有全新结构的磷系抗植物病毒新农药——毒氟磷,历经长达8年的卫生毒理学、合成工艺、分析方法、田间药效、环境毒理学、环境代谢行为、残留试验等系统研究,在国际上首次发现其激活植物免疫系统的作用机制,提出了针对植物抗病激活而发现的绿色抗病毒剂的新思路,创建了基于植物抗病激活而发现的抗植物病毒剂的筛选方法。
水稻旱青立病发生原因田间试验研究

水稻旱青立病发生原因田间试验研究戚士胜唐桂林*胡凤桂周宗玲陶伟余承良权循东林剑俞林(寿县农业技术推广中心,安徽寿县232200)摘要本试验采用分蘖末期及时烤田和长期淹水2种处理进行水稻大田旱青立病发生原因区组对比试验,2021年8个参试品种,2022年4个参试品种。
结果表明:及时烤田加干干湿湿管理,各品种水稻均未发生旱青立病,且结实率和千粒重明显增加,在防病的同时有一定的增产作用;长期淹水不烤田条件下水稻易发生旱青立病,且成熟后谷粒中有机砷含量也较高,其发病原因可能是植株有机砷含量增高。
不同品种之间抗性差异明显,荃优822和皖稻68较抗病。
淹水处理的水稻孕穗中后期植株地上部分铜、锌含量极显著降低,铁含量极显著增加,硫、镁、硅无明显变化。
因此,可以排除铜中毒和缺硅、缺硫等因素导致发病,锌、铁元素含量变化对旱青立病的影响有待进一步研究。
关键词水稻旱青立病;发病原因;发病率;砷含量;中微量元素含量;产量中图分类号S435.111.4文献标识码A文章编号1007-5739(2023)16-0091-06DOI:10.3969/j.issn.1007-5739.2023.16.024开放科学(资源服务)标识码(OSID):旱青立病(又称水稻颖壳不闭合、张嘴瘪、直穗病)是我国长江流域稻区普遍发生的一种水稻生理性病害,在美国、日本、澳大利亚、东南亚国家都有发生。
寿县于1962年报道发现该病,并在20世纪80年代后连年发生,其中2006年、2007年、2009年、2014年及2020年均大发生,给水稻生产带来很大危害[1]。
其致病原因目前尚不明确,国内学者有的认为该病是除草剂及植物激素造成的;有的认为是土壤养分不平衡及缺素造成的;有的认为是旱改水造成的砷中毒,但砷中毒机制不清楚;有学者认为是药害;也有认为是高温造成的;还有认为是污水灌溉造成的[2]。
国外通过盆栽试验和在大田土壤或水中添加有机砷试验,证明致病原因是有机砷中毒[3],但土壤中有机砷是如何形成和降解的以及水稻如何吸收及代谢尚不清楚。
水稻生物安全事故案例

水稻生物安全事故案例水稻作为世界上最重要的粮食作物之一,其生物安全问题备受关注。
下面列举了10个水稻生物安全事故案例,以便更好地了解和防范这些问题。
1. 高蛋白水稻事件:2005年,中国农业科学院发布了一项名为“超级稻”的研究成果,称其在蛋白质含量上有显著提高。
然而,随后的调查发现,这种高蛋白水稻由于基因突变而导致对人类肝脏有害。
这一事件引发了对转基因水稻的争议和质疑。
2. 糯米变异事件:2013年,菲律宾政府宣布,他们在一批从中国进口的糯米样本中发现了一种变异基因,该基因使糯米具有抗草甘膦的能力。
这引发了国际社会对转基因水稻的担忧,因为草甘膦是一种广泛使用的除草剂。
3. 水稻病害扩散:水稻病害是水稻生物安全的主要问题之一。
例如,稻瘟病、稻瘿蚊病、纹枯病等病害的爆发和扩散,给农民的收成带来了严重的损失。
这些病害的扩散往往与环境条件、种植管理和病原体的遗传变异等因素有关。
4. 水稻虫害暴发:水稻虫害是水稻生物安全的另一个重要问题。
例如,稻飞虱、稻纵卷叶螟等虫害的暴发会导致水稻产量的急剧下降。
这些虫害通常与气候变化、农药滥用和转基因水稻的抗虫性有关。
5. 水稻转基因污染事件:转基因水稻的种植和销售在一些国家和地区是被禁止的。
然而,由于种子流通和农民种植管理不善,转基因水稻的污染事件时有发生。
这些事件不仅导致农民的经济损失,还可能对环境和人类健康造成潜在风险。
6. 水稻品种失传事件:随着农业现代化的推进,许多传统水稻品种逐渐被高产、抗病虫的杂交水稻取代。
然而,这种单一的水稻种植模式使得许多传统水稻品种逐渐失传。
这不仅导致了遗传资源的丧失,还可能影响农作物的抗逆性和可持续发展。
7. 水稻生物技术研发滞后:水稻生物技术的研发滞后是水稻生物安全的另一个问题。
相比于其他作物,水稻的基因组复杂,转基因技术的应用较为困难。
这导致了水稻生物技术的进展相对缓慢,限制了其在抗病虫、抗逆性和品质改良等方面的应用。
8. 水稻种质资源管理不善:水稻种质资源是水稻研究和育种的重要基础。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
种业资讯 水稻基因组研究取得重要进展 等
水稻基因组研究取得重要进展
近日,国际基因组研究权威刊物《基因组研究》在线刊登了福建省农科院与厦门大学合作的最新研究成果“全基因组范围内分析水稻选择性腺苷化位点的动态变化”。
该项成果主要研究了水稻发育过程中选择性多聚腺苷化(APA)动态变化以及对水稻产量性状关键基因表达调控的影响,为理解基因表达转录后调控提供崭新的认识,同时也对水稻基因组注释提供更加精确数据,从而可为水稻新品种选育提供重要信息和基因资源。
(福建省科技厅网)耐盐小麦高产栽培试验示范项目通过验收“耐盐小麦在墨玉县高产栽培的试验示范”项目由中国科学院新疆生态与地理研究所承担,其针对和田地区墨玉县土壤盐渍化问题,开展了耐盐小麦“新冬26号”和“新冬34号”高产栽培示范和推广。
项目组以墨玉县加汗巴格乡为核心试验区,建立良种繁育基地,先后在喀尔赛镇、奎牙镇、扎瓦乡、阿克萨拉依乡、萨依巴格乡、芒来乡、阔依其乡、雅瓦乡等地建立了8个推广试验示范区,累计推广面积超过万亩,小麦增收150万公斤。
项目在墨玉县加汗巴格乡乡政府召开了验收会,并组织专家赴麦田进行了现场勘验,专家一致认为,该项目为盐碱地改良利用做出了创新性工作,有效提升了盐碱地生产力,促进了农民增收,希望该项目能够形成系统,从而在和田乃至新疆的盐碱地开展示范推广。
验收会结束后,召开了“核桃林下小麦种植技术示范与推广”项目研讨会,和田地区是新疆核桃的主产区之一,核桃林荫下经济作物的种植一直是困扰农业发展的瓶颈问题。
探索适合核桃林下种植模式的小麦材料以及培育林荫下小麦新品系是当前的主要需求之一,新疆生地所专家任巍已在该乡开展了前期的引种试验工作。
专家认为,下一步需要对筛选出的小麦材料进行进一步的试验论证。
(中国科学院新疆分院网)
中国农业科学院植物保护研究所
揭示水稻程序性细胞死亡新机制
日前,中国农业科学院植物保护研究所王国梁科研团队通过图位克隆方法鉴定到一个负调控水稻程序性细胞死亡的DRP类蛋白(死亡抵抗蛋白),揭示了该蛋白通过调控水稻细胞色素c从线粒体的释放而控制细胞程序性死亡发生的新机制。
相关成果于近日在线发表在国际期刊《科学公共图书馆—病原学》上。
据悉,动植物中普遍存在的程序性细胞死亡(Programmed Cell Death,PCD)是生物体自我调节细胞存亡的典型形式,在生长发育与生物或非生物胁迫过程中起重要作用。
植物中假病斑突变是在没有外来病原物侵染情况下细胞自发形成的程序性细胞死亡,这种遗传学上的突变会带来抗性相关基因的诱导表达和植物对病虫害的抗性增强。
本研究对在水稻组织培养过程中得到的一个水稻假病斑突变体dj-lm进行了遗传、生化和细胞生物学分析。
该突变体从苗期到水稻成熟全生育期均呈假病斑表型,植株的抗病性增强,并伴有明显的活性氧簇产生。
利用图位克隆方法成功克隆了控制该假病斑表型的基因OsDRP1E,发现OsDRP1E的E409V点突变导致细胞死亡。
进一步的功能分析表明,OsDRP1E的E409V突变影响其鸟苷三磷酸酶活性、线粒体定位的丢失和细胞色素c的不正常释放。
本研究首次在水稻中发现细胞色素c从线粒体释放调控程序性细胞死亡的现象,为进一步深入了解DRP类蛋白参与程序性细胞死亡分子机制与水稻抗病反应信号传导奠定了重要基础。
该研究以中国农业科学院植物保护研究所为第一完成单位,湖南农业大学联合培养博士研究生李智强和中国农业科学院植物保护研究所副研究员丁波是文章的共同第一作者,王国梁研究员与丁波副研究员为文章通讯作者,周雪平教授为文章的共同作者。
该研究得到国家自然科学基金面上项目的资助。
(中国科学报)
・35・。