5-氮杂胞苷对小麦生长发育及 DNA 甲基化的影响
DNA甲基化在肿瘤发生中的作用

DNA甲基化在肿瘤发生中的作用肿瘤是人类健康的头等大事,它是由基因突变和表观遗传学变化引起的遗传疾病。
DNA甲基化是一种常见的表观遗传学变化,它是指DNA分子在胞内繁殖时,通过在在五碳脱氧核糖核苷酸的C5位加上一个甲基基团而产生的一种修饰,它在正常组织中具有调控基因表达,维护基因稳定性,参与细胞分化和应答外源性刺激等多种功能。
但是,在肿瘤发生中,DNA甲基化的模式发生改变,造成癌基因的高度表达或肿瘤抑制基因的沉默,这种表观遗传学的改变往往会引起肿瘤的发生和发展。
因此,深入了解DNA甲基化在肿瘤发生中的作用,对于治疗肿瘤有着重要的意义。
DNA甲基化的机制DNA甲基化是一种简单的化学修饰,它是由甲基转移酶催化丙烷基单元(C1)从S-腺苷甲硫氨酸(AdoMet)转移到细胞内DNA链合成过程中的胞嘧啶(Cyt)的C5核苷酸上。
DNA甲基转移酶(DNMT)是DNA甲基化的关键酶,它包括DNMT1, DNMT3a和DNMT3b三个亚型。
DNMT1是在细胞分裂期间能够保证分子和细胞的遗传稳定性,通过识别和甲基化前一代细胞从父本获得的甲基化DNA,维持其在细胞分裂后的遗传稳定性。
DNMT3a/b通过识别新的DNA序列元素来甲基化胞苷。
然而,过度的DNA甲基化也可能触发继承性的表观遗传学改变,从而引起肿瘤的发生。
DNA甲基化对于肿瘤的发生和发展,具有重要的作用。
它可以通过多种方式参与调节肿瘤细胞的基因表达和功能。
首先,DNA甲基化可以引起癌的基因高度表达,包括促细胞分裂和生长的基因和转录激活因子。
例如,在结肠直肠癌和胃癌中,印迹基因CDKN2A的启动子区域的甲基化状态的改变被认为是这些肿瘤的重要驱动因素。
此外,在癌症中经常出现的促细胞分裂和生长信号通路基因的DNA甲基化也是引起癌症的重要机制之一。
其次,DNA甲基化还可压制肿瘤抑制基因的表达。
肿瘤抑制基因损失或其功能异常的情况下,细胞将失去对癌症的抵抗能力。
例如,在人类胃癌和乳腺癌中,肿瘤抑制基因BRCA1的基因沉默与BRCA1启动子区域的甲基化增加有关联。
植物DNA甲基化研究进展
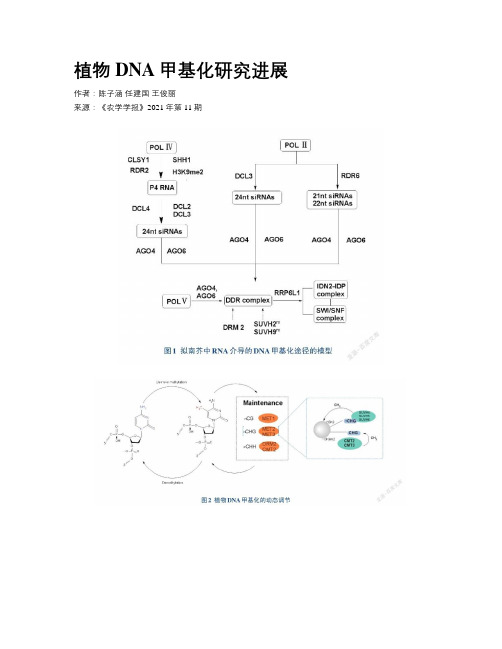
植物DNA甲基化研究进展作者:陈子涵任建国王俊丽来源:《农学学报》2021年第11期摘要:DNA甲基化是一種重要的表观遗传修饰,能够有效调控基因组稳定性。
为了了解DNA甲基化对植物生长发育的影响,本文归纳了近年来植物DNA甲基化的模式,总结了植物DNA甲基化的生物学功能,概括了DNA甲基化的研究方法,最后总结了植物DNA甲基化研究中存在的问题,并指明了研究方向,为后续植物基因组研究提供理论依据。
关键词:植物;DNA甲基化;表观遗传;修饰;生长发育;逆境胁迫;基因组;稳定性中图分类号:S184文献标志码:A论文编号:cjas2020-0152Research Advances on Plant DNA MethylationChen Zihan, Ren Jianguo, Wang Junli(School of Public Health, the key Laboratory of Enviromental Pollution Monitoring and Disease Control,Ministry of Education, Guizhou Medical University, Guiyang 550025, Guizhou, China)Abstract: DNA methylation is an important epigenetic modification that can effectively regulate genome stability. In order to understand the impact of DNA methylation on plant growth and development, this article summarizes plant DNA methylation patterns, concludes the physiological functions of plant DNA methylation, and reviews the research methods of DNA methylation. At last, this article sums up the problems in the study of plant DNA methylation and points out the research directions in the future, providing a theoretical basis for subsequent plant genome research.Keywords: Plants; DNA methylation; Epigenetic; Modification; Growth and Development; Adversity Stress; genome; stability0引言DNA甲基化(DNA methylation)是目前表观遗传学研究较为清晰的机制之一,广泛存在于生物界中,是真核细胞中最为常见的一种基因组修饰方式,它在调节基因组功能的同时不改变DNA的碱基序列。
DNA甲基化对小麦盐胁迫应答基因的影响研究开题报告

DNA甲基化对小麦盐胁迫应答基因的影响研究开题报告一、研究背景小麦是世界上最主要的粮食作物之一,可以种植在全球各地的干旱、高温、低温等极端环境中。
然而,盐胁迫是导致小麦减产的主要因素之一。
在盐胁迫下,小麦的生长和发育受到明显的抑制,其中许多基因起着重要的作用。
DNA甲基化是一种影响基因表达和细胞分化的基本遗传机制。
越来越多的研究表明,DNA甲基化对植物的生物逆境反应和耐受性有很大的影响。
然而,到目前为止,DNA甲基化对小麦盐胁迫应答的影响尚不清楚。
因此,本研究旨在探讨DNA甲基化在小麦盐胁迫应答基因中的作用,为小麦抗逆育种提供理论依据和数据支持。
二、研究内容和方法1. 研究目标探究DNA甲基化对小麦盐胁迫应答基因的影响。
2. 研究内容(1)提取小麦盐胁迫处理前后的基因组DNA,并进行高通量测序。
(2)分析小麦盐胁迫前后基因的甲基化水平及变化。
(3)筛选出与小麦盐胁迫应答相关的甲基化基因。
(4)构建甲基化基因与非甲基化基因的表达模型,比较其表达差异,并验证差异基因是否与盐胁迫应答相关。
3. 研究方法(1)实验材料:小麦普通小麦“高粱18”品种。
(2)盐胁迫处理:在12~14天生育期,将小麦盆栽浸泡在含5% NaCl的水中,经过48小时后取样。
(3)DNA甲基化测序:采用Bisulfite-Seq方法,对小麦盆栽的基因组DNA进行甲基化测序,并对比分析盐胁迫前后甲基化水平的变化。
(4)全转录组测序:采用RNA-Seq方法,对小麦盆栽在盐胁迫前后的转录组进行测序,并将测序结果与甲基化数据进行比对和分析。
(5)数据分析:使用R软件及相关生物信息学数据库,对测序结果进行基因差异分析、聚类分析、通路富集分析等。
三、预期结果和意义1. 预期结果(1)比较盐胁迫前后小麦基因组的甲基化水平,筛选出与盐胁迫应答相关的甲基化基因。
(2)构建甲基化基因与非甲基化基因的表达模型,比较其表达差异,并验证差异基因是否与盐胁迫应答相关。
4.2基因表达与性状的关系-表观遗传教学设计2021-2022学年高一下学期生物人教版必修2

4.2《基因表达与性状的关系-表观遗传》教学设计-人教版高中生物(2019)必修二教材分析《表观遗传》选自高中生物人教版教材必修二《遗传与进化》第 4章第 2节。
本节课内容为现代遗传学研究的前沿内容,与人类生活较密切的部分相关实例已得到了清晰阐释,如柳穿鱼花的花色遗传,小鼠的毛色遗传等,通过对其遗传物质的研究最终得到“存在基因中碱基序列不变但表型改变的表观遗传现象”的概念。
在此基础上延申甲基化和组蛋白修饰等表观遗传修饰在细胞分化过程中的作用,细胞癌变和诱导多动能干细胞过程中的表观遗传修饰改变现象。
从教材本节课的安排来看,前期课程先介绍了基因表达的过程,再描述了基因表达与性状的关系,最后讲表达过程中进行影响的表观遗传现象,体现了教材编写的逻辑性,从微观到宏观,从机制到表现,符合学生的认知规律。
本节课的重点在于影响基因表达的因素,表观遗传的特点、表观遗传的实例、基因与表达产物、基因与环境之间的相互作用对生物体性状产生何种影响。
能力要求更加偏重于对现象的解释和机制的探究。
学情分析本节内容安排在经典遗传学之后,目的是通过“细胞分化的本质是基因选择性表达的结果”和“表观遗传机制控制基因表达”两部分内容的学习,加深对遗传信息传递和表达过程的理解,构建完整的生物学大概念,为形成遗传与进化的生命观念做好分子层面的铺垫。
学生之前已经掌握了经典遗传学,以及基因对性状对常规控制,已经有了一定的学习基础。
作为课标和教材新加入的教学内容,“表观遗传”的引入是最新研究成果进入高中课堂的生动体现,也是本节课内容的重点和难点。
为突破重点和难点,本节课以问题为导向,充分考虑学生的接受能力,以科学研究证据为资料,从讨论父母给的遗传信息都一样么,以及“细胞分化”与“基因表达”的关系入手,分析科学研究数据,概述细胞的分化是基因选择性表达的结果;然后提出新问题:“基因选择性表达是如何调控的?”以科学研究进展为材料,构建表观遗传的概念并总结出表观遗传的调控方式;再以现实问题创设情境,引导学生在层层递进真实情境中解决问题,符合学生的认知规律,促进生物学学科核心素养的落实。
丁酸钠和5-氮杂胞苷对大麦、水稻、玉米、小麦种子萌发及芽苗期生

麦、 水稻 、 玉米及小麦进行处 理 , 统 计种子萌发率 、 幼 苗株 高 、 根系发 育情况 以及植株 的干质量 等指标 , 系统 比较 这 4种禾
本科作物对丁酸钠和 5一 A C等 2 种表观遗传试剂处理的敏
感性差异 , 为研究 D N A去 甲基化 、 组蛋 白乙酰化水平 升高对
发育 、 非生物逆境抗性 等方面起着 关键作用 … , 对油菜 、 大
1 . 2 试 验 方 法
选取饱满的种子 , 用7 5 % 的 乙醇消毒 3 0 S , 清水润洗 后
下降 , 且其表 型效应 与处 理浓度相 关 。丁酸钠处理 能促进
水稻花培绿苗 的形 成 , 但 不能 显著 提 高大麦 苗 期 的耐湿
性 。与 5一A C相 比 , 关 于 丁 酸 钠 对 种 子 萌 发 和 幼 苗 发 育 影 响还知之甚少 , 缺 乏 丁 酸 钠 与 5一A C对 作 物 种 子 萌 发 及 幼 苗 生 长 效 应 的系 统 对 比研 究 。
供试 材 料 为 大 麦 苏 啤 3号 ( H o r d e u m口 “ a r e L .C V . S u p i 3 ) 、 水稻丰美 占( O r y z a s a t i v a L .C V .F e n g m e i z h a n ) 、 玉米 郑单 9 5 8( Z e a m a y s L .C V .Z h e n g d a n 9 5 8 ) 、 小麦湘麦 1 0号 ( T r i t i c u m a e s t i v u m L . e v .X i a n g ma i l O ) 。供试大麦 、 水稻 、 小麦 材料 由笔者所在单位保存 , 玉米购于种子市场 。
胞苷 ( 5一a z a c y t i d i n e , 简称 5一 A c) 溶液对 大麦 、 水稻 、 玉米和小麦 等 4种作物种 子萌发 和幼 苗生长 发育 的影 响。结果 表明 , 丁酸钠 、 5一A c处理均 可抑制种子萌发 、 植株 株高及根长的生长 。受 害指数分析结果表 明, 4种作物对 5一A C处
5-氮杂胞苷对SMMC-7721细胞增殖与侵袭的影响及机制探讨的开题报告

5-氮杂胞苷对SMMC-7721细胞增殖与侵袭的影响及机制探讨的开题报告开题报告:一、研究背景肝癌是全球范围内常见的一种恶性肿瘤,预计在21世纪中肝癌仍将是全球死亡率最高的癌症之一。
目前,尽管在治疗方面取得了一定的进展,但肝癌的治疗仍然面临巨大挑战。
因此,需要寻找新的治疗方法和药物。
5-氮杂胞苷是一种新型的核苷酸类似物,具有抗肿瘤活性。
研究表明,5-氮杂胞苷能够抑制多种恶性肿瘤的细胞增殖、促进肿瘤细胞凋亡,但对于SMMC-7721的作用机制研究较为缺乏。
二、研究目的本研究的目的是探讨5-氮杂胞苷对SMMC-7721细胞增殖和侵袭的影响以及其可能的作用机制,为其在肝癌治疗中的应用提供理论和实践基础。
三、研究内容与方法研究内容:1. 评估5-氮杂胞苷对SMMC-7721细胞增殖和侵袭的影响;2. 探究5-氮杂胞苷对SMMC-7721细胞周期和凋亡的影响;3. 调查5-氮杂胞苷对SMMC-7721恶性转移相关分子、细胞信号通路和相关受体的影响。
研究方法:1. 采用MTT法检测肝癌SMMC-7721细胞的生长抑制率和半数抑制浓度(IC50);2. 采用Transwell姜筛法评估细胞侵袭和迁移能力;3. 利用PI染色和Annexin V-FITC/PI双染法检测细胞周期和凋亡的变化;4. 采用Western blot技术检测5-氮杂胞苷对SMMC-7721细胞恶性转移相关分子、细胞信号通路和相关受体的影响。
四、研究预期结果本研究预计将获得5-氮杂胞苷对SMMC-7721细胞增殖和侵袭方面的新实验结果,以及其在细胞周期、凋亡和信号转导通路等方面的作用机制。
结果有望为临床治疗提供新的靶向肝癌细胞的治疗方法和药物。
五、研究意义本研究将填补关于5-氮杂胞苷对SMMC-7721细胞作用的缺失,并为肝癌的治疗提供新的思路。
同时,本研究将丰富关于5-氮杂胞苷的作用机制的认识,为进一步应用5-氮杂胞苷的研究提供理论和实践基础。
江西省南昌市第三中学2024-2025学年高三上学期10月月考生物学试题(含答案)

南昌三中 2024—2025 学年度上学期 10 月考试高三生物试卷一、单选题(本大题共 12 小题,每题 2 分,共 24 分)1.刚挤出的鲜牛奶加热后有层“厚厚的奶皮 ”,奶皮的组成一般为:蛋白质10%~20%,脂肪40%~50%,水分10%以下,其余为碳水化合物和矿物质(钙、铁、锌、磷、钾等)。
下列相关叙述错 误的是 ( )A. 奶皮中含多种蛋白质,不一定都由 21 种氨基酸组成B. 奶皮中的碳水化合物最可能含乳糖,不能被细胞直接吸收C. 奶皮中的脂肪,因其氧原子含量高成为细胞中良好的储能物质D. 奶皮中的铁、锌属于微量元素,缺铁导致血液运输氧气的能力下降2.氮元素是植物必需的营养元素之一,植物根系从土壤中吸收的硝酸根(NO −3)是植物体内氮元 素的主要来源。
根系吸收NO −3依赖于转运蛋白(NRT1.1),蛋白激酶 CIPK23 是调控 NRT1.1 蛋白 磷酸化状态的关键酶,可引起 NRT1.1 第 101 位苏氨酸(T101)磷酸化,从而引起 NRT1.1 结构的改变,促进根细胞吸收NO −3 。
图甲、乙为不同浓度的NO −3时,根细胞对NO −3的吸收过程图解。
下 列分析正确的是 ( )A. 低浓度的NO − 3可引起 CIPK23 磷酸化,加速细胞吸收NO −3B. NRT1.1 与NO −3高低亲和性的转换与其蛋白结构的变换有关C. NRT1.1 基因的突变,若不影响 T101 磷酸化,则不会影响NO −3的运输D. 推测 NRT1.1 吸收NO −3时可能消耗能量,CIPK23 会抑制根细胞吸收NO −33.2017 年初,我国科学家完成了酿酒酵母 16 条染色体中的 4 条的人工合成,开启了人类“设计生命、再造生命和重塑生命 ”的新纪元。
下列有关分析正确的是( )A. 正常情况下,酿酒酵母细胞可能会出现 32 个以上的 DNA 分子B. 姐妹染色单体中含一个亲代 DNA 分子和一个子代 DNA 分子C. 人工合成染色体,需要核苷酸、氨基酸、磷脂和 ATP 等原料D. 酵母菌细胞有复杂的生物膜系统,氨基酸的脱水缩合在此进行4.人体消化道中消化淀粉的酶有多种,胰淀粉酶是其中一种。
5-氮杂胞苷(5-azaC)对盐胁迫下小麦幼苗生理特性及DNA甲基化的影响

5-氮杂胞苷(5-azaC)对盐胁迫下小麦幼苗生理特性及DNA甲基化的影响5-氮杂胞苷(5-azaC)对盐胁迫下小麦幼苗生理特性及DNA甲基化的影响引言:盐胁迫是世界范围内对农作物生产产生不利影响的主要因素之一。
小麦是世界上主要的粮食作物之一,在盐胁迫条件下的生长受到很大限制。
研究发现,DNA甲基化可以影响基因的表达,进而影响植物对盐胁迫的响应。
5-氮杂胞苷(5-azaC)是一种常用的DNA甲基化抑制剂,通过抑制DNA甲基转移酶的活性,可以改变DNA甲基化状态,并对植物的生理特性产生影响。
本研究旨在探究5-氮杂胞苷对盐胁迫下小麦幼苗生理特性及DNA 甲基化的影响。
材料与方法:实验使用小麦(Triticum aestivum L.)幼苗作为研究对象。
将小麦种子在灭菌状态下播种于含有不同浓度盐溶液的培养基中,同时加入不同浓度的5-azaC。
通过观察小麦幼苗的形态特征、根长、叶绿素含量、活性氧含量和DNA甲基化水平的变化,来研究5-azaC对盐胁迫下小麦幼苗的影响。
结果与讨论:在盐胁迫条件下,小麦幼苗受到了明显的抑制,生长缓慢且发育异常。
然而,与对照组相比,添加5-azaC后,小麦幼苗的生长状况有所改善。
根长显著增加,植株形态更加健壮。
此外,叶绿素含量也有所增加,表明叶绿素的合成得到了促进。
活性氧含量的测定结果显示,5-azaC能够显著减少盐胁迫引起的氧化应激。
这表明5-azaC可能通过调节氧化还原平衡,减轻了盐胁迫对小麦幼苗的伤害。
进一步的研究发现,5-azaC还能够影响小麦幼苗的DNA甲基化水平。
在盐胁迫的条件下,小麦幼苗的DNA甲基化水平显著增加。
然而,添加5-azaC后,DNA甲基化水平明显下降。
这说明5-azaC通过抑制DNA甲基转移酶的活性,改变了DNA的甲基化状态,进而影响了基因的表达,提高了小麦幼苗对盐胁迫的耐受性。
结论:本研究结果表明,5-氮杂胞苷(5-azaC)可以改善盐胁迫引起的小麦幼苗生长受限情况。
DAC

5-杂氮胞苷(5-aza-dC)是一种特异性甲基转移酶抑制剂,作用机制是与DNA 甲基转移酶共价结合,降低酶的生物活性,它可以降低DNA 甲基化水平并使得染色体的结构松散,从而影响基因的表达。
本文研究结果说明,通过5-aza-dC 降低外源基因的甲基化水平后,基因的表达水平明显提高。
说明降低外源基因的甲基化水平是提高外源基因表达水平的有效手段。
大量研究均说明外源基因在宿主染色体中的整合位点会影响到这个基因的表达水平。
如果外源基因整合到染色体的疏松区域,这个基因将会处于一个相对活化状态,表现出较高的表达水平,相反,如果这个外源基因整合到染色体的致密区域或异染色质区,基因将会被失活,表达水平将比较低甚至发生沉默现象。
实验中虽然说明了5-aza-dC 可以通过降低RP 启动子的甲基化水平来提高外源基因的表达水平,但对不同的细胞克隆,5-aza-dC 的作用不完全相同,说明5-aza-dC 的作用还受到外源基因在宿主染色体中的整合位点的影响。
整合在不同位点的外源基因对5-aza-dC 的敏感性不同。
氮杂胞苷L1210、AK小鼠淋巴细胞白血病和ECA等均有明显抗肿瘤作用,是主要作用于S期周期特异性药。
其抗肿瘤作用与其他嘧啶类拮抗剂不同,与嘧啶核苷酸代谢并无关系,而是以伪代谢物身份替代胞嘧啶搀入DNA及RNA,干扰DNA、RNA的生理功能而产生细胞毒作用及抗肿瘤作用。
近年来更注意到氮杂胞苷可搀人DNA甲基化过程。
许多基因的转录功能与嘧啶以甲基化状态密切相关,在整个细胞分裂过程中,通过甲基转移酶的作用,这种甲基化作用始终保持无缺,氮杂胞苷由于可搀入DNA,抑制DNA甲基化,因此影响基因的表达及分化。
但是氮杂胞苷并不是影响所有基因的表达,而是对某些基因的表达有影响,对另外一些基因的表达则无影响,原因不明。
对这一机制的探讨,亦可能会产生新的治疗上的重要意义。
本品静脉注射后,血中半衰期为3~5h,略长于阿糖胞苷,于48H内排出85%的药物。
甲基化抑制剂5-杂氮胞苷对T淋巴细胞株程序性死亡受体-1基因启动子区域甲基化水平及其表达的影响

[ 摘要 ] 目的 : T淋 巴细胞株 Mo 4 细胞为模 型 , 以 h 探讨 甲基化抑制 剂 5杂 氮胞苷 ( - ayii ,-a ) 一 5a ct n 5Z c z de
对 淋 巴细 胞 表 面 程 序 性 死 亡 受 体 一( rga m dda eet , D 基 因启 动 子 的 去 甲基 化 作 用及 其 诱 导 1 porm e et rcpo 1 P 一) h r
,
J ANG nga g I Yo f n ,XU Yun ,GONG o h n Gu z o g
( . eatetff cos i ae, eo dXag aH si l C nrl ot n e i , h nsa40 1 ; 1D p r n o t u Ds s Scn i y o t , et uhU i r t C a gh 10 m i e s n pa aS v sy 1
中南大学学报 ( 医学版 ) .c n Su^ 西 ( ds 2 1 ,6 1 ) , ef o t c) 0 13 ( 2
ht:/ w cu e . ;h p:/b x ・ 。 t / w w・sm d 。g / xy ’y “ p
16 l3
・
AR I L S・ TC E
Efe to e h l to nh b t r o m e h l to f c fm t y a i n i i io n de t y a i n
pa t r ft e PD- e e i r m o e e i n a d te n o h l g n n p o t r r go n
的 P — 因表达 的改 变, D 基 并进一步研 究去 甲基化作 用与 肋 一 因表达之 间的关 系。方法 : J基 以不 同浓度的 5 一 Z c分组( m lL组 、 i lL组 、0 p lL组 ) 用于体 外培 养的 M l4细胞 7 , a 0 o / 5x / mo 1  ̄ / mo 作 ot - 2h 流式 细胞仪 (l y f w c— o
浙江省杭州市学军中学西溪校区2021学年高二下学期中考试生物试题

浙江省杭州市学军中学西溪校区2021学年高二下学期中考试生物试题1.下列关于组成细胞的物质的叙述正确的是()A.Mg 2+是所有光合色素合成所必需的B.补充I -可预防甲亢D.磷脂不参与调节生命活动C.蛋白质的空间结构改变,则一定变性失活2.下列关于细胞结构的叙述正确的是()A.大量碱性物质进入溶酶体不会改变其中酶的活性B.观察叶绿体形态时应选用黑藻基部成熟叶片C.高倍显微镜下可见叶绿体有双层膜结构D.囊泡运输过程中存在膜成分的更新3.下列相关叙述错误的是()A.ATP的合成是一个吸能反应,与此同时必定正在发生一个放能反应B.植物细胞吸水膨胀达到平衡状态时,细胞液浓度大于细胞外溶液浓度C.转运蛋白在跨膜运输物质时会发生形变D.植物根尖分生区细胞较难发生质壁分离现象4.以下关于探究酶促反应影响因素的实验叙述正确的是()A.探究温度对酶促反应速率影响的实验中有空白对照B.以蔗糖和蔗糖酶为材料探究温度对酶促反应速率影响的实验,可选用本尼迪特试剂作为检测试剂C.改变酶的浓度可通过改变酶与底物的接触几率来影响酶促反应速率D.探究pH对酶促反应速率影响实验中将酶与底物混合后迅速至于相应pH缓冲溶液中处理5.曹操于《短歌行》中写道:慨当以慷,忧思难忘。
何以解忧?唯有杜康。
其中“杜康”借代美酒。
酒进入人体后主要在肝脏细胞中代谢,先利用乙醇脱氢酶将乙醇氧化为乙醛,再利用乙醛脱氢酶将乙醛氧化为乙酸,最终将乙醇代谢为CO2和H2O。
下列说法不正确的是A.乙醇代谢产生的[H]可与氧结合生成水,并释放能量B.这两种脱氢酶数量多和活性高的人,一般解酒能力较强C.饮酒后,酒精在人体内的代谢会导致内环境的pH明显降低D.某醉酒者血液中乙醇脱氢酶含量明显超标,其肝脏可能出现病变6.表观遗传中生物表型的改变可能是通过DNA甲基化、RNA干扰等多种机制来实现的。
某基因在启动子上存在富含双核苷酸“C-G”的区域,其中的胞嘧啶在发生甲基化后转变成5-甲基胞嘧啶,仍能在DNA复制过程中与鸟嘌呤互补配对,甲基化会抑制基因的表达。
DNA甲基化的机制和影响

DNA甲基化的机制和影响DNA甲基化是指DNA分子中碳氢化合物甲基基团与胞嘧啶或腺嘌呤的氮原子结合,形成5-甲基胞嘧啶或6-甲基腺嘌呤的化学修饰。
这种化学修饰在多个生物过程中起着重要的作用。
甲基化修饰和基因表达在哺乳动物细胞中,基因表达和甲基化紧密相连。
大部分情况下,DNA甲基化是在启动子区域的CpG岛(即CG序列重复出现的区域)进行的。
这些岛状结构通常位于基因的5'末端,指示该基因是否会被转录为RNA,从而编码蛋白质。
在启动子区域进行的甲基化通常是通过DNA甲基转移酶(DNMT)介导完成的。
甲基化是一种遗传学上易于传递的标记,这也使得它成为细胞遗传记忆的基础。
例如,如果在胚胎发育时存在过多的甲基化,那么这些修饰可能会导致基因在以后的发育过程中被关闭。
此外,甲基化标记还可以区分不同类型的细胞,从而使细胞分化和成熟。
甲基化异常和疾病DNA甲基化的异常可能会引起多种疾病。
例如,在某些癌症类型中,CG岛区域的甲基化被显著改变,导致基因的表达模式发生变化。
此外,如果胚胎发育过程中存在过多的甲基化,可能会影响胚胎着床和细胞分化,进而导致容易发生儿童畸形和神经管缺陷。
过度甲基化范围广泛,包括自闭症、精神障碍、抑郁症等多种疾病。
这种过度甲基化有可能与环境因素有关,比如毒性物质的暴露和激素的使用。
此外,永久地过度甲基化还被认为会导致基因不稳定,从而增加患癌风险。
DNA甲基化和年龄DNA甲基化也和年龄紧密相关。
事实上,通过CG岛区域的DNA甲基化状态可以推断一个人的年龄。
这种反应通常会以线性方式随时间推移而发生,并且受到生活方式因素(如饮食和锻炼)的影响。
此外,研究表明,DNA甲基化状态的模式与特定的生理过程明显相关,例如细胞凋亡和发育。
DNA甲基化的可逆性DNA甲基化可以被移除,从而导致某些基因表达模式的变化。
然而,这种过程是非常复杂的,并且具体涉及到多个分子和生物过程。
现在的某些研究表明,某些高血汞患者使用硫氨酸和B族维生素的补充剂,可能有助于逆转部分甲基化异常。
低温和5—氮胞苷对油菜花芽分化和DNA甲基化的影响

低温和5—氮胞苷对油菜花芽分化和DNA甲基化的影响引言
油菜是一种重要的油料作物,其花芽分化是其生长发育的关键阶段。
在这一过程中,DNA甲基化是一种重要的表观遗传学修饰,在植物生长发育中发挥着重要的调控作用。
目前对低温和5-氮胞苷对油菜花芽分化和DNA甲基化的影响的研究还比较有限。
本文旨在探讨低温和5-氮胞苷对油菜花芽分化和DNA甲基化的影响,为进一步了解油菜生长发育中的调控机制提供理论依据和实践指导。
低温对油菜花芽分化的影响
低温是油菜生长过程中常见的一种胁迫因子,它会对油菜花芽分化产生一定的影响。
研究表明,适度的低温可以促进油菜花芽分化,提高花蕾数量和产量。
长时间的低温胁迫则会抑制花芽分化,影响油菜的产量和品质。
在实际生产中需要合理控制低温胁迫的时间和强度,以促进油菜花芽分化的良好发育。
低温对DNA甲基化的影响
研究发现,低温胁迫可以影响植物的DNA甲基化水平。
在一定程度的低温胁迫下,植物的DNA甲基化水平会发生变化,进而影响基因的表达和调控。
这提示低温可能通过调控DNA甲基化来影响油菜花芽分化的过程。
进一步研究低温对油菜DNA甲基化的影响机制,有助于探索低温胁迫对植物生长发育的影响机制。
5-氮胞苷对油菜花芽分化的影响
5-氮胞苷是一种重要的DNA甲基化抑制剂,它能够干扰DNA的甲基化过程,从而影响基因的表达和调控。
研究发现,在适度的5-氮胞苷处理下,油菜的花芽分化受到一定的促进,花蕾数量和产量均有所提高。
过量的5-氮胞苷处理则会对油菜的花芽分化产生抑制作用,影响其产量和品质。
低温和5—氮胞苷对油菜花芽分化和DNA甲基化的影响

低温和5—氮胞苷对油菜花芽分化和DNA甲基化的影响引言
油菜花是一种重要的农作物,其花期的形成和开花对产量和品质具有重要影响。
研究油菜花芽的分化过程对于提高油菜的产量和品质具有重要意义。
而DNA甲基化是一种重要的表观遗传学调控方式,对于植物的生长和发育具有重要影响。
本研究旨在探究低温和5-氮胞苷对油菜花芽分化和DNA甲基化的影响,从而为提高油菜产量和品质提供理论基础。
材料和方法
1. 植物材料:油菜花种子。
2. 处理方法:分为对照组、低温处理组和5-氮胞苷处理组。
3. 低温处理:将油菜花种子在4℃条件下处理7天。
4. 5-氮胞苷处理:将油菜花种子浸泡在含有5-氮胞苷的培养基中处理7天。
5. 采样:对照组、低温处理组和5-氮胞苷处理组的油菜花种子在处理结束后进行采样。
6. 分析方法:利用组织切片观察油菜花芽的分化情况;利用全基因组测序技术分析DNA甲基化水平。
结果
1. 组织切片观察结果显示,低温处理组和5-氮胞苷处理组的油菜花芽分化程度显著高于对照组。
2. 全基因组测序结果显示,低温处理组和5-氮胞苷处理组的油菜花芽DNA甲基化水平显著高于对照组。
讨论
本研究结果表明,低温和5-氮胞苷处理对油菜花芽的分化和DNA甲基化水平均有显著影响。
低温处理可以促进油菜花芽的分化,同时提高DNA甲基化水平;而5-氮胞苷处理同样可以促进油菜花芽的分化,同时提高DNA甲基化水平。
这表明低温和5-氮胞苷可能通过调控DNA甲基化水平来影响油菜花芽的分化过程。
这些结果为进一步探究油菜花芽分化和DNA甲基化的调控机制提供了重要的参考。
5-氮杂胞苷和丁酸钠对大麦苗期耐湿性的影响

5-氮杂胞苷和丁酸钠对大麦苗期耐湿性的影响徐延浩;侯泽豪;张文英;刘晓雪;杨飞【期刊名称】《广东农业科学》【年(卷),期】2015(042)024【摘要】以耐湿性具有差异的大麦泰兴9425和Naso Nijo为材料,采用盆栽方式研究外源5-氮杂胞苷和丁酸钠对湿害下大麦不定根、干重、叶片脯氨酸和丙二醛(MDA)含量的影响.湿害指数分析表明泰兴9425湿害受害指数较低,耐湿能力更强.外源5-氮杂胞苷处理使湿害胁迫后泰兴9425和Naso Nijo湿害指数分别降低4.90%和15.22%,MDA含量分别降低12.83%和34.68%;显著降低了Naso Nijo 脯氨酸的累积,而泰兴9425脯氨酸积累增加,但并不显著.外源丁酸钠对两个供试品种湿害指数没有显著影响,与淹水组相比,显著增加了两个品种叶片MDA和脯氨酸的累积.研究结果表明5-氮杂胞苷在一定程度上能减轻大麦苗期湿害,并对其具有一定生理调节作用,而丁酸钠则不适用于大麦湿害缓解体系.【总页数】5页(P26-30)【作者】徐延浩;侯泽豪;张文英;刘晓雪;杨飞【作者单位】主要粮食作物产业化湖北省协同创新中心,湖北荆州434025;长江大学农学院,湖北荆州434025;主要粮食作物产业化湖北省协同创新中心,湖北荆州434025;长江大学农学院,湖北荆州434025;主要粮食作物产业化湖北省协同创新中心,湖北荆州434025;长江大学农学院,湖北荆州434025;主要粮食作物产业化湖北省协同创新中心,湖北荆州434025;长江大学农学院,湖北荆州434025;主要粮食作物产业化湖北省协同创新中心,湖北荆州434025;长江大学医学院,湖北荆州434025【正文语种】中文【中图分类】S512.3【相关文献】1.丁酸钠和5-氮杂胞苷对大麦、水稻、玉米、小麦种子萌发及芽苗期生长的影响[J], 侯泽豪;杨飞;商水根;方汉顺;张文英;徐延浩2.大麦苗期耐湿性的鉴定筛选 [J], 杨建明;沈秋泉;汪军妹;朱靖环3.大麦品种苗期湿害试验及品种耐湿性研究 [J], 陶红;沈会权;陈和;陈健;陈晓静;徐相宏;张昌松4.5-氮杂胞苷逆转SARI基因甲基化对人乳腺癌细胞生长和侵袭的影响 [J], 朱文斌; 孙艳; 张春晶; 蒋瑞雪; 陈绍仁; 胡芳; 刘继鑫; 孙艳宏; 姚淑娟; 吕丽艳; 张畅5.5-氮杂胞苷对高表达转玉米C4-PEPC基因水稻耐旱性的影响 [J], 严婷;李霞;曹悦;吴博晗;王净;张嫚嫚因版权原因,仅展示原文概要,查看原文内容请购买。
5azaC对菠菜生长发育及DNA甲基化的影响

5-azaC 对菠菜生长发育及 DNA甲基化的影响菠菜 (Spinacia oleracea L.)为藜科(Chenopodiaceae)菠菜属(Spinacia)一年生或两年生草本植物。
菠菜作为一种常有的园艺蔬菜作物已经被广泛种植,且菠菜一般为雌雄异株 , 很少雌雄同株 , 其染色体组为 2n=2x=24(XY)。
所以 , 对于菠菜的研究目前主要会合在新品种选育、种植技术优化以及性别决定及分化体系三个方面, 对其余方面研究特别是菠菜表观遗传特色特别是性别发育的表观调控体系的研究鲜有报导。
DNA甲基化作为最早发现的基因组表观遗传修饰门路之一 , 已有凭据表示该体系可以植物基因的表达进行表观调控, 从而影响植物的生长发育。
为了探究 DNA甲基化在菠菜营养生长阶段及性别发育阶段的作用, 本文以甲基化克制剂 5-azaC 办理及未办理的菠菜为资料, 经过比较观察 , 解析菠菜的生长发育过程及性比的变化 , 同时 , 采纳 MS-ISSR、MS-RAPD和 HPLC法对基因组 DNA 甲基化水平进行检测 , 解析菠菜表型变化和基因组DNA甲基化水平之间的相关关系 , 从而试试揭露 DNA甲基化对于菠菜生长发育及性别过程中的作用体系。
主要研究结果以下 :1. 使用不一样浓度的 5-azaC 对菠菜种子进行办理 , 解析甲基化克制剂5-azaC 对于菠菜萌芽势、萌芽率、株高、叶长、根长、开花时间及性比的影响。
结果表示 ,5-15 μ M的 5-azaC 对菠菜办理后 , 使萌芽率、萌芽势、根长、苗长、叶长、株高与比较对比 , 跟着浓度的增高而降低 , 开花时间提早 , 但是与比较对比相差不明显 ; 菠菜对 30μM浓度办理不敏感 , 各项生长指标与比较相差很少 ;50-1000 μM的 5-azaC 对菠菜办理后 , 跟着浓度的增添与比较对比各项生长指标都降低 , 此中高浓度有显然的克制作用 , 如株高与比较对比显然变矮, 开花时间内提早 1d 以上。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
5-azaC处理水稻幼苗对基因组DNA 甲基化及水稻生长发育的影响表观遗传学:不需要基因序列改变而产生的可遗传的基因表达改变;主要包括DNA的甲基化修饰和组蛋白氨基酸的末端修饰。
表观遗传变异包括: X-染色体失活、转基因沉默、核仁显性和基因组印记等方面。
表观遗传变异的原因:是由于表观遗传密码的改变,如DNA 甲基化、组蛋白的公家修饰改变等。
DNA 甲基化:在DNA 复制后,经DNA甲基转移酶(DMT)催化,将S-腺苷酰-L-甲硫氨酸(SAM) 上的甲基基团连接到DNA 分子腺嘌呤碱基或胞嘧啶碱基上,进行DNA修饰的过程。
DNA 甲基化的方式胞嘧啶甲基化腺嘌呤甲基化DAM (DNA 腺嘌呤甲基转移酶)识别回文序列GATC, 在此位置两条链的腺嘌呤N-6位置上同时甲基化,同时SAM 转变为SAH(S-腺苷高半胱氨酸)在DMT(DNA甲基转移酶)的作用下,CpG(CNG,CCGG)位点胞嘧啶C5被甲基化DNA 甲基化作用DNA甲基化的生物学意义影响基因表达状态:通过该表基因调控区DNA 甲基化程度调控基因转录参与基因组防御:通过高度甲基化而使外源DNA(如转座子)处于沉默状态提高环境适应能力:在不改变基因型的情况下产生可遗传的新表型DNA 胞嘧啶甲基化的种类和形成从化头甲基化(de novo methylation)DNMT3a ,DNMT3b在完全没有甲基化的DNA 上加上甲基。
维持化甲基化(maintenance methylation)Met1 ,DNMT1较特异地识别半甲基化状态的DNA ,负责在细胞分裂过程中依据DNA 母链的甲基化状态给新合成的DNA 链上加上甲基。
DNA 胞嘧啶甲基化检测方法使用对胞嘧啶5’- 甲基化状态敏感的限制性内切酶进行酶切处理,然后检测多态性:甲基化敏感扩增多态性(MSAP, Methylation SensitiveAmplification Polymorphism)胞嘧啶转化为其它碱基,然后对比测序:重亚硫酸盐测序(Bisulfite sequencing)GCmTACT重亚硫酸钠GCmTATT测序此外,还有单核苷酸法,甲基化接受力的检测法,CpG 岛分离法和基因活性测量法,基因芯片法等表1 甲基化敏感酶对不同甲基化状态DNA 的切割结果CCm GG不切割CCmGG不切割CC GG Msp I不切割CC GG Hpa IICmCGG CCGG 原始序列内切酶? MSAP 流程:提取基因组DNA(注意发育时期和成长状态相同)………ATCCGGTGGCTTGCCmGGAC……………TAGGCCACCGAACGG CCTG……Msp I 酶切+ 接头Hap II 酶切+ 接头ATCC GGTGGCTTGCCm GGAC ATCC GGTGGCTTGCCmGGACTAGG CCACCGAACGG CCTG TAGG CCACCGAACGG CCTGPCR PCRPAGE 电泳PAGE 电泳Msp I5’…CC(m)G G…3’3’…GG C(m)C…5’Hpa II5’…CCGG…3’3’…GGCC…3’? 重亚硫酸盐测序实验目的使用不同浓度的5-azaC 溶液,处理生长早期的水稻幼苗,使其基因组DNA 胞嘧啶甲基化水平发生显著的可遗传改变,使用MSAP 方法检测DNA 胞嘧啶甲基化水平的改变;水稻幼苗因DNA 甲基化水平改变而影响基因表达水平,产生明显的表观遗传表型。
实验材料1、水稻:日本晴(Oryza sativa var japonica, nipponbare)2、所需仪器离心机、PCR仪、气浴摇床、水浴摇床、植物生长箱电泳仪、电泳槽、微量移液器、紫外分析仪、分析天平3、试剂(1)植物基因组DNA 改良CTAB 提取液:buffer A: 0.35mol/L Sorbitol ,0.1mol/L Tris-HCl pH7.5 ,0.5mmol/L EDTA;buffer B: 0.2mol/L Tris-HCl pH7.5 ,0.05mol/L EDTA ,2mol/L NaCl ,2%CTAB;buffer C: 5%N-lauroylsarcosine(2)PCR试剂:Takara rTaq ,10X PCR buffer ,ddH2O ,①预扩增引物:H/M+0: (5´-GACTGCGTACCAATTCA-3´)PCR 引物EcoRI+0: (5´-ATCATGAGTCCTGCTCGG-3´)②选择性扩增引物:H/M+AN: 5´-GACTGCGTACCAATTCAA(T, C, G)-3´EcoRI+AN: 5´-ATCATGAGTCCTGCTCGGA(T, C, G)-3´。
(3)酶切-连接试剂:EcoRI (NEB) ,Hap II (NEB) ,Msp I (NEB) ,T4 DNA ligase ,T4 10 xreaction buffer(4)接头:EcoRI adapter: 5’-CTCGTAGACTGCGTACC-3’CATCTGACGCATGGTTAAHapII/MspI adapter: 5-‘GATCATGAGTCCTGCT-3’AGTACTCAGGACGAGC(5)PAGE 试剂:聚丙烯酰胺,尿素,AgNO 3,Na 2 CO 3,溴酚蓝(6)银染固定/终止液(10% 冰乙酸) ,染色液(2升双蒸水预冷,用时加2克硝酸银,3ml 37% 甲醛) ,显影液(2升双蒸水中溶解60克碳酸钠,冷却至10-12ºC,用时,加入3ml 37% 甲醛,400ul 硫代硫酸钠(10mg/ml)) 。
(7)其它试剂:5-azaC(100mg/mL) ,蒸馏水,PVP (20%) ,氯仿: 异戊醇(24:1),异丙醇,70% 乙醇,RNA 酶A (DNase-free) ,TE ,TAE ,琼脂糖,EB ,未酶切的λDNA ,1xTBE (0.089M Tris-borate, 0.089 M Boric Acid, 0.002 M EDTA) ,3x loading buffer(300 mM NaOH, 97% formamide, 0.2% bromophenol blue)。
实验方法1、水稻的培养(26 ℃,暗培养)取水稻水稻种子20粒X 7组,将水稻种子用75% 的酒精消毒后用无菌水浸泡数小时,当水稻种子露嘴时,分别置于90mm 培养皿内的双层滤纸上,加15mL蒸馏水浸种24h(第一天)。
2、5-azaC处理水稻幼苗:(1)6组种子分为6个实验组和1个对照组,实验组分别用20mg/mL,40mg/mL, 60mg/mL、80mg/mL、100mg/mL、120mg/mL5-azaC溶液处理,对照组用蒸馏水培养,共处理10天,每2天换处理液1次(第二~ 十一天)。
(2)10 天后每个处理随机取10 株幼苗嫩叶混合提取DNA,其余麦苗移载大田并观察田间生长情况。
2、植物基因组DNA 的提取与纯化改良CTAB法(Kidwell and Osborn, 1992)[8](1)取不同处理的叶1g,将其在液氮中研磨成粉后转入50ml离心管。
(2)向离心管中加入等体积DNA 提取缓冲液(A:B:C=1:1:0.4混合,加1%PVP于B液)。
(3)置于65℃恒温水浴下摇床,40-50rpm振摇90分钟。
(4)加入等体积的氯仿:异戊醇(24:1) ,轻轻上下颠倒10-15分钟后,4 ℃3000rpm离心10-15分钟。
(5)取上层水相加入2/3 体积的预冷异丙醇,上下颠倒5分钟并置4℃冰箱内10min。
(6)待DNA 沉淀后于4 ℃3000rpm离心收集DNA于管底。
(7)DNA于70%乙醇中过夜。
(8)取DNA沉淀并风干(第十二天)。
(9)在2ml离心管中加1ml TE溶解沉淀。
(10)待DNA 完全溶解后,加入5ul不含DNA酶的RNA酶,37℃保温30分钟以除去DNA 中的RNA。
(11)加入等量氯仿: 异戊醇(24:1) 纯化DNA ,轻轻上下颠倒10-15分钟后,4℃3000rpm 离心10-15分钟。
(12)取DNA沉淀,用70%乙醇过夜洗涤DNA,并将其溶于DNA 于200-500ul TE中。
(13)取基因组DNA溶液,0.8% 琼脂糖凝胶电泳,检查DNA的质量并据已知的DNA Marker (未酶切的λDNA)估计其浓度;调整DNA 浓度,置于-20℃备用。
3、水稻基因组DNA 酶切(1)甲基化分析采用两种甲基化敏感酶HapⅡ和MspⅠ(NEB)。
在限制性酶切和连接前,先把两个单链的接头复性为双链结构( 序列见试剂部分) ,即100℃变性5分钟,缓慢冷却至室温,-20℃冷冻保存备用。
(2)水稻MSAP限制性酶切和连接体系如下:表2 水稻MSAP限制性酶切、连接体系T4 10 x buffer 2.5ulBSA (10mg/L) 0.1ulEcoR I adaptor (5pM) 0.5ulH/M adaptor (50pM) 0.5ulEcoR I (20U/ul) 0.5ulH/MMspI (10U/ul) 0.5ul HpaII (20U/ul) 0.25ulT4 ligase 0.1ulGenomic DNA 300ng20ul AFLP-Water 补足总反应体积(3)混匀体系,37℃,6小时,8℃,4小时,4℃保存。
(4)共四个处理组,即水稻25mg/mL 处理组,水稻50mg/mL处理组,水稻100mg/mL 处理组,水稻对照组;每组分别用HapII/MspI双酶切,共需切8管(第十五天)。
4、预扩增(1)预扩增:水稻AFLP 预扩增体系如右表(预扩增引物PCR引物部分):混匀体系,按下列参数进行PCR 扩增反应:表3 AFLP 预扩PCR体系10 x PCR reaction buffer 2.5uldNTPs (2.5mM) 2ulEcoR I primer (5 uM) 1ulH/M primer (5 uM) 1ulTaq DNA polymerase (5U/ uL) 0.2ulPCR water 16.3ulDNA 模板(预扩增稀释产物)2ulTotal volume 25ul(2)PCR 程序:94℃预变性2min, 94℃变性30秒,56℃复性30秒,72℃延伸60 秒, 共进行30 个循环,最后72℃延伸10分钟(第十七天)。
(3)根据检测结果,用0.1x TE buffer稀释预扩增产物1/10~1/40,作为PCR 选择性扩增的DNA模板。
按以下参数进行PCR 扩增反应:表4水稻AFLP 选择性扩增体系10 x PCR reaction buffer 1.5uldNTPs (2.5mM) 0.6ulEcoR I primer (5 uM) 0.6ulH/M primer (5 uM) 0.6ulTaq DNA polymerase (5U/ uL) 0.12ulPCR water 9.58ulDNA 模板(预扩增稀释产物) 2 2ulTotal volume 15ul94℃预变性2 分钟, 94 ℃变性30 秒,65℃复性30 秒,72℃延伸80 秒,以后每轮循环温度递减1℃,扩增10 轮。