钙调素拮抗剂与Cat+对茄子幼苗抗冷性的影响
有效预防茄子冷害

有效预防茄子冷害
植株遭受低温冷害,生理代谢失调,生长发育受阻。
因受害程度和受害时间不同,症状也有差异,一般表现为叶片扭曲、叶面出现淡褐色或白色斑点,叶缘干枯等,严重时整株枯死。
一、病因温室保温性能差,冬季温度低。
遇到了寒流、雨雪和短期低温等天气状况。
定植期过早。
二、防治方法
① 正确确定播种期、定植期选择寒尾暖头的无风晴天播种或定植。
② 培育壮苗采用营养钵育苗,黑色塑料营养钵具有白天吸热,夜温保温护根的作用。
在外界温度-10℃,苗床温度6~7℃时,营养钵内温度在10℃左右,幼苗能缓慢生长,不受冻害。
进行低温锻炼,增强体内抗寒能力,往往会收到良好的效果。
③叶面喷肥在低温逆境下,根系吸收能力差,叶面上喷光合微肥,可补充因根系吸收营养不足而造成的缺素症。
叶面喷米醋,可抑菌驱虫。
米醋与白糖和过磷酸钙混用,可增加叶肉含糖量,提高叶片硬度,提高抗寒性。
喷醋的浓度为100~300倍。
注意,低温季节不要使用生长素类生长调节剂,以防降低抗寒性。
④喷抗寒剂目前市场出售植物抗寒剂、低温保护剂、防冻剂等叶面喷肥,都属于生长调节剂一类物质,其有增加幼苗抗性,保苗促长的作用,可以应用。
另外,通过对外源ABA、氧化钙、多效唑等化学物质作试验,表明这三种物质也具有抗低温功能,如果用50~100毫克/升的ABA、0. 5%氧化钙或者50~100毫克/升多效唑对甜椒植株喷雾,可有效预防低温伤害。
此外,于傍晚喷巴姆兰丰收液膜250倍液,或27%高。
茄子嫁接苗根系对低温环境胁迫的响应

茄子嫁接苗根系对低温环境胁迫的响应
高青海;吴燕;徐坤;高辉远
【期刊名称】《应用生态学报》
【年(卷),期】2006(17)3
【摘要】以‘济农2000’为对照,研究了低温胁迫下不同抗冷性茄子砧木嫁接苗根系的生理生化变化及其与抗冷性的关系.结果表明,在10℃(昼)/3℃(夜)低温处理初期,根系对低温最敏感,表现为MDA含量、冷害指数、POD活性、脯氨酸含量、可溶性蛋白含量等迅速增加,而根系呼吸速率、SOD、CAT活性等迅速降低.随着低温胁迫时间的延长,根系呼吸速率持续降低,而冷害指数持续升高,渗透调节物质基本呈增加的趋势.常温恢复3 d后,呼吸速率、渗透调节物质、SOD活性等升高,但均以抗冷性较强的赤茄砧木嫁接苗恢复能力较强,而抗冷性较弱的台茄砧木嫁接苗与自根苗较差.利用抗冷性较强的砧木进行嫁接,可显著增强茄子根系的活性,进而提高其抗冷性.
【总页数】5页(P390-394)
【关键词】茄子;嫁接;低温胁迫;根系特性
【作者】高青海;吴燕;徐坤;高辉远
【作者单位】山东农业大学;菏泽学院
【正文语种】中文
【中图分类】S641.1
【相关文献】
1.甘蔗幼苗根系形态结构及保护系统对低温胁迫的响应 [J], 孙波;刘光玲;杨丽涛;李杨瑞
2.低温胁迫下砧穗互作对茄子嫁接苗叶片抗氧化酶系统的影响 [J], 张晓艳;徐坤
3.NaCl胁迫对茄子嫁接苗根系多胺代谢的影响 [J], 刘正鲁;朱月林;魏国平;张古文;杨立飞;胡春梅
4.高、低温胁迫对雾培嫁接苗根系生长发育的影响 [J], 鲁雪利;于海业;赵红星;张蕾
5.茄子对低温胁迫的生理响应及不同品种耐冷性比较 [J], 任国三;程加祥;王红对;赵传麟;张峰;李修燕;徐传铭
因版权原因,仅展示原文概要,查看原文内容请购买。
茄子耐低温材料的筛选及其耐低温生理响应研究

植物遗传资源学报2013ꎬ14(6):1161 ̄1166JournalofPlantGeneticResources茄子耐低温材料的筛选及其耐低温生理响应研究包崇来ꎬ杜黎明ꎬ胡天华ꎬ朱琴妹ꎬ胡海娇ꎬ何群燕ꎬ毛伟海(浙江省农业科学院蔬菜研究所ꎬ杭州310021)㊀㊀摘要:以100份茄子种质资源中筛选出的耐低温基因型材料E659和低温敏感基因型材料E646为试材ꎬ研究了低温处理对不同耐寒性茄子叶片的光抑制能力㊁丙二醛含量㊁电导率㊁过氧化氢酶(CAT)㊁抗坏血酸酶(APX)和超氧化物歧化酶(SOD)活性的影响ꎮ结果表明ꎬ在4ħ处理5d后ꎬ与低温敏感材料E646相比ꎬ低温胁迫下筛选获得的极耐低温基因型材料E659ꎬFV/FM比值下降较少ꎬ表明其PSII系统受损较少ꎬ而丙二醛含量和电导率上升幅度较低则表明膜脂过氧化反应引起的氧化损伤较轻ꎮ4ħ低温处理24h之内ꎬ发现E659植株中抗坏血酸酶活性增高ꎬ过氧化氢酶和超氧化物歧化酶的活性损害减弱或时间推迟ꎮ可见ꎬ茄子耐低温基因型材料E659在低温胁迫下能快速启动应激反应ꎬ增强氧化系统清除能力ꎬ减轻低温对植株生长发育的伤害ꎮ㊀㊀关键词:茄子ꎻ低温胁迫ꎻ光氧化反应ꎻ生理机制ScreenofLow ̄temperatureTolerantCultivarandAnalysisofPhysiologicalMechanisminEggplantBAOChong ̄laiꎬDULi ̄mingꎬHUTian ̄huaꎬZHUQin ̄meiꎬHUHai ̄jiaoꎬHEQun ̄yanꎬMAOWei ̄hai(TheInstituteofVegetableꎬZhejiangAcademyofAgriculturalSciencesꎬHangzhou310021)Abstract:Inthestudyꎬweobtainedalow ̄temperaturetolerantcultivarE659andacoldsensitivecultivarE646byscreeningfrom100eggplantgermplasmresourcesunderlowtemperaturestress.Andthetwocultivarsweremeasuredthephotochemicalcapacityꎬoxidativedamageꎬhydrogenperoxideaccumulationꎬandantioxidantenzymeactivities.TheresultsshowedthatcomparedwithE646ꎬE659hadlessdecreaseofFV/FMratioandlessincreaseofmalondialdehyde(MDA)contentandelectrolyteleakageafter4ʎtreatmentfor5daysꎬwhichsuggestteddemonstra ̄tingminordamageofPSIIsystemandoxidativedamagebylipidperoxidationinthiscultivarꎬrespectively.Thea ̄scorbateperoxidase(APX)ofE659increasedandatalase(CAT)activitysuperoxidedismutase(SOD)reducedordelayedwithin24hoursof4ħtreatment.Theseconclusionsindicatedthatthelow ̄temperaturetolerantcultivarE659couldpossessesarapidresponsemechanismandhadhigherreactiveoxygenspeciessystem(ROS)clearancecapacitytolowercoldinjury.Keywords:eggplantꎻlowtemperaturestressꎻphotoxidativeꎻphysiologicalmechanism收稿日期:2013 ̄05 ̄07㊀㊀修回日期:2013 ̄05 ̄29㊀㊀网络出版日期:2013 ̄10 ̄22URL:http://www.cnki.net/kcms/detail/11.4996.S.20131022.1548.029.html基金项目: 十二五 农村领域国家科技计划项目(2012AA100103)ꎻ浙江省重大科技项目(2012C12903)ꎻ浙江省农科院创新提升工程项目(2012R23Y01E01)第一作者研究方向为茄子育种与种质创新ꎮE ̄mail:baocl@126.com通信作者:毛伟海ꎬ研究方向为蔬菜育种与种质创新ꎮE ̄mail:maowh@126.com茄子是起源于热带的喜温蔬菜作物ꎬ多数茄子品种在温度低于7~8ħ时ꎬ种子发芽和植株营养生长及开花结果都会受到阻碍ꎬ低温制约了茄子的生长发育ꎬ致使其产量和品质降低[1]ꎮ阎世江等[2]研究了低温处理下茄子成株期的一些农艺性状ꎬ结果表明低温处理对茄子的结果数影响较大ꎬ进而严重影响产量ꎮ因此ꎬ选育耐低温茄子材料㊁探讨其耐低温的生理生化机制ꎬ对研究茄子耐低温分子调控机理㊁选育耐低温品种具有非常重要的意义ꎮ研究显示ꎬ持续9d的夜间低温会造成番茄不可逆的PSII光抑制作用[3]ꎮ水稻营养生长阶段受到低温胁迫时ꎬ通过PSII系统的电离子运植㊀物㊀遗㊀传㊀资㊀源㊀学㊀报14卷输会受到抑制ꎬ从而引起光抑制[4]ꎮ因此维持较高的光化学反应能力ꎬ从而保持PSII系统的运输能力是耐低温所必须的ꎮPSII系统的损坏可能是由于氧化应激造成的ꎬ通过形成活性氧造成氧化损伤[5]ꎮ植物体内有多种机制可以减少或防止氧化应激及其引起的不良影响ꎮ一个是通过无害辐射扩散过剩能量来防止活性氧形成ꎮ由玉米黄素和花药黄素的合成过程形成的叶黄素循环ꎬ是过剩能量无害辐射扩散的主要生理机制[6]ꎮ另一个反应就是通过类胡萝卜素或抗氧化酶对活性氧(ROSꎬreactiveoxygenspecies)进行解毒ꎮ抗氧化酶包括超氧化物歧化酶(SODꎬsuperoxidedis ̄mutase)㊁抗坏血酸过氧化物酶(APXꎬascorbateper ̄oxidase)和过氧化氢酶(CATꎬcatalase)[7 ̄8]ꎮ目前ꎬ已有多个报道通过研究抗氧化酶活性变化来探讨茄子耐低温生理机制ꎮ王凤华等[9]研究表明低温胁迫下APX活性先上升后下降ꎬ胁迫处理72h时酶活性最高ꎮ张泽煌等[10]报道在0~5ħ低温下ꎬ茄子叶片细胞膜透性增大ꎬMDA含量上升ꎬ细胞膜脂过氧化加剧ꎬ细胞膜损伤ꎬSOD和CAT活性上升ꎬPOD活性保持相对稳定ꎮ张晓艳等[11]研究表明ꎬ低温胁迫下不同组合茄子嫁接苗中ꎬ耐低温材料的SOD㊁POD和CAT活性均高于不耐低温材料ꎮ这些研究结果均表明茄子在低温胁迫时ꎬ体内的抗氧化酶活性会发生一系列的变化ꎬ但是在不同的品种材料之间ꎬ变化趋势是不同的ꎮ虽然已有多个研究探讨了茄子的耐低温生理机制ꎬ但还未见获得极耐低温茄子基因型材料的报道ꎮ本研究获得了1份耐低温性非常好的优异种质资源ꎬ对选育耐低温茄子新品系具有非常重要的意义ꎮ本研究主要通过测定茄子耐低温与低温敏感品种之间的光化学反应能力和光抑制及氧化损伤反应的区别ꎬ旨在探讨该耐低温基因型材料的耐低温机理ꎬ为耐低温基因的QTL定位㊁克隆奠定基础ꎮ1㊀材料与方法1 1㊀试验材料的培养与处理供试材料为本课题组自主选育的100份茄子高代纯合自交系ꎮ在人工气候室中采用50孔穴盘育苗ꎮ育苗期间平均昼夜温度为25ħ/18ħꎬ白天最大光强为800μmol/μm2 sꎮ幼苗长至4叶1心时进行低温处理ꎮ处理时采取逐步降温的方法ꎮ先在昼/夜温度为15ħ/10ħ条件下处理1dꎬ再置于昼/夜温度为10ħ/5ħ下处理1dꎬ最后在昼夜温度为4ħ下处理10dꎮ分别挑选极耐低温材料E659和低温敏感材料E646ꎬ再一次播种生长后于4叶1心时期进行低温处理ꎬ处理方式与筛选时期相同ꎮ在昼夜温度为4ħ处理后ꎬ分别于0h㊁6h㊁12h㊁24h㊁48h㊁5d取样ꎬ用于叶绿素荧光参数和生理指标的测定ꎮ每次每品种各取2株ꎬ摘取叶片后迅速用液氮保存ꎬ贮存于-72ħ超低温冰箱ꎮ1 2㊀叶绿素荧光参数的测定用荧光计(英国Lynn公司FMSIHansatech)分别在对照和处理植株充分扩展的叶片上进行叶绿素荧光测量ꎮ叶片在黑暗下预处理30minꎬ8000μmol/m2 sphotons曝光0 8sꎬ收集数据ꎮ光系统Ⅱ(PSⅡ)最大光化学量子产量(Fv/Fm)ꎬ内在量子产量FVᶄ/FMᶄꎬ光化学淬灭qP(photochemi ̄calquenching)由荧光计自动计算得出ꎮFVᶄ/FMᶄ与qP的比例为PSII效率Pꎮ1 3㊀电导率的测定分别取对照和处理样品2片完全舒展的叶片浸泡在20mLddH2O中ꎬ25ħ下震荡处理2hꎬ然后用mettlertoledo公司SecenEasyConductivity电导率仪测量水电导率ꎮ样品在沸水中煮30minꎬ冷却到室温ꎬ再次测量水电导率ꎮ煮沸后电导率/煮沸前电导率的比率为电导率ꎮ1 4㊀抗氧化酶活性、膜脂过氧化反应和过氧化氢含量测定㊀㊀准确称取0 1g叶片组织ꎬ液氮研磨后加入100mmol/LK2HPO4 ̄KH2PO4缓冲液900μLꎬ制成10%的组织匀浆ꎬ5000g离心10minꎬ取上清ꎬ待测ꎮ超氧化物歧化酶SOD㊁过氧化氢酶CAT㊁丙二醛含量和过氧化氢H2O2的测定方法按照南京建成生物工程研究所研制的试剂盒说明书进行ꎬ并按照各自说明书上提供的公式进行计算ꎮ抗坏血酸过氧化物酶APX测定采用上海博耀生物公司生产的APX定量检测试剂盒(ELISA)ꎬ按照试剂盒测定方法准备试剂㊁样品和标准品ꎮ加入准备好的样品㊁标准品和酶标试剂后ꎬ37ħ反应60minꎬ洗板5次ꎬ加入显色液ꎬ37ħ显色15minꎬ加入终止液ꎬ15min内在450nm处测定OD值ꎮ1 5㊀数据分析3次独立试验测定ꎬ每次测定试验重复3个样品ꎬ共9个数据进行分析ꎮ利用Excel2003和DPSv9 50统计软件对获得的数据进行统计分析ꎮ2611㊀6期包崇来等:茄子耐低温材料的筛选及其耐低温生理响应研究2㊀结果与分析2 1㊀耐低温材料的筛选100份高代纯合自交系在4叶1心时经4ħ低温处理10d后ꎬ基因型E659植株形态正常ꎬ生长良好ꎬ没有出现冷害症状(图1)ꎬ而其他基因型几乎都出现叶片萎缩ꎬ植株倒伏ꎬ呈现典型的冷害特征ꎮ基因型E646在4ħ低温处理后第4天叶片逐渐萎缩ꎬ第10天时所有植株叶片全部倒伏ꎬ症状最严重(图1)ꎮ重复3次试验结果基本一致ꎬ说明基因型E659是1份耐低温材料ꎮ图1㊀茄子幼苗低温处理10d后植株生长状况Fig 1㊀TheseedlingsgrowthofgenotypesE659andE646afterlowtemperaturestressfor10daysE659生长势强ꎬ株高100~110cmꎬ第1雌花节位9~10节ꎬ花蕾紫色ꎬ中等大小ꎮ平均单株座果数28 8个ꎬ果长31 2cmꎬ果粗3 0cmꎬ单果重110g左右ꎬ果形直ꎬ果皮紫红色ꎬ光泽好ꎬ外观漂亮ꎬ商品性好ꎬ耐低温性强ꎬ是一个很好的耐低温紫红长茄的亲本材料ꎮ2 2㊀低温胁迫对茄子光化学反应能力的影响FV/FM代表PSII系统的最高效率ꎬ是光抑制反应的最佳体现指标ꎮ在低温处理24h之前ꎬ2种基因型的FV/FM比值没有明显变化ꎮ直到低温处理5d后(图2A)ꎬ不耐冷基因型E646中ꎬFV/FM比值明显比未经低温处理的对照降低60%左右ꎬ而耐低温基因型E659中ꎬFV/FM比值比未处理的样品下降幅度较小ꎮ表明耐低温品系中光化学反应受损较少ꎬ光抑制能力较强ꎬPSII系统中最大效率P值与PSII光化学反应中的光吸收能力相对应ꎮ图2B表明ꎬ低温处理5d后ꎬ与常温对照的材料相比ꎬE646中P值下降了将近60%ꎬ而E659中P值只稍微下降ꎬ而且低温处理后ꎬE659中的P值明显高于不耐低温材料E646ꎬ说明E659的光合系统在低温处理下受损较小ꎬ能维持较强的光化学反应能力ꎮA:FV/FM值ꎻB:PSII效率ꎬ下同A:FV/FMratioꎬB:PSIIefficiencyꎬthesameasbelow图2㊀茄子幼苗低温处理5d后基因型E646和E659的FV/FM值和PSII效率Fig 2㊀FV/FMratioandPSIIefficiencyineggplantseedlingsofgenotypesE646andE659afterlowtemperaturetreatmentfor5days2 3㊀低温胁迫对质膜稳定性的影响机体通过酶系统产生氧自由基ꎬ攻击生物膜中的不饱和脂肪酸ꎬ从而引发膜脂过氧化反应ꎬ并因此形成膜质过氧化产物ꎬ如醛基丙二醛等ꎮ因此ꎬ可以通过测定丙二醛反应底物巴比妥酸TBA的含量变化来显示丙二醛的含量ꎬ从而反应膜脂过氧化反应ꎬ揭示细胞损伤情况ꎮ图3A表明ꎬ低温处理5d后ꎬ不耐低温基因型E646中ꎬ丙二醛含量比未经低温处理的对照升高将近7倍ꎻ而在耐低温材料E659中ꎬ丙二醛含量与未经低温处理的对照相比ꎬ只升高了3 5倍左右ꎬ升高幅度为E646的一半ꎬ说明耐低温基因型中膜脂过氧化反应较弱ꎬ细胞损伤情况相对较轻ꎮ图3B表明ꎬ当低温处理5d后ꎬ不耐低温基因型E646的电导率升高了7倍ꎬ而基因型E659约只升高了4倍ꎬ表明在耐低温基因型中ꎬ细胞膜的受损程度较低ꎬ离子渗漏程度较轻ꎬ而E646中ꎬ细胞膜破坏及离子渗漏严重ꎮ2 4㊀低温胁迫对过氧化氢酶和抗氧化酶活性的影响已有的研究结果表明ꎬ低温处理初期氧化反应指标非常重要[5]ꎮ如图4显示ꎬ胁迫6h后ꎬH2O2产量在耐冷基因型E659中快速增加ꎬ但是不耐冷材料E646中H2O2产量只见微弱升高ꎬ变化不大ꎮ基因型E659中的H2O2产量虽然从12h就开始下降ꎬ3611植㊀物㊀遗㊀传㊀资㊀源㊀学㊀报14卷图3㊀茄子幼苗低温处理5d后对基因型E646和E659的丙二醛含量(A)和电导率(B)的影响Fig.3㊀OxidativedamageineggplantseedlingleavesofgenotypesE646andE659subjectedtolowtemperaturestressfor5days.(A)Malondialdehydeproductionmeasured. (B)Membraneintegritymeasuredasrelativeionleakage但是到24h时ꎬ一直都比基因型E646中的H2O2产量高ꎬ到了48h时ꎬ2种材料中的H2O2产量才达到几乎一致ꎮ结果表明E659可能通过增加H2O2产量诱发某种调控机制来抵抗低温胁迫ꎮ图4㊀茄子幼苗低温处理下基因型E646和E659中过氧化氢含量的变化Fig.4㊀H2O2productioninseedlingsofthegenotypesE646andE659subjectedtolowtemperaturestress2种基因型在低温处理24~48h时ꎬSOD㊁CAT和APX的活性变化不大(图5)ꎮ但在胁迫处理24h之前ꎬ各种抗氧化酶的活性及变化程度都是不一样的ꎮ未经低温处理时ꎬ2种基因型中的同一种抗氧化酶活性差不多ꎮ超氧化物歧化酶SOD对机体的氧化与抗氧化平衡起着至关重要的作用ꎬ能清除超氧阴离子自由基ꎬ保护细胞免受损伤ꎮ如图5A所示ꎬ在2种基因型中ꎬ处理6~48h期间ꎬSOD酶活性较未处理时均下降ꎬ但是不耐低温基因型E646中酶活性下降量显著多于耐低温基因型E659ꎬ表明基因型E646中SOD酶活性受低温影响较大ꎬ失活较严重ꎮ过氧化氢酶CAT能分解H2O2ꎬ消除毒性ꎮ在开始低温处理6h后ꎬ耐低温基因型E659中酶活性显著上升ꎬ几乎达到了2倍ꎬ12h后开始缓慢下降到原来水平ꎬ到48h时下降了40%左右ꎮ在不耐低温基因型E646中ꎬ低温处理开始时CAT酶活性就开始下降ꎬ12h后下降了40%ꎬ到48h时几乎下降了70%左右(图5B)ꎮ低温处理后ꎬ抗坏血酸过氧化A:SOD活性ꎻB:CAT活性ꎻC:APX活性A:SODactivityꎬB:CATactivityꎬC:APXactivity图5㊀茄子幼苗低温处理下基因型E646和E659的活性氧清除系统抗氧化酶活性的变化Fig 5㊀ActivityofROSscavengingenzymesinseedlingsofthegenotypesE646andE659subjectedtolowtemperaturestress4611㊀6期包崇来等:茄子耐低温材料的筛选及其耐低温生理响应研究物酶APX的活性也呈现下降ꎬ开始处理6h时基因型E646中酶活性下降将近50%ꎬ到48h时下降了70%左右ꎮ而E659中ꎬ虽然酶活性到48h时也下降了70%左右ꎬ但是到6h之前ꎬ下降速度明显比E646慢ꎮ说明在不耐低温的材料中酶活性受低温影响较快且较强烈(图5C)ꎮ3 讨论目前ꎬ已有部分关于茄子耐低温研究的报道[9ꎬ12]ꎬ但尚未见苗期在4ħ低温处理10d后仍能保持正常生长状态ꎬ没有受到明显损害的种质材料ꎮ本研究获得的茄子耐低温基因型E659ꎬ为茄子耐低温抗性种质资源的创制和耐低温新品种的选育提供了新的基因源ꎬ并可作为茄子耐低温分子调控机制研究的新材料ꎮ植物受到低温胁迫时ꎬ会发生一系列生理生化反应[13 ̄16]ꎮ研究表明ꎬ在不耐冷水稻品系中ꎬ冷胁迫处理下的响应系统是PSII系统而不是PSI系统ꎬ最大效率因子FV/FM是光抑制反应的最佳指示剂[17 ̄19]ꎮV.Bonnecarrere等[5]研究表明ꎬ在耐低温水稻品种中ꎬ与冷敏感材料相比ꎬFV/FM比值下降幅度相对较小ꎮ不同的耐低温水稻品种中ꎬ抵抗性更强的基因型ꎬFV/FM比值更能维持在较高水平[18]ꎬ因此FV/FM比值可用来量化植物的耐低温抗性程度ꎮ本研究表明ꎬ茄子幼苗在低温胁迫下ꎬ随着处理时间的延长ꎬFV/FM和P值均呈下降的变化趋势ꎬ表明低温处理均会损坏茄子的光反应系统ꎻ低温敏感材料E646的FV/FM和P值比耐低温材料E659下降幅度大ꎬ表明E659中的光反应系统PSII受损情况较轻ꎬ光化学反应能力较强ꎬ因此可抵抗低温胁迫ꎬ在长时间的低温处理下都能维持植株的正常生长ꎮ抗氧化反应和细胞膜的完整性可衡量耐低温基因型的耐低温机制ꎬ如M.R.Morsy等[13]研究表明耐低温水稻基因型材料的抗氧化损害程度比冷敏感基因型材料相对较低ꎬ反应细胞膜完整性的电导率也是在冷敏感基因型中多升高了6倍ꎮ本研究表明ꎬ在低温处理后ꎬ2种基因型均出现膜脂过氧化反应ꎬ但基因型E659过氧化程度明显比低温敏感材料E646低ꎬ丙二醛含量在低温处理后升高倍数为E646的一半ꎮ电导率的测定结果也显示E646中电导率升高倍数是E659的1.8倍ꎮ表明在低温胁迫后ꎬ耐低温基因型E659细胞膜的受损程度明显要低于低温敏感基因型E646ꎬ因此能保持比较完整的细胞性状ꎬ保证离子通道的完整性ꎬ使离子能正常运输ꎬ防止离子的大量渗漏ꎬ从而确保植物能正常发育生长ꎮ而不耐低温基因型中由于细胞膜不完整ꎬ可能会出现离子大量外漏ꎬ从而导致细胞破碎ꎬ植株呈现倒伏㊁萎蔫ꎬ甚至死亡ꎮ李建设等[20]的研究结果也表明ꎬ低温下耐寒性强的品种能保持更好的细胞膜完整性和更低的MDA含量ꎮ张素勤等[12]研究表明ꎬ低温胁迫下耐冷性强的品种细胞受伤害程度较轻ꎬ耐冷性弱的品种则相反ꎮ在正常情况下细胞内自由基的产生和清除处于一种动态平衡的状态ꎬ自由基过高或者过低都能引起机体损失[21]ꎮ当H2O2浓度较低的时候ꎬ会阻止叶绿体sulfydryl ̄containingenzymes的产生及抑制PSII系统蛋白的合成ꎬ光氧化系统受到损害ꎮH2O2还是诱导氧化胁迫抵抗机制启动的信号因子[5ꎬ22]ꎮK.Y.Yun等[23]研究表明ꎬ冷害处理的开始阶段ꎬ氧化胁迫会引起相应基因表达ꎬH2O2浓度的升高是引导这种反应的信号ꎮV.Bonnecarrere等[5]也表明水稻冷害胁迫时ꎬ主要在前6h会发生H2O2浓度的急剧升高ꎬ而24h后浓度恢复到原来表达水平ꎮ尽管这种变化在低温胁迫过程中的功能还未知ꎬ但可能正是由于这种H2O2浓度的急剧升高ꎬ启动ROS系统的一些响应调控途径[5]ꎮ本试验中结果表明ꎬ低温处理6h基因型E659中H2O2产量迅速增加ꎬ12h后H2O2产量仍显著高于未处理情况下的产量ꎮ而E646低温处理12h前H2O2产量变化不大ꎮ到24h后ꎬ2种基因型中的H2O2产量区别不大ꎮ结果表明耐低温材料可能在低温处理开始的12h之间ꎬ通过H2O2浓度的升高来启动响应的低温防御机制ꎬ与上述研究一致ꎮ细胞在低温胁迫过程中会受到不同程度的氧化损伤ꎬ促进自由基的产生ꎬ引发膜脂过氧化作用ꎬ加速细胞内有害代谢产物的积累ꎬ造成次生伤害ꎮ氧化损伤是由于植物叶绿体中的ROS产量升高引起的ꎬ保持稳定ROS水平对应答胁迫十分重要[21]ꎮB.Demming ̄Adams等[6]提出植物通过过量能量的安全释放及ROS系统清除的抵抗机制来抵御低温胁迫ꎮ细胞中也存在着清除这些自由基的多种途径ꎬ如ROS清除系统ꎬ包括超氧化物歧化酶SOD㊁过氧化氢酶CAT和谷胱甘肽转移酶GST等在内的保护酶系统及内源抗氧化剂ꎬ防止或终止脂质过氧化ꎮ以往研究表明ꎬ许多物种中ꎬ如水稻和茄子ꎬ低温处理后很多耐低温和低温敏感材料中都会导致这些活性氧酶活性产生不同的变化[7ꎬ8ꎬ13ꎬ24]ꎬ表明活性氧导致的抗氧化代谢和氧化胁迫是植物应答低温胁迫的重要组成部分ꎮ人们认为ꎬ较高水平的酶活性能有5611植㊀物㊀遗㊀传㊀资㊀源㊀学㊀报14卷利于植物抵抗非生物胁迫[25]ꎮ李建设等[20]表明ꎬ低温下耐寒性强的品种能保持更高的SOD和POD等保护酶活性ꎮ张素勤等[12]研究表明ꎬ低温胁迫下ꎬ耐冷性强的品种能维持较高的酶活性ꎮ本研究结果显示ꎬ当茄子经低温胁迫时ꎬ耐低温基因型E659中ꎬCAT的活性在开始的6h内迅速升高ꎬ之后开始下降ꎬ但活性一直强于基因型E646ꎮSOD和APX的活性在低温处理的时候开始下降ꎬ但E659中下降明显较少ꎬ一直到48h后ꎬ都比E646中的酶活性强ꎮ因此本研究的结果也表明ꎬE659基因型能通过升高抗氧化酶活性如CAT或者尽量减少酶活性损耗如SOD和APX来维持ROS系统清除能力ꎬ抵抗低温胁迫ꎬ保证茄子植株正常生长ꎮ本试验通过分析耐低温材料和低温敏感材料之间的光抑制和氧化损伤表明ꎬ茄子耐低温基因型E659也是通过过量能量的安全释放及ROS清除系统的抵抗机制来抵御低温胁迫ꎮ推断在低温胁迫开始的早期阶段能通过累积更多的H2O2产量ꎬ启动基因表达抵抗低温胁迫ꎮ然后增强抗氧化物酶CAT的酶活性ꎬ尽量抵抗SOD和APX的失活ꎬ维持较强的ROS系统清除能力ꎬ进一步来抵抗低温胁迫ꎮ而随着胁迫时间的增长ꎬ则是通过保护系统PSIIꎬ维持较强的光反应能力和保存细胞膜完整性来抵御低温危害ꎬ维持植株正常生长ꎬ因此能在低温胁迫处理10d时还未出现冷害症状ꎮ参考文献[1]㊀日本农业渔村文化协会.蔬菜生物生理学基础[M].北京农业大学ꎬ译.北京:农业出版社ꎬ1985:281 ̄294[2]㊀阎世江ꎬ张继宁ꎬ刘洁.低温下茄子成株期几个农艺性状的遗传分析[J].植物遗传资源学报ꎬ2012ꎬ13(5):884 ̄889[3]㊀LiuYFꎬQiMFꎬLiTL.Photosynthesisꎬphotoinhibitionꎬandan ̄tioxidantsystemintomatoleavesstressedbylownighttemperatureandtheirsubsequentrecovery[J].PlantSciꎬ2012ꎬ196:8 ̄17[4]㊀JeongSWꎬChoiSMꎬLeeDSꎬetal.Differentialsusceptibilityofphotosynthesistolight ̄chillingstressinrice(OryzasativaL.)dependsonthecapacityforphotochemicaldissipationoflight[J].MolCellꎬ2002ꎬ13:419 ̄428[5]㊀BonnecarrereVꎬBorsanibOꎬDiazbPꎬetal.Responsetophotoxi ̄dativestressinducedbycoldinjaponicariceisgenotypedepend ̄ent[J].PlantSciꎬ2011ꎬ180:726 ̄732[6]㊀Demming ̄AdamsBꎬAdamsIIIWW.Photoprotectionandotherresponsesofplantstohighlightstress[J].AnnuRevPlantPhysi ̄olPlantMolBiolꎬ1992ꎬ43:599 ̄626[7]㊀SaruyamaHꎬTanidaHM.Effectofchillingonactivatedoxygen ̄scavengingenzymesinlowtemperature ̄sensitiveandtolerantcul ̄tivarsofrice(OryzasativaL.)[J].PlantSciꎬ1995ꎬ109:105 ̄113[8]㊀HuangMꎬGuoZ.Responsesofantioxidativesystemtochillingstressintworicecultivarsdifferinginsensitivity[J].BiolPlanta ̄rumꎬ2005ꎬ49:81 ̄84[9]㊀王凤华ꎬ李光远ꎬ刘宝国ꎬ等.低温胁迫对茄子幼苗APX活性的影响及cDNA序列克隆研究[J].安徽农业科学ꎬ2008ꎬ36(13):5272 ̄5277[10]㊀张泽煌ꎬ黄碧琦ꎬ陈钟佃ꎬ等.低温胁迫对茄子的伤害及茄子抗寒机理[J].福建农业学报ꎬ2000ꎬ5(1):40 ̄42[11]㊀张晓艳ꎬ徐坤.低温胁迫下砧穗互作对茄子嫁接苗叶片抗氧化酶系统的影响[J].山东农业科学ꎬ2009(6):39 ̄41ꎬ45[12]㊀张素勤ꎬ程智慧ꎬ耿广东.低温胁迫对茄子幼苗生理特性的影响[J].湖南农业大学学报:自然科学版ꎬ2006ꎬ32(4):393 ̄396[13]㊀MorsyMRꎬJouveLꎬHausmanJFꎬetal.Alterationofoxidativeandcarbohydratemetabolismunderabioticstressintworice(OryzasativaL.)genotypescontrastinginchillingtolerance[J].JPlantPhysiolꎬ2007ꎬ164:157 ̄167[14]㊀ZbierzakAMꎬPorfirovaSꎬGriebelTꎬetal.ATIR ̄NBSproteinencodedbyArabidopsisChillingSensitive1(CHS1)limitschlo ̄roplastdamageandcelldeathatlowtemperature[J].PlantJꎬ2013ꎬ75(4):539 ̄552[15]㊀刘春英ꎬ陈大印ꎬ盖树鹏ꎬ等.高㊁低温胁迫对牡丹叶片PSⅡ功能和生理特性的影响[J].应用生态学报ꎬ2012ꎬ23(1):133 ̄139[16]㊀MiuraKꎬFurumotoT.Coldsignalingandcoldresponseinplants[J].IntJMolSciꎬ2013ꎬ14(3):5312 ̄5337[17]㊀HirotsuNꎬMakinoAꎬUshioAꎬetal.Changesinthethermaldis ̄sipationandtheelectronflowinthewater ̄watercycleinricegrownunderconditionsofphysiologicallylowtemperature[J].PlantCellPhysiolꎬ2004ꎬ45:635 ̄644[18]㊀HirotsuNꎬMakinoAꎬYokotaSꎬetal.Thephotosyntheticproper ̄tiesofriceleavestreatedwithlowtemperatureandhighirradiance[J].PlantCellPhysiolꎬ2005ꎬ46:1377 ̄1383[19]㊀MaxwellKꎬJohnsonGN.Chlorophyllfluorescence:apracticalguide[J].JExpBotꎬ2000ꎬ51:659 ̄668[20]㊀李建设ꎬ耿广东ꎬ程智慧.低温胁迫对茄子幼苗抗寒性生理生化指标的影响[J].西北农林科技大学学报:自然科学版ꎬ2003ꎬ31(1):90 ̄96[21]㊀陈晓玲ꎬ张金梅ꎬ辛霞ꎬ等.植物种质资源超低温保存现状及其研究进展[J].植物遗传资源学报ꎬ2013ꎬ14(3):414 ̄427[22]㊀ChengCꎬYunKYꎬRessomHWꎬetal.Anearlyresponseregula ̄toryclusterinducedbylowtemperatureandhydrogenperoxideinseedlingsofchilling ̄tolerantjaponicarice[J].BMCGenomicsꎬ2007ꎬ8:175 ̄193[23]㊀YunKYꎬParkMRꎬMohantyBꎬetal.Transcriptionalregulatorynetworktriggeredbyoxidativesignalsconfigurestheearlyre ̄sponsemechanismsofjaponicaricetochillingstress[J].BMCPlantBiolꎬ2010ꎬ10:16[24]㊀GuoZꎬOuWꎬLuSꎬetal.Differentialresponsesofantioxidativesystemtochillinganddroughtinfourricecultivarsdifferinginsensitivity[J].PlantPhysiolBiochemꎬ2006ꎬ44:828 ̄836[25]㊀TurkanIꎬMelikeBꎬOzdemirFꎬetal.Differentialresponsesoflip ̄idperoxidationandantioxidantsintheleavesofdrought ̄tolerantP.acutifoliusGrayanddrought ̄sensitiveP.vulgarisL.subjectedtopolyethyleneglycolmediatedwaterstress[J].PlantSciꎬ2005ꎬ168:223 ̄2316611。
钙对热激诱导黄瓜幼苗抗冷性的影响

( olg f h mi r n i c n e C l eo e s y a d L f S i c ,Ti j r l iest ,T a j 0 3 7 hn ) e C t e e a i No ma Unv ri nn y ini 3 0 8 ,C ia n
中 圈分 类 号 : 4 . Q9 5 1 文献标识码 : A
Ef e to l i m n H e ts c n u e f c fCa c u o a - ho k I d c d Ch li g Re it n e i c m b r S e i g iln ss a c n Cu u e e dln s
c li g r ss a c he c c m be e d i s hiln e it n eoft u u rs e lng .
维普资讯
第 2 7卷
第 3 期
天 津 师 范 大 学学 报 ( 自然 科 学 版 )
J u n lo a j r l iest Nau a ce c iin o ra fTini No ma v ri n Un y( tr l in eEdt ) S o
摘 要 :用 C C 和 钙 的 螯合 剂 Z A 浸 种 , 究 了热 激诱 导黄 瓜 幼 苗抗 冷 性 过 程 中的 钙 效 应.结 果表 明 : 激 处 a1 GT 研 热
理 能 够提 高冷 胁 迫 条 件 下 黄 瓜 幼 苗 的 主要 抗 氧 化 酶 S D, OD 和 C T 活 性 , 低 膜 脂 过 氧 化 产 物 丙 二 醛 的含 量 O P A 降 和质 膜 透 性 , 因而 增 强 了 黄瓜 幼 苗的 抗 冷 性 ; 处 理 可 增 强 热 激 诱 导 这 一 效 应 , 由钙 的 螯 合 剂 E TA 处 理 的 效 钙 但 G 果 则相 反 , 说 明钙 参 与 了热 激 诱 导 黄 瓜 幼 苗抗 冷性 的调 控 . 这 关键 词 : 瓜 幼 苗 ;热 激 ; 冷 性 ; 黄 抗 钙
低温驯化对低温胁迫下茄子幼苗生理的影响

低温驯化对低温胁迫下茄子幼苗生理的影响摘要:茄子是一种温带蔬菜,对低温胁迫分外敏感。
本探究通过低温驯化处理茄子幼苗,探究了。
探究结果表明,经过低温驯化处理的茄子幼苗在低温胁迫下表现出更好的适应性,包括叶片叶绿素含量的增加、叶片相对电导率和丙二醛含量的降低,以及抗氧化酶活性的提高。
此外,低温驯化处理还能够增进茄子幼苗的生长和发育,提高根系活力和光合能力,从而缓解低温胁迫对茄子幼苗的不利影响。
本探究为茄子种植中低温胁迫的防治提供了重要的参考。
关键词:低温驯化;低温胁迫;茄子幼苗;生理影响引言:茄子(Solanum melongena L.)是一种温带蔬菜,对低温胁迫分外敏感。
在温度低于15℃时,茄子幼苗往往会出现“裂心”、“硬化”和生长迟缓等现象,严峻影响了茄子的生长和产量。
因此,探究低温胁迫下茄子幼苗的生理响应以及寻找适合的防治方法是茄子种植中急需解决的问题。
低温驯化是一种经过适度低温处理后提高植物抗寒性的方法。
通过低温驯化处理,植物可以在一定程度上增强对低温胁迫的适应能力,减轻低温对植物生长发育的不良影响。
然而,目前对于低温驯化对于茄子幼苗生理影响的探究还较少。
因此,本探究旨在探究,为茄子种植中低温胁迫的防治提供理论基础。
材料与方法:1. 试验材料:本试验使用大棚内栽培的茄子(品种:紫罗兰)。
茄子幼苗的生长状况良好,无病虫害。
2. 试验设计:将茄子幼苗分为两组,分别为低温驯化组和比较组。
低温驯化组的茄子幼苗在4℃下连续低温处理72小时,而比较组的茄子幼苗则在常温条件下生长。
处理结束后,将两组茄子幼苗放置在15℃下进行低温胁迫处理。
3. 生理指标测定:3.1 叶绿素含量测定:取茄子幼苗叶片样品,用乙醇提取叶绿素,并用紫外分光光度计测定其吸光度。
3.2 相对电导率测定:取茄子幼苗叶片样品,用蒸馏水浸泡后测定其电导率。
3.3 丙二醛含量测定:接受硫代巴比妥酸法测定茄子幼苗叶片中丙二醛的含量。
3.4 抗氧化酶活性测定:测定茄子幼苗叶片中超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性。
茄子抗冷性的研究进展

引起 的膜脂 过 氧化 和蛋 白质破 坏有关 。植 物体 内 的 自由基 与活性 氧具 有很 强的氧化 能力 ,对许 多 ④
茄子细胞在低温逆境 中超氧物 自由基 的增加 , 防
21 年第 1 00 期
现 代 日 艺
止膜脂过氧化 , 保持膜结构的稳定性 , 是增强植物 抗寒 性 的关键 所 在 。
生物功能分子有破坏作用。茄子体内也同时存在 些 清 除 自由基 和 活性 氧 的酶 类 和非 酶 类物 质 。 自由基 、活性 氧 和清除它们 的酶 类 和非 酶类 物质 在正 常条件 下维持平 衡状态 ,在 一定 的低 温范 围
一
寒 害的临界 温度为 721 茄子 在进行保 护地栽 培 . ̄。 2 时 , 自温度 的伤 害主要是 冷 害。 害使蔬 菜作 物 来 冷
1 渗 透调 节物质 与茄 子抗 冷性 的 关 系 . 2 l 21脯氨 酸 与 茄子抗 冷性 的关 系 。脯氨 酸 是一 …
抗冷性并不是由单基 因决定的, 而是 由一系列相 关 的直接 或 间接作 用 的基 因形 成一 个 复 杂 的调控
网 络,所 以分 子标 记成 为 广大 学 者研 究 植 物抗 冷
感 的 , 目前 在茄 子抗 冷性 生理 学研 究 的热点 主 要
王孝宣等 以耐寒性不 同的番茄 品种为材料研究 ,
结果 表 明 , 随处 理 温度 的 降低 , 溶性 糖 的含 量逐 可 渐增 加 。 温 下 ( ℃和 I ̄ 可溶性 糖 的含 量与 低 8 2C)
品种的耐寒性成显著或极显著正相关 ,且苗期与 开花 期表 现 一致 。 陈 权龙 等研 究 认 为 , 稻低 温 水
时 候 , 子 中糖 分 照 常积 累 , 溶 性 糖增 加 , 茄 可 可提
Ca^(2+)与钙调素拮抗剂对辣椒幼苗抗冷性的影响

Ca^(2+)与钙调素拮抗剂对辣椒幼苗抗冷性的影响
张化生;郭晓冬;王萍
【期刊名称】《内蒙古农业大学学报:自然科学版》
【年(卷),期】2007(28)3
【摘要】研究Ca2+和钙调素(CaM)拮抗剂W 7[N-(6-am inohexyl)-5-chloro-1-naphthalene su lfonam ide]对辣椒幼苗抗冷性的影响,结果表明:Ca2+浸种处理显著降低了低温胁迫下叶片电解质渗透率和MDA含量,提高了SOD、POD活性和可溶性蛋白质、可溶性糖的含量,而W 7浸种处理显著提高了低温胁迫下辣椒幼苗叶片电解质渗透率和MDA含量,降低了SOD、POD活性和可溶性蛋白质、可溶性糖的含量,说明Ca2+.CaM信使系统在辣椒幼苗抗冷调控过程中起着重要的作用。
【总页数】4页(P209-212)
【关键词】钙调素;Ca^2+;CaM拮抗剂;辣椒;抗冷性
【作者】张化生;郭晓冬;王萍
【作者单位】甘肃农业大学农学院;甘肃省农业科学院蔬菜所
【正文语种】中文
【中图分类】S641.3
【相关文献】
1.PEG胁迫下Ca 2+和钙调素拮抗剂W7对不同品种番茄幼苗抗氧化系统的影响[J], 张春梅;邹志荣;黄志;张志新
2.H_2O_2和Ca^(2+)对受低温胁迫香蕉幼苗抗冷性的影响 [J], 康国章;陶均;孙谷畴;王正询
3.钙通道拮抗剂和钙调素拮抗剂对白血病耐药细胞的耐药基因和细胞内Ca^(2+)浓度的影响 [J], 陈同辛;沈蕾;林梓;王耀平;应大明
4.外源钙离子(Ca~2+)对万寿菊幼苗抗冷性的影响 [J], 刘玲;周桃华
5.钙调素拮抗剂与Ca^(2+)对茄子幼苗抗冷性的影响 [J], 高洪波;陈贵林
因版权原因,仅展示原文概要,查看原文内容请购买。
简析不同处理方式对茄子连作障碍缓解效果
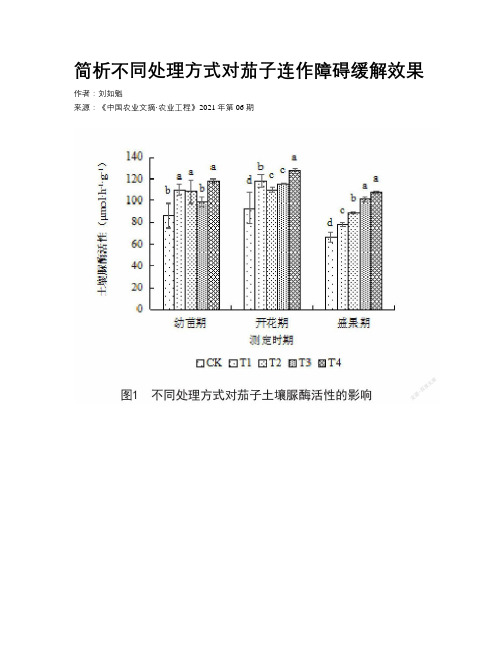
简析不同处理方式对茄子连作障碍缓解效果作者:***来源:《中国农业文摘·农业工程》2021年第06期摘要:为研究不同处理方式对连茄子缓解效果,设置5种处理方式,分别为对照(CK)、增施有机肥(T1)、增施硅肥(T2)、高温闷棚(T3)和高温闷棚配施微生物有机(T4),研究土壤微生物数量、土壤酶活性以及茄子病害发生指数,结果表明,增施有机肥、增施硅肥、高温闷棚和高温闷棚配施微生物有机处理和对照相比均能够显著提高土壤脲酶、磷酸酶、酚氧化酶和过氧化物酶活性,且显著改善茄子品质、提高产量、降低发病指数,其中高温闷棚配施微生物有机处理产量最高,发病率最低,是克服茄子连作障碍的有效措施。
关键词:茄子;连作障碍;处理方式茄子属于茄科茄属,在我国广泛种植,含有丰富的蛋白质、氨基酸和維生素等营养成分,是我国主要的栽培蔬菜之一[1]。
随着生活质量的提高,对茄子的需求逐渐增加,为满足市场需求,增加茄子产量,茄子连作现象频发,由于长期单一种植,导致严重的连作障碍,土壤元素向两极变化,供作物吸收的养分不足,土壤质量下降,长期连作还会使土壤积累盐分,影响植物细胞渗透平衡,导致养分运输和水分吸收受阻,病虫害加重,产量降低和品质变劣等问题[2]。
王芳、王敬国研究表明,茄子连作导致干物质积累量下降、根结线虫危害加重、出苗率降低[3]。
王梦怡研究表明,茄子连作第3年时土壤中蔗糖酶、脲酶和碱性磷酸酶活性降低,叶片中丙二醛(MDA)含量增高,超氧化物歧化酶(SOD)和过氧化物酶(POD)活性低于对照,长势减弱,同时茄子产量降低,出现一系列连作障碍问题[4]。
大量研究表明,土壤改良、增强植株抗性是减轻连作障碍的基本思路。
近年来连作障碍治理新方法不断增加,曲成闯等研究表明,生物有机肥可改善黄瓜连作过程中土壤理化性质变劣的现象,提高土壤酶活性和黄瓜产量,从而减缓黄瓜连作障碍的发生[5]。
吕海龙等研究表明,在连作栽培条件下,硅肥可以改善土壤微生物区系,延缓土壤微生态结构的转变,提高土壤酶活性,进而缓解设施茄子连作障碍[6]。
两种化感物质对茄子幼苗生理特性的影响

两种化感物质对茄子幼苗生理特性的影响两种化感物质对茄子幼苗生理特性的影响茄子(Solanum melongena L.)是一种重要的蔬菜作物,被广泛种植于很多地区。
茄子幼苗是茄子生长过程中的重要阶段,其生理特性直接影响到其生长和发育的结果。
然而,茄子幼苗常受到各种环境和化感物质的影响,因此研究茄子幼苗对不同化感物质的生理特性反应对于茄子的生产和可持续发展具有重要意义。
本研究选取了两种常见的化感物质,分别为赤霉素和乙烯。
赤霉素是一种植物激素,能够促进茄子生长和发育,而乙烯则具有抑制植物生长的作用。
通过在实验室条件下进行实验,观察这两种化感物质对茄子幼苗生理特性的影响。
首先,我们观察了茄子幼苗的株高和根长。
实验结果显示,赤霉素处理组的茄子幼苗株高显著高于对照组,而乙烯处理组的株高明显低于对照组。
这说明赤霉素能够促进茄子幼苗的生长,而乙烯则具有抑制茄子幼苗生长的作用。
另外,赤霉素处理组茄子幼苗的根长也显著大于对照组,而乙烯处理组的根长较短。
这表明赤霉素能够促进茄子幼苗的根系发育,而乙烯则对茄子幼苗的根系发育产生抑制作用。
其次,我们研究了茄子幼苗的叶片光合特性。
实验结果显示,赤霉素处理组的茄子幼苗的净光合速率和光合有效辐射利用效率均显著高于对照组,而乙烯处理组的茄子幼苗则呈现出相反的结果。
这说明赤霉素能够提高茄子幼苗的光合效率,从而增加光合产物的合成和积累,而乙烯则对茄子幼苗的光合产物合成产生负面影响。
最后,我们研究了茄子幼苗的抗氧化酶活性。
抗氧化酶是植物应对氧化胁迫的重要防御系统,能够抑制有害氧化物的积累。
实验结果显示,赤霉素处理组茄子幼苗的超氧化物歧化酶和过氧化物酶活性显著高于对照组,而乙烯处理组的抗氧化酶活性则较低。
这表明赤霉素能够增强茄子幼苗的氧化胁迫抵抗能力,而乙烯则对茄子幼苗的氧化胁迫抵抗能力产生负面影响。
综上所述,赤霉素和乙烯对茄子幼苗的生理特性产生明显影响。
赤霉素能够促进茄子幼苗的生长和发育,增强其光合能力和抗氧化能力,而乙烯则具有抑制茄子幼苗生长和发育的作用,降低光合效率和抗氧化酶活性。
不同来源茄子耐低温性的鉴定

不同来源茄子耐低温性的鉴定
阎世江;张继宁;刘洁
【期刊名称】《河北科技师范学院学报》
【年(卷),期】2013(027)001
【摘要】对不同遗传来源的3,4,6,9,10,12,14,40,250,325号等10份茄子高代自交系进行低温处理,处理方法为:前2d白天15℃,夜间10℃,光照度4 000 lx,从第3天开始,白天10℃,夜间7℃,光照度4 000 lx,处理10 d.然后调查耐低温性、株高、茎粗、POD活性、SOD活性、MDA的质量摩尔浓度、脯氨酸的质量分数等指标.结果表明,常温下上述性状差异不显著,经低温处理差异显著,耐低温性与处理后的株高、茎粗、POD活性、SOD活性密切相关.材料14,4号的耐低温性较强;材料325,9号的耐低温性较弱;材料3,250,10,40,12号的耐低温性居中.
【总页数】4页(P9-12)
【作者】阎世江;张继宁;刘洁
【作者单位】山西省农业科学院科技情报研究所
【正文语种】中文
【中图分类】S641.1
【相关文献】
1.不同来源黄瓜亲本苗期耐低温鉴定 [J], 闫世江;司龙亭;马志国;杨佳明
2.不同茄子品种田间病害调查及其抗病性鉴定 [J], 朱丽梅;崔群香;蔡元琴;郭迎迎
3.茄子苗期耐低温性鉴定方法初步研究 [J], 易金鑫;陈静华
4.茄子不同品种耐低温特性室内鉴定初报 [J], 高志奎;武占会;孙会军
5.不同耐低温性茄子果实发育过程中主要营养成分变化的研究 [J], 芮三亚;成玉富;王建忠
因版权原因,仅展示原文概要,查看原文内容请购买。
三种植物提取物对茄子黄萎病抗性效果的研究的开题报告

三种植物提取物对茄子黄萎病抗性效果的研究的开题报告
一、选题背景
茄子是重要的蔬菜之一,但茄子黄萎病(Verticillium wilt)对茄子的生长和发展产生了严重的影响,甚至会导致茄子的死亡。
目前,植物提取物已被广泛应用于农业生产中的病害防治,其中包括对茄子黄萎病的防治。
因此,本研究旨在探讨三种植物提取物对茄子黄萎病的抗性效果。
二、研究目的
1. 研究三种植物提取物对茄子黄萎病的抗性效果;
2. 探究三种植物提取物影响茄子生长和发育的影响;
3. 为农业生产提供茄子黄萎病的抗性控制策略。
三、研究内容
1. 收集茄子黄萎病病原体;
2. 准备三种植物提取物;
3. 测定三种提取物对茄子黄萎病的抗性效果;
4. 观察病理变化和测定茄子生长发育指标;
5. 数据统计和数据分析。
四、研究方法
1. 茄子黄萎病病原体的收集:从受感染的茄子植株上采集病原体;
2. 植物提取物的制备:分别制备三种植物提取物;
3. 治疗效果的测定:采用人工接种法,分别施以三种提取物,观察茄子黄萎病的发病情况;
4. 叶片颜色和立枯节数量的测定;
5. 茄子重量和花果长度的测定;
6. 统计学分析。
五、预期成果
1. 研究三种植物提取物对茄子黄萎病的抗性效果,明确其防治效果;
2. 探究三种植物提取物对茄子生长和发育的影响,分析其优劣;
3. 为农业生产提供茄子黄萎病的抗性控制策略。
钙调素拮抗剂与Ca 2+对茄子幼苗抗冷性的影响

细 胞 质膜 透性 的测 定 按 杨根 平 等 】 的方 法 ,以相对 电解 质渗 透 率 表示 。丙 二 醛 ( A MD )含 量 的测
定采用硫代 巴比妥酸法 ,以 t o・ I L lg1刚 表示 M A含量。超 氧化物歧化酶 (O )活性 的测定按 m D SD
对茄子幼 苗抗冷性 的影响 ,结 果表 明 :聊 浸种处 理 显著 提高 了低 温胁 迫 下茄 子幼 苗 叶片 电解 质渗 透率 和 M A含量 ,降低 了 S D、C T活性 和可溶性 蛋 白质含量 ;而 C2 D O A a 浸种 处理显著 降低 了低温胁 迫 下叶 片电解
质渗透 率和 M A含量 ,提 高 了 S D、C T活性 和可 溶性蛋 白质 含量 ,说 明 C 2 CM 信使 系统 在 茄子 幼苗 D O A a+- a 抗冷调 控过程 中起着重 要的作用 。 关 键词 :钙 调素 ;CM拮 抗剂 ;C 2 a a ;茄子 ;抗 冷性
1 材 料 与 方 法
试验 于 20 00年 2~4月 和 20 年 2~4月 分 两 期 在 河 北 农 业 大 学 进 行 。 茄 子 ( o nm m l ea 01 S/ u e gn a o
L )品种为丰研 2 (ega ) . 号 Fny 2 。种子经 0 1 K n d n .% M O 消毒后 ,分成 3 份。分别设 4 m l CC2 0m o・ L a1
中图分类 号 :S6 12 4 . 文献标识 码 :A 文章 编号 :0 1.5 X (O2 30 4—4 5333பைடு நூலகம்20 )0 . 30 2
茄子 属 典 型 的喜 温 蔬 菜 ,低 温冷 害 是 制 约 茄 子 越 冬 栽 培 的主 要 障 碍 因子 。研 究 表 明 ,第 二 信 使 C2 在调节植物细胞对低温¨2 a ’ 、高温 ̄4、盐害 等环境胁迫信号转导过程 中起着重要的作用。CM 】 33 , 】 a 作 为 C2 a 的受 体 ,通 过 与 c2 合 而 激 活 一 系 列 的靶 酶 和 非 酶 蛋 白 ,从 而 调 控 生 理 代 谢 及 基 因表 a 结 达 J 。w7 [ 一(-mnhx1 5clr 1npta n lnmi )作 为 一 种 新 的 C 拮 抗 剂 ,可 导 致 N 6a i ey )-一h o 一ahhl es f a d o o- e uo e M a c2 C 信使 系统 功 能发 生 障碍 ,与 C Z ( 丙 嗪 ) F ( 氟拉 嗪 )等其 它 C 拮抗 剂 相 比 ,W7 a ・ a M P 氯 、T P 三 M a 不 影 响细 胞膜 的结 构 和 流动 性 【 】 因此利 用 w7和 c2 理 阻 碍 和增 强 c 2 C 信 使 系统 是 目前 7 。 , a 处 a ・ a M 探 索活 体植 物 cs C 信 使功 能 的重 要 手 段 。 目前 国 内外 对 c 2 a ・a M a 和 C 的 研 究 多 是 以 玉米 、水 稻 M a 等 农作 物 为 材料 ,尚未见 有 关 c2 C 拮 抗 剂 与茄 子 抗冷 性 的研 究 报 道 。 a 和 a M 本试 验 利用 w7和 c 2 a 浸种 处 理 研究 c2 C 对 低温 胁 迫下 茄 子 幼 苗膜 脂 过 氧化 、保 护 酶 活 性 a ・a M 及保 护性 物 质含 量 等方 面 的影 响 ,从 阻碍 和 增 强 c2 C 信 使 系 统 转 导 两 个 角 度 揭 示 c2 C 信 a ・a M a a M 使 在抗 冷 中的作 用 机 理 ,为 茄 子 乃至 蔬 菜设 施 栽 培 防止 低温 冷 害 提供 理 论 依据 。
不同茄子砧木幼苗抗冷性的筛选

不同茄子砧木幼苗抗冷性的筛选
高青海;徐坤;高辉远;吴燕
【期刊名称】《中国农业科学》
【年(卷),期】2005(038)005
【摘要】以济农2000为对照,研究了低温胁迫下不同茄子砧木幼苗的形态及生理
生化变化.结果表明,低温胁迫初期,试材对低温最敏感,叶片电解质渗漏率、丙二醛含量、SOD活性和光补偿点等迅速增加,而POD活性、CAT活性、AQY、光合速率、Fv/Fm、ФPSⅡ等迅速降低;之后,叶片的保护酶活性、光合速率、Fv/Fm逐渐升高.随低温胁迫时间延长,叶片的保护酶活性和光合速率又有所下降,但赤茄与托鲁巴姆
下降幅度较小,仍维持较高水平.通过综合比较筛选,试材中以赤茄与托鲁巴姆的抗冷性较好,台茄和耐病VF的抗冷性较差,但仍比对照品种济农2000强.
【总页数】6页(P1005-1010)
【作者】高青海;徐坤;高辉远;吴燕
【作者单位】山东农业大学园艺科学与工程学院;山东农业大学园艺科学与工程学院;山东农业大学生命科学学院,泰安,271018;山东农业大学园艺科学与工程学院【正文语种】中文
【中图分类】S6
【相关文献】
1.壳聚糖对茄子幼苗抗冷性的影响 [J], 刘术均;张青;惠成章;杨光;方伟;鲁娇娇;姜闯
2.不同砧木嫁接对苦瓜幼苗抗冷性的影响 [J], 杜卓涛;朱白婢;赖彩余;商桑;田丽波;
邹凯茜;朱国鹏;林师森
3.5-氨基乙酰丙酸对茄子幼苗抗冷性的影响 [J], 刘辉;黄娟
4.茄子抗冷砧木的筛选和嫁接苗抗冷性研究 [J], 乜兰春;陈贵林;高洪波
5.VA菌根菌抗冷菌株的筛选及其对茄子抗冷性的影响 [J], 柏素花;董超华;刘新因版权原因,仅展示原文概要,查看原文内容请购买。
茄子采后冷害发生生理机制及前处理措施的研究

茄子采后冷害发生生理机制及前处理措施的研究茄子是典型的冷敏性蔬菜,贮运不当极易遭受冷害,严重影响其品质。
本研究以长茄和圆茄为试材,通过测定冷害指数、相关生理生化指标及质构指标,研究不同贮藏温度对茄子冷害发生的影响,了解其冷害发生的生理机制;以长茄为试材,分析研究不同前处理方法对抑制茄子冷害发生的作用,再通过测定冷害指标、质构指标、微生物指标及扫描电镜观察,探究了 UV-C结合草酸处理对抑制茄子冷害发生及抑菌效果的作用,为了解茄子采后冷害发生的机制和茄子贮运提供理论依据。
研究结果表明:(1)不同贮藏温度下茄子的冷害发生程度不同。
2℃时两种茄子在贮藏3d时出现冷害症状,5℃时在6d时出现冷害症状。
2℃时茄子的冷害指数极显著高于5℃,细胞膜透性和丙二醛含量均显著高于5℃,失重率、呼吸强度和SOD、POD、CAT活性显著低于5℃,粘附性显著高于5℃,弹性极显著低于5℃;8℃贮藏的茄子在18d不发生冷害,并且较好地保持了茄子的TSS含量、SOD、POD、CAT活性、粘附性、内聚性、弹性与咀嚼性。
在相同贮藏温度下,长茄子冷害程度相对比圆茄子严重。
(2)不同前处理对抑制茄子冷害发生效果不同。
冷激(0℃冷水浸果30min)、热激(45℃热水浸果30s)、UV-C(7kJ/m2)、草酸(6mmol/L草酸浸果10 min)、水杨酸(1.5mmol/L水杨酸浸果10 min)、壳聚糖(1%壳聚糖浸果1 min)处理均可减轻冷害程度,抑制失重率的升高,维持TSS含量和POD活性,其中UV-C处理效果最佳,草酸处理其次。
(3)UV-C结合草酸处理有效抑制茄子冷害的发生,保持果实TSS含量,抑制细胞膜透性和MDA含量的升高,维持POD、SOD、CAT活性,保持茄子粘附性、内聚性、弹性、咀嚼性和细胞完整性,抑制茄子表面霉菌和酵母菌的生长,降低菌落总数。
低温胁迫程度对茄子幼苗生理特性的影响

低温胁迫程度对茄子幼苗生理特性的影响张素勤;耿广东;程智慧【期刊名称】《山地农业生物学报》【年(卷),期】2008(027)002【摘要】通过5℃和8℃的低温处理,研究了低温对茄子幼苗的生理效应.结果表明,随着温度的下降,茄子叶片内SOD活性增大.在5℃低温下,SOD活性增加得最多;在低温条件下,茄子幼苗叶片内POD活性被激活,经5℃和8℃低温处理后,其体内POD活性分别比对照增加292%和113%,且差异均达到极显著水平;CAT活性随温度下降而呈现先降后升的变化趋势;MDA含量随温度的下降而增加,经5℃和8℃处理后,茄子幼苗体内MDA含量比对照分别增加24.94%和30.22%;脯氨酸和可溶性蛋白质含量也随温度的下降而增加.【总页数】4页(P119-122)【作者】张素勤;耿广东;程智慧【作者单位】贵州大学,农学院,贵州,贵阳,550025;西北农林科技大学,园艺学院,陕西,杨凌,712100;贵州大学,农学院,贵州,贵阳,550025;西北农林科技大学,园艺学院,陕西,杨凌,712100;西北农林科技大学,园艺学院,陕西,杨凌,712100【正文语种】中文【中图分类】S641.101【相关文献】1.低温胁迫对茄子幼苗生理特性的影响 [J], 张素勤;程智慧;耿广东2.低温胁迫对茄子幼苗耐冷生理特性的影响 [J], 高秀瑞;潘秀清;武彦荣;李冰3.外源NO对低温胁迫下茄子幼苗生长及生理特性的影响 [J], 刘旭;林碧英;吴宏琪;李小琼;祖颖达;钟路明;王爱红4.外源褪黑素对低温胁迫下大白菜幼苗生长及生理特性的影响 [J], 李欠敏;杨琳;段巧红;黄家保5.低温胁迫对茄子单性结实材料生理特性的影响 [J], 高秀瑞;李冰;高洪波;武彦荣;潘秀清因版权原因,仅展示原文概要,查看原文内容请购买。
一种茄子及提高茄子抗冷性的处理方法[发明专利]
![一种茄子及提高茄子抗冷性的处理方法[发明专利]](https://img.taocdn.com/s3/m/fee852c52af90242a995e5ca.png)
专利名称:一种茄子及提高茄子抗冷性的处理方法
专利类型:发明专利
发明人:赵云峰,吴玲燕,王梦影,韩亮亮,谢锦明,姜骅峰,吕晴申请号:CN201710326817.1
申请日:20170510
公开号:CN107156282A
公开日:
20170915
专利内容由知识产权出版社提供
摘要:本发明公开了一种茄子及提高茄子抗冷性的处理方法,涉及农业技术领域。
该提高茄子抗冷性的处理方法包括:将茄子在水杨酸溶液中浸泡18‑20min,然后进行辐照处理30‑60min;将经过辐照处理的茄子进行预冷处理2‑3h,然后将茄子放入保鲜袋中,并置于3‑5℃的温度下冷藏。
本发明中的处理方法可以有效控制茄子采后细胞膜透性的升高,抑制丙二醛含量的积累,减缓可溶性糖、可溶性蛋白含量的下降,促进游离脯氨酸含量的升高。
经过上述处理方法处理过的茄子在冷藏过程中的抗冷性能明显提升。
申请人:盐城工学院
地址:224000 江苏省盐城市希望大道中路1号
国籍:CN
代理机构:北京超凡志成知识产权代理事务所(普通合伙)
代理人:赵丽娜
更多信息请下载全文后查看。
钙提高茄子幼苗抗寒力的研究

钙提高茄子幼苗抗寒力的研究
王凤华;林德清;王贵学
【期刊名称】《四川农业大学学报》
【年(卷),期】2005(023)002
【摘要】以茄子幼苗为试材,研究了茄苗越冬期间施用CaCl2对茄苗抗寒力的影响.结果表明:CaCl2处理可以显著提高茄子幼苗的抗寒力.进一步的研究表明CaCl2处理提高茄苗抗寒力主要表现在促进了可溶性糖、热稳定蛋白质和游离Pro的积累.【总页数】4页(P192-194,222)
【作者】王凤华;林德清;王贵学
【作者单位】重庆大学,生物工程学院,重庆,400044;西南农业大学,园艺学院,重庆,400716;重庆大学,生物工程学院,重庆,400044
【正文语种】中文
【中图分类】S641.1;Q945
【相关文献】
1.PLTPA提高西葫芦幼苗细胞抗寒力的生理生化基础 [J], 郝联芳;王小婵;金黎
2.硝酸钙胁迫对茄子幼苗生长及生理生化指标的影响 [J], 郑兴莲;申书兴;王彦华;陈雪平;罗双霞;郭丽娟
3.亚适温催芽对提高西瓜幼苗根系抗寒力的效应 [J], 陈垣;郭凤霞;李妮;熊祥云
4.抗寒剂CR-4提高玉米幼苗抗寒力及质膜5'-核苷酸酶冷稳定性的研究 [J], 孙龙华;简令成;王瑞萍
5.植物抗寒剂提高黄瓜幼苗抗寒力及细胞膜系统冷稳定性的研究 [J], 张红;简令成;李广敏
因版权原因,仅展示原文概要,查看原文内容请购买。
茄子低温障碍的防治方法

茄子低温障碍的防治方法茄子在育苗或者提前、延晚栽培过程中,常常会遇低温冷害和冻害的侵袭,我们将这种现象称之为低温障碍。
它会严重影响到茄子的生长发育,对后期的产量也会造成严重的影响,那么该怎么防治低温障碍呢?随松鼠哥一起来了解了解吧。
1、近地面覆盖在温室栽培时,可以用薄膜、草帘、保温被、地膜等多种材料进行组合覆盖,从而达到保温的目的。
因为不同的保温材料的增温效果不同,一般每增加一层薄膜可以提高2-3度,减少热损耗30%-50%,而覆盖第二层即可增温6度注意。
如果是露地栽培,早期可以采用地膜近地面覆盖的形式覆盖幼苗,使得幼苗整体处于地膜以下,这样起到的保温效果更好。
2、补充加温在寒流来临前,仅仅依靠温室和多层覆盖条件是不能满足茄子对于温度的需求的,这时就需要立即采取加温措施。
根据自身的条件可以采取火道加温、电热线、小型水暖锅炉等多种手段补充热量。
但是一定不能用明火熏烟的方式,以免烟雾熏苗,同时还会影响到生命安全。
如果有地热、电厂废热等条件地区可输管道利用这些热源提高土壤温度。
3、播种育苗在播种时应该确定好播种时期和定植日期,尽量选择寒尾暖头的无风晴天播种或定植,确保幼苗能安全出苗和成活。
在育苗时建议营养钵育苗,因为黑色塑料营养钵具有白天吸热,夜温保温护根的作用,一般营养钵的温度和外界温度相差20度注意,能确保幼苗不受冻害的生长,而且还能进行低温炼苗,增强抗寒能力。
4、叶面喷肥在低温环境,幼苗的根系吸收能力较差,这时需要在叶面上喷施光合微肥,确保幼苗因营养不足而造成缺素症。
同时还可以叶面喷施米醋,能起到抑菌驱虫的主要,米醋和白糖以及过磷酸钙混合使用,能增加叶肉含糖量,提高叶片硬度,提高抗寒性。
在低温时要注意不要棚室任何生长类的生长调节剂,以免将其幼苗的抗寒性。
以上是茄子低温障碍的防治方法介绍,希望能帮助到你,xx。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
23 7 C + . W 与I 对冷胁迫下茄子幼苗叶片S D活性的影响 W O 较高的S 活性是植物抵抗逆境胁迫的生理基础。图3 D O 表明, 低温胁迫前, 种处理叶片SD活 3 O 性并无显著差异。在低温胁迫 0 1 h 一 2 范围内 C'处理和对照 SD活性同步上升,在低温胁迫 1- a. O 2 4h 8 范围内C 十 a 处理SD t O 活性一直显著高于对照。随低温处理时间的延长, 7 W 处理叶片SD活性 O 呈下降趋势且显著低于C2处理和对照。C + a . a 处理在低温胁迫2 h SD 2 4 O 活性达最高峰,比 时 对照提 前 1h 2 。在胁迫2 h W 处理和对照 SD 4 时, 7 O 活性分别比C 十 a 处理降低 7. %和 2. %。 ' 24 1 28 0 这说明 W 通过抑制 C2" 7 a CM形成降低了SD ' a O 活性, a 处理能显著促进 C ' M形成并提高SD C2 十 a -. 2C O 活性, 并使之维持在较高的水平, 减轻了呀 自 基对茄子幼苗的伤害。 由 2 W 与。2对冷胁迫下茄子幼苗叶片 C T . 4 7 + A 活性的影响 C] A, 对清除组织中的氏仇等有害自 基有重要的作用。由图4 由 可以看出,随低温胁迫时间的延 长, + 份 处理和对照叶片CT A 活性均呈现先上升后下降的变化趋势, 并且 C2处理在整个胁迫过程 a + 中显著高于对照和W 处理。在整个胁迫范围内W 处理 C T 7 7 A 活性呈缓慢下降的变化趋势。在低温胁
(屠 山 1 笋 n
2 结果与分析
21 与Ct对低温胁迫茄子幼苗叶片细胞质膜透性的影响 . W7 a' 从图1 看出, 可以 随低温胁迫时间的 延长, 种处理电解质渗透率均表现为先上升后下降, 3 其中 W 处理始终高于Ct处理和对照 ( 7 a 十 去离子水处理) 在低温胁迫3 h 3 ; 6 种处理电解质渗透率均达 时 最高峰, 此时W 处理分别比Ct处理和对照增加 1 . %和 4. %; 7 a + 36 46 75 8 低温胁迫3 h 3 6 种处理电 后 解质渗透率均呈下降的 变化趋势。 这表明W 处理使茄子幼苗细胞膜渗透率显著增加, 7 加重冷害对幼 苗的伤害,而Ct处理却明显降低膜的渗透率, a + 提高 幼苗抗冷性。 22 , . W 和。2对冷胁迫下茄子幼苗叶片M A含f的影响 + D 不同 处理茄子幼苗经。 4 h 一 8 低温胁迫后,叶片中过氧化产物 M A D 含量变化见图2 。低温处理 前, a , 、 Ct W 对照的M A ' 7 D 含量均较低,随低温胁迫时间的延长, 种处理 M A含量均呈上升趋 3 D 势。 ,处理在整个胁迫过程中M A a D 含量显著增加。与 W 和对照相比, a 处理显著降低 了M A 7 C' . D 的 含量。在胁迫4 h 8 时,C + a 处理叶片M A t D 含量分别比W 处理和对照降低了 5. %和 2. %. 7 1 2 7 58 6 这表明W 通过抑制 C " 的形成 7 a - t . C M 使茄子幼苗体内h)含量增加, t i 人 加剧了冷害对幼苗的伤害,而 c+ a 通过促进 份 "CM的产生减缓了 l -a 低温的伤害, 提高了 幼苗的 抗冷性。
子排列于 铺有滤纸的培养皿中, 8 催芽7 h 将发芽整齐的 2' C 2 。 种子分别播于装有蛙石的7 孔穴盘中, 2 在日 光温室内 培育幼苗。 昼温2 - 0 夜温 1- 0 其它管理与一般日 5 9, 2 8 5 9, 1 7 光温室相同。子叶展平 后每隔2 用 1 营养液 ( 本山 d / 2 按日 崎茄子营养液配方配制)浇灌 1 7 后每隔2 用完全营养液 次, d d
6
‘C' .处理 C " e m m * .hr e i
0对照 c n o o t l一 r
曰产 洲
(居 娇
月
1‘ 曰
. 处理 W7 川 曰. W, 卜 . .对暇 c n o ot l r
几J
罗 。日 盆 )
几 1 ,
山
0 4 1 2 2 4 肠 8
细 质膜 性的 按杨 平等a 方 以 对电 渗透 表 丙二 (D) 的 胞 透 测定 根 (的 法, 相 解质 率 示。 醛 M A 含量 测 )
定采用硫代巴比 妥酸法[,以,洲--F ( 9 ) 川 1 ' g M表示 M A D 含量。超氧化物歧化酶 ( D S )活性的测定按 O
收祠 日期: 01 - 20 - 1;修回日 : 02 0 - 7 0 6 7 期 20 - 1 0 若金项目:河北省自然科学基金资助项 目 (91 ) 392 ;农业部 “ 5 蕊菜遗传与生理实验室 资助项目
浇 次。 灌1 待幼苗 达三叶一 选生长正常的 心时 移人LH5 G R2 - 光照培养箱内 ( 0 光强印iom ,‘ u l - , n. . 2一
每天光照 1 h, 2 先于1 1 0( 夜)环境下预处理 1 , ) 2 0 昼/ /9 d 再分别置于5 ℃下处理 。 2 2, ,1, 3, 4 6 4 h 然后每天取样测定。 8 ,
对茄子幼苗抗冷性的影响, 结果表明: 7 W 浸种处理显著提高了低温胁迫下茄子幼苗叶片电解质渗透率和 M A含量,降低了SD C T D O , 活性和可溶性蛋白质含量; C'浸种处理显著降低了低温胁迫下叶片电解 A 而 a 质渗透率和 M A D 含量, 提高了SD CT O , 活性和可溶性蛋白质含量, A 说明 Cz " a' M信使系统在茄子幼苗 C . 抗冷调控过程中起着重要的作用 关健词: 钙调素; a Cm拮抗剂; a 茄子; Ct; 抗冷性 中圈分类号: 612 文献标识码: S 4, A 文幸编号: 53 5X 02 0- 4- 01 33 ( 0) 0 3 4 - 2 32 0
卜1
勺
么c 犷处理 c r tet 犷ta n em
‘ 0
绷 绷
" ' c 处理 C 'ra e a at t m "e m
M 处理 W ra et W7 7 t n t m e .对照 c nrl ot o
『卜 留吕 0 0 协 攀 职 (I
月
. 处理 W7 a et W7 r m t t n e .对照 C n o ot l r
晃
艺 学 报 20, () 3 6 02 2 3: - 4 9 2 - 4 2
Xoa utre rc l a 5 u -
钙调素拮抗剂与 C t 对茄子幼苗抗冷性的影响 a +
高洪波 陈贵林 ‘
( 河北农业大学园艺学院, 保定 01 1 70 ) 0
摘 要: 究 调素 ( 拮 剂W (- n hy -co latl som e 和C ' 钙 研 C ) 抗 7 (av el 5hr -phe un i) a a M N 6 a x) l nha e ad - - a n l f Z
不影响细 胞膜的 结构和流 动性(1 因 7 3 此利用W 和CZ处理阻碍和 , 4 7 a 十 增强C -a a CM信使系统是目 " 前
探索活体植物 C 十 : a 《a t M信使功能的重要手段。目 前国内外对 C + CM的研究多是以玉米、 a和 a t 水稻 等农作物为材料, 尚未见有关 C ' a a 和CM拮抗剂与茄子抗冷性的研究报道。 t 本试验利用W 7 和 CZ浸种处理研究 C" a a ' a - CM对低温胁迫下茄子幼苗膜脂过氧化、保护酶活性 及 保护性物质含量等方面的影响, 从阻碍和增强 C" . a - CM信使系统转导两个角度揭示 C' ・a a CM信 十 使在抗冷中的作用机理,为茄子乃至蔬菜设施栽培防止低温冷害提供理论依据。
3 期
高洪波等:钙调素拮抗剂与Ct对茄子幼苗抗冷性的影响 ‘
迫2 I W 处理和对照C T 4 t 时, 7 A 活性分别比C ‘ a 处理降低 7. %和 3.1 t 49 0 1 %。这表明 W 通过抑制 7 7 C 'CM的形成降低了CT a ". ? A 活性,而 Ct处理由于促进 CZ . a + a CM信使系统的形成增强 了CT ' . A活 性, 减缓 了HO 对茄子幼苗的伤害, Z2 提高植株抗冷力。
书 通讯作者 (uo6 c ea m e. Ah r s d a tr , o p r a )
园
艺
学
报
2 9卷
Gnpi 等’ 方 测 以 制 蓝 噢 (B) 还 0 为 个 活 单 过 化 i ol 〔的 法 定, 抑 氮 四 NT 光化 原5 一 酶 性 位。 氧 氢 a oi 。 n t 〕 。 %
- 一
0‘ - s e 一 - 0 1 2 2 4
肠
一 4 8
时间 t e旬 i m 时间 . e i () m h '" 圈1 ” 和〔'处扭对冷容茄苗 , a 皿质扭性的影 晌 W 和 C'处理对冷容药苗 N 含t的影晌 , .小 M
茄子属典型的 喜温蔬菜, 低温冷害是制约茄子越冬栽培的主要障碍因子。研究表明, 第二信使
C ‘ 调节 a 在 植物细 ` 胞对低温() 高温( )盐害5 1 , 2 3 , 4 (等环境 ) 胁迫信号转导过程中 重要的 起着 作用。C a M
作为 C + a 的受体, t 通过与 CZ结合而激活一系列的靶酶和非酶蛋白, a 十 从而调控生理代谢及基因表
1 材料与方法
试验于20 年2 4 ( - 月和2 1 - 月分两期在河北农业大学进行。茄子 (on mle 0》 0 年2 4 0 Sau egn l m o r a
L) 种为 研2 ( na2。 经。I K n4 毒 分 份。 别设4 n oL’C 品 丰 号 F gn 种子 . MO消 后, 成3 分 e y ) % 0 l C 1 l " a2 m 一 ( 析 、 0 f 1 -W (g 公司 ) 去 水 〔 分 纯) 3 n " 1 S m 产品 和 离子 对照) 种3 处 浸种1h 0 L 7 a } i 浸 个 理。 2 后将种
达s W ( (.nel - hm -phe some 作 一 新的C 拮 剂, 导 ( 7 6 uhy 5 l - atl u n i〕 为 种 ) . N - ox ) - o l ha e ad - c n n l f a 抗 可 致 M