南京农业大学 生命科学学院
进入本专业毕业时专业技行政职
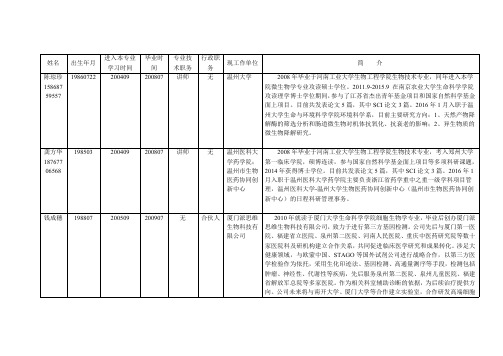
姓名出生年月进入本专业学习时间毕业时间专业技术职务行政职务现工作单位简介陈琼珍158687 5955719860722200409200807讲师无温州大学2008年毕业于河南工业大学生物工程学院生物技术专业,同年进入本学院微生物学专业攻读硕士学位。
2011.9-2015.9在南京农业大学生命科学学院攻读理学博士学位期间,参与了江苏省杰出青年基金项目和国家自然科学基金面上项目。
目前共发表论文5篇,其中SCI论文3篇。
2016年1月入职于温州大学生命与环境科学学院环境科学系,目前主要研究方向:1、天然产物降解酶的筛选分析和肠道微生物对机体抗氧化、抗衰老的影响;2、异生物质的微生物降解研究。
龚方华187677 06568198503200409200807讲师无温州医科大学药学院;温州市生物医药协同创新中心2008年毕业于河南工业大学生物工程学院生物技术专业,考入郑州大学第一临床学院,硕博连读,参与国家自然科学基金面上项目等多项科研课题,2014年获得博士学位。
目前共发表论文5篇,其中SCI论文3篇。
2016年1月入职于温州医科大学药学院主要负责浙江省药学重中之重一级学科项目管理,温州医科大学-温州大学生物医药协同创新中心(温州市生物医药协同创新中心)的日程科研管理事务。
钱成穗198807200509200907无合伙人厦门派思维生物科技有限公司2010年就读于厦门大学生命科学学院细胞生物学专业,毕业后创办厦门派思维生物科技有限公司,致力于进行第三方基因检测。
公司先后与厦门第一医院、福建省立医院、泉州第二医院、河南人民医院、重庆中医药研究院等数十家医院科及研机构建立合作关系,共同促进临床医学研究和成果转化。
涉足大健康领域,与欧蒙中国、STAGO等国外试剂公司进行战略合作,以第三方医学检验作为依托,采用生化印迹法、基因检测、高通量测序等手段,检测包括肿瘤、神经性、代谢性等疾病,先后服务泉州第二医院、泉州儿童医院、福建省解放军总院等多家医院。
植物耐铝的生物化学与分子机理

摘要:某些耐铝植物在铝胁迫下分泌有机酸被认为是 一个重要的抗性机制。从根系分泌出来的有机酸能与 根际的 >-$ ? 结合, 形成无毒性的螯合物, 从而减轻了 铝对根系的毒害。但是, 铝诱导有机酸分泌的中间环 节及调节机制至今仍不清楚。一些证据表明, 铝能激 活根尖细胞质膜内的阴离子通道, 因而可以调节有机 酸的分泌。近年来, 人们开始注意一些信号分子如蛋 白激酶、 水杨酸等介导铝诱导有机酸的分泌, 已经获得 一些成果。同时, 铝胁迫基因的分离和鉴定也为人们 从分子水平上研究和认识铝胁迫下植物的抗性机制奠 定了基础。 关键词:铝; 有机酸; 蛋白激酶; 水杨酸; 铝诱导基因 中图分类号:@=&<
G<0
植物生理与分子生物学学报\ \ \ \ \ \ \ \ \ \ \ \ \ \ \ \ \ \ 0* 卷
时, 我们也测定了柠檬酸的含量, 结果显示铝处理 的根尖柠檬酸的含量高出对照 ! 倍左右。铝刺激 有机酸相关合成酶活性提高的这种机制得到转基 因实验的支持。有人将大肠杆菌柠檬酸合酶基因 转入到烟草幼苗, 结果显示转基因植株具有较高的 柠檬酸合酶活性, 柠檬酸分泌量增加, 植株表现出 较强的抗性 ( "# $% &’#()# 等 !**+ ) 。之后, ,#-.%/# 等 ( 011! ) 用豆科植物根瘤促进型 ( (2"’$#3#(4%(5#" .267-) 的 89: 基因导入苜蓿后, 发现转基因植株 89: 活性提高 !; < 倍, 根尖内苹果酸含量提高了 =; 0 倍, 有机酸的分泌提高了 +; ! 倍。但也有一些 结果不支持铝诱导相关酶活性提高的有机酸分泌 机制。例如, 在以铝诱导苹果酸分泌为主的抗性小 麦上, 观察不到 89: 活性的提高, 表明铝诱导苹 果酸的分泌并不一定与根系内相应酶活性的提高 有直接关系 ( >/%( 等 !**? ) 。
2019南京农业大学生命科学学院硕士拟录取名单

249.80
598.80
章文华
非定向就业
全日制
生命科学学院
统考
071001
植物学
王文磊
103079210000339
348
240.80
588.80
徐益峰
非定向就业
全日制
生命科学学院
统考
071001
植物学
李建民
103079210000999
344
240.00
584.00
陈亚华
非定向就业
全日制
生命科学学院
370
223.00
593.00
曹慧
非定向就业
全日制
生命科学学院
统考
071005
微生物学
王颖
103079210007882
358
234.00
592.00
王伟武
非定向就业
全日制
拟录取学院
考试方式
拟录取专业代码
拟录取专业名称
姓名
考生编号
初试成绩
复试成绩
总成绩
拟录取导师
拟录取类别
拟录取培养方式
备注
生命科学学院
212.00
522.00
李新华
非定向就业
全日制
生命科学学院
统考
071001
植物学
唐永伟
103079210009031
340
181.90
521.90
陈晨
非定向就业
全日制
生命科学学院
统考
071001
植物学
苏有科
103079210000234
316
188.50
南京农业大学016生命科学学院2019级硕士拟录取名单
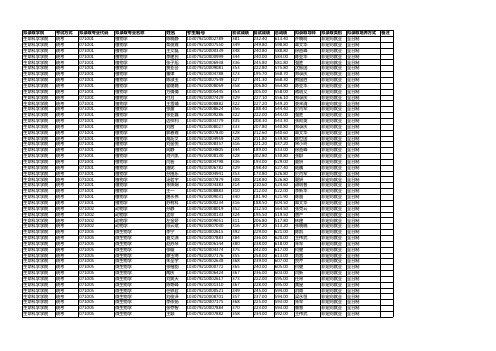
生命科学学院统考071001植物学陈晓静103079210002789381232.40613.40许晓明非定向就业全日制生命科学学院统考071001植物学栾俊霞103079210007550349249.80598.80章文华非定向就业全日制生命科学学院统考071001植物学王文磊103079210000339348240.80588.80徐益峰非定向就业全日制生命科学学院统考071001植物学李建民103079210000999344240.00584.00陈亚华非定向就业全日制生命科学学院统考071001植物学张子旭103079210006938336245.80581.80强胜非定向就业全日制生命科学学院统考071001植物学贾亚会103079210009081353222.80575.80沈振国非定向就业全日制生命科学学院统考071001植物学董律103079210004788373195.70568.70郑录庆非定向就业全日制生命科学学院统考071001植物学陈淑玉103079210007549327241.30568.30熊国胜非定向就业全日制生命科学学院统考071001植物学翟璐璐103079210008069358206.80564.80陈亚华非定向就业全日制生命科学学院统考071001植物学方倩倩103079210005445353205.00558.00蒋明义非定向就业全日制生命科学学院统考071001植物学付月103079210007429329227.10556.10郑录庆非定向就业全日制生命科学学院统考071001植物学王雪靖103079210008882322227.20549.20娄来清非定向就业全日制生命科学学院统考071001植物学张露103079210008624356188.40544.40於丙军非定向就业全日制生命科学学院统考071001植物学张亚鑫103079210009286322222.00544.00强胜非定向就业全日制生命科学学院统考071001植物学边祥利103079210003779335208.30543.30张阿英非定向就业全日制生命科学学院统考071001植物学刘茜103079210008027333207.80540.80郑录庆非定向就业全日制生命科学学院统考071001植物学郭春霞103079210007830328212.60540.60章文华非定向就业全日制生命科学学院统考071001植物学高沥文103079210004959328211.80539.80陈世国非定向就业全日制生命科学学院统考071001植物学刘金悦103079210008357316221.20537.20宋小玲非定向就业全日制生命科学学院统考071001植物学闻静103079210004805344189.00533.00徐益峰非定向就业全日制生命科学学院统考071001植物学肖兴凯103079210008130328202.80530.80张群非定向就业全日制生命科学学院统考071001植物学闫晋103079210004798336193.00529.00夏妍非定向就业全日制生命科学学院统考071001植物学潘妮103079210006782329198.40527.40陆巍非定向就业全日制生命科学学院统考071001植物学孙施乐103079210004941353173.80526.80於丙军非定向就业全日制生命科学学院统考071001植物学汤茗宇103079210007879308218.80526.80夏妍非定向就业全日制生命科学学院统考071001植物学朱姝娴103079210004383314210.60524.60谭明普非定向就业全日制生命科学学院统考071001植物学王一103079210008883310212.00522.00李新华非定向就业全日制生命科学学院统考071001植物学唐永伟103079210009031340181.90521.90陈晨非定向就业全日制生命科学学院统考071001植物学苏有科103079210000234316188.50504.50章文华非定向就业全日制生命科学学院统考071002动物学孙静103079210008019352212.50564.50张克云非定向就业全日制生命科学学院统考071002动物学孟珍103079210000133324195.50519.50曾严非定向就业全日制生命科学学院统考071002动物学左金娇103079210009051311206.80517.80林建非定向就业全日制生命科学学院统考071002动物学段云斌103079210007030316197.20513.20张晓晓非定向就业全日制生命科学学院统考071005微生物学李宁103079210002615392229.00621.00陈凯非定向就业全日制生命科学学院统考071005微生物学夏文潇103079210007883384236.00620.00王伟武非定向就业全日制生命科学学院统考071005微生物学赵苏琴103079210006144380238.00618.00朱军非定向就业全日制生命科学学院统考071005微生物学张璇103079210004374375242.00617.00何健非定向就业全日制生命科学学院统考071005微生物学廖玉绮103079210007176355258.00613.00刘蓉非定向就业全日制生命科学学院统考071005微生物学朱全宇103079210002638368239.00607.00贺芹非定向就业全日制生命科学学院统考071005微生物学张楷韵103079210000772365240.00605.00何健非定向就业全日制生命科学学院统考071005微生物学陶庆103079210006424367236.00603.00闫新非定向就业全日制生命科学学院统考071005微生物学刘笑天103079210002617373222.00595.00任昂非定向就业全日制生命科学学院统考071005微生物学陈奇峰103079210001310367228.00595.00黄星非定向就业全日制生命科学学院统考071005微生物学任轶君103079210000521349245.00594.00洪青非定向就业全日制生命科学学院统考071005微生物学刘俊泽103079210008701357237.00594.00梁永恒非定向就业全日制生命科学学院统考071005微生物学李依扬103079210007175368225.00593.00朱军非定向就业全日制生命科学学院统考071005微生物学张存智103079210007884370223.00593.00曹慧非定向就业全日制生命科学学院统考071005微生物学王颖103079210007882358234.00592.00王伟武非定向就业全日制生命科学学院统考071005微生物学裴冬梅103079210004951346244.00590.00闫新非定向就业全日制生命科学学院统考071005微生物学商娜103079210009054350238.00588.00黄星非定向就业全日制生命科学学院统考071005微生物学李晓阳103079210007416359229.00588.00何琳燕非定向就业全日制生命科学学院统考071005微生物学王令帅103079210008071377210.00587.00赵明文非定向就业全日制生命科学学院统考071005微生物学曹伟苗103079210008070348238.00586.00徐希辉非定向就业全日制生命科学学院统考071005微生物学吴川徽103079210005749351228.00579.00钟增涛非定向就业全日制生命科学学院统考071005微生物学纪彦晗103079210008078346229.00575.00蒋建东非定向就业全日制生命科学学院统考071005微生物学苗永豪103079210001312363211.00574.00朱军非定向就业全日制生命科学学院统考071005微生物学王晓雯103079210007342343229.00572.00黄彦非定向就业全日制生命科学学院统考071005微生物学王三乐103079210000522339229.00568.00钟增涛非定向就业全日制生命科学学院统考071005微生物学董文103079210009631345223.00568.00盛下放非定向就业全日制生命科学学院统考071005微生物学李悦103079210006421341226.00567.00曹慧非定向就业全日制生命科学学院统考071005微生物学孙海荣103079210009088345220.00565.00盛下放非定向就业全日制生命科学学院统考071005微生物学吴毅103079210006937349214.00563.00徐冬青非定向就业全日制生命科学学院统考071005微生物学游霞103079210007331332230.00562.00梁永恒非定向就业全日制生命科学学院统考071005微生物学周格帆103079210007887342218.00560.00刘蓉非定向就业全日制生命科学学院统考071005微生物学樊启文103079210007174331224.00555.00崔中利非定向就业全日制生命科学学院统考071005微生物学徐祖凤103079210005755339214.00553.00冉婷婷非定向就业全日制生命科学学院统考071005微生物学丁丽琴103079210007180344208.00552.00王卉非定向就业全日制生命科学学院统考071005微生物学杨政艳103079210008072356195.00551.00于汉寿非定向就业全日制生命科学学院统考071008发育生物学高尚雨103079210001452328240.90568.90朱昌华非定向就业全日制生命科学学院统考071008发育生物学李梦烁103359000912612358163.50521.50强胜非定向就业全日制调剂生命科学学院统考071008发育生物学张宇杰103079210004799325191.00516.00戴伟民非定向就业全日制生命科学学院统考071008发育生物学李雷103079210003781299175.00474.00宋小玲非定向就业全日制生命科学学院统考071009细胞生物学江晶晶103079210006292343236.00579.00蒋明义非定向就业全日制生命科学学院统考071009细胞生物学刘承健103079210007596331197.20528.20张群非定向就业全日制生命科学学院统考071009细胞生物学崔珍珍103079210009201306168.00474.00蒋明义非定向就业全日制生命科学学院统考071010生物化学与分子生物学王雨童103079210003778396264.00660.00腊红桂非定向就业全日制生命科学学院统考071010生物化学与分子生物学王曰桥103079210008073363249.00612.00沈文飚非定向就业全日制生命科学学院统考071010生物化学与分子生物学赵迪迪103079210007889359226.00585.00谢彦杰非定向就业全日制生命科学学院统考071010生物化学与分子生物学胡丽丽103079210009094361219.00580.00王心宇非定向就业全日制生命科学学院统考071010生物化学与分子生物学李苗苗103079210007179338239.00577.00郭晶晶非定向就业全日制生命科学学院统考071010生物化学与分子生物学徐若然103079210007419350213.00563.00芮琪非定向就业全日制生命科学学院统考071010生物化学与分子生物学赵琦103079210010576342214.00556.00李盛本非定向就业全日制生命科学学院统考071010生物化学与分子生物学吴梦瑶103079210000237327226.00553.00张炜非定向就业全日制生命科学学院统考071010生物化学与分子生物学徐浩月103079210004785345205.00550.00鲍依群非定向就业全日制生命科学学院统考071010生物化学与分子生物学叶杰103079210002625327214.00541.00刘峰非定向就业全日制生命科学学院统考071010生物化学与分子生物学殷兴华103079210009096332207.00539.00张炜非定向就业全日制生命科学学院统考071010生物化学与分子生物学徐彦103079210006374308228.00536.00郭晶晶非定向就业全日制生命科学学院统考071010生物化学与分子生物学汪金103079210006426330192.00522.00鲍依群非定向就业全日制生命科学学院统考071010生物化学与分子生物学陈威仲103079210005353309208.00517.00陈熙非定向就业全日制生命科学学院统考071010生物化学与分子生物学夏辉辉103079210006146325191.00516.00腊红桂非定向就业全日制生命科学学院统考071010生物化学与分子生物学翟丰超103079210008429310205.00515.00谢彦杰非定向就业全日制生命科学学院统考071010生物化学与分子生物学马利国103079210000820309206.00515.00李盛本非定向就业全日制生命科学学院统考071010生物化学与分子生物学刘永刚103079210008806315190.00505.00熊国胜非定向就业全日制生命科学学院统考085238生物工程桂鹏103079210006571390241.20631.20钟增涛非定向就业全日制生命科学学院统考085238生物工程王海燕103079210005407394232.00626.00何健非定向就业全日制生命科学学院统考085238生物工程花少伟103079210004129397216.40613.40王卉非定向就业全日制生命科学学院统考085238生物工程徐铭阳103079210008336387222.30609.30黄星非定向就业全日制生命科学学院统考085238生物工程姚晓亮103079210005409388216.70604.70闫新非定向就业全日制生命科学学院统考085238生物工程戴建成103079210007302377223.50600.50赵明文非定向就业全日制生命科学学院统考085238生物工程吴琦103079210007304363233.10596.10於丙军非定向就业全日制生命科学学院统考085238生物工程汪岚103079210009912370215.90585.90陈世国非定向就业全日制生命科学学院统考085238生物工程魏鑫怡103079210005454355229.90584.90顾向阳非定向就业全日制生命科学学院统考085238生物工程陈欣然103079210005919377207.80584.80徐剑宏非定向就业全日制生命科学学院统考085238生物工程孙仁杰103079210007801353231.20584.20甘立军非定向就业全日制生命科学学院统考085238生物工程周慧敏103079210006425334245.90579.90陈新非定向就业全日制调剂生命科学学院统考085238生物工程吕娜103079210000784365214.60579.60蒋建东非定向就业全日制生命科学学院统考085238生物工程张东豪103079210008595368210.70578.70陈晨非定向就业全日制生命科学学院统考085238生物工程王浩103079210004868367207.70574.70张克云非定向就业全日制生命科学学院统考085238生物工程侯雅楠103079210004436343230.90573.90陈亚华非定向就业全日制生命科学学院统考085238生物工程姚世敏103079210005920360212.10572.10王伟武非定向就业全日制生命科学学院统考085238生物工程张皓文103079210008967362205.20567.20陈亚华非定向就业全日制生命科学学院统考085238生物工程吴兴103079210005408357209.50566.50李新华非定向就业全日制生命科学学院统考085238生物工程陈皖松103079210005486341224.40565.40强胜非定向就业全日制生命科学学院统考085238生物工程关立康103079210000092346218.30564.30夏妍非定向就业全日制生命科学学院统考085238生物工程刘维佳103079210007512334224.10558.10张群非定向就业全日制调剂生命科学学院统考085238生物工程金勤103079210010180351201.90552.90于汉寿非定向就业全日制调剂生命科学学院统考085238生物工程孙宇103079210007303332219.10551.10娄来清非定向就业全日制生命科学学院统考085238生物工程路书山103079210006803339212.10551.10曹慧非定向就业全日制生命科学学院统考085238生物工程徐兵103079210005750343205.50548.50师亮非定向就业全日制调剂生命科学学院统考085238生物工程冯冰103079210008801330217.30547.30钟增涛非定向就业全日制调剂生命科学学院统考085238生物工程曹悦103079210004866328215.90543.90李霞非定向就业全日制生命科学学院统考085238生物工程葛立刚103079210006570355188.90543.90贺芹非定向就业全日制生命科学学院统考085238生物工程方波103079210006535336206.90542.90崔瑾非定向就业全日制调剂生命科学学院统考085238生物工程刘宇昊103079210009085329211.80540.80沈文飚非定向就业全日制调剂生命科学学院统考085238生物工程杨图南103079210009510334204.80538.80陈凯非定向就业全日制生命科学学院统考085238生物工程张静103079210001230321217.60538.60强胜非定向就业全日制调剂生命科学学院统考085238生物工程邱钰丰103079210009352329209.00538.00徐剑宏非定向就业全日制生命科学学院统考085238生物工程李震103079210009465332205.50537.50宋小玲非定向就业全日制调剂生命科学学院统考085238生物工程张治荣103079210004561334203.40537.40谢彦杰非定向就业全日制调剂生命科学学院统考085238生物工程丁佩103079210009522311223.80534.80陈新非定向就业全日制调剂生命科学学院统考085238生物工程刘威103079210004375320213.70533.70刘蓉非定向就业全日制调剂生命科学学院统考085238生物工程叶森柯103079210005410320209.10529.10谭明普非定向就业全日制生命科学学院统考085238生物工程孙茜茜103079210004944312216.90528.90戴伟民非定向就业全日制调剂生命科学学院统考085238生物工程徐珩103079210002624310218.30528.30郑录庆非定向就业全日制调剂生命科学学院统考085238生物工程黄波103079210009612317207.60524.60陈明杰非定向就业全日制生命科学学院统考085238生物工程严斌103079210004435317203.20520.20陈新非定向就业全日制生命科学学院统考085238生物工程王曙103079210006075309202.40511.40沈文飚非定向就业全日制生命科学学院统考085238生物工程周亚飞103079210008710300198.50498.50许晓明非定向就业全日制生命科学学院推荐免试071001植物学魏田慧1030791022300580张阿英非定向就业全日制生命科学学院推荐免试071001植物学张德瑞1030791069402040崔瑾非定向就业全日制生命科学学院推荐免试071001植物学宋梦萦1030791030703190甘立军非定向就业全日制生命科学学院推荐免试071001植物学牛梦洋1030791046403560崔瑾非定向就业全日制生命科学学院推荐免试071001植物学刘泽1030791030704270沈振国非定向就业全日制生命科学学院推荐免试071005微生物学刘亚莉1030791022300570洪青非定向就业全日制生命科学学院推荐免试071005微生物学钟玲丽1030791030700770崔中利非定向就业全日制生命科学学院推荐免试071005微生物学张馥1030791030702400蒋建东非定向就业全日制生命科学学院推荐免试071005微生物学范君佩1030791019303600赵明文非定向就业全日制生命科学学院推荐免试071005微生物学吴高月1030791030704350顾向阳非定向就业全日制生命科学学院推荐免试071010生物化学与分子生物学王俊杰1030791030703580沈文飚非定向就业全日制生命科学学院推荐免试085238生物工程王苗1030791030703570沈振国非定向就业全日制。
棉铃虫中肠钙黏蛋白Cry1A结合区编码基因的原核表达及多克隆抗体制备

本研究旨在通过原核表达棉铃虫中肠钙黏蛋白 Cy A的 rl 毒素结合区的 基 因片段 , 命名 为 C d—t, a b 获得 重组蛋 白纯 品, 以此免疫动物 , 制备 以该融合蛋白为抗原的抗兔多克隆抗 体, 为进一步探索棉铃虫对 转基因棉的抗性机理及制定抗
~
2 6一
江苏农业科学
21 0 0年第 2期
黄晶晶, 常菊花 , 陈楚芹, 等.棉铃虫中肠钙黏蛋 白c)A结合 区编码基因的原核表达及 多克隆抗体制备[ ] r1 r J .江苏农业科学, 1( )2 2 2 0 2 :6- 9 0
棉铃虫 中肠钙黏蛋 白 CyA结合 区编码基 因 rl 的原核表达及 多克 隆抗体制备
检测其效价 。结果显示 , pT一 0 — a t表达载体转化 B 2 菌后 , II 导成功获得 分子量为 3 u 以 E 3a Cd—b L1 经 P ̄诱 ' 7k 左右
的重组 蛋白 , 单抗 一Hs Wet l 鉴定其正确表达 。纯化后的蛋白成功免疫新西兰大 白兔 , i的 s r bo e n t 获得 了该蛋白的多克
京 农 业 大学 生 命 科 学 学 院 生 物 化 学 与 分 子 生 物 学 实 验 室 保
药产生抗性 , B 农药与 转基 因棉 间存在交互抗性 , 且 t 这为 棉铃虫对 转基因棉产 生抗性 提出了早期预警 。B 活化毒 t 素能 决 定其 杀 虫 特 异 性和毒力的主要 因素 。钙黏蛋 白是 B 毒素 Cy A的主 t rl
黄 晶晶 ,常菊花 ,陈楚 芹 ,沈晋 良 沈 文飚 。 ,
(. 1南京农业大学生命科学学 院, 江苏南京 20 9 ; . 10 5 2 南京农业大学植 物保护学院 , 江苏南京 2 09 10 5; 3 南京农业 大学食品科技学院 , . 江苏南京 20 9 ) 10 5
南京农业大学2014年SRT计划项目一览表

生命科学学院 1410A10 学校资助 万晓辉 章文华 利用 Na+荧光探针比较研究野生型和突变体水稻 Na+浓度差异
1200
生命科学学院 1410A11 学校资助 章钰婷 张阿英 玉米中 JHDM 在 BR 信号转导中的功能分析
1200
生命科学学院 1410A12 学校资助 董丹玥 任 昂 氧化胁迫条件下转录因子 AP1 对灵芝三萜含量的影响
农学院
1411A07 学校资助 倪天界 赵团结 大豆开花期叶黄突变体 NT-y2 的光合生理特点与基因定位研究
经费 (元) 1200 1200 1200 1200 1200 1200 1200 1200 1200 1200 1200
学院
农学院 农学院 农学院 农学院 农学院 农学院 农学院 农学院 农学院 农学院 农学院
学院
项目编号
资助类别
项目主持 指导教师 项目名称
人
植物保护学院 1412A16 学校资助 高庆
赵弘巍 小分子 RNA 结合蛋白 argonaute 在病原细菌侵染后的定位变化研究
资源与环境科 1413A01
学学院
学校资助
田丁好
潘剑君
面向粮食安全的农用地质量综合评价技术研究
资源与环境科 1413A02
1200
生命科学学院 1410A13 学校资助 王 雪 黄 明 肉种类的检测与研究
1200
生命科学学院 1410A14 学校资助 高 岩 沈振国 重金属铜污染土壤修复植物的处理和利用
1200
生命科学学院 1410A15 学校资助 王 谨
张克云
昆虫病原线虫强毒品系 Heterorhabditidoides rugaoensis RG081015 1200
南京农业大学2011年SRT计划项目一览表-南京农业大学教务处
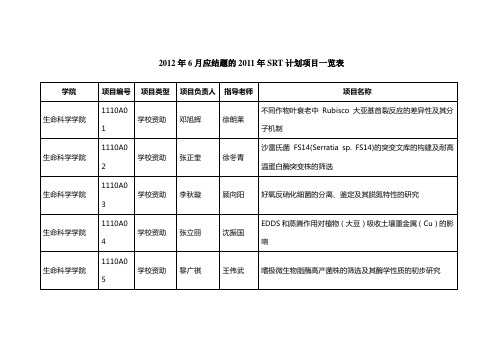
学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 项目编号 1110A01 1110A02 1110A03 1110A04 1110A05 1110A06 1110A07 1110A08 1110A09 1110A10 1110A11 1110A12 1110A13 1110A14 1110A15 1110A16 1110A17 1110A18 项目类型 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 项目负责人 邓旭辉 张正奎 李秋璇 张立丽 黎广祺 陈亚男 牟尚婕 赵泽 王周圆 潘雨博 周雪晨 张中南 刘鹏宇 吴慧 毛学伟 唐芮 吉家正 李星星 指导老师 徐朗莱 徐冬青 顾向阳 沈振国 王伟武 许晓明 黄星 盛下放 赖仞 钟增涛 刘永杰 于汉寿 沈标 王卉 鲍依群 陈亚华 赵团结 张炜 项目名称 不同作物叶衰老中 Rubisco 大亚基首裂反应的差异性及其分子 机制 沙雷氏菌 FS14(S反硝化细菌的分离、鉴定及其脱氮特性的研究 EDDS 和蒸腾作用对植物(大豆)吸收土壤重金属(Cu)的影响 嗜极微生物脂酶高产菌株的筛选及其酶学性质的初步研究 腐植酸与 PSⅡ除草剂的交互作用 高效纤维素降解菌的固定化研究 不同污染程度超积累植物内生产 ACC 脱氨酶细菌多样性研究 圆眼珍珠蛙皮肤活性肽的分离纯化 百脉根根瘤菌与宿主共生结瘤过程中时序性表达基因的筛选 南京地区犬细小病毒感染的调查及病原基因型分析 绿色荧光蛋白基因标记蜜蜂螺原体致病机理研究 黄瓜枯萎病病原菌拮抗菌的筛选及其生物防治效应 霍乱弧菌基因 vc0787 对其生物膜形成的影响 Patellin 蛋白家族的表达谱分析 外生菌根菌对宿主植物耐盐碱性影响的研究 大豆叶形态建成关键基因 alf 的图位克隆与表达分析 鸭疫里氏杆菌亚单位疫苗的研制
蛋白质结构分析原理及工具-文献综述

蛋白质结构分析原理及工具(南京农业大学生命科学学院生命基地111班)摘要:本文主要从相似性检测、一级结构、二级结构、三维结构、跨膜域等方面从原理到方法再到工具,系统地介绍了蛋白质结构分析的常用方法。
文章侧重于工具的列举,并没有对原理和方法做详细的介绍。
文章还列举了蛋白质分析中常用的数据库。
关键词:蛋白质;结构预测;跨膜域;保守结构域1 蛋白质相似性检测蛋白质数据库。
由一个物种分化而来的不同序列倾向于有相似的结构和功能。
物种分化后形成的同源序列称直系同源,它们通常具有相似的功能;由基因复制而来的序列称为旁系同源,它们通常有不同的功能[1]。
因此,推测全新蛋白质功能的第一步是将它的序列与进化上相关的已知结构和功能的蛋白质序列比较。
表一列出了常用的蛋白质序列数据库和它们的特点。
表一常用蛋白质数据库网址可能有更新氨基酸替代模型。
进化过程中,一种氨基酸残基会有向另一种氨基酸残基变化的倾向。
氨基酸替代模型可用来估计氨基酸替换的速率。
目前常用的替代模型有Point Accepted Mutation (PAM)矩阵、BLOck SUbstitution Matrix (BLOSUM)矩阵[2]、JTT模型[3]。
序列相似性搜索工具。
序列相似性搜索又分为成对序列相似性搜索和多序列相似性搜索。
成对序列相似性搜索通过搜索序列数据库从而找到与查询序列相似的序列。
分为局部联配和全局联配。
常用的局部联配工具有BLAST和SSEARCH,它们使用了Smith-Waterman 算法。
全局联配工具有FASTA和GGSEARCH,基于Needleman-Wunsch算法。
多序列相似性搜索常用于构建系统发育树,这里不阐述。
表二列举了常用的成对序列相似性比对搜索工具表二成对序列相似性比对搜索工具网址可能有更新2 蛋白质一级结构分析(含保守结构域)蛋白质结构的基本信息来源于它的一级结构,分析蛋白质一级结构的第一步是将它们分成其组成部分,然后处理每个部分的结构[4]。
《植物生理学》中外教材的比较

《植物生理学》课程双语、多媒体教学引出的思考陈亚华夏凯沈振国蔡庆生*南京农业大学生命科学学院,南京210095摘要:双语教学、课件制作和教材建设是课程改革的重要组成部分,它们各自改革的侧重点不同,其中,双语教学主要侧重提高学生应用外语的能力,课件制作侧重提高授课效率,而教材建设则主要体现教学大纲所涵盖的内容,并且伴随着科学的发展不断予以更新和充实。
双语教学和课件制作离不开相匹配的教材建设。
自2001年教育部颁发《关于加强高等学校本科教学工作提高教学质量的若干意见》、提出加强大学教学质量的12项措施、要求各高校在三年内开设5%-10%的双语课程以来,各高校越来越注重本科教学与国际接轨,优先在专业基础课实施、探讨和推广双语教学,进行课程教学改革,旨在满足培养具有国际竞争力高层次人才的需要。
作为南京农业大学校级精品建设课程之一,近年我们在《植物生理学》本科教学过程中,在包括双语教学、课件制作和国外教材选用等课程改革方面作了一些尝试。
一、以专业词汇为切入点,进行行之有效的双语教学《植物生理学》的授课对象通常为大学2年级学生,他们中的绝大多数已通过了大学英语4、6级考试,公共基础英语达到较高水平。
然而,《植物生理学》作为一门专业基础课,它涵盖有诸多专业学术词汇,这些是学生们在学习这门课程之前从未接触过的,在双语教学过程中,要想让学生弄懂和掌握《植物生理学》基本理论和知识,首先必须引导他们熟悉专业术语。
因此,处于起步阶段的《植物生理学》课程双语教学,通常以用双语讲解专业词汇为切入点和主要形式,配之以英语图表的讲解。
实施这种最为基本的双语教学形式,一方面可以因材施教,在引导学生规范地用英文表达基本概念和关键知识点的基础上,拓宽专业视野,逐步提高学生们的植物生理学及其相关学科的学习能力、专业英语应用和国际学术交流能力;另一方面,可以缓解和避免在现阶段实际双语教学过程中所存在的教学内容和课时的矛盾和授课教师用英语自如表达的困难。
南京农业大学2012年SRT计划项目一览表-南京农业大学大学生
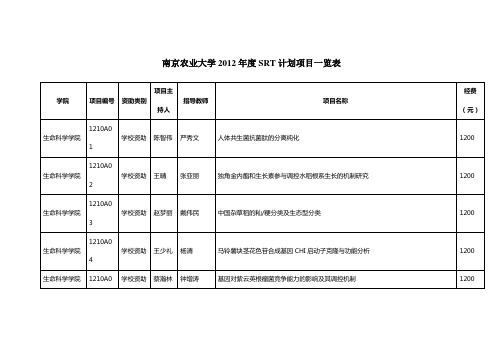
不结球白菜突变体库的构建 济宁点大豆杂优组合筛选及其遗传基础研究 大豆异交结实性影响因子的探究 利用大豆代换系进行茸毛性状定位和抗虫机理研究 花后喷施脱落酸对下代小麦高温耐性的诱导效应研究 库源对稻米品质的影响及其生理机制 一个控制小麦旗叶宽主效 QTL 的精细定位 水旱轮作不同种植模式下农药投入调查及减量技术对策研究 水稻着粒密度 QTLqGn3 回交背景选择与定位研究 水稻种间杂种花粉不育基因的定位与图位克隆 我国南方 SMV 流行株系抗病基因的精细定位 以陆地棉 TM-1 为背景的达尔文氏棉染色体片段导入系的培育 水稻抗旱指标鉴定及耐旱品种筛选
连续进料条件下温度对餐厨垃圾厌氧发酵效率及发酵产物的影响 菜豆根瘤菌群体感应系统对其结瘤、固氮的影响 不同品种棉花纤维伸长与壁加厚过程的生长发育曲线分析及探索细胞壁标志性蛋白质 土壤耐铅、镉、铜微生物的分离筛选及其吸附机理研究 呋喃丹水解酶基因的克隆 一株胶质类芽孢杆菌 N342 的产生物絮凝剂特性研究 建立数学模型预测几种常见行道树的叶面积及其生长 烟草青枯病拮抗菌的筛选及抗菌物质的分离鉴定 霍乱弧菌致病性与趋化性相关研究 氯化血红素对盐胁迫下不同品种水稻种子萌发的影响 蜱螨体内螺原体的分离与特性研究 PHB 高产芽孢杆菌菌株的分离及其鉴定 粳型抗除草剂转基因水稻与籼、粳型杂草稻杂交 F3 代的遗传特性、杂草性、适合度研
学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助 学校资助
王梦琴 张淼燚 周洋 徐莹 王珺 高权 赵越 吴强 黄思宇 孙晔 王哲 孙前成 刘建兴
ห้องสมุดไป่ตู้
顾向阳 郑会明 刘康 何琳燕 洪青 崔中利 陆巍 沈标 王卉 谢彦杰 于汉寿 闫新 宋小玲
遗传学排名

071007 遗传学遗传学的研究范围包括遗传物质的本质、遗传物质的传递和遗传信息的实现三个方面。
遗传物质的本质包括它的化学本质、它所包含的遗传信息、它的结构、组织和变化等;遗传物质的传递包括遗传物质的复制、染色体的行为、遗传规律和基因在群体中的数量变迁等;遗传信息的实现包括基因的原初功能、基因的相互作用,基因作用的调控以及个体发育中的基因的作用机制等。
遗传学中的亲子概念不限于父母子女或一个家族,还可以延伸到包括许多家族的群体,这是群体遗传学的研究对象。
遗传学中的亲子概念还可以以细胞为单位,离体培养的细胞可以保持个体的一些遗传特性,如某些酶的有无等。
对离体培养细胞的遗传学研究属于体细胞遗传学。
遗传学中的亲子概念还可以扩充到DNA脱氧核糖核酸的复制甚至mRNA的转录,这些是分子遗传学研究的课题。
一个受精卵通过有丝分裂而产生无数具有相同遗传组成的子细胞,它们怎样分化成为不同的组织是一个遗传学课题,有关这方面的研究属于发生遗传学。
由一个受精卵产生的免疫恬性细胞能够分别产生各种不同的抗体球蛋白,这也是遗传学的一个课题,它的研究属于免疫遗传学。
从噬菌体到人,生物界有基本一致的遗传和变异规律,所以遗传学原则上不以研究的生物对象划分学科分支。
人类遗传学的划分是因为研究人的遗传学与人类的幸福密切相关,而系谱分析和双生儿法等又几乎只限于人类的遗传学研究。
微生物遗传学的划分是因为微生物与高等动植物的体制很不相同,因而必须采用特殊方法进行研究。
此外,还有因生产意义而出现的以某一类或某一种生物命名的分支学科,如家禽遗传学、棉花遗传学、水稻遗传学等。
更多的遗传学分支学科是按照所研究的问题来划分的。
例如,细胞遗传学是细胞学和遗传学的结合;发生遗传学所研究的是个体发育的遗传控制;行为遗传学研究的是行为的遗传基础;免疫遗传学研究的是免疫机制的遗传基础;辐射遗传学专门研究辐射的遗传学效应;药物遗传学则专门研究人对药物反应的遗传规律和物质基础,等等。
生命科学学院南京农业大学学生管理制度考试

25分)、、、、_________。
(含一天)的,提出书面申请,由或________批准(含一周)的,提出书面申请,由指定的批准;请假在一周以上的,提出书面申请由指定的签署意见,报批准。
3、学生缺席数达到某门课程一学期上课时数的(体育课每学期缺课者),需重修该课程。
4、根据本科人才培养方案要求,自2007级学生(除基地班学生外)起,本科学生应修满学分的公共选修课,其中文化素质教育类课程不少于学分。
5、违反宪法,反对四项基本原则、破坏安定团结、扰乱社会秩序者,给予___________处分;对组织、策划、参加非法游行的或者煽动闹事的,情节严重造成恶劣影响的,给予___________处分;对违反《治安管理处罚条例》,性质恶劣的,给予____________处分;对严重影响学习教育教学秩序、生活秩序,造成严重后果的,给予______________ 处分6、大学生社会实践一般安排在暑假期间,少数四年级学生可以安排在寒假期间进行,连续实践时间不得少于天.社会实践报告的规范化要求:(一)字数不少于字/篇。
(二)一律由学生本人用计算机。
(三)评阅教师应该按要求认真填写评阅意见、成绩并签名。
7、违章用电,私接电源的,除外,给予处分;造成严重后果的,给予处分。
8、计算机管理:(1)学生宿舍原则上放置计算机;(2)确因学习需要,需在宿舍内使用计算机的,必须向提出申请,并办理相关手续遵守有关规定.在学校规定的学习时间内(周一至周五上午8:00 -、下午14:00-、晚上18:30-),严禁使用计算机从事与无关的活动。
使用计算机,不得妨碍他人休息。
9、一学期内累计旷课学时以上尚不满学时的,视情节给予记过或留校察看处分;累计旷课达学时的,予以退学.10、学生宿舍卫生,每周定期检查_____次,抽查______次,检查结果予以公布并记录在册,同时折成相应分值计入学生个人综合素质测评总成绩中。
宿舍卫生检查包括_______________和______________两部分。
菠萝蛋白酶研究与应用

酶在预防和治疗单胃动物的腹泻, 尤其是在预防和治疗仔 。菠萝蛋白酶利用其蛋白 水解功能, 抑制病原菌株肠毒源性大肠杆菌在动物体内 , 尤其是肠道等处的受体活性 , 从而降低其在体内的粘附程 度。这对于种猪养殖的技术更新不无裨益。 5 1 2 在制药方面的应用 生物活性肽具有免疫调节、 抗菌、 抗肿瘤等多种生理活性作用。与氨基酸相比 , 它更 易被人体吸收, 并且可获得更高的生物效价和营养价值 , 是目前研究的热点之一。吴鑫颖等
压榨 去杂志 超滤浓缩 降温、 加有机溶剂、 沉淀
以啤酒废酵母为原
料, 经菠萝蛋白酶酶解、 分离、 纯化、 干燥等工序, 制得了啤 酒废酵母活性肽 , 并探索出了水解获得啤酒废酵母活性肽 的最佳条件。 5 1 3 在人体临床上的应用 菠萝蛋白酶作为药物 , 目 前大多情况下均是通过同其他物质按一定配比使用而实 现临床上最佳的治疗效果。扑尔敏是一类抗过敏药物, 当 其同菠萝蛋白酶混合使用时 , 能起到很好的抗炎效果 , 可
菠萝下脚料
冷冻干燥
汁液 酶制品
澄ቤተ መጻሕፍቲ ባይዱ液
浓缩液
湿酶
图 3 超滤法提取菠萝蛋白酶的工艺流程
[ 11 ]
防止因组织胺所引起的毛细血管通透性增高
[ 15]
; 庞然
[ 16]
4 菠萝蛋白酶的固定化技术
菠萝蛋白酶的固定化不仅能使其稳定性增强, 在长时 间内反复使用 , 而且能使底物与酶极易分开, 提高酶的使 用效率和产物得率 , 从而大大简化产物的提纯工艺, 还能 大幅降低生产成本。 固定菠萝蛋白酶的方法主要有吸附法、 交联法和共价 结合法等。吸附法无需借助特殊的化学试剂, 简便价廉, 但结合力是弱键作用 , 当环境中温度、 pH、 离子强度等改 变或底物存在时, 极易使其从载体上解析下来。交联法能 使低分子质量的底物通过扩散自由进入凝胶颗粒, 酶和高 分子量的终产物不能从凝胶颗粒中渗漏出去并且结合牢 固 , 但是所得的酶活低。共价结合法固定化酶结合牢固, 酶不易泄漏, 但是一部分酶会因起载体作用而失去催化能 力。故在实际应用中通常将交联法和吸附法联 合使用。 近年来 , 利用凹凸棒土作为载体实现菠萝蛋白酶的固定化 成为一种新兴技术。凹凸棒土结构内部含有大量的结构 水、 结晶水和吸附水。这一点对于作为酶载体 , 可为酶提 供一定的水分 , 对保持酶的结构完整性非常具有优势。另 一方面, 凹凸棒土作为一种无机矿物, 不仅价廉易得, 而且 具备了一般无机载体均具有的优点即机械强度高、 热稳定 性好、 抗有机溶剂、 抗微生物腐蚀等
生物化学检测技术

02
原理:琼脂糖是一种天然聚合长链状分子,可以形成具有刚性的滤孔,凝胶孔径的大小决定于琼脂糖的浓度。琼脂糖凝胶电泳法分离DNA,主要是利用分子筛效应,迁移速度与分子量的对数值成反比关系。因而就可依据DNA分子的大小使其分离。该过程可以通过把分子量标准参照物和样品一起进行电泳而得到检测。
电泳槽和大分子分离结果
电泳技术分类
从实际和实用出发的分类
根据分离样品样品的数量和目的分制备和分析电泳 根据结合配套的技术分免疫、层析、等电聚焦、转移、双向、脉冲梯度电场、垂直交替凝胶电泳等 根据使用电压分常压(<500)和高压电泳。 根据电泳系统pH连续与否分连续和不连续pH电泳 根据介质使用与否分自由和区带电泳(滤纸、薄层、凝胶电泳) 根据电泳槽的形式分有 垂直的、水平的、柱状的、板状的、毛细管的、湿小室及幕状的。 毛细管电泳是20世纪80年代研制出的一种新型的区带电泳方法,具有分辨率好、灵敏度高、检测快捷等特点。
带电颗粒在电场中泳动的速度称迁移率或泳动度。 电泳时,带电颗粒(球形)在介质中受力平衡匀速运动,则有: v = F阻/f = FE/f = E·q/f = E·q/(6π·r·η) v — 泳动度 F阻 — 颗粒所受阻力 FE — 颗粒所受电场力 f — 摩擦系数 由上式可见:泳动度与球形分子半径、介质粘度、颗粒所带电荷以及电场强度有关。 非球形分子(如线状DNA)在电泳过程中受到更大的阻力,即粒子的泳动度与粒子形状有关。
试剂与设备
操作方法
上样buffer
01
02
03
04
0.25%溴酚蓝 ,0.25%二甲苯青 ,40%蔗糖水溶液
0.25%溴酚蓝 ,0.25%二甲苯青 ,30%甘油水溶液
4离心技术

第三节 离心的基本方法
离心分离是制备生物样品广泛应用的重要手段。如分离活
体生物、细胞器、生物大分子、小分子聚合物等。
根据这个公式,相对离心力(RCF)和每分钟转数(N)
之间便可以互换,这种互换关系是很有实用价值的。
2019/1/7
6
南京农业大学 生命科学学院
第一节 离心技术原理
1. 相对离心力(RCF)
一般情况低速离心转速单位以 rpm 表示,高速离心则以重力加
速度g表示。 在计算颗粒的相对离心力时,应注意离心管与中心轴之间的距
(详见本页备注)
15 南京农业大学 生命科学学院
2019/1/7
高速离心机:
2019/1/7
16
南京农业大学 生命科学学院
超速离心机:
组成:
驱动和速度控制 温度控制 真空系统 转头
增加真空系统,这是它与高速离 心机的主要区别。 常用于分离亚细胞器、病毒粒子 、 DNA 、 RNA 和蛋白质分子 。 在分离时无须加入可能引起被分 离物质结构改变的物质。
ω — 粒子旋转的角速度 r — 粒子的旋转半径 Fc — 离心力
2019/1/7
5
南京农业大学 生命科学学院
第一节 离心技术原理
1. 相对离心力(RCF)
由于转头有不同的制造商制造,半径不同,故用相对离心
力来表示离心力的大小。 RCF (×g) = Fc/F重力 = mG/mg = r· ω2/g = 1.119×10-5· N2· r =11.2r(N/1000)2 也有:
缺点是样品和介质直接接触转头,耐腐蚀要求高,操 作复杂。
γ-氨基丁酸对高温胁迫下高羊茅耐热性的调控

γ-氨基丁酸对高温胁迫下高羊茅耐热性的调控
夏倩倩;甘立军
【期刊名称】《生物学杂志》
【年(卷),期】2018(035)006
【摘要】以高羊茅爱瑞3号为试验材料,采用γ-氨基丁酸(GABA,25、50、100、200和400 mg/L)预处理,研究外源γ-氨基丁酸对高温胁迫下高羊茅耐热性的影响.结果表明:在42℃/30℃(昼/夜)的高温胁迫下,喷施不同浓度的GABA能够缓解高温对高羊茅幼苗的伤害.进一步的探究表明,高温胁迫下喷施外源GABA不仅能够降低幼苗中丙二醛(MDA)和H2 O2的积累,还能增强幼苗抗氧化酶系统中超氧化物歧化酶(SOD)、过氧化物酶(POD)和抗坏血酸过氧化物酶(APX)的活性.同时,还能够提高幼苗体内可溶性蛋白含量以及降低幼苗体内可溶性糖含量和叶片外渗液的相对电导率.当GABA的喷施浓度为100 mg/L时,效果最佳.
【总页数】5页(P50-54)
【作者】夏倩倩;甘立军
【作者单位】南京农业大学生命科学学院,南京210095;南京农业大学生命科学学院,南京210095
【正文语种】中文
【中图分类】Q945.78
【相关文献】
1.高温胁迫下修剪高度对高羊茅生长的影响 [J], 陈煜;张礼军;杨志民;李志华
2.不同氮源对高温胁迫下高羊茅抗氧化酶活性的影响 [J], 李良霞;李建龙;王艳;潘永年;李高扬;图雅
3.高温胁迫下草坪草高羊茅差异表达基因的分子研究 [J], 李卉;王艳;王海宏;梁珂珂;李建龙
4.高温胁迫下高羊茅生理生化特性研究 [J], 张庆峰;徐胜;李建龙
5.高温胁迫下不同氮肥处理对凌志高羊茅耐热性的调控效果 [J], 薛峰;李建龙;李良霞;王艳
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2014-6-7
第二节 乳糖操纵子
1961年,Jacob和Monod提出了操纵子模型,这是与特殊 代谢途径有关的基因转录的协同调控模型。
操纵子是基因表达和调控的单元,典型的操纵子包括:
结构基因(除调节基因以外的所有基因),编码那些在 某一特定的生物合成途径中起作用的、其表达被协同调 控的酶。 调控元件,如操纵序列,是调节结构基因转录的一段 DNA序列。
2014-6-7
20
南京农业大学 生命科学学院
第二节 乳糖操纵子
1. 酶的诱导——lac体系受调控的证据
用 32P 标记的 mRNA 与模板 DNA 进行定量分子杂交,表明 培养基中加入乳糖 1 ~ 2分钟后,编码 β- 半乳糖苷酶和透过 酶的lac mRNA量就迅速增加,去掉乳糖后,量立即下降。
属于这种调节方式的有:大肠杆菌中的色氨酸操纵子、苯
丙氨酸操纵子、苏氨酸操纵子、异亮氨酸操纵子和缬氨酸
操纵子以及沙门氏菌的组氨酸操纵子和亮氨酸操纵子、嘧
啶合成操纵子等等。
2014-6-7 14 南京农业大学 生命科学学院
第一节 原核基因表达调控总论
4. 降解物对基因活性的调节
有葡萄糖存在的情况下,即使在细菌培养基中加入乳糖、 半乳糖、阿拉伯糖或麦芽糖等诱导物,与其相对应的操纵 子也不会启动,不会产生出代谢这些糖的酶来,这种现象 称为葡萄糖效应或称为降解物抑制作用。
因表达调控(gene regulation,gene control)。
基因调控是现阶段分子生物学研究的中心课题。
2014-6-7 7 南京农业大学 生命科学学院
第一节 原核基因表达调控总论
基因表达调控主要表现在以下二方面: 转录水平上的调控 转录后水平上的调控: mRNA加工成熟水平调控 翻译水平调控 不同的生物使用不同的信号来指挥基因调控。
2014-6-7
21
南京农业大学 生命科学学院
第二节 乳糖操纵子
1. 酶的诱导——lac体系受调控的证据
实验室常用两种乳糖类似物 —— 异丙基巯基半乳糖苷( IPTG)和巯甲基半乳糖苷(TMG),在酶活性分析中常用 发色底物 O- 硝基半乳糖苷( ONPG )。因为它们都不是半
乳糖苷酶的底物,所以又称为安慰性诱导物。
原核生物中,营养状况和环境因素对基因表达起着举足 轻重的影响。 在真核生物尤其是高等真核生物中,激素水平和发育阶 段是基因表达调控的最主要手段,营养和环境因素的影 响力大为下降。
8 南京农业大学 生命科学学院
2014-6-7
第一节 原核基因表达调控总论
在转录水平上对基因表达的调控决定于DNA的结 构、RNA聚合酶的功能、蛋白因子及其他小分子 配基的相互作用。
区(O)之间,不能单独起始半乳糖苷酶和透过酶基因的
高效表达。
操纵区是 DNA上的一小段序列(仅为26bp),是阻遏物 的结合位点。
2014-6-7
23
南京农业大学 生命科学学院
当阻遏物与操纵区相结合时, lacmRNA 的转录起始受 到抑制。 诱导物通过与阻遏物结合,改变其三维构象,使之不能 与操纵区相结合,诱发lac mRNA的合成。
在负转录调控系统中,调节基因的产物是阻遏蛋白 (repressor)。根据其作用特征又可分为负控诱导系统和 负控阻遏系统二大类。
在正转录调控系统中,调节基因的产物是激活蛋白 (activator) 。也可根据激活蛋白的作用性质分为正控诱 导系统和正控阻遏系统。
阻遏蛋白 转录激活 转录抑制 负控诱导 负控阻遏
当氨基酸饥饿时,细胞中便存在大量的不带氨基酸的 tRNA, 这种空载的tRNA会激活焦磷酸转移酶,使ppGpp大量合成。 ppGpp 的 出 现 会 关 闭 许 多 基 因 , 以 应 付 这 种 紧 急 状 况 。 ppGpp 影响RNA聚合酶与这些基因转录起始位点的结合,使 基因被关闭。 ppGpp与pppGpp的作用范围十分广泛,它们影响一大批操纵 子而被称为超级调控因子。
第七章 基因的表达与调控(上)
细菌中所利用的大多数基本调控机制一般执行如下
规律:一个体系在需要时被打开,不需要时被关闭。这
种“开 - 关”( on-off )活性是通过调节转录来建立的
,也就是说 mRNA 的合成是可以被调节的。实际上, 当我们说一个系统处于“off ”状态时,也可能有本底 水平的基因表达,常常是每世代每个细胞只合成 1 或 2 个 mRNA 分子和极少量的蛋白质分子。为了方便,我
解释:在没有诱导物的情况下,转运酶和β-半乳糖苷酶仍有 本地水平的表达。阻遏物的结合并非绝对紧密,偶尔会掉下 ,使得转录可以进行。表达量足以使诱导过程得以启动。
2014-6-7
25
南京农业大学 生命科学学院
第二节 乳糖操纵子
2.2. 大肠杆菌对乳糖的反应
在以甘油为碳源的培养基中 加乳糖以前, lac 操纵子本底 水平表达 加乳糖后,阻遏物失活, mRNA大量表达 乳糖耗尽,阻遏物浓度逐渐大 于异构乳糖,阻遏状态重新形 成,mRNA水平下降。 β-半乳糖苷酶半衰期长,其活 性下降滞后。
2014-6-7
24
南京农业大学 生命科学学院
第二节 乳糖操纵子
2.1. lac操纵子的本底水平表达
两个矛盾:
诱导物需要穿过细胞膜才能与阻遏物结合,而转运诱导物 需要转运酶,转运酶的合成有需要诱导。
人们发现乳糖并不与阻遏物相结合,真正的诱导物是异构
乳糖,而后者是在β-半乳糖苷酶的催化下由乳糖形成的。
白质。
另 一 类 则 被 称 为 适 应 型 或 调 节 型 ( adaptive or regulated),因为这类蛋白质的合成速率明显地受环境的 影响而改变。如大肠杆菌细胞中一般只有 15 个 β- 半乳糖苷 酶,但若将细胞培养在只含乳糖的培养基中,每细胞中这个 酶的量可高达几万个分子。
2014-6-7 4 南京农业大学 生命科学学院
13 南京农业大学 生命科学学院
2014-6-7
第一节 原核基因表达调控总论
3. 弱化子对基因活性的影响
在这种调节方式中,起信号作用的是有特殊负载的氨酰 tRNA的浓度,在色氨酸操纵子中就是色氨酰 -tRNA的浓度 。当操纵子被阻遏, RNA合成被终止时,起终止转录信号 作用的那一段DNA序列被称为弱化子。
2014-6-7
18
南京农业大学 生命科学学院
第二节 乳糖操纵子
P为启动子,O为操纵区,lacI编码阻遏子
3个结构基因各决定一种酶: Z编码β-半乳糖苷酶; Y编码 β-半乳糖苷透过酶;A编码β-半乳糖苷乙酰基转移酶。键的专一性酶,除能将 乳糖水解成葡萄糖和半乳糖外,还能水解其他β-半乳糖 苷(如苯基半乳糖苷)。 β-半乳糖苷透过酶的作用是使外界的 β-半乳糖苷透过大 肠杆菌细胞壁和原生质膜进入细胞内。 β-半乳糖苷乙酰基转移酶的作用是把乙酰辅酶 A上的乙 酰基转移到-半乳糖苷上,形成乙酰半乳糖。
降解物抑制作用是通过提高转录强度来调节基因表达的,
是一种积极的调节方式。
2014-6-7
15
南京农业大学 生命科学学院
第一节 原核基因表达调控总论
5.细菌的应急反应
细菌有时会碰到紧急状况,比如氨基酸饥饿 —— 氨基酸的 全面匮乏。细菌会产生一个应急反应 —— 停止包括生产各 种RNA、糖、和蛋白质的几乎全部生物化学反应过程。 实施这一应急反应的信号是鸟苷四磷酸 (ppGpp) 和鸟苷五 磷酸(pppGpp)。产生这两种物质的诱导物是空载tRNA。
2014-6-7
12
南京农业大学 生命科学学院
第一节 原核基因表达调控总论
2. 原核基因调控的主要特点:
原核生物通过特殊代谢物调节的基因活性主要分为可诱导 和可阻遏两大类:
可诱导调节。是指一些基因在特殊的代谢物或化合物的 作用下,由原来关闭的状态转变为工作状态,即在某些 物质的诱导下使基因活化。这类基因中最突出的例子是 大肠杆菌的乳糖操纵子。 可阻遏调节。这类基因平时都是开启的,处在产生蛋白 质或酶的工作过程中,由于一些特殊代谢物或化合物的 积累而将其关闭,阻遏了基因的表达。比如大肠杆菌中 的色氨酸操纵子。
调节基因,其产物能够识别调控元件,例如阻抑物,可 以结合并调控操纵基因序列。
17 南京农业大学 生命科学学院
2014-6-7
第二节 乳糖操纵子
大肠杆菌能利用乳糖作为碳源,而利用乳糖作为碳源的酶 只有当乳糖成为惟一的碳源时才会被合成。 大肠杆菌乳糖操纵子( lactose operon)包括3个结构基 因:Z、Y和A,以及启动子、控制子和阻遏子等。转录的 调控是在启动区和操纵区进行的。
们常常使用“off ”这一术语,但必须明白所谓“关”
实际的意思是基因表达量特别低,很难甚至无法检测。
2014-6-7 5 南京农业大学 生命科学学院
第七章 基因的表达与调控(上)
内容: 1. 原核基因表达调控总论 2. 乳糖操纵子与负控诱导系统 3. 色氨酸操纵子与负控阻遏系统 4. 其他操纵子 5. 转录水平上其它调控 6. 转录后调控
2014-6-7
6
南京农业大学 生命科学学院
第一节 原核基因表达调控总论
随着生物个体的发育, DNA 分子能有序地将其
所承载的遗传信息,通过密码子-反密码子系统转变
成蛋白质,执行各种生理生化功能。
科学家把从 DNA 到蛋白质的过程称为基因表达
(gene expression) ,对这个过程的调节就称为基
分 子 生 物 学
南京农业大学 生命科学学院
第七章 基因的表达与调控(上)