11-核糖体
细胞生物学(11 核糖体)

3 多聚核糖体(polyribosomes)
在蛋白质合成过程中,同一条mRNA分子能够同多个核糖 体结合,同时合成若干条蛋白质多肽链,结合在同一 条mRNA上的核糖体就称为多聚核糖体(polysome 或 polyribosomes
起始复合物形成,蛋白 质的合成随即开始,此 过程称为蛋白质合成的 延伸。延伸涉及四个重 复的步骤∶①氨酰tRNA 进入核糖体的A位点; ②肽键形成;③转位;④ 脱氨酰tRNA释放。上述 四步的循环,使肽链不 断延长。在整个过程中, 需要GTP和一些延长因 子的参与
■ 终止(termination)
■ 原核生物rRNA基因及转录
2 核糖体的装配
人细胞中核糖体装配的主要过程
第三节、核糖体的功能-蛋白质的合成
1、 核糖体的功能位点
A、新掺入的氨酰tRNA (aminoacyl-tRNA )结合的位点 P、肽酰tRNA位点(peptidyl-tRNA site) E、脱氨酰tRNA(deaminoacyl-tRNA) 离开P位点到完全从核糖体释放 出来的一个中间停靠点
NORs in human chromosomes: 13\14\15\21\22
Code for (in eukaryotes): 18s, 28s, 5.8s rRNA
■ 真核生物18S、5.8S、28S rRNA和5SrRNA基因
真核生物有四种rRNA基因, 其中18S、5.8S和28S rRNA 基因是串联在一起的,每个基因被间隔区隔开, 5S rRNA 基因则位于不同染色体上。
第十一章多聚核糖体与蛋白质的合成

第十一章核糖体● 核糖体是细胞质中普遍存在的一种非膜性细胞器,由RNA和蛋白质组成,是细胞内蛋白质合成的场所。
● 多聚核糖体是由多个甚至是几十个核糖体串联在一条mRNA上构成的,能高效的进行肽链的合成。
● 蛋白质合成是以各种氨基酸为原料,mRNA为模板,tRNA 作为“搬运工具”以及核糖体作为“装配机” 合成肽链的过程。
● RNA可能是生命起源中最早的生物大分子。
关键词:核糖体;多聚核糖体;蛋白质合成第二节多聚核糖体与蛋白质的合成核糖体(ribosome)是合成蛋白质的细胞器,其功能是以mRNA为模板,以氨基酸为原料高效且精确地合成蛋白质多肽链。
在真核细胞中,核糖体以多聚核糖体的形式存在能高效的进行肽链的合成。
一、多聚核糖体核糖体往往并不是单个独立地执行功能,而是由多个核糖体串连在一条mRNA 分子上高效地进行肽键的合成。
这种具有特殊功能与形态的核糖体与mRNA的聚合体称为多聚核糖体(polyribosome)。
图11-2-1多聚核糖体二、蛋白质的合成蛋白质合成是以各种氨基酸为原料,mRNA为模板,tRNA 作为“搬运工具”以及核糖体作为“装配机” 合成肽链的过程。
原核细胞蛋白质合成的过程已比较清楚,包括3个阶段:肽链合成的起始,延伸和终止。
在起始之前还要进行氨基酸的活化(一)氨基酸的活化1. 定义氨基酸的活化是指各种参加蛋白质合成的AA与携带它的相应的tRNA结合成氨酰- tRNA的过程。
活化反应在氨酰-tRNA 合成酶的催化下进行。
2.过程活化反应分两步进行:活化:AA-AMP-E复合物的形成转移:氨酰-tRNA形成20种氨基酸中每一种都有各自特异的氨酰-tRNA合成酶。
氨酰-tRNA合成酶具有高度的专一性,它既能识别相应的氨基酸(L-构型),又能识别与此氨基酸相对应的一个或多个tRNA 分子;即使AA识别出现错误,此酶具有水解功能,可以将其水解掉。
这种高度的专一性保证了氨基酸与其特定的tRNA准确匹配,从而使蛋白质的合成具有一定的保真性。
细胞生物学-11_真题-无答案

细胞生物学-11(总分100,考试时间90分钟)一、判断题1. 核糖体存在于一切细胞内。
A. 正确B. 错误2. 在真核细胞内,除5S rRNA外,所有的核糖体rRNA都是在核仁区合成的。
A. 正确B. 错误3. 直径约为15nm的核糖体的亚基是通过直径为9nm的核孔运输到胞质中的。
A. 正确B. 错误4. 有丝分裂前期,核纤层蛋白去磷酸化使核膜破裂。
A. 正确B. 错误5. 细胞在有充足营养条件下,从G1期进入M期。
A. 正确B. 错误6. 酵母细胞“DNA复制是否完成”控制点位于G2期,若DNA复制未完成,则细胞能进入G2期,但不能完成G2期的事件。
A. 正确B. 错误7. 中心粒和基体均不具有自我复制性质。
A. 正确B. 错误8. 细胞周期中,仅在G1/S有检验点(checkpoint)。
A. 正确B. 错误9. 有丝分裂后期开始,所有姐妹染色单体同时分裂,各单体分别向两极移动。
A. 正确B. 错误二、填空题1. 分泌性蛋白质首先在______合成,当肽链延长到80个氨基酸左右时,信号肽与______结合,肽链延长暂时停止。
2. 核糖体中各组成成分是以______键结合在一起的,其组装方式称为______。
3. 在多肽合成过程中,合成一条多肽链所需要的时间主要取决于______,对不同相对分子质量的多肽来说,单位时间内所合成的各种多肽的分子数目______。
4. 细胞周期可以通过某些方式同步化,最重要的是人工细胞周期同步方法包括______和______。
5. 植物细胞没有______和星体。
其纺锤体称为无星纺锤体,后期分裂时会形成______。
6. 细胞松弛素B处理对动物细胞有丝分裂的影响是胞质______,结果形成______细胞。
7. 测定细胞周期各期的时间,可采用3H-TdR脉冲标记法,利用放射自显影方法,统计______占总细胞的百分数加以测定。
8. 几乎所有细胞的染色质在有丝分裂期间都是可见的,它们通过螺旋化形成高度聚缩的结构,称为______;它们在有丝分裂中期的特征,被称为______。
高一生物必修1(1-3章)试卷

高一生物试卷一.选择题:(1—30每题1分,31—40每题2分,只有一个选项正确,共计50分)1.细胞内含量最多的有机化合物和无机化合物分别是()A.糖类、水B.脂质、无机盐C.蛋白质、水D.蛋白质、纤维素2.下列属于群落层次的是()A.一个池塘中全部的草鱼B.一片枫叶林C.一个被杂菌污染的培养皿上的全部生物D.一个草原上全部的动植物3.酸奶中的一个乳酸菌属于生命系统的哪个层次()A.细胞B.器官C.种群D.生态系统4.淀粉、脂肪、胰岛素和DNA共有的化学元素是()A.C、H、O B.C、H、O、NC.C、H、O、N、P D.C、H、O、N、P、S5.下列四组生物中,都属于原核生物的一组是()A.病毒和青霉菌 B.破伤风杆菌和蓝藻C.衣藻和变形虫 D.草履虫和酵母菌6.所有的原核细胞都具有()A.核糖体和线粒体B.细胞膜和叶绿体C.内质网和中心体D.细胞膜和核糖体7.下列叙述与细胞学说不符的是()A.细胞是一个有机体,一切动植物都由细胞发育而来,并由细胞和细胞产物所构成B.细胞是一个相对独立的单位,既有它自己的生命,又对其他细胞共同组成的整体的生命起作用C.所有生物体都是由细胞构成的D.新细胞可以从老细胞中产生8.下列选项中,属于动植物细胞共有的糖类是()A. 葡萄糖、核糖、脱氧核糖B. 葡萄糖、淀粉、果糖C. 淀粉、脱氧核糖、乳糖D. 麦芽糖、果糖、乳糖9.谷物中含量丰富的多糖是()A.糖原和纤维素B.淀粉和糖原C.淀粉和纤维素D.蔗糖和淀粉10.2007年3月22日是第十五届“世界水日”,3月22-28日是第二十届“中国水周”联合国确定今年“世界水日”的宣传主题是“应对水短缺”。
以下有关水的说法错误的是()A.自由水是良好溶剂B.衰老细胞水分减少C.能参与生物化学反应,维持生物体生命活动D.新陈代谢旺盛时,结合水与自由水的比值升高11.已知一种氨基酸的R基是—CH2—OH,在这种氨基酸分子中,含有C、H、O的原子数分别是()A.3、4、7 B.3、7、3 C.7、1、4 D.4、3、712.下列用于鉴定组织中是否有脂肪存在的试剂及其颜色反应是()A.碘液、蓝色B.斐林试剂、砖红色C.苏丹Ⅲ染液、橘黄色D.双缩脲试剂、紫色13.下列关于实验操作步骤的叙述中,正确的是()A.用于鉴定还原糖的斐林试剂甲液和乙液,可直接用于蛋白质的鉴定B.脂肪的鉴定实验中需用显微镜才能看到被染成橙黄色的脂肪滴C.鉴定可溶性还原糖时,要加入斐林试剂甲液后再加入乙液D.用于鉴定蛋白质的双缩脲试剂A液和B液,要混合均匀,再加入含蛋白质的试管中,且现配现用14.病毒、蓝藻和酵母菌都具有的物质或结构是()A.细胞壁 B.细胞质 C.细胞膜 D.核酸15.下列物质中有的是氨基酸,有的不是,以下所有的氨基酸分子中,经缩合反应形成的物质可能是()①NH2—CH2—COOH ②NH2—CH2—CH2OH③NH2—CH (C00H)—(CH2)2—COOH ④ NH2—CH2—CH2—C00HA.一肽B.二肽C.三肽D.四肽16.电子显微镜下观察到的细微结构属于()A.低倍结构B.高倍结构C.显微结构D.亚显微结构17.观察DNA和RNA在细胞中的分布,运用的原理是()A.单独利用甲基绿对细胞染色可显示DNA在细胞中的分布,从而推知RNA分布B.单独利用吡罗红对细胞染色可显示RNA在细胞中的分布,从而推知DNA分布C.利用甲基绿和吡罗红混合染色剂对细胞染色,同时显示DNA和RNA在细胞中分布D.在细胞核、细胞质内可以分别提取到DNA和RNA18.生物体内的蛋白质种类繁多、千差万别,其原因不可能的是()A.组成肽键的化学元素不同B.组成蛋白质的氨基酸种类、数量不同C.氨基酸排列顺序不同D.肽链的空间结构不同19.科学家常用哺乳动物成熟红细胞作材料来研究细胞膜的组成,是因为()A.哺乳动物成熟红细胞容易得到B.哺乳动物成熟红细胞内没有核膜、线粒体膜等膜结构C.哺乳动物成熟红细胞在水中容易涨破D.哺乳动物成熟红细胞的细胞膜在光学显微镜下容易观察到20.在人体内能将白细胞吞噬的病菌杀死和消化分解的结构是()A.核糖体B.线粒体C.中心体D.溶酶体21.已知Mn是许多酶的活化剂,例如能激活硝酸还原酶。
第11章 核糖体

15
E.coli核糖体小亚单位中rRNA与r蛋白的相互关系示意图 线条表示相互作用及作用力的强(粗线)与弱(细线) (引自Alberts et al,1989)
16
16SrRNA的一级结构是非常保守的 16SrRNA的二级结构具有更高的保守性:臂环结构(stemloop structure) rRNA臂环结构的三级结构模型
100s
120s
10
对核糖体结构进行精细分析的必
要手段是:获得高质量的核糖体晶
二、核糖体的结构
体,然后进行X射线衍射分析
1、对核糖体的X射线衍射图谱分析表明:
每个核糖体有供tRNA结合的3个位点(A、P、E),横跨大小亚
单位的结合面;
11
1、对核糖体的X射线衍射图谱分析表明:
每个核糖体有供tRNA结合的3个位点(A、P、E),横跨大小亚
化的rRNA进行核糖体的重组装,显示核糖体中r蛋白与 rRNA的结构关系。
₪同一生物中不同种类的r蛋白的一级结构均不相同 ₪不同生物同一种类r蛋白之间具有很高的同源性,并在进化上
非常保守。
₪蛋白质结合到rRNA上具有先后层次性。 ₪核糖体的重组装是自我装配过程
13
70S核糖体的小亚单位中rRNA与全部的r蛋白关系的空间模型 对rRNA,特别是对16SrRNA结构的研究
第十一章 核糖体
第一节 核糖体的类型与结构
第二节
多聚核糖体与蛋白质的合成
1
1953年Robinsin和Brown用电镜在植物细胞中发现; 1955年Palade在动物细胞中观察到;
核糖体是合成蛋白质的细胞器,其唯一的功能是按照mRNA的指令
由氨基酸高效且精确地合成多肽链。
细胞生物学答案10-11

答案:一.名词解释1.细胞核:是遗传信息的储存场所,在这里进行基因复制、转录和转录初产物的加工过程,从而控制着细胞的遗传与代谢活动。
2.核被膜:位于细胞核的最外层,是细胞核与细胞质之间的界膜。
3.核纤层:紧贴内层核膜下,是一层由纤维蛋白构成的网络结构,它与胞质中间丝、核内骨架有密切联系。
4.核孔:内外膜常常在某些部位相互融合形成环状开口,称为核孔。
5.亲核蛋白:是指在细胞质内合成后,需要或能够进入细胞核内发挥功能的一类蛋白质。
6.核定位序列:NLS亲核蛋白一般都含有特殊的氨基酸序列,这些内含的特殊短肽保证了整个蛋白质能够通过核孔复合体被转运到细胞核内。
这段具有“定向”、“定位”作用的序列被命名为核定位序列。
7.染色质:是指间期细胞核呃逆由DNA、组蛋白、非组蛋白及少量RNA组成的线性复合结构,是间期细胞遗传物质存在的形式。
8.染色体:是指细胞在有丝分裂或减数分裂的特定阶段,由染色质聚缩而成的棒状结构。
9.组蛋白:是构成真核生物染色体的基本结构蛋白,富含带正电荷的Arg和Lys 等碱性氨基酸,等电点一般在pH10.0以上,属碱性蛋白质,可以喝酸性 dDNA 紧密结合,而且一般不要求特殊的核苷酸序列。
10.非组蛋白:主要是指与特异DNA序列相结合的蛋白质,所以又称序列特异性DNA结合蛋白。
含有较多的Asp、Glu,带负电荷,属酸性蛋白质。
11.核小体:是一种串珠状结构,由核心颗粒和连接线DNA两部分组成,通过酶消化实验建立。
12.常染色质:是指间期细胞核内染色质纤维折叠压缩程度低,相对处于伸展状态,用碱性染料染色时着色浅的那些染色质。
13.异染色质:是指间期核中,染色质纤维折叠压缩程度高,处于聚缩状态,用碱性染料染色时着色深的那些染色质。
14.结构异染色质:指的是各种类型的细胞中,除复制期以外,在整个细胞周期均处于聚缩状态,DNA组装比在整个细胞周期中基本没有较大变化的异染色质。
15.兼性异染色质:是指在某些细胞类型或一定的发育阶段,原来的常染色质聚缩,并丧失基因转录活性,变为异染色质。
细胞生物学第11章 15章习题

细胞生物学第11章 15章习题细胞生物学第11章-15章习题第一一章核糖体一、名词解释1.核糖体2。
5SrRNA基因SD3序列4多核糖体5 N端规则6蛋白酶体7核酶8肽基转移酶2。
填空1、核糖体是一种的颗粒装结构,其主要成分是和。
2、在细胞内核糖体的存在形式主要包括和。
3.蛋白质合成的初始过程非常复杂,包括一系列催化步骤。
4.每个氨基酸都可以与其对应的tRNA分子偶联形成一个。
5.包括tRNA分子的两个结合位点:P位点,它与连接多肽链延伸的tRNA分子紧密结合;,位点a,与一个氨基酸的tRNA分子结合。
6、催化肽键的形成,一般认为这个催化反应是由核糖体大亚基上的分由…调解。
7、在所有细胞中,都有一种特别的密码子aug,它携带一种特别的氨基酸,即,作为蛋白质合成的起始氨基酸。
核糖体沿着mRNA前进,这需要另一个延伸因子,需要水解。
当核糖体遇到终止代码(,)时,延长结束,核糖体和新合成的多肽被释放。
翻译的最后一步叫做,需要一系列因素。
三、选择题1.核糖体在原核细胞和真核细胞中的沉降系数分别为()a.30s和50sb,40s和60sc 50s,60sd 70s和80s2,核糖体的e位点为()。
a.真核mrna加工位点b.trna离开原核生物核糖体的位点c.核糖体中受ecori限制的位点d.电化学电势驱动转运的位点3、真核起始因子eif-3的作用是()。
a、帮助形成亚单位起始复合物(eIF-3,GTP,met-tRNA,40s)B.帮助亚单位起始复合物(三元复合物,40s)与mRNA 5'端C结合。
如果与40s亚单位结合,防止40s与60s亚单位D结合。
与mRNA 5'端帽结构结合以解锁二级结构4。
以下哪种配合物不是初始反应的产物?()a.gtp+pib、atp+pic。
起始因子D.多肽IV.判断问题1.在trna分子中普遍存在的修饰核苷酸是在掺入trna转录物结合前由标准核苷酸共价修饰而来。
核糖体的结构和功能

核糖体的结构和功能核糖体是细胞负责蛋白质合成的重要分子机器,在这个过程中,它起到了至关重要的作用。
本文将介绍核糖体的结构和功能,解释核糖体的作用原理。
一、核糖体的结构核糖体是由核糖核酸、核糖蛋白和其他蛋白质组成的复合物,它们配合着完成蛋白质的合成。
核糖体的组成包括大剂量细胞组分,约占细胞整体重量的40%~50%,它的直径为15至20纳米。
核糖体主要由两个亚单位组成,一个是大的亚单位和一个小的亚单位,它们协作完成蛋白质的合成。
在人类体内,大型亚单位由约50种蛋白质和三个不同的rRNA成分组成,而小型亚单位则由约30种蛋白质和一个rRNA成分组成。
二、核糖体的功能核糖体在生命过程中具有多种功能。
它的主要功能是蛋白质合成,在这个过程中,核糖体通过识别和翻译mRNA中的基因序列,将其翻译成相应的氨基酸序列,组成蛋白质。
具体地,核糖体的功能可以勾画为以下的步骤:1. RNA转运:在核糖体开始转录mRNA之前,mRNA必须先进入细胞质,并被tRNA(转运RNA)伴侣所携带。
由于tRNA 中含有一个特定的氨基酸,因此该分子的翻译是由氨基酸的可用性和核糖体的位置决定的。
2. 初始结合:大型核糖体亚单位将mRNA和tRNA中的氨基酸一起招募进去。
核糖体的结构使得它为特定mRNA序列提供高亲和力,因此在这个过程中出现错误是非常少的。
3. 翻译:随着新的tRNA分子加入,核糖体不断扩大,以将氨基酸序列拼接起来。
在在这个步骤中,核糖体小亚单位起到了重要的作用。
它扫描mRNA上的密码子,以推进氨基酸的合成。
在这个过程中,每个相应的氨基酸序列都必须与对应的mRNA序列相配。
这就是蛋白质合成的关键所在,它决定了蛋白质的三维结构。
如果核糖体由于某种原因无法完成上述步骤,就会导致蛋白质合成的错误,并可能导致细胞功能受损。
三、结论综上所述,核糖体是一个极其重要的细胞分子机器,对人体正常运转发挥了非常重要的作用。
然而,在人的生命嵌合的过程中,由于糖体的功能所受到环境、基因和其他因素的影响,因此可能会导致内部调节出现问题。
核糖体的结构,分布及其作用

核糖体的结构,分布及其作用
核糖体是一种细胞内质体,其结构和功能在不同生物中有些微差异。
在真核生物中,核糖体主要存在于细胞质中,由大约
80个不同的核糖体蛋白和四种RNA组成,包括核糖体RNA (rRNA)和转运RNA(tRNA)。
在细菌中,核糖体主要位
于细胞质中,并且由约55个蛋白质和三种rRNA组成。
核糖体的结构可分为两个亚基:大亚基和小亚基。
大亚基含有A、P和E三个位点,用于合成多肽链。
小亚基含有转运RNA 结合位点,用于将氨基酸转运到核糖体上。
核糖体的主要作用是参与蛋白质合成。
它通过将mRNA中的
密码子与tRNA中的反密码子互补配对,将氨基酸顺序合成成
多肽链。
此过程包括三个主要步骤:起始、延长和终止。
在起始阶段,核糖体与mRNA和启动转运RNA结合,并选择正确的A位点来起始蛋白质合成。
在延长阶段,核糖体依次
从mRNA上读取密码子信息,并将tRNA上的氨基酸加入到
蛋白质链中,形成多肽链。
在终止阶段,核糖体识别到终止密码子,导致蛋白质链的释放,并使核糖体复原以便进入下一个循环。
总结来说,核糖体通过解码mRNA中的密码子并合成多肽链,参与了蛋白质合成过程。
核糖体在各种细胞中广泛存在,并且在分子结构和功能上表现出一定的差异。
11种鲈形目鱼类的核糖体基因GC含量及其与硬骨鱼类的特征比较

11种鲈形目鱼类的核糖体基因GC含量及其与硬骨鱼类的特征比较司李真;武宝生;孔晓瑜;杨敏;龚理;时伟【摘要】核糖体基因为串联重复多拷贝的基因,包括3个编码基因(18S,5.8S,28S)和两个间隔区ITS1(internal tran-scribed spacer 1)和ITS2(internal transcribed spacer 2).目前,对核糖体基因的相关报道主要集中在个体内不同拷贝间的多态特征,以及其作为分子标记在系统演化关系中的应用,GC含量作为一项非常重要的核苷酸序列指标,而鲜有报道.为了探讨鱼类的核糖体基因GC含量特征以及间隔区是否也存在GC平衡现象,本研究选择了鲈形目(Perciformes)5科11种鱼类5个片段的核糖体基因进行研究,包括尖吻鲈科(Latidae)、射水鱼科(Toxotidae)、军曹鱼科(Rachycentridae)、剑鱼科(Xiphiidae)、鲹科(Carangidae).获得了1651个单克隆序列,通过分析并比较已有的其他硬骨鱼序列片段的GC含量变化特征,结果发现:本研究鱼类的18S的GC含量为52.6%~57.1%(平均54.6%),5.8S为55.6%~58.9%(平均57.4%),28S为64.2%~65.8%(平均64.6%),ITS1为56.5%~73.0%(平均65.0%),ITS2为62.3%~77.5%(平均69.1%).编码区的GC含量相对较保守,变异范围较小,18S和5.8S变化范围明显小于间隔区,28S则位于间隔区的最低值和最高值之间.因此,我们发现硬骨鱼核糖体ITS高于60%的GC含量是该类群的一个特征,并且高GC含量的ITS1和ITS2序列中不存在明显的高GC 富集区,其含量高低的变化与序列长度也没有相关性.本研究11种鱼类的ITS1和ITS2的GC含量在种内的相似性既有大于也有小于种间相同片段的相似性,因此GC平衡现象只存在部分种类中.本研究结果可为鱼类核糖体基因序列特征的进一步研究及利用提供科学依据.%The ribosomal RNA gene (rDNA) cluster consists of multiple units of three coding genes (18S, 5.8S, and 28S) as well as twointernal transcribed spacers (ITS1 and ITS2) separating the coding regions. Thus far, stu-dies on these five fragments mainly focused on the polymorphism of different copies within each individual sam-ple and identification of useful markers for phylogenetic relationship analysis. However, there are limited studies related to the features of GC content, which is a very important characteristic of ribosomal RNA gene. The cha-racteristics of rDNA GC content and whether the GC balance phenomenon exists in the coding regions in fishes were investigated by selecting 11 species from five families of Perciformes, including Latidae, Toxotidae, Rachy-centridae, Xiphiidae, and Carangidae. In all, 1651 monoclones from the five fragments mentioned above were obtained. The GC content features were analyzed based on the sequences from the 11 species or the datasets from other teleostean fishes retrieved from GenBank. The results from the above two analyses were compared. The fol-lowing results were obtained. First, the GC content of 18S, 5.8S, 28S, ITS1, and ITS2 rangedfrom 52.6% to 57.1% (average, 54.6%), 55.6% to 58.9% (average, 57.4%), 64.2% to 65.8% (average, 64.6%), 56.5% to 73.0%(average, 65.0%), and 62.3% to 77.5% (average, 69.1%), respectively. Second, compared with non-coding regions, coding regions were relatively conserved. The GC contentof the coding genes varied in smaller ranges than those of the internal transcribed spacers. The GC contents of 18S and 5.8S were lower than those of ITS1 and ITS2, but that of 28S was between the lowest and highest values of ITS1 and ITS2. Therefore, we found that the GC contentof non-coding regions was higher than 60%, which was a remarkablecharacteristic of these fishes, and no correla-tion was found between fragment length and higher GC content. Further, no obvious G, C, or GC rich block was found in the high-GC-content regions of ITS1 and ITS2 sequences. Third, the similarity of GC content between ITS1 and ITS2 within the same species could be higher or lower than that of the same fragment among different species within the 11 species. Therefore, the GC balance phenomenon is not universal and only exists in species whose intra-species GC content similarity is lower than the inter-species GC content similarity. The results of this study might provide a scientific basis for further studies and facilitate the utilization of the ribosomal gene cha-racteristics of fish.【期刊名称】《中国水产科学》【年(卷),期】2017(024)004【总页数】12页(P657-668)【关键词】编码基因;间隔区;18S;5.8S;ITS1;ITS2【作者】司李真;武宝生;孔晓瑜;杨敏;龚理;时伟【作者单位】中国科学院热带海洋生物资源与生态重点实验室, 广东广州 510301;中国科学院大学, 北京 100049;中国科学院热带海洋生物资源与生态重点实验室, 广东广州 510301;中国科学院大学, 北京 100049;中国科学院热带海洋生物资源与生态重点实验室, 广东广州 510301;中国科学院热带海洋生物资源与生态重点实验室, 广东广州 510301;中国科学院大学, 北京 100049;中国科学院热带海洋生物资源与生态重点实验室, 广东广州 510301;中国科学院大学, 北京 100049;中国科学院热带海洋生物资源与生态重点实验室, 广东广州 510301【正文语种】中文【中图分类】S963核糖体基因在生物中有几十个甚至上万个串联重复单元, 每个单元包括编码区与非编码区,编码区包括18S、5.8S和28S 3个基因, 非编码区则由位于18S与5.8S之间的ITS1及5.8S与28S之间的 ITS2两个内转录间隔区组成。
12 生物化学习题与解析--蛋白质的生物合成

蛋白质的生物合成一、选择题(一) A 型题1 .蛋白质生物合成A .从 mRNA 的 3 ' 端向 5 ' 端进行B .由 N 端向C 端进行C .由 C 端向 N 端进行D .由 28S-tRNA 指导E .由 5S-rRNA 指导2 .蛋白质生物合成的延长阶段不需要A .转肽酶B . GTPC . EF-Tu 、 EF-Ts 、 EFGD . mRNAE . fMet-tRNA fMet3 .有关蛋白质合成的叙述正确的是A .真核生物先靠 S-D 序列使 mRNA 结合核糖体B .真核生物帽子结合蛋白复合物( eIF -4F 复合物)在起始过程中发挥作用C . IF 比 eIF 种类多D .原核生物和真核生物使用不同的起始密码E .原核生物有 TATAAT 作为起始序列,真核生物则是 TATA4 .关于氨基酸密码子的描述错误的是A .密码子有种属特异性,所以不同生物合成不同的蛋白质B .密码子阅读有方向性,从 5 ' 端向 3 ' 端进行C .一种氨基酸可有一组以上的密码子D .一组密码子只代表一种氨基酸E .密码子第 3 位( 3 ' 端)碱基在决定掺入氨基酸的特异性方面重要性较小5 .遗传密码的简并性是A .蛋氨酸密码可作起始密码B .一个密码子可编码多种氨基酸C .多个密码子可编码同一种氨基酸D .密码子与反密码子之间不严格配对E .所有生物可使用同一套密码6 .遗传密码的摆动性正确含义是A .一个密码子可以代表不同的氨基酸B .密码子与反密码子可以任意配对C .一种反密码子能与第三位碱基不同的几种密码子配对D .指核糖体沿着 mRNA 从 5 ' 端向 3 ' 端移动E .热运动所导致的 DNA 双螺旋局部变性7 .一个 tRNA 的反密码子为 5 '- IGC-3 ' ,它可识别的密码是A . GCAB . GCGC . CCGD . ACGE . UCG8 .信号肽识别颗粒( signal recognition particles , SRP )可识别A . RNA 聚合酶B . DNA 聚合酶C .核小体D .分泌蛋白的 N 端序列E .多聚腺苷酸9 .下列关于多聚核糖体( polysome )叙述正确的是A .是一种多顺反子B .是 mRNA 的前体C .是 mRNA 与核糖体小亚基的聚合体D .是核糖体大、小亚基的聚合体E .是一组核糖体与一个 mRNA 不同区段的结合物10 .关于蛋白质生物合成的描述哪一项是错误的A .氨基酸必须活化成活性氨基酸B .氨基酸的羧基被活化C . 20 种编码氨基酸各自有相应的密码D .活化的氨基酸靠相应的 tRNA 搬运到核糖体E . tRNA 的反密码子与 mRNA 上的密码子严格按碱基配对原则结合11 .核糖体结合位点( ribosomal binding site , RBS )A .也称 Pribnow 盒B .在原核生物 mRNA 上C .真核生物转录起点D .由 Meselson-stahl 首先发现E .在 tRNA 分子上12 .翻译延长的进位A .指翻译起始复合物的生成B .肽酰 -tRNA 进入 P 位C .由延长因子 EFG 带领,不需消耗能量D .是下一位氨基酸的氨基酰 -tRNA 进入核糖体的 A 位E .多肽链离开核糖体13 .翻译延长需要A .氨基酰 -tRNA 转移酶B .磷酸化酶C .氨基酸合成酶D .肽链聚合酶E .转肽酶14 .蛋白质生物合成中多肽链的氨基酸排列顺序取决于A .相应 tRNA 的专一性B .相应氨基酰 -tRNA 合成酶的专一性C .相应 tRNA 上的反密码D .相应 mRNA 中核苷酸排列顺序E .相应 rRNA 的专一性15 .肽链合成终止的原因是A .翻译到达 mRNA 的尽头B .特异的 tRNA 识别终止密码C .释放因子能识别终止密码子并进入 A 位D .终止密码子本身具有酯酶功能,可水解肽酰基与 tRNA 之间的酯键E .终止密码子部位有较大阻力,核糖体无法沿 mRNA 移动16 .蛋白质合成终止时,使多肽链从核糖体上释出的因素是A .终止密码子B .转肽酶的酯酶活性C .释放因子D .核糖体解聚E .延长因子17 .蛋白质合成中,有关肽链延长叙述正确的是A .核糖体向 mRNA5 ' 端移动三个核苷酸距离B .肽酰 -tRNA 转位到核糖体的 A 位C . GTP 水解成 GDP 和 H 3 PO 4 以提供能量D .空载的 tRNA 从 P 位进入 A 位E . ATP 直接供能18 .多聚核糖体中每一核糖体A .从 mRNA 的 3 ' 端向 5 ' 端前进B .可合成多条多肽链C .可合成一条多肽链D .呈解离状态E .可被放线菌酮抑制19 .氨基酸通过下列哪种化学键与 tRNA 结合A .糖苷键B .酯键C .酰胺键D .磷酸酯键E .氢键20 .信号肽的作用是A .保护 N- 端的蛋氨酸残基B .引导分泌性蛋白进入内质网腔C .保护蛋白质不被水解D .维护蛋白质的空间构象E .传递蛋白质之间的信息21 .下列那一项是翻译后加工A .加 5 ' 端帽子结构B .加 3 ' 端 poly ( A )尾C .酶的激活D .酶的变构E .氨基酸残基的糖基化22 .干扰素抑制蛋白质生物合成是因为A .活化蛋白激酶,而使 eIF-2 磷酸化B .抑制肽链延长因子C .阻碍氨基酰 -tRNA 与小亚基结合D .抑制转肽酶E .使核糖体 60S 亚基失活23 .下列哪一种物质抑制氨基酰 -tRNA 与小亚基结合A .土霉素B .氯霉素C .红霉素D .链霉素E .林可霉素24 .哺乳动物细胞中蛋白质生物合成的主要部位在A .细胞核B .线粒体C .核糖体D .高尔基复合体E .核仁25 .靶向输送到细胞核的蛋白多肽链含有特异信号序列,下列叙述错误的是A .多肽链进细胞核定位后不被切除B .位于 N 末端C .不同多肽链的特异信号序列无共同性C .富含赖、精及脯氨酸 E .也称为核定位序列26 .下列哪种物质直接抑制真核生物核糖体转肽酶A .放线菌酮B .四环素C .土霉素D .链霉素和卡那霉素E .利福平27 .氯霉素可抑制原核生物的蛋白质合成,其原因是A .特异性的抑制肽链延长因子( EFT )的活性B .与核糖体的大亚基结合,抑制转肽酶的活性,而阻断翻译延长过程C .活化一种蛋白激酶,从而影响起始因子( IF )磷酸化D .间接活化一种核酸内切酶使 mRNA 降解E .阻碍氨基酰 -tRNA 与核糖体小亚基结合28 .白喉毒素的作用是A .抑制信号肽酶B .与位于内质网膜表面的受体蛋白结合C .使延长因子 -2 ( eEF-2 )发生 ADP 糖基化而失活,阻断多肽链延长D .加速肽酰 -tRNA 从 A 位移到 P 位,造成氨基酸缺失,从而生成无功能的蛋白质E .通过抑制 GTP 和 fMet-tRNA fMet 在小亚基上的结合,抑制蛋白合成的起始29 .出现在蛋白质分子中的氨基酸,下列哪一种没有遗传密码A .色氨酸B .蛋氨酸C .谷胺酰胺D .脯氨酸E .羟脯氨酸30 .在体内,氨基酸合成蛋白质时,其活化方式为A .磷酸化B .与蛋氨酸结合C .生成氨基酰辅酶 AD .生成氨基酰 -tRNAE .与起始因子结合31 .不属于蛋白质合成后加工修饰的过程为A .肽链 N 端修饰B .亚基聚合C .疏水脂链的共价连接D .多肽链折叠为天然构象的蛋白质E .酶的化学修饰(二) B 型题A .进位B .成肽C .转位D .终止E .释放1 .氨基酰 -tRNA 进入核糖 A 位称为2 .肽酰 -tRNA-mRNA 与核糖体位置的相互变更称为3 . P 位上的肽酰基与 A 位上的氨基酰 -tRNA 的氨基形成肽键称为A .链霉素B .氯霉素C .林可霉素D .嘌呤霉素E .白喉毒素4 .对真核及原核生物蛋白质合成都有抑制作用的抗生素是5 .主要抑制真核细胞蛋白质合成的是A .蛋白质 6- 磷酸甘露糖基化B .滞留信号序列C .囊泡D .分泌小泡E .前体形式6 .靶向输送至溶酶体信号是7 .靶向输送至内质网的蛋白质多肽链 C- 端含8 .质膜蛋白质的靶向输送需要A .信号肽B .信号肽酶C .信号肽识别颗粒D .分泌性蛋白E .对接蛋白9 .有碱性 N 端、疏水核心和加工区三个区域的是10 .属于蛋白核酸复合体的是A .肽键B .酯键C .氢键D .磷酸二酯键E .糖苷键11 .核苷酸之间的连接键12 .氨基酸之间的连接键13 .碱基与核糖之间的连接键14 .氨基酸与 tRNA 之间的连接键A . RNase 抑制因子B .干扰素C .嘌呤霉素D .红霉素E .链霉素15 .抑制 RNase 活性的是16 .能诱导合成 2 ' -5 ' 寡聚腺苷酸的是17 .与酪氨酰 - tRNA 结构相似的是(三) X 型题1 .参与蛋白质合成的物质是A . mRNAB . GTPC .转肽酶D .核糖体E .聚合酶2 .翻译后加工包括A .剪切B .共价修饰C .亚基聚合D .加入辅基E .水解修饰3 .蛋白质合成后可靶向运输到A .留在胞液B .线粒体C .细胞核内D .内质网E .溶酶体4 .引起读码错误的抗生素有A .巴龙霉素B .链霉素C .潮霉素 BD .新霉素E .嘌呤霉素5 .真核生物的 hnRNA 要具有模板作用,必须进行A .剪接B .首尾修饰C .插入稀有碱基D .切除内含子E .碱基甲基化6 .关于 S-D 序列的叙述,正确的是A .也称核糖体结合位点B .与 16S rRNA 3 ' 端 --UCCUCC-- 互补C .碱基序列 --AGGAGG-- 为核心D .位于起始密码上游E .即起始序列7 .翻译的准确性与下列哪些因素有关A .氨基酰 -tRNA 合成酶对底物氨基酸和相应 tRNA 都有高度特异性B .氨基酰 -tRNA 分子中 tRNA 的反密码可通过碱基配对识别 mRNA 分子的遗传密码C .氨基酰 -tRNA 合成酶具有校正活性D .延长因子 EFG 有转肽酶活性E .核糖体对氨基酰 -tRNA 的进位有校正功能8 .关于分子伴侣A .高温应激可诱导该蛋白合成增加B .与分泌性蛋白同在C .能加快多肽链折叠速度D .增加功能性蛋白折叠产率E .可促进需折叠的多肽链折叠为天然构象的蛋白质9 .在蛋白质生物合成中A . 20 种编码氨基酸是原料B . tRNA 携带氨基酸C . mRNA 起模板作用D . rRNA 是合成的场所E .氨基酰 -tRNA 合成酶识别并结合相应的氨基酸和 tRNA10 .干扰素的作用是A .调解细胞生长分化B .激活免疫系统C .抗病毒D .间接诱导核酸内切酶E .诱导使 eIF-2 磷酸化的蛋白激酶活化11 .关于蛋白质二硫键异构酶A .主要在内质网发挥作用B .促二硫键变构C .胞液中活性高D .催化错配的二硫键断裂并形成正确二硫键E .促蛋白质变性二、是非题1 .蛋白质生物合成所需的能量都由 ATP 直接供给。
第11章 核糖体与核酶

E.coli (a)核糖体小亚单位中的部分r蛋白与rRNA的结合位点) (b)及其在小亚单位上的部位
(三)核糖体的装配
• 1、rRNA的转录与加工
• (1)真核生物:18S、5.8S和28S基因是串联一起, 每个基因被间隔区隔开。该基因在RNA聚合酶Ⅰ 作用下,首先转录成一个45S的前rRNA,然后 被加工成41S前rRNA,再被切割成20S前rRNA 和32S前rRNA,20S rRNA加工为成熟的18S rRNA,32S rRNA加工为28S和5.8S rRNA • 5SrRNA基因位于不同染色体上,由RNA聚合酶Ⅲ 转录,仅进行简单加工或不加工
原核生物与真核生物核糖体成分的比较
二、核糖体的结构
(一)核糖体结构与功能的分析方法
离子交换树脂可分离纯化各种r蛋白; 纯化的r蛋白与纯化的rRNA进行核糖体的重组装, 显示核糖体中r蛋白与rRNA的结构关系 双向电泳技术可显示出E.coli核糖体在装配各阶段中, 与rRNA结合的蛋白质的类型 双功能的交联剂和双向电泳分离可用于研究r蛋白在 结构上的相互关系 电镜负染色与免疫标记技术结合,研究r蛋白在核糖 体的亚单位上的定位。 70S核糖体小亚基16S rRNA结构及与全部r蛋白关系
(二)蛋白质合成的基本过程
1、链的起始: • (1)30S亚基与mRNA的结合:在原核生物, mRNA依靠SD序列只和游离的30S小亚基结合; 真核生物,小亚基识别mRNA5’端甲基化帽结构, 然后沿mRNA滑动,直到遇到识别序列,典型的 识别序列为5’-CCACCAUGC-3’,含AUG • 小亚基与mRNA结合还需要起始因子(initiation factor, IF)的帮助.原核生物命名为IF,有IF1,IF2和 IF3,IF3帮助mRNA与核糖体结合;真核生物为eIF, 至少有10个,eIF4帮助mRNA与核糖体结合.
11-14核糖体

~49种蛋白质
A
18S RNA
~33种蛋白质
5S RNA
5S RNA
第三节 核糖体的生物发生与自组装
真核细胞核糖体rRNA几乎都是在染色体的核仁 组织区(nucleolus organizer region, NOR) 内转录合 成的。
18S rRNA+ 33r蛋白质
•rDNA
45S rRNA
(前体分子)
+Mg++ —Mg++
二聚体 120S
核糖体以游离的形式存在于细胞质中,以可附着在内质网上。
第二节 核糖体的基本类型与 化学组成
一、核糖体的基本类型 原核细胞核糖体
类 种 真核细胞质核糖体
叶绿体核糖体
真核细胞器核糖体
线粒体核糖体
二、核糖体的化学组成
核糖体的类型 原核细胞核糖体 真核细胞核糖体 真核细胞器核糖体
Termination
终止包括两步:
1、RF与mRNA终止信号结合,激活转肽酶; 2、水解肽链和 tRNA间的键,新合成肽链、tRNA、
mRNA离开核糖体,后者即解离成大、小亚基, 进入下一轮反应。
(三)合成的蛋白质类型
结构蛋白: 又称内源性蛋白(endogenous protein),主 要由游离核糖体合成,多分布细胞基质中, 是用于细胞本身或参与组成细胞自身结构 的蛋白质,是细胞内代谢所需的蛋白质。
•rRNA - ribosomal RNA —Is what the ribosomes are made of
•tRNA - transfer RNA —Brings the amino acids to the mRNA to string them together to make a protein
11-核糖体-细胞生物学
形态结构
7
核糖体上有五个重要的功能活性部位:
①. A位点 氨酰基-tRNA结合位点
②. P位点 肽酰基-tRNA结合位点
③. T因子 肽酰基转移酶位点
④. G因子 GTPase位点
⑤. E位点 空载tRNA离开核糖体前暂时停留的位点
.
形态结构
8
三、核糖体的类型
1.附着核糖体:
合成外输性蛋白, 如分泌蛋白;以及膜 蛋白、溶酶体酶及驻 留蛋白等。
核糖体的功能
.
29
肽链延长
核糖体的功能
.
30
肽链合成终 止
核糖体的功能
.
31
核糖体的功能
进位
肽链的形成
核糖体移 位
.
32
密码子及其特性
起始密码子——AUG;
终止密码子——UAA(ochre 赭石密码子) UAG(amber 琥珀密码子) UGA(opal 蛋白石密码子)
.
✓ 无标点、不重叠
✓ 简并(degeneracy)
✓ 1958年Roberts根据化学成份命名为 核糖核蛋白体,简称核糖体Ribosome。
.
35
2009 Nobel prize for studies of the structure and function of the ribosome
Venkatraman
Thomas A. Steitz
在急性药物中毒性(四氯化碳)
肝炎和病毒性肝炎后,以及肝硬化
病人的肝细胞中,经常可见到大
量多聚核糖体解聚呈离散单体状,
固着多聚核糖体脱落,分布稀疏、
导致分泌蛋白合成下降。所以病
人血浆白蛋白含量下降。
动物细胞的构成

动物细胞的构成动物细胞是构成动物体的基本结构单位,是一种复杂的生物单元。
它由许多不同的细胞器组成,每个细胞器都承担着特定的功能。
下面我们将详细介绍动物细胞的构成。
1. 细胞膜:细胞膜是动物细胞最外层的结构,它由脂质双层组成。
细胞膜起到保护细胞内部结构的作用,同时它还是物质进出细胞的通道。
2. 线粒体:线粒体是动物细胞内的能量工厂,它通过细胞呼吸过程产生的ATP 供给细胞的能量需求。
线粒体内含有DNA,具有自我复制的能力。
3. 核:核是细胞的控制中心,它包含着细胞的遗传物质DNA。
核内的DNA编码着细胞的遗传信息,并通过转录和翻译过程控制细胞的生物合成。
4. 核膜:核膜是包围核的双层膜,它具有选择性通透性,控制物质进出核的过程。
核膜上还有许多核孔,通过核孔,RNA可以从核内运输到细胞质中。
5. 核仁:核仁是核内的一个小结构,它参与细胞的蛋白质合成。
核仁内含有rRNA和蛋白质。
6. 溶酶体:溶酶体是动物细胞内的一种液泡结构,它含有多种水解酶,可以降解各种细胞内外的废物物质。
溶酶体还参与细胞的自噬过程。
7. 内质网:内质网是一种细胞内膜系统,它分为粗面内质网和平滑内质网两种。
粗面内质网上有许多附着的核糖体,参与蛋白质的合成和修饰。
平滑内质网参与脂质代谢、药物代谢等。
8. 高尔基体:高尔基体是细胞内的一个复合细胞器,它参与蛋白质的修饰、分拣和包装。
高尔基体还参与细胞膜的合成和分泌蛋白的运输。
9. 微管和微丝:微管和微丝是细胞骨架的组成部分,它们由蛋白质聚合而成。
微管参与细胞的分裂、细胞器的定位和运输等过程。
微丝参与细胞的运动和细胞骨架的维持。
10. 高尔基体:高尔基体位于细胞核附近,它是细胞内的一个复合细胞器,参与细胞膜的合成、分泌蛋白的加工和运输等过程。
11. 核糖体:核糖体是细胞内的蛋白质合成工厂,它由rRNA和蛋白质组成。
核糖体通过转录和翻译过程合成蛋白质,参与细胞的生物合成。
12. 非细胞器结构:除了上述细胞器外,动物细胞还包含一些非细胞器结构,如细胞质基质、细胞核基质等。
核糖体分子结构

核糖体分子结构核糖体是细胞内重要的蛋白质合成机器,起着决定蛋白质合成速率和准确性的关键作用。
它是由核糖核酸(rRNA)和蛋白质组成的复杂大分子结构。
在细胞质内,核糖体固定在内质网上,参与到翻译过程中。
核糖体的结构相当复杂,可以分为大亚基、小亚基和rRNA三个部分。
大亚基(50S)由两种不同的rRNA分子和多种蛋白质组成,小亚基(30S)也由rRNA和蛋白质构成。
两个亚基结合在一起形成70S核糖体。
核糖体rRNA是构成核糖体的重要组成部分,因其与翻译中的基因密码子相互配对,发挥着重要的功能。
细菌核糖体中有三种不同的rRNA:16S、23S和5S。
16S rRNA参与到识别启动子和选择正确的AUG密码子,从而使翻译正确开始;23S rRNA参与肽链合成的催化作用,并与药物相互作用,抑制细菌生长;5S rRNA担任支撑和稳定核糖体结构的角色。
另外,核糖体中的蛋白质也起着重要的功能。
它们可以稳定核糖体结构、帮助核糖体与mRNA进行相互作用、促进肽链的合成等。
核糖体蛋白质非常多样,每个细胞种类可能有几十种不同的核糖体蛋白质。
核糖体具有高度有序的结构,形成了一个功能完备的物理平台,使得蛋白质合成能够高效进行。
核糖体的结构不仅在这个生物系统中是高度保守的,也在不同生物种类中存在着相似性。
研究人员利用高分辨率的电子显微镜技术,揭示了核糖体在蛋白质合成过程中的精确分子结构。
这一发现为我们理解细胞中蛋白质合成的机制提供了独特的视角。
通过对核糖体结构的深入研究,我们可以更好地了解核糖体与药物相互作用的机制,为疾病治疗提供新的思路。
此外,深入研究核糖体结构还有助于发展新型抗菌药物,应对细菌耐药问题。
总之,核糖体是细胞内不可或缺的重要机器,其结构与功能的研究对于我们深入理解蛋白质合成的机制具有重要意义。
我们应当进一步加强对核糖体的研究,深入挖掘其在生命活动中的奥秘,为生物医学领域的科研和药物研发做出更大的贡献。
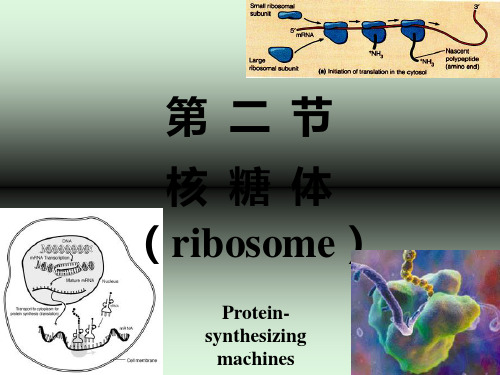
第十一章核糖体

第11章核糖体● 核糖体是细胞内一种核糖核蛋白颗粒(ribonucleoprotein particle), 其惟一功能是按照mRNA的指令将氨基酸高效且准确地合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
● 按核糖体存在的部位可分为三种类型:细胞质核糖体、线粒体核糖体、叶绿体核糖体。
按存在的生物类型可分为两种类型:真核生物核糖体和原核生物核糖体。
原核细胞的核糖体较小, 沉降系数为70S,相对分子质量为2.5×103 kDa,由50S和30S两个亚基组成;而真核细胞的核糖体体积较大,沉降系数是80S,相对分子质量为3.9~4.5×103 kDa, 由60S和40S两个亚基组成。
● 在真核细胞中,核糖体进行蛋白质合成时,既可以游离在细胞质中,称为游离核糖体,也可以附着在内质网的表面,称为膜旁核糖体或附着核糖体。
真核细胞含有较多的核糖体,每个细胞平均有106 ~107 个,而原核细胞中核糖体较少每个细胞平均只有15×102 ~18×103 个。
● 典型的原核生物大肠杆菌核糖体是由50S大亚基和30S小亚基组成的。
在完整的核糖体中,rRNA约占2/3, 蛋白质约为1/3。
50S大亚基含有34种不同的蛋白质和两种RNA分子,相对分子质量大的rRNA的沉降系数为23S,相对分子质量小的rRNA为5S。
30S小亚基含有21种蛋白质和一个16S的rRNA分子。
● 真核细胞核糖体的沉降系数为80S,大亚基为60S,小亚基为40S。
在大亚基中,有大约49种蛋白质,另外有三种rRNA:28S rRNA、5S rRNA和5.8S rRNA。
小亚基含有大约33种蛋白质,一种18S的rRNA。
关键词:核糖核蛋白颗粒;游离核糖体;附着核糖体;第一节核糖体的类型与结构核糖体 (ribosome)是合成蛋白质的细胞器,其惟一功能是按照mRNA的指令将氨基酸高效且准确地合成蛋白质多肽链。
【doc】蛋白质生物合成中核糖体延伸循环的变构三位点模型

蛋白质生物合成中核糖体延伸循环的变构三位点模型'生命的化学》1991年11卷第d期蛋白质生物合成中核糖体延伸循环的变构三位点模型蒋达和(武汉太学病毒及分子生物学系)自60年代提出了蛋白质生物台成中核糖体延伸循环的tRNA与梭糖体结合的两位点模型以来,迄今国内外各种生化教科书都一直采用这一模型[见图1A].现在,已有许多证据表明大肠杆菌的70s核糖体上除了A稻P位外,还存在结台tRNA的第3个位点即E位点(Exit),并证实在移位前,后的核糖体是处于两种不同的构象状态.据此而提出了一种核糖体延伸循环的变构三位点模型(见图lB).该模型的主要特征为:①核糖体含有A,P和E3个tRNA结台位点,E位特异地结台脱酰基的tRNA分子}@在移位过程中,脱酰基的tRNA不离开梭糖体,而是从P位移到E位,移位的顺序方向为tA—P—E}⑧移位前后,结合在核糖体上的两种tRNA都进行密码子一反密码子相互作用l④核糖体具有两种构象状态,一种是移位前状态,其A位和P位对tRNA具有高亲和力,而E位的亲和力较低,另一种为移位后状态,其P位和E位对tRNA具高亲和力,而A位的亲和力则减弱.原来低亲和力结台位点的被占据可以诱导两种构象的各自相互转变.可见,棱糖体A位和E位的构象变化是一种负协同效应,结果伴随着氨酰tRNA结台到A位的是脱酰基的tRNA从E位离开核糖体.变构3位点模型不同于两位点模型之处主7呈反向重复,导致反义RNA的产生Tosic等(1990)的进一步研究结果认为,mld小鼠正是由于反义RNA与有义RNA形成双链而阻止MBP的表达,印存在转录后的调控作用. Heywood等(1986)在鸡中发现,与肌动蛋白重链mRNA5t端有同源性的两个tcRNA 102,而且这些RNA之间有着密切关系.认为tcRNA102是特定肌mRNA的自然反义RNA.三,檀物中自然存在蚋反义RNA近年研究表明,植物中也自然存在有反义RNA.它们与在动物中发现的反义RNA有所不同.动物中发现的大多数反义RNA与目标mRNA仅有部分序列互补.但对大麦的研究发现,n一淀粉酶的反义P,:NA序列覆盖了A型和B型a一淀粉酶mRNA的全长序列.而且发现两型淀粉酶的mRNA之反义RNAs是由不同位点转录的.这两型淀粉酶mRNA及其反义RNA的表达在发育上受到调控,而且它们在糊粉组织中都以等量存在.另一特别反义RNA的存在形式是在启动子区中发现的.有一种Antirrhinummajus花的突变表型,该花的色素沉着受到改变.色素的遗传呈半显性形式.此种突变是由于编码CHS酶的Nivea位点受到转座子诱导的重排.CHS在类黄酮的台成过程中起着中心作用,它的失活将阻断花的色素沉着.对重排的分子遗传学分析发现,CHS的启动子区存在反向重复.由此将产生一个CHS的反义转录物,从而阻止CHS的转录.虽然上面讨论了真核生物中反义RNA的存在,但对它们自然存在的机理以及它们的生物学功能的了解还甚少.采用诱发反义基因突变和人工构建反义基因以及人工台成反义寡核苷酸等研究手段可对反义RNA的生物学功能进行深入研究.这一领域正受到国内外学者的广泛关注.4^t两位点嵌驵《生命的化学>1991年11卷第4期Bl查杓三位点辑塑图1核糖体延伸循环的三个主要反应示意图1.氨酰tRNA与A位结合,2.肽基转移;3.移位,方形接糖体表示移位前构象状老,移位后的接糖体构象状态以圆形表示.+网l2黯聃让畦E命的化学~1991年儿卷第4蜘人肿瘤细胞的多抗药性(MDR)徐湘民(第一军医大学生化教研室)自50年代应用化疗至今,化疗法已是人类对付癌症的主要手段之一但另一方面,化疗又始终受到肿瘤细胞抗药性的困扰.它们或表现为天然固有的抗药性,即癌细胞对化疗药几乎不敏感|或表现为从最初的高敏感转化为用药后的低敏感甚至不敏感,即所谓获得性抗药性.从而使化疗的应用范围和疗效受到了很大的制约.近年来多抗药性(multipl—drugre—sistancc,MDR)基因的发现及其抗药机理的研究,为解除这一困扰带来了希望.一,MDR和MDR基因像细菌可对一种以上的化学杀(抑)菌剂具有抗药性一样,肿瘤细胞的MDR是指其对多种化疗药物的交叉抗药性.迄今巳在动物的MDR细胞中(包括人肿瘤细胞)发现的抗性化疗药见附表.MDR细胞为什么会对化学结构和作用机制完全不同的抗癌药产生交叉抗药性,是一个长期使人迷惑不解的问题.解开此迷的最早研究始于在中国仓鼠细胞(CHC)膜上一种膜通透性糖蛋白(分子量170kd)的发现,此蛋白命名为P一糖蛋白或P170.在MDR细胞中,P170含量与抗药水平相平行,即抗药性越强,P170含量越高.P170的作用是将细胞内的某些外来或内生的亲脂性"毒物"清除出细胞,使其难以在细胞内发挥效应.在人肿瘤细胞一KB细胞系的研究中,以秋水仙碱为选择剂,使细胞获得了对秋水仙碱,长春碱,阿霉素及放线菌素D的MDR,细胞的MDR与P170的相关性同动物细胞中观察结果相一致. 无疑,P170只是参与MDR的一种表型.在明确了MDR与P170的基本关系后,人们的兴趣转向了MDR基因.细胞遗传学观察到MDR细胞总是伴有基因放大现象.利用胶内复性技术,对MDR的CHC的放大基因医进行克隆制备探针,用它钓取人KB细胞基因组中的同源序列获得成功.将人基因组的该DNA 序列制成探针,在人肿瘤细胞中找到了与MDR 相关的4.5kbmRNA,该mRNA在MDR高抗性细胞中总是处于高水平以KB细胞中提取的基因转导小鼠,小鼠在表达4.5kbmRNA的同时获得了MDR,在CHC药敏细胞中没能找到这种mRNA.而MDR细胞中提取的该mRNA能与放大基因区杂交上.至此,MDR从基因到蛋白的轮廓清楚了,即编码P170的基因——MDR基因包含在基因放大序列当中,以成,表明只有E拉存在正确的脱酰基tRNA时才能降低A位的亲和力,这些都意味着E位及E位上的密码子一反密码子相互作用对蛋白质的生物合成及其翻译精确性也超重要作用. 目前,变构3位点模型已在大肠杆菌和古细菌(Halobaetcriumlmlobium)等生物中得到证实,真核系统的大鼠肝80s核糖体上也发现有3个tRNA结合位点存在该模型可能与生物体内的实际较相符,但是否为生物界的普遍现象仍需进一步研究证实.图2为根据变构3位点模型绘制的核糖体延伸循环示意图.要是多子一个E位点,而且A位与E位之阔存在负协同的变构效应,移位前后核糖体上都存在有两个tRNA.最近发现,大肠扦菌核糖体的E位对蛋白质生物台成具有重要作用,除了为脱酰基tRNA离开核糖体时提供出口外,还有实验表明,E位被正确的脱酰基tRNA占据时,A位对不正确氨酰tRNA的结合可降到本底值,但对正确氨酰tRNA的结台几乎不受影响,说明E位被同种脱氨酰tRNA占据后可防止不正确氨酰tRNA结合到A位.当E位存在不正确的脱酰基tRNA时,则可增加不正确的氮酰tRNA与A位结台,参与蛋白质生物台。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
在蛋白质合成中肽酰转移酶的活性研究
核糖体蛋白
在核糖体中rRNA是起主
要作用的结构成分
r 蛋白质的主要功能
核糖体蛋白
很难确定哪一种蛋白具有催化功能: 在E. coli中核糖体蛋白突变甚至缺失对蛋 白质合成并没有表现出“全”或“无”的影 响。 多数抗蛋白质合成抑制剂的突变株,并非由于 r 蛋白的基因突变而往往是 rRNA基因突变。 在整个进化过程中rRNA的结构比核糖体蛋白 的结构具有更高的保守性。
离子交换树脂可分离纯化各种 r 蛋白; 纯化的 r 蛋白与纯化的rRNA进行核糖体的重组装,显 示核糖体中 r 蛋白与rRNA的结构关系 双向电泳技术可显示出 E.coli核糖体在装配各阶段中, 与rRNA结合的蛋白质的类型 双功能的交联剂和双向电泳分离可用于研究 r 蛋白在结 构上的相互关系 电镜负染色与免疫标记技术结合,研究 r 蛋白在核糖体 的亚单位上的定位。 对rRNA,特别是对16S rRNA结构的研究 70S核糖体的小亚单位中rRNA与全部的 r 蛋白关系的空 间模型
Comparison of ribosome structures: E.coli 70S (upper row in each panel, structure from 18,19) and yeast 80S (lower row in each panel) ribosomes, both limited to 35 Å resolution. (A–J) Rotational series, with a rotational increment around the vertical axis of 36. Scale bar 100 Å. (K) Exploded views of E.coli and yeast ribosomes, showing close correspondence of features of interface surfaces of ribosomal subunits, most notably the interface canyon (IC). CP, central protuberance; S, stalk; L1 or ‘L1’, L1 or L1 analog arm; h, head; g, neck groove; p, platform. Note that the lower right portion of the small subunit and the lower left portion of the large subunit are surfaces where the sectioning plane has cut a mass that is merged between the two subunits at this resolution. (adapted from Nucleic Acids Research, 1998, 26(2):655-661)
E. Coli 核糖体小亚单位中rRNA与 r 蛋白的相互关系示意图 线条表示相互作用及作用力的强(粗线)与弱(细线) (引自Alberts et al, 1989)
16S rRNA的一级结构是非常保守的;
16S rRNA的二级结构具有更高的保守性:
臂环结构 (stem-loop structure)
rRNA臂环结构的三级结构模型。
核糖体小亚单位rRNA的二级结构
(a) E. coli 16S rRNA;(红色为高度保守区); (b) 酵母菌18S rRNA,它们都具有类似的40个臂环结构(图中1~40),其长度和位置往 往非常保守;P、E分别代表仅在原核或真核细胞中存在的rRNA的二级结构。
与mRNA的结合位点; 与新掺入的氨酰-tRNA的结合位点 —— 氨酰基 位点,又称A位点; 与延伸中的肽酰-tRNA的结合位点 —— 肽酰基 位点,又称P位点; 肽酰转移后与即将释放的tRNA的结合位点 —— E位点(exit site); 与肽酰-tRNA从A位点转移到P位点有关的转移酶 (即延伸因子EF-G) 的结合位点; 肽酰转移酶的催化位点; 与蛋白质合成有关的其它起始因子、延伸因子和 终止因子的结合位点。
多聚核糖体的生物学意义
细胞内各种多肽的合成,不论其分子量的大小或是 mRNA的长短如何,单位时间内所合成的多肽分子 数目都大体相等。 以多聚核糖体的形式进行多肽合成,对mRNA的利 用及对其浓度的调控更为经济和有效。
Visualizing transcription and translation
蛋白质的合成 (synthesis of the protein) RNA在生命起源中的地位及其演化过程
一、多聚核糖体 (polyribosome or polysome)
概念
核糖体在细胞内并不是单个独立地执行功能,而 是由多个甚至几十个核糖体串连在一条mRNA分子上高 效地进行肽链的合成,这种具有特殊功能与形态结构 的核糖体与mRNA的聚合体称为多聚核糖体。
第十一章 核糖体(ribosome)
核糖体 (ribosome) 是核糖核
蛋白体的简称,是合成蛋白质的
细胞元件,其唯一的功能就是按
照mRNA的指令由氨基酸高效且 精确地合成出多肽链。
第十一章 核糖体(ribosome)
核糖体的基本类型
多聚核糖体与蛋白质的合成
作业
第一节 核糖体的基本类型
类型与组成成分 核糖体蛋白质与 rRNA的功能分析
一、核糖体的基本类型与成分
基本类型
附着核糖体
游离核糖体 70S的核糖体 80S的核糖体
存在于原核细胞还是真核细胞 游离于细胞质还是附着于ER
主要成分 r-protein: 40%,核糖体表面
NA:
60%,核糖体内部
Free and membrane-bound ribosomes
能的作用机制,从而为揭开核糖体这一具有30多亿年历
史的、古老的、高度而复杂的分子机器的运转奥秘迈出
了极重要的一步。
L11rRNA
复 合 物 的 三 维 结 构
二、核糖体蛋白质与 rRNA 的功能分析
核糖体上具有一系列与蛋白质合
成有关的结合位点与催化位点
在蛋白质合成中肽酰转移酶的活
性研究
核糖体上具有一系列与蛋白质 合成有关的结合位点与催化位点
在核糖体中rRNA是起主要作用的结构成分
具有肽酰转移酶的活性; 为tRNA提供结合位点(A位点、P位点和E 位点);
为多种蛋白质合成因子提供结合位点;
在蛋白质合成起始时参与同mRNA选择性地 结合以及在肽链的延伸中与mRNA结合; 核糖体大小亚单位的结合、校正阅读(proof reading)、无意义链或框架漂移的校正、以 及抗菌素的作用等都与rRNA有关。
(a)
(b)
mRNA Large subunit Small subunit
Large subunit Small subunit Completed polypeptide
(a)
(b)
Polyribosomes
(a) Schematic drawing of a polyribsome (polysome). (b) Electron micrograph of a grazing section through the outer edge of a rough ER cisterna. The ribosomes are aligned in loops and spirals, indicating their attachment to mRNA molecules to form polysomes. (c) Electron micrograph of metal-shadowed polysomes isolated from reticulocytes engaged in hemoglobin synthesis. Most of these polysomes have between four and six ribosomes.
mammalian
80S; 相对分子量: 4.2×106; 60%RNA.
60S 28S=4718 bp 5.8S=160 bp 5S=120 bp
40S 18S=1874 bp
49
33
核糖体的结构
结构与功能的分析方法 蛋白质合成过程中很多重要步 骤与 50S 核糖体大亚单位相关
结构与功能的分析方法
Comparison of Ribosomal Components between Prokaryotes and Eukaryotes species
bacterium
ribosome
70S; 相对分子量: 2.5×106; 66%RNA.
subunit
rRNA
r-protein
31 21
50S 23S=2904 bp 5S=120 bp 30S 16S=1542 bp
蛋白质合成过程中很多重 要步骤与50S核糖体大亚单位相关
涉及的多数因子为G蛋白(具有GTPase活性),核糖体
上与之相关位点称为GTPase相关位点。 最近人们成功地制备L11-rRNA复合物的晶体,获得了 其空间结构高分辨率的三维图象。 这一结果证实了前人用各种实验所获得的种种结论。
提出直观、可靠而且比人们的预料更为精巧复杂和可
(a)
(b)
Important Binding Sites on the Prokaryotic Ribosome
This model of ribosome structure shows the A (aminoacyl) and P (peptidyl) sites as cavities on the ribosome where charged (amino acid-carrying) tRNA molecules bind during polypeptide synthesis. The more recently postulated E (exit) site is the site from which discharged tRNAs leave the ribosome. The mRNA-binding site binds a particular nucleotide sequence near the 5’ end of the mRNA, placing the mRNA in the proper position for the translation of its first codon. (a) The diagrammatic representation of a ribosome. The pair of horizontal dashed lines indicate where the mRNA molecule lies. (b) A more realistic representation. The binding sites are all located at or near the interface between the large and small subunits.