古细菌RNase HII与金属离子结合的热力学研究
国家自然科学基金委员会生命科学部2014年度青年基金项目

国家自然科学基金委员会生命科学部2014年度青年基金项目第26卷第12期2014年12月生命科学Chinese Bulletin of Life SciencesV ol. 26, No. 12Dec., 2014文章编号:1004-0374(2014)12-1342-62国家自然科学基金委员会生命科学部2014年度青年基金项目项目名称申请人依托单位1微生物学植物乳杆菌激活NHE8信号通路并在炎症性肠病中发挥保护作用可能的机制探究刘畅上海交通大学南极乔治王岛土壤中产蛋白酶细菌和胞外蛋白酶的多样性以及细菌新种属和新型适冷蛋白酶资源周明扬齐鲁工业大学II类NAD-依赖型异柠檬酸脱氢酶的功能与进化机制研究王鹏安徽师范大学黄河三角洲盐生植物内生菌多样性差异及功能基因分析夏志洁山东师范大学活性污泥中稀有微生物类群BFB的多样性及其环境响应研究郗丽君中国石油大学(华东)北冰洋产多糖细菌多样性及多糖特性张炳照广州中国科学院先进技术研究所青海油田原油与采出液水相微生物差异机制及原油来源菌种资源多样性研究蔡曼中国科学院微生物研究所一个特殊新疆鹰嘴豆根瘤菌种群自然进化规律及其进化机制的研究张俊杰郑州轻工业学院新疆藜科5种盐生植物内生细菌多样性及耐盐促生菌株评价王宏飞中国科学院新疆生态与地理研究所红树林生态系统中弗兰克氏菌的多样性及其数量分布研究刘敏中国热带农业科学院中国发网菌科黏菌分类及分子系统学研究张波吉林农业大学中国座坚壳属真菌的分类与分子系统学研究马海霞中国热带农业科学院基于转录组测序的腹黏菌亚纲和发网菌亚纲黏菌核糖体序列测定及分子系统学研究亓宝吉林农业大学中国链格孢菌大孢子种的形态学与分子系统学研究邓建新长江大学中国假网衣科地衣系统分类学研究张璐璐山东师范大学松乳菇多糖(LDG-A)调控巨噬细胞免疫应答活性的分子机制侯怡铃西华师范大学茶树病原真菌和内生真菌的多样性及其分布规律刘芳中国科学院微生物研究所丝状子囊菌ITS分子进化研究李熠福建农林大学武夷山国家自然保护区凋落枯枝暗色丝孢真菌分类研究马立国山东省农业科学院中国菌寄生属资源、分类及该属的分子系统学研究曾昭清中国科学院微生物研究所不同生境条件下齿瓣石斛不同生长时期菌根真菌多样性研究邵士成中国科学院西双版纳热带植物园中国担子地衣的分类及分子系统研究王欣宇中国科学院昆明植物研究所中国珀扎若拉属和偏脚菇属分类及分子系统学研究何晓兰四川省农业科学院裂丝盖伞复合群的隐存多样性及生物地理学研究范宇光长白山科学研究院疫霉属(Phytophthora)DNA条形码的选择与评价兰成忠福建省农业科学院毛醌素生物合成途径解析及其调控殷华中国科学院天津工业生物技术研究所阴沟肠杆菌SDM中2,3-丁二醇手性形成及代谢调控机制研究李理想上海交通大学产碱假单胞菌中龙胆酸代谢直接水解途径的研究刘琨上海交通大学沙门氏菌DNA磷硫酰化修饰蛋白复合物的功能研究成秋香上海交通大学聚醚类盐霉素的生物合成机理解析姜春艳上海交通大学寡糖转运在嗜碱芽孢杆菌 N16-5 半纤维素利用中的作用研究宋亚囝天津科技大学SpTrz2调控粟酒裂殖酵母线粒体介导的细胞凋亡机制的研究商巾杰南京师范大学DOI: 10.13376/j.cbls/2014186第12期1343国家自然科学基金委员会生命科学部2014年度资助项目铜绿假单胞菌合成鼠李糖脂对环境胁迫条件的响应机制研究姜天翼山东建筑大学开发适用于链霉菌的新型诱导表达系统王为善中国科学院微生物研究所激活蛋白AP1参与茉莉酸甲酯诱导灵芝酸合成的调控机理任昂南京农业大学代谢产物调控碳降解物阻遏作用和蛋白质组资源分配尤从慧深圳大学温度对法夫酵母MK19虾青素合成调控机制的研究苗莉莉中国科学院微生物研究所高山被孢霉脂质合成与苯丙氨酸代谢相关性及调控机制研究王鸿超江南大学蛋白乙酰化修饰对天蓝色链霉菌发育分化的调控机制研究赵维中国科学院上海生命科学研究院枯草芽孢杆菌解聚酶YwtD调节γ-聚谷氨酸链长的结构基础研究曾菊梅中国科学院成都生物研究所大肠杆菌表达异源蛋白质时的多态性研究赵云中国科学院生物物理研究所全局性调控子GacA对假单胞菌L-肉碱生物合成的调控机制研究黄娇芳中国科学院上海高等研究院hmsT 3'非翻译区介导的mRNA稳定性研究朱慧中国医学科学院病原生物学研究所微生物细胞外还原制备纳米粒子过程中的电子传递机制研究王敏聊城大学基于次级代谢产物活性和结构的重楼内生菌多样性及与宿主方晓梅中国医学科学院医药生物技术研究所植物相关性研究肠出血性大肠杆菌效应分子NleF通过抑制caspase-4参与其致病宋婷中国人民解放军军事医学科学院的分子机制研究微生物菊粉酶C末端结构域的功能研究刘光磊中国海洋大学谷氨酰胺蓝靛素合成酶的催化机制研究秦华中国科学院微生物研究所槐糖脂的结构修饰及其联合依托泊苷对食管癌细胞增殖马晓静合肥工业大学的影响和机制研究氨基糖苷类抗生素核糖开关翻译调控机制的深入研究张静复旦大学多烯聚酮细胞色素P450的区域及立体特异性研究季俊杰中国科学院过程工程研究所Thienodolin中噻唑并吲哚环的生物合成机制研究马宏敏武汉大学维生素K2合成关键酶的结构解析与基于结构的抑制剂设计徐铮南京工业大学集胞藻PCC6803中sll1981基因编码蛋白的功能研究李志敏中国科学院青岛生物能源与过程研究所沙雷氏菌灵菌红素的前体MBC合成途径中相关蛋白结构解析冉婷婷南京农业大学及反应机制的研究粘细菌新型β-1,3-葡聚糖酶的催化机制研究黄彦南京农业大学恶臭假单胞菌YL19产2种环脂肽的结构鉴定及合成机制研究孙燕陕西师范大学Bacillus subtilis双精氨酸转运系统中信号肽定向识别的分子机制崔文璟江南大学电子歧化转氢酶NfnAB的分子进化及结构与功能关系的研究黄海燕山东省医学科学院极端嗜热细菌Caldicellulosiruptor中双功能纤维素水解酶吕明中国科学院青岛生物能源与过程研究所作用机制的研究里氏木霉内切葡聚糖酶EGI催化纤维素水解的机理研究宋乡飞中国科学院青岛生物能源与过程研究所毕赤酵母转录因子Fhl1p功能鉴定及其促进外源蛋白表达的机理梁书利华南理工大学莱茵衣藻膜蛋白Cr-FAX在脂肪酸跨叶绿体膜运输过程中的作用研究李楠楠西南大学镉胁迫下MAPKs对通道蛋白的调控机理张丽琳天津大学构巢曲霉Calcineurin和CchA参与菌丝极性生长的分子调控机制王莎湖州师范学院blaOKP β-内酰胺酶耐药基因进化研究邹立扣四川农业大学多基因协同调控里氏木霉高效分泌表达蛋白的研究苏小运中国农业科学院饲料研究所潘多拉菌中氯苯代谢的两个基因簇的转录调控研究晁红军中国科学院武汉病毒研究所ClpX ATPase在蜡样芽孢杆菌生物防治小麦土传病害中的作用张颖河南大学酿酒酵母耐受玉米秸秆水解液抑制物基因的功能鉴定刁刘洋中国科学院上海生命科学研究院病原细菌受体激酶PcrK特异识别寄主植物激素分子信号的机制王芳芳中国科学院微生物研究所深海链霉菌SCSIO ZJ46来源的抗菌环肽desotamides的生物合成研究李青连中国科学院南海海洋研究所稻瘟菌可长距离移动的效应蛋白MoSDT1在与水稻互作中杨静云南农业大学的分子鉴定寒地根瘤菌III型效应因子对大豆宿主特异性的影响辛大伟东北农业大学1344生命科学第26卷RAS-2调控粗糙脉孢菌生物钟的分子机制研究胡启文中国人民解放军第三军医大学固氮施氏假单胞菌非编码RNA crcZ和crcY在碳代谢抑制中战嵛华中国农业科学院生物技术研究所的协同作用机制利用CRISPR/Cas系统建立新型乳酸菌基因组编辑技术郭婷婷山东大学纳豆激酶高效分泌表达系统的构建与最适pH偏酸性改造刘中美江南大学普鲁兰酶的理性设计及其在枯草芽孢杆菌中的分泌表达研究谢能中广西科学院建立土曲霉高效基因打靶平台的研究黄雪年中国科学院青岛生物能源与过程研究所大肠杆菌人工合成功能寡糖2-岩藻化乳糖的系统研究黄笛南开大学组合调控解淀粉芽孢杆菌胞内辅因子NADH/NAD+强化2,3-丁二醇杨套伟江南大学生物合成的分子机制磷霉素生物合成中Fom3催化的sp3碳原子甲基化机制研究陈允亮中国科学院上海生命科学研究院肠杆菌中以葡萄糖为原料利用苯丙酮酸途径合成苯乙醇张海波中国科学院青岛生物能源与过程研究所的代谢调控研究基于微流控液滴的放线菌高通量筛选培养和分离何湘伟北京林业大学基于双底物平行标记实验的13C代谢通量分析的新方法姚瑞莲上海交通大学微流控芯片上微生物培养的溶氧梯度控制方法与基因表达研究甘明哲中国科学院苏州纳米技术与纳米仿生研究所基于单细胞拉曼分选的新型酵母突变株筛选方法研究王婷婷中国科学院青岛生物能源与过程研究所单细胞拉曼技术在微生物群落动态监测中的方法学研究黄适中国科学院青岛生物能源与过程研究所新型宿主免疫调节抗菌化合物的作用机制研究崔金辉中国科学院微生物研究所根际微生物在壶瓶碎米荠超积累硒过程中的作用及其机制袁林喜中国科学技术大学Aquamicrobium sp. hun6降解皮考啉酸及其衍生物的机理研究吴志国天津科技大学喜温嗜酸硫杆菌中sigma因子对硫代谢和环境压力适应性的调控陈林旭山东大学铜绿假单胞菌胞外多糖和鼠李糖脂交互调控的分子机理王世伟中国科学院微生物研究所一株鞘脂菌多环芳烃降解代谢网络的转录组学研究丁国春中国农业大学天然来源腐殖质作为电子媒介体促进多氯联苯生物降解的研究章春芳浙江大学双启动子级联调控荧光蛋白基因表达载体的构建及环境监测应用李琴中国环境科学研究院Sphingobium sp.MEA3-1降解烷基取代苯胺类化合物2-甲基-6-乙基侯颖河南科技大学苯胺分子机理研究芽孢杆菌B1菌株降解4-羟基苯甲酸途径中NIH重排反应研究冯昭中江苏师范大学磷酸激酶slt2调控黄曲霉毒素产生的分子机制张峰福建农林大学中国桦木属植物外生菌根真菌多样性及分布格局研究王琴辽宁省林业科学研究院海洋枯草芽孢杆菌C01抗菌物质合成调控的信号通路研究穆大帅山东大学沉水植物与磷细菌的联合对水-沉积物间磷循环的作用规律研究李海峰河南工业大学生物阴极加速氯代烃污染物还原脱氯及作用机制李智灵哈尔滨工业大学菌株Pigmentiphaga sp.H8对3,5-二溴-4-羟基苯甲酸的降解陈凯南京农业大学及脱溴机制大兴安岭地区火山口湖好氧不产氧光合菌群落多样性及系统分类李爱华中国科学院微生物研究所海洋弧菌菌群感应信号分子N-acyl homoserine lactones对NK 细胞付凯飞中国人民解放军海军总医院的调控作用研究基于噬菌体随机肽库研制新型酿酒酵母表面吸附剂张海燕河南大肠道微生物组在转基因鲤糖代谢中的作用及调控机制研究颜庆云中国科学院水生生物研究所特殊生境中地衣内生菌次生代谢产物及其抗辐射作用研究元超中国医学科学院药用植物研究所基于“质粒宏基因组”研究饲料中铜、锌、砷对猪粪细菌耐药邓祖军广东药学院质粒的选择与转移微生物协同利用石油烃作用的机理研究胡冰北京大学氧调控奥奈达希瓦氏菌降解偶氮染料的机理研究杨玉义中国科学院武汉植物园假诺卡氏菌CB1190降解四氢呋喃的代谢机理及关键酶研究方倜中国科学院武汉病毒研究所Cupriavidus basilensis B-8对木质素降解机制的研究石岩河南师范大学第12期1345国家自然科学基金委员会生命科学部2014年度资助项目嗜酸微生物协同作用浸出黄铁矿的分子机制韩一凡中国科学院天津工业生物技术研究所c-di-GMP受体Filp与PXO_02715蛋白互作及其对水稻白叶枯杨凤环中国农业科学院植物保护研究所病菌毒性的共调控作用结核分枝杆菌LrpA蛋白调控基因转录的机制研究宋宁宁中国农业科学院哈尔滨兽医研究所控制致病性大肠杆菌DegP表达的调控蛋白及其调控机制研究李兆利中国农业科学院哈尔滨兽医研究所水貂铜绿假单胞菌强弱毒菌株差异表达基因的鉴定及其与毒力齐静山东省农业科学院的相关性研究一株糖霉菌属放线菌抑菌活性成分及作用机理研究张越锋塔里木鱼类病原菌迟钝爱德华氏菌功能RNA组研究杨敏俊上海人类基因组研究中心福氏志贺菌2a新毒力蛋白Pic细胞毒性的分子机理研究张俊琪复旦大学巴尔通体效应蛋白BepC与宿主p53蛋白互作诱导细胞凋亡机制研究袁聪俐上海交通大学malS-5'UTR调节伤寒沙门菌耐酸力和侵袭力的作用机制研究张盈江苏大学噬菌体裂解酶细胞壁结合域抑制金黄色葡萄球菌生物膜形成杨航中国科学院武汉病毒研究所的机理与应用结核分枝杆菌耐药相关非编码基因区的系统发现和耐药新机制研究张泓泰中国科学院生物物理研究所血红素氧化酶独特的ND9/14位点促进空肠弯曲菌感染定植张睿中国人民解放军第三军医大学及机制研究头束霉及其近似属的分子系统学分析耿月华河南农业大学水稻纹枯病菌诱导水稻程序化死亡的作用机制郑爱萍四川农业大学SreE介导的铁调节对皮炎外瓶霉形态发生、药物敏感性高露娟复旦大学及致病性的影响钙信号系统介导白念珠菌形态发生分子机制的研究喻其林南开大学隐球菌转录因子Frt1功能和调控机理研究刘同宝山东大学烟曲霉Rho1蛋白对其细胞壁生物学特性、致病力及诱发宿主田曙光中国人民解放军军事医学科学院天然免疫应答的调控机制研究白念珠菌开关蛋白S?1和S?2在菌丝发育和致病过程中的调控戴宝娣中国科学院上海生命科学研究院机制研究昆虫RNAi抗病毒免疫途径调控南方水稻黑条矮缩病毒与不同贾东升福建农林大学介体间存在亲和性差异的机制研究细胞自噬在伪狂犬病毒复制感染中的作用孙明霞中国农业科学院哈尔滨兽医研究所口蹄疫病毒基因组3'非编码区通过DDX21诱导I型干扰素产生杨德成中国农业科学院哈尔滨兽医研究所的分子机制GCRV内衣壳的自组装及与细胞相互作用机制研究张付贤中国科学院武汉病毒研究所杆状病毒核心基因gp41在BV和ODV形成过程中的作用机制研究沈姝中国科学院武汉病毒研究所质型多角体病毒衣壳蛋白VP5参与病毒RNA复制机制的研究杨洁武汉大学杆状病毒ODV表面刺突的组分鉴定和功能研究侯典海中国科学院武汉病毒研究所蝙蝠正呼肠孤病毒对实验动物的致病性研究杨兴娄中国科学院武汉病毒研究所应激颗粒对新城疫病毒复制与先天性免疫的调控机制研究孙英杰中国农业科学院上海兽医研究所日本乙型脑炎病毒逃逸神经系统CD8+ T细胞免疫清除的机制刘珂中国农业科学院上海兽医研究所猪瘟兔化弱毒疫苗株适应家兔的关键基因定位李永锋中国农业科学院哈尔滨兽医研究所基于HIV-1病毒感染必需因子Vif和其相互作用宿主蛋白的高通量周小红吉林大学小分子药物筛选体系Hedgehog信号通路在人呼吸道合胞病毒感染过程中作用机制研究邹罡中国科学院上海巴斯德研究所纤毛杆影响嵌合型腺病毒感染T淋巴细胞效率的机制研究张文峰广东药学院I型单纯疱疹病毒UL2蛋白核质转运信号的鉴定及其在病毒蔡铭升广州医科大学感染中功能的研究流感NS1蛋白通过改变细胞骨架帮助病毒释放姜威中国科学院微生物研究所内质网蛋白SEC62调控登革病毒感染的新机制研究柳恒中国科学院上海生命科学研究院1346生命科学第26卷PI3K/Akt信号通路在DHEA衍生物抗EV71中的作用研究魏艳红湖北工业大学中东呼吸综合征冠状病毒刺突蛋白N-端结构域及S1亚基的结构逯光文中国科学院微生物研究所与功能研究麻疹病毒N蛋白诱导细胞自噬及其分子机理研究刘鑫武汉大学人腺病毒E1B55K与E4orf6蛋白相互作用的型特异性研究及应用邹小辉中国疾病预防控制中心病毒病预防控制所乙型脑炎病毒入侵环节中包膜糖蛋白关键氨基酸残基和肽段刘海滨中国科学院武汉病毒研究所的识别与功能分析受MicroRNA调控的重组EV71在恶性神经胶质瘤模型上的溶瘤研究张晓玮中国科学院武汉病毒研究所肠道病毒71型(EV71)诱导线粒体自噬的分子机制研究王蓓中国医学科学院病原生物学研究所来自丙型肝炎亚基因组复制子的外体活化炎症小体的机制研究徐咏芬中国科学院上海巴斯德研究所霍乱弧菌分型噬菌体VP2吸附和注入分子机制的研究徐嘉良北京工商大学同一原噬菌体在两株希瓦氏菌中的作用及H-NS对其切离调控的研究刘晓晓中国科学院南海海洋研究所ORF48催化铜绿假单胞菌噬菌体PaP1甲基化的作用与机制卢曙光中国人民解放军第三军医大学猪肺炎支原体果糖二磷酸醛缩酶的致病机制研究华利忠江苏省农业科学院靶向单基因模型构建与新型小分子化合物CB抗衣原体分子包小峰南通大学作用机制研究2植物学龙胆属植物中雌雄异位和雌雄异熟的功能分异和进化式样研究李肖夏中国林业科学研究院百合属的花进化和传粉生态学刘长秋中国科学院昆明植物研究所拟南芥RSU3调控花粉管顶端生长的分子机理周利明河北联合大学水稻CRC2蛋白在减数分裂联会复合体形成中的分子机理研究纪剑辉淮阴师范学院玉米胚胎发生相关基因EMB4的功能研究李翠玲山东大学双靶向半胱氨酸蛋白酶抑制剂NtCYS调控胚柄细胞程序性死亡赵鹏武汉大学的分子机理研究野生大豆/栽培大豆胚胎发生的比较研究及其重要经济性状刘媛中国科学院东北地理与农业生态研究所的分子调控机制探讨暖温带不同功能型植物的水分利用策略及对降水变化的响应杜宁山东大学TBL在植物细胞壁多糖乙酰化修饰中的功能研究刘香玲中国科学院遗传与发育生物学研究所一个新的水稻每穗粒数主效QTL Gn2的图位克隆和功能分析朱金燕江苏省农业科学院拟南芥MUP24.5基因维持种子黏液质结构的功能研究于丽中国科学院青岛生物能源与过程研究所拟南芥AtFH14与微丝及微管骨架的相互作用机制王姣姣北京师范大学中国紫珠属(唇形科)的分类修订马仲辉广西大学中国千金藤属(防己科)的分类学研究张紫刚南京农业大学虾脊兰属及其近缘类群(兰科)的系统分类学研究翟俊文福建农林大学广义九里香属的分类修订及系统学研究牟凤娟西南林业大学中国山矾属(山矾科)的分类学修订刘博中央民族大学中国蛛毛苣苔属的分类学研究许为斌广西壮族自治区中国科学院广西植物研究所爵床科恋岩花属的系统位置及其物种分化研究高春明滨州学院中国淡水隐藻类的分类学研究夏爽中南民族大学中国海洋绿藻门刚毛藻目的分子系统发育学及其DNA条形码库构建黄冰心汕头大学木腐菌对粗木质残体附生苔藓植物多样性的影响及其机制闫晓丽中国科学院成都生物研究所木灵藓属(Orthotrichum hedw.)的形态演化、系统发育和分类王庆华中国科学院植物研究所拟蕨藓属的分类修订及其与近缘属的关系于宁宁中国科学院植物研究所中国细鳞苔属(Lejeunea)植物的分类修订韦玉梅广西壮族自治区中国科学院广西植物研究所耐旱复苏植物旋蒴苣苔的分子谱系地理学研究李晶首都师范大学小叶栒子复合体谱系地理及物种界定研究李飞飞中央民族大学卫矛科南蛇藤属(Celastrus L.)分子系统学与生物地理学研究沐先运北京林业大学国家自然科学基金委员会生命科学部2014年度资助项目第12期1347基于RAD-Tag技术:特提斯孑遗洲际间断分布植物穗菝葜祁哲晨浙江理工大学(菝葜科)的亲缘地理学研究伞形科山芹属的系统发育与生物地理学研究廖晨阳四川大学世界凤尾蕨属及凤尾蕨亚科的系统学研究张良中国科学院成都生物研究所中国—喜马拉雅特有属蓝钟花属的系统进化与生物地理研究周卓中国科学院昆明植物研究所花柱二型性水生植物莕菜的适应性遗传进化研究岳晓丽湖北工业大学番荔枝科独子木属的分子系统发育研究:探索高分化速率薛彬娥中国科学院华南植物园与性状演化及生物地理的相关性东亚特有药用植物玄参复合种系统发育和物种形成机制研究陈川杭州植物园植物RPW8-NBS-LRR类抗病信号传导基因的起源、演化邵珠卿南京大学及功能化机制水稻亚种间新合成四倍体早期世代基因组变异徐春明东北师范大学特异响应冷胁迫的DREB1/CBF基因亚家族在陆生植物中的演化康菊清陕西师范大学黑种草属植物中AP3-3基因表达量的调控进化及其对花瓣形态张睿中国科学院植物研究所多样性的贡献栽培大豆茎杆直立驯化性状的分子机制研究董阳中国科学院植物研究所南果梨花发育分子机制研究张吉斯鞍山师范学院蕨类植物叶绿体RNA编辑及其适应性进化研究高磊中国科学院。
古菌的化学生物学特性及其在环境修复中的应用研究

古菌的化学生物学特性及其在环境修复中的应用研究古菌(Archaea)是一类独特的微生物,其细胞结构、生理过程和基因组组成都与细菌和真核生物有很大的差异。
近年来,随着对古菌研究的不断深入,人们发现古菌在环境修复、产生有用化学物质等方面有着潜在的应用价值。
本文将介绍古菌的化学生物学特性以及其在环境修复中的应用研究。
一、古菌的化学生物学特性1.基于RNA的分类目前,所有生物被分为三个域:细菌(Bacteria)、真核生物(Eukarya)和古菌(Archaea)。
古菌的分类主要基于RNA序列的相似性,这是比较古菌与其他生物之间亲缘关系的一种最为有效的方法。
2.细胞结构古菌细胞壁不含有真菌和细菌的主要成分——封闭多糖,而是由一种特殊的合成多糖和一层糖蛋白组成的膜状结构。
此外,古菌细胞膜的脂质组成也与细菌和真核生物有所不同。
3.代谢途径古菌具有多样的代谢途径,其中包括光合作用、化能和化合物作为能源的氧化、还原和甲烷作为能源的化学反应。
古菌还能够在极端条件下生存,如高温、高盐度、高压和强酸碱性环境。
4.基因组组成古菌的基因组结构比较简单,通常只有数百万个碱基对,比真核生物要小得多。
古菌的基因组大小和复杂度与生活方式和栖息环境密切相关。
二、古菌在环境修复中的应用1.重金属污染修复古菌能够在高温、高压和强酸碱性等极端环境下生长,因此对于重金属污染的修复具有一定的应用前景。
研究人员发现,古菌能够吸附、还原和沉淀重金属离子,如汞、铅、铬等。
同时,古菌还能够产生多个对重金属具有降解和还原性质的酶类物质,如硫酸盐还原酶和底物级磷酸酰化酶等。
2.有机污染修复除了重金属污染,有机污染也是环境修复中的重要问题。
古菌在生产有机物的过程中还能够产生许多酶,这些酶可以分解和利用有机污染物。
古菌能够利用多种有机废料为基础生产产物,其中含有大量的有机污染物,如苯、酚、酮等。
古菌利用这些有机物产生氨基酸、聚酯、纤维素和其他有用化学物质,从而实现了有机废物资源化的环保效益。
热泉微生物化石的识别研究及其科学意义

热泉微生物化石的识别研究及其科学意义微体古生物2005年6月;22(2):136—142 ActaMicropalaeontologicaSinica22(2):136—142;June,2005热泉微生物化石的识别研究及其科学意义*冯军李江海牛向龙(北京大学地球与空间科学学院,教育部造山带与地壳演化重点实验室,北京100871)提要现代海底热液喷I=I以及许多陆地热泉周围生活着密集的生物群落.热液生态系统的初级生产者嗜热细菌和古细菌(Archaea),其初级能量来源是由地球深部上升喷出流体提供的化学能.围绕现代热泉微生物及其与地史时期热泉微生物化石的对比研究表明,它们具有相似结构特征.研究微生物成矿机制和微生物化石化作用,以及沉积物中由生物化学作用产生的生物标志,不仅有助于探讨海底热液活动的规律性和成矿机制,也可以为鉴别古老岩石和地外矿物中生命现象提供更多更详细的鉴定标志,对于理解生命起源和地外生命都有重要的理论意义.关键词微生物化石热泉热液块状硫化物鉴别标志1前言现代海底热液喷口以及许多陆地热泉周围生活着密集的生物群落.特别是海底黑烟囱周围发现极端环境下的生命活动及其独特的生命体系,并以此提出生命起源于海底黑烟囱周围的假说(Corliss,1990).尽管在现代深海热液喷口以及陆地热泉发现了生机勃勃的热液生物群落,但是热液生物的化石却不多,而且大部分是显生宙的,前寒武纪的热液生物化石特别是微生物化石很少(Littleeta1., 1998).微生物化石的鉴别非常困难并且存在很多争议.本文重点探讨热泉微生物化石的标志,形成过程及其研究意义等,以期为鉴别古老岩石和地外矿物中生命现象提供更多更详细的鉴定标志.2热泉微生物化石记录自二十世纪70年代末在太平洋发现第一个海底黑烟囱以后,对现代以及古代海底黑烟囱的研究已经取得了很大的进展,20多年里世界各地对不同时代的块状硫化物(被认为海底热液成因)进行了广泛的调查研究,先后在澳大利亚,俄罗斯,爱尔兰,塞浦路斯,阿曼等地找到了古海底黑烟囱的残片,以及相关的块状硫化物.但是,时代大于6亿年的尚不很多,迄今为止发现时代最老的海底黑烟囱是在我国华北中元古代硫化物矿床中发现的硫化物黑烟囱(李江海等,2003),这也是世界范围内首次发现的前寒武纪完整的黑烟囱构造.目前围绕该黑烟囱的野外调查和实验测试以及相关的微生物化石研究工作正在进行中.尽管很多现代海底热液系统发现了生机勃勃的热液群落(DemingBaross,1993;Nisbet,2000),但是热液生物化石却不多,仅仅在俄罗斯乌拉尔南部志留纪块状硫化物矿床(Herringtoneta1., 1997),塞浦路斯的晚白垩世特罗多斯蛇绿岩的硫化物中(Littleeta1.,1999),加利福尼亚侏罗纪块状硫化物(Littleeta1.,1999),爱尔兰晚石炭纪黄铁矿矿床(Russell,1996)等二十余处地点发现管状蠕虫类,腹足类,腕足类和双壳类等类似现代海底黑烟囱周围生物的化石(Littleeta1.,1999),证实地史时期黑烟囱对生命活动具有重要影响,引起科学界的强烈反响.块状硫化物中找到的最古老的细菌化石是在澳大利亚Pilbara克拉通火山块状硫化物(3235Ma)中发现的黄铁矿化的丝状体,可能是丝状微生*基金项目国家自然科学基金项目"河北兴隆中元古代硫化物黑烟囱形成机理及其意义"(编号:40472097)和北京大学985项目资助.收稿日期2004—12—29}5军等:热敏懈生物化,』的i{{别删.究驶J科学意义137IVMS'II的'j体娃儆f片c弓l由lt1mlT,2l¨1I.IPEul¨1lroRraphs【)ff】hlP*t-n{finn1IheSniphl=ru'hIVM,]epnsil(froltl1llH?I.!fill(})物的化i<]'{asrflusset【-J)f))采光学显微镜和于1捕r乜镜研究表叫+这蝗状体呈蝗状.没有分枝.粗耋l1{均匀(径c】.52m1.I=乇洲luI11)有自'的,弯曲【l}】和I抽等形态.在同方向上柏一缠绕.密集分.数量{II!多.jnl丌]1¨Hlnl的区域巾(厚:{『I,㈨】扭过:螂1个反射光下,丝状体似乎总是由微米级的黄铁矿『体组成这状体.破认为足原牛物的遗迹.足,I活在太古代洋底的嗜热细苗c!仃j是化学自养的,段有光台作J/j,牛活住ll:【'或者接近flc的环境中i果上述解释成.么它们将r泛地分布在,时的深海火II【系统.j活海底热液系统(海底黑烟嘲)周(Nisbet.2E100).庄永一化太平津热破堆物巾封"常被硅质重I5rh氧氧化物以]亚少Mn的氧氧化物矿化十I描电镜研究走叫大部分的矿化堆织物中坦状体细闰有硅和h-氢氧化物包壳或抒被它们拯~列菌避状体有弯I】,赢的,独n或告舒枝的..Ⅱ形成多孔站佝矿物包壳螭细的丝状体小于I,tztn,有丝状体为表而有硅厩加厚可以超过2Iznt厚许多n乎滑们和ⅡI片状表丽的球状体附着而呈绳状这状体细菌都被硅lf空结(Hein"f.,j1jJ这些丝状体的形惫与Chile北部和新四兰的陆地热泉巾的照状怍中H同.与其它海底热泉的蝗状体也似刘太平洋Expl{)rer南部热液喷【=I研究发现.儿于所f『的矿化细菌和细胞外聚台体都被不太则的富硅的铁氧化物(1JT能是水铁矿),锰氧化物,铁硅酸盐(可能足绿脱石)包裹E们很好的犄状(2201ln1),针状(2l,lO0mn),片状,规则球状(l1锄nttm)和丝状体形式仔氲cllLI【riper&Fouquet.1988)东太平洋洋隆(旷'9.32N)罔2状悱矿化细茼的扫描电镜丑c{片=A)妊们.分杖丝状捧;uJ似菌丝的闸状体(Philoso1)her喷广lJ(引白Juniper&Fo,-t{uet.1洲j:硅化的地状lj奉具打平滑和披状丧面,一避状悼冉硅质球过度生长;I))E 质丝状悼由球状体结形成(引自】teil1el",.【).Fig.2SEMimageilluslralllvarialioninfilamenlmorl~hO[ogy.AIx)ng.branching[ilriTnenl s:H)Hyp]meHikeril~aonlnelwork[Phiios~~pherV en1.ExpklrerRidge)(frifillJ1】】tiper&tr~'{}tl(1Ile11'igs):(,】Silkifiedfilamemwithn1cn【hsurfaceandundu]alingsur[ace;st)mt1filanmntsaovergrownbysillc{.s【】hrL:D】SilicafiLamentsandaggregatescomposedoffusedSl~eruid([romHelnetⅡ,..1999).报道"吹"热液__喷的向色梨斗托物和挫状体.珏遵堆积彤成n厚nq微生物这些喷n勺丝状体1]j能硫氧化的f绑茼(h)…1ih{,..¨i98,谯茭美丽同家公同盟泉.一此微物在|#沉积物一lJ.其一}f还有多秆状细菌.以及一硅藻~一种硫氰化细tfmⅢ,th"")住富硫化物与卒气丽处形成几雁米}乇的灶状体肝离喷I]JLlrli米区域水较低斤水含气体.这早!!譬的始状体}乇满l殳石这些微生物形成颜色鲜艳的馓q一糊.封1]_普遍发生硅化作刚一活在碳酸盐热泉LfJfi:Ji+V}物,l胞外壤合作物质(由活的细胞+liflll胞残余组成的犟质).倒圳生物脱(Labrenz,cf,.20~iuJ物膜经常做+b+ll铣氰化物包裹.从而其能长到保存微生l物体迅速被沉淀物缀盏.形成弛特的矿物结构(Car[1on,.,200(I】.{A.【{细肯帕1描电镜豫(f)t'to[1ll~;热喂j(引力I+lank",..鲫?!J:¨谜状桃怍胞(l也fii7,指1J宽l业J:1)1典物脾的¨描电髓I像c引Jl1.])renz,",..!"lJFig.{A.1{I卜=Mhllag{-{¨let+i+lrfltotyj.,eM(nI.ILF,Ilrin~)(fmmBlank,20(:2);L,)SEMimage.】Ichainnr川一rlllⅧ1")L(wkhhindkiIIedbywhiwa~r1"ows;1j】St:Milnat(elhlypica]art2+tlj__hebiofiln+s(from[.abrer,z.2o00J3微生物化石标志一艘认,}物化百H保存在L#{岩l{】.在奸伦比亚河玄武岩巾世发现了微生物化百(stcv㈨8Mckiul~Y,ij).j千证南非r『1太代巴侗顿绿岩带中枕状熔巾保留有地球早期海底微乍物活曲的证据(FiIrneh,",..2oo1)因此.化石末只存=)二沉积寄rIl1怍为微生物化必须保存有荚结构,确实足L物战冈的:还有,这些样-6必颁还【:J以重复采刮实验表Il』{小旧利一类i:i~TflN的矿化途径不(WestalIef..).自然中以及蛮验筝p得到的细茼被物换后.血构f.,大小,肜态,细胞复杂性,绷1胞壁结构,微生物的聚集等方而与原来的ff=I_机体1f14同(Weslallf【.,..2cj【)】)凶此这些形态上的特点口以为鉴圳微牛物化石提供鉴别标(Wt.s1.all,f.,2c=1㈨|_2lJ).根据许多微生物化石的观测资料和深人研究,[1『以概括{{激,I-物化帕士要特征.井可作为寻找地史时删徵生物化石的自效标.对于探索地外牛命都有露的理论意义与肜惫有咒的土耍特征概括曳【l下(上匝过光学或背高分辨宰显微镜扫描电镜千n透射电镜等):3l大小尺寸现代微生物的尺寸范围r『_H『儿J个微米到≤o.1¨1(Kajander『{)lu.I998:Uwins,"f..】998).几部分现代微't-拗郁很小(直径≤!!xni)(Westall,.,2J】化r化的敞生物纳胞具有原始钏胞的形态和大小所以微生物化亓的大小范围是l101Xli1.直径的不同反映出分类学或2期冯军等:热泉微生物化石的识别研究及其科学意义者保存机制上的不同(HofmannFarmer,2000).3.2形态具有特殊的形态,包括:球状,丝状体(曲线或者螺旋形等),杆状,管状等,以及细胞繁殖的证据:包括细胞的生长,分裂和出芽等.对于迅速矿化的石灰华和硅质热泉体系中的化石化过程研究表明,表面普遍有简单的包壳(成型后有机物快速分解留下的),并被含金属沉淀物聚集加厚,从而使其在成岩作用和重结晶作用期间也能很好的保存(HofmannFarmer,2000).这一过程使得丝状体化石具有管状结构,并且管的直径尺寸和原始的丝状细菌大小类似.在深海热液喷口,丝状细菌包壳为硅质,有机物被硫化物和铁氢氧化物代替(ZierenbergSchiffman,1990;Littleea1., 1999).在沉积物中保存的微生物化石中,相互缠绕,拉长,无规律的丝状体超过球状体以及无定形(Banfieldela1.,2000).丝状体在形态学上优于球菌,可以组成复杂的微生物席,在湍流边界层,丝状体组成的微生物席对其周围的局部微环境(如()一HS梯度)具有重要的调节作用(TaylorWirsen, 1997),并且可以在快速流动的环境中保存下来.丝状体可能是发生在硫化物流体中的特征现象.3.3群落.必须有聚集现象细菌通常是集群生活的,很难分离出个体,常常数以百万计(尽管也有些群落数量较少),因此化石化细菌也应该多呈集群保存,由不同种类微生物形成群落.3.4与细菌产物有关的集合体(生物膜)由活细胞和细胞残余形成的三维网状结构作为生物膜,可以为微生物提供适宜的微环境使其与外界不利环境隔开.很多地方都发现了碳酸盐沉积物中的生物膜,其与细胞以及细胞残余密切相关,是一种独特的物理生物指示标志.生物膜可以是多种成分的,其结构和构造发生矿化后可以保存下来(Carltoneta1.,2000;Westalleta1.,2000).3.5生物侵蚀除了直接的化石证据以外,微生物可以留下证明其存在过的间接证据.比如细菌的钻孔作用而留下的孔洞,孔壁圆滑,直径(1—2m)和形状与细菌一致,有些孔中还可能保留有细菌化石.另外微生物分泌的酶可以形成侵蚀标记(BarkerBan—fiekd,1996).微生物可以促进矿物的成岩作用和分解作用.当微生物生活在岩石或者矿物表面时, 其代谢产物可能导致侵蚀作用;侵蚀作用表现为岩石(或者矿物)的组分(Fe,Mn,硫化物,硫酸盐等)的氧化或者还原.微生物对于矿物组分的侵蚀作用是有选择性的,只有一种或者几种矿物组分可以溶解, 这样未被微生物侵蚀的不溶组分残余将变得富集(Henry,1998;Carltoneta1.,2000).作为微生物化石必须是原生的也就是说它们不是污染的,必须鉴别出微生物化石与非化石和成岩后的矿物,人工污染等的区别.另外,处理样品时必须特别小心,防止现代的污染,以及识别出后期含微生物流体的侵入.微生物化石主要依靠形态进行鉴别和分类,并且依据形态对比推测其生存环境.然而,因为现代的和化石化的细菌,蓝藻,海藻的尺度, 形态无法区分,所以这样的分类非常笼统,并且形态相似,其代谢方式以及生存环境未必相似(Alter—mann,2001)4热液微生物化石化机制在提出如何识别细菌化石之前,需要了解细菌如何形成化石,以及它们在岩石记录中的特征.细菌化石化试验研究证明,细菌的细胞壁还有细胞质和细菌产生的胞外多聚糖物质,它们作为矿物结晶成核的位置,最终细胞被矿物完全置换(有时在矿物中也形成印模的孔洞).细菌化石能够被保存下来, 主要是由于细胞表面被可溶的重金属固化(生物成矿作用),包括离子交互作用或者新陈代谢产物产生的化学反应(westalleta1.,2001).生物从外界摄取离子,沉淀在细胞内,外或者发生化学反应,矿物析出,生长,聚集,残留于细胞的作用被称为生物矿化作用(biomineralization),与微生物矿化作用有关的矿物最少有250种以上(Lowenstam,1981;田崎和江,1995).这些矿化反应大致可分为两种:主动矿化和被动矿化(Southam&Donald,1999): 4.1主动矿化主要是细菌细胞代谢产物反应生成金属成分(Fortineta1.,1994)或者直接被细菌转化(GorbyLovley,1992).细菌活动也可以促使矿物在晶核表面富集.在饱和溶液情况下,细菌充当地球化学反应的固体颗粒,并且可以增加矿物沉淀的速度(Konhauser,1998).4.2被动矿化主要发生在细菌细胞表面,这是因为细菌细胞壁是呈负电荷并包含氨基,可以吸附各种离子和矿物,并且导致各种矿物结晶,如氧化物,硅酸盐,碳酸盐,硫化物等(Fortineta1.,1998;Kon—hauser,1998).在细胞表面存在两种矿化作用:(1)140微体古生物22卷从溶液中带出和捕获颗粒;(2)金属粘结和沉淀在细胞表面以提高金属浓度(Maginneta1.,2002).微生物由于与热液频繁接触,使得金属离子可以进入细胞膜,一旦细胞死亡或者降解,在细胞膜上的金属离子可以作为核促进矿物结晶(Caryeta1.,1998).金属沉淀在细菌表面通常为含水无定形聚合体.但是,它们可以随着时间的推移形成晶核(Beveridge, 1988),晶核较稳定很少发生分解,因为在细胞壁上介于液态与晶核之间的界面张力使得其呈还原性. 细菌细胞矿化的另一个重要特征是其有非常大的表面(体积比)(Beveridge,1988),并且死的细胞相对于活的细胞可以粘结吸附更多的金属离子(Urrutiaeta1.,1992).细胞表面的金属包壳可以防止重新活化,并且可以长期保留下细胞的组分.细胞的形态控制细菌化石的大小和形状,包括:球形,杆状,丝状体,螺旋体和茎状结构等.采用透射电镜对超薄切片研究表明,矿化的细菌细胞具有原始细胞的形态和大小,即使细胞膜和细胞质都发生矿化,细胞膜也可以与细胞质区别开(Southam& Donald,1999)5讨论:研究热液微生物化石的科学意义海底黑烟囱及周围生物群落的发现,极大地改变了人类对海洋中矿产和生物资源的认识,是研究金属成矿过程与极端环境下生命活动的理想实验室.科学家通过深潜器到海底可以实地观察和监测黑烟囱地质活动及其对生物的影响.黑烟囱周围的化能自养古细菌,代表了早期生命的遗孓.海底黑烟囱提供了与早期地球及其生命活动类似的环境和条件,如大量气体,高温环境,深水隐蔽等,为研究生命起源以及演化提供了良好例证(冯军等,2004).为此,在开拓海底黑烟囱研究领域的同时,开展它们与陆上保存块状硫化物的地质对比,将揭示深海黑烟囱成矿过程,早期生命起源与演化的重要线索.热液微生物化石的识别标志包括:大小,形态,细胞繁殖,群落聚集和生物膜等直接标志以及生物侵蚀等间接标志.地史时期热泉周围相关生物化石,是生命起源与演化的重要记录.我国分布不同时代,不同构造环境下形成的块状硫化物,具有研究上述问题得天独厚的优势.研究微生物成矿机制和微生物化石化作用,以及沉积物中由生物化学作用产生的生物标志物,不仅有助于探讨海底热液活动的规律性和成矿机制,也可以为鉴别古老岩石和地外矿物中生命现象提供鉴定标志,对于理解生命起源和地外生命都有重要的理论意义.参考文献冯军,李江海,陈征等,2004.海底黑烟囱与生命起源综述.北京大学(自然科学版),40(2):318325李江海,冯军,牛向龙等,2003.华北中元古代硫化物黑烟囱发现的初步报道.岩石,19(1):167168田崎和江,1995.微生物力<弓铖物.地耍=一灭,489:173O AITERMANNW.2001.TheoldestfossilsofAfricaabriefreap—praisalofreportsfromtheArchean.AfricanEarthSciences,33:427~436BANFIELDJF,WELCHSA,ZHANGHengzhongeta1.,2000. Aggregation——basedCrystalgrowthandmicrostructuredevelop——mentinnaturalironoxyhydroxidebiomineralizationproducts. Science,289:751754BARKERWW,BANFIELDJF,1996.Biologicallyversusinor~ganicallymediatedweatheringreactions:relationshipsbetween mineralsandextracellularmicrobialpolymersinlithobiontiecommunities.ChemicalGeology,132:55~69 BEVERIDGETJ,1988.Thebacterialsurface:generalconsidera~tionstowardsdesignandfunction.Can.J.Microbio1.,34: 363372BLANKCE,CADYSL,PACENR,2002.Microbialcomposition ofnear-boilingsilica-depositingthermalspringsthroughoutYel~lowstoneNationalPark.AppliedandEnvironmentalMicrobio- logy,68(10):51235135CARLT0NCA,AIERTFG,CHAFETZHSn.,2000.Mi~croscopicphysicalbiomarkersincarbonatehotsprings:Implica~tionsinthesearchforlifeonMars.Icarus,147:4967 CARYSC,SHANKT,STEINJ,1998.Wormsbaskinextreme temperatures.Nature,391:545546CORLISSJB,1990.Hotspringsandtheoriginoflife.Nature, 347:624DEMINGJW,BAR0SSJA,1993.Deep-seasmokers:Windowsto asubsurfacebiosphere?Geochim.Cosmochim.Acta,57: 32193229FENGJun,LIJianghai,CHENZhengeta1.,2004.Areviewon "BlackSmokers"anditsimplicationfororiginoflife.ActaSci~entiarumNaturaliumUniversitatisPekinensis,40(2):318325F0RTIND,S()UTHAMG,BEVERIDGETJ,1994.Anexamina~tionofironsulfide,iron-nickelsulfideandnickelsulfideprecipi~rationbyaDesulfotomaculumspecies:anditsnickelresistance mechanisms.FEMSMicrobio1.Eco1.,14:121—132F0RTIND,FERRISFG,SC0TTSD,1998.FormationofFe-sili—caresandFe-oxidesonbacterialsurfacesinsamplescollected nearhydrothermalventsontheSouthernExplorerRidgeinthe northeastPacificOcean.AmericanMineralogist,83:1399—14082期冯军等:热泉微生物化石的识别研究及其科学意义141 FURNESH,BANERJEEN2004.EarlyLiferecorded304:578581R,MUEHLENBACHSKela1., inArcheanPillowLavas.Science,G0RBYYA,L0VLEYDA,1992.Enzymaticuraniumprecipitati—on.Environ.Sci.Techno1.,26:2O5~2O7HEINJR.KOSKIRA,EMBIEYRWela1.,1999.Diffuse-flow hydrothermalfieldinanoceanicfracturezonesetting,North—eastPacific:depositcomposition.ExplorationsinMiningGeo- logy,8:299322HELMUTK.2001.Archaea.Encyclopediaoflifesciences.Nature PublishingGroup.1—11HENRYLE,1998.Geomicrobiology:itssignificanceforgeology. Earth-ScienceReviews,45:45--60HERRINGT0NRJ,LITTLECTS,MASLENNIK0VVV eta1.,1997.Silurianhydrothemnal—ventcommunityfromthe southernUralsRussimNature,385:146148 HOFMANNBA,FARMERJD,2000.Filamentousfabricsinlow- temperaturemineralassemblages:aretheyfossilbiomarkers? Implicationsforthesearchforasubsurfacefossilrecordonthe earlyEarthandMars.PlanetaryandSpaceScience,48:lO77—1086JOHNBC,1990.Hotspringsandtheoriginoflife,Nature,347: 624JUNIPERSK,FOUQUETY,1988.Filamentousiron-silicadepo- sitsfrommodernandancienthydrothermalsites.CanadianMi—neralogist,26:859—869KAJANDEREO,CIFTCI()GLUN.1998.Nanobacteria:Analter—nativemechanismforpathogenicintra-andextracellularcalcifi—A,95: 82748279K0NHAUSERK0,1998.Diversityofbacterialironmineraliza—tion.Earth-ScienceReviews,43:91—121LABRENZM,DRUSCHELGK,TAMARATEnZ.,2000. FormationofSphalerite(ZnS)DepositsinNaturalBiofilmsof Sulfate-ReducingBacteria.Science,290:1744—1747 LIJianghai,FENGJun,NIUXianglongela1.,2003.Theprelimi—naryreportonthediscoveryofblacksmokerchimneywithinthe MesoproterozoicsulphidedepositofNorthChina.ActaPetro—logicaSinica,19(1):167l68UTTLECTS,HERRINGT0NRJ,MASLENNIK0VVV以口£., 1998.Thefossilrecordofhydrothermalventcommunities. GeologicalSociety,148:259—27OLITTLECTS,HERRINGT0NRJ,HAYM0NRMnZ.,1999.EarlyJurassichydrothermalventcommunityfromthe FranciscanComplex,SanRafaelMountains,California.Geolo—gY,27:167—17OLITTLECTS,CANNJR,HERRINGT0NRJ.,1999. LateCretaceoushydrothermalventcommunitiesfromtheTroo—dosophiolite,Cyprus.Geology,27:1027--1030L0WENSTAMHA,1981.Mineralsformedbyorganisms.Na ture,211:1126一n31MAGINNEJ,LITTLECTS,HERRINGTONRJela1.,2002. Sulphidemineralisationinthedeepseahydrothermalventpoly—chaete,Alvinellapompejana:implicationsforfossilpreserva—tion.MarineGeology,181:337356NISBETE,2000.TherealmsofArchaeanlife.Nature,405:625—626PRIEURD,1997.Microbiologyofdeep-seahydrothermalvents. MarineBiotechnology,15:242—244RASMUSSENB,2000.Filamentousmicrofossilsina3235million- year-oldvolcanogenicmassivesulphidedeposit.Nature,405: 676—679R0NAPA,ScoTrSD,1993.Aspecialissueonsea-floorhydro thermalmineralization:newperspectives,preface.Economic Geology,88:1935--1975RUSSELLMJ.1996.Thegenerationathotspringofsedimentary oredepositsmicrobialitesandlife.OreGeologyReviews,1O: 199214SC0rrS,2002.Mineralsonland,mineralsinthesea.Geotimes, 47:18S0UTHAMG,D0NALDR,1999.Astructuralcomparisonof bacteria1microfossilsvs'nanobacteria'andnanofossils.Eart}卜ScienceReviews,48:251—264STEVENST().MCKINLEYJP,1995.Lithotrophicmicrobial ecosystemsindeepbassaltaquifers.Science,270:450--453 TAYIRCD.WIRSENC0,1997.MicrobiologyandEcologyof FilamentousSulfurFormation.Science,277:1483—1485 URRUTIAM,KEMIPERM,DoYLER以.,1992.Themem—brane-inducedprotonmotiveforceinfluencesthemetalbinding abilityofBacillussubtiliscellwalls.App1.Environ.Mierobi—o1.,58:3837—3844UWINSPJR,WEBBRI,TAYL0RAP,1998.NovelnanO-Or—ganismsfromAustraliansandstones.AnLMineral,83:15411550WESTAILF,B0NIL,GUETZ0NIME,1995.Thesilicificationofmicroorganisms.Palaeontology,38:495—528WESTALLF,BRACKA,H0FMANNBela1.,2000.AnESA studyforthesearchforlifeonMars.PlanetaryandSpaceSCi—ence,48:181—2O2WESTALLF,WITMJ,DANNJ以a1.,2001.EarlyArcheanfos—silbacteriaandbiofilmsinhydrothermallyinfluencedsediments fromtheBarbertongreenstonebelt,SouthAfrica.PrecambrianResearch,lO6:93一l16ZIERENBERGRA,SCHIFFMANP,1990.Microbialcontrolof silvermineralizationataseafloorhydrothermalsiteonthenorthernGordaRidge.Nature,348:155—157l42微体古生物22卷THEIDENTIFICATIoNoFTHEMICROBEI1OSSILANDITSSCIENTIFICIMPLICA T IoNFENGJun,LIJiang—haiandNIUXiang-long (SchooloJEarthandSpaceSciences,PekingUniversity,MOEKeyLaboratoryofOrogenic BeltsandCrustalEwdution.Beijing100871) AbstractHydrothermalcommunitiesonthedeepseafloorlivearoundblacksmokersites.The primary producersofhydrothermalecosystemsarethermophilesandarchaea.Bacteriaconvertchem icals(fromthesulfur—richfluidspewedoutofvents)toenergy,inaprocesscalledchemosynthesis.Thecomparison inmodernhotspringmicrobeandancienthydrothermalsystemsindicatesthattheyhavesimil arstructuralcharacteristics.Studiesinmechanismofmineralization,theprocessofbacterialfossilization andthebio—chemicalmicrobialbiosignaturesinsedimentnotonlyhascrucialimplicationforthefurtherp robeintotherulesoftheactivitiesofhydrothermalfluidindeepseaandminerogenicmechanismbutalsoho ldimpor—tanceforexplorationofsignaturesoflifeol"prebioticchemistryonotherbodiesinthesolarsys tem,as wellasprovidingmorerobustcriteriaforrecognizingbiogenicfeaturesinancientterrestriala ndextrater—restrialmaterials.Inaddition,ithasimportanttheoreticalmeaningfortheunderstandingofast robiologyandtheoriginoflife.Keywordsmicrobefossils,hotspring,hydrothermalfluid,massivesulfide,identification。
安徽大学2016年大学生科研训练计划项目师生互选情况一览表

资助经费 (万元) 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0 2.0
KYXL2016020 物理与材料科学学院 高效远程量子态制备方案的设计 KYXL2016021 物理与材料科学学院 氢氧化钴基纳米结构在外加磁场驱动下的物相转变机理研究 KYXL2016022 物理与材料科学学院 稀土铁氧体单晶磁热效应的研究 KYXL2016023 化学化工学院 KYXL2016024 化学化工学院 KYXL2016025 化学化工学院 KYXL2016026 化学化工学院 KYXL2016027 化学化工学院 KYXL2016028 化学化工学院 KYXL2016029 化学化工学院 KYXL2016030 化学化工学院 KYXL2016031 化学化工学院 KYXL2016032 化学化工学院 KYXL2016033 化学化工学院 KYXL2016034 生命科学学院 KYXL2016035 生命科学学院 KYXL2016036 生命科学学院 KYXL2016037 生命科学学院 KYXL2016038 生命科学学院 KYXL2016039 生命科学学院 氮、铁和钴多元素掺杂多孔碳微球材料制备与电催化性能研究
王章银 讲师 吴明在 教授 李秋菊 讲师 陈平 副教授 副教授
许非凡 刘瑶瑶 奚坚超,黄京城 刘闯,薛震泳 孙帅,钱鹏飞 纪迎港,刘小虎 洪峰,印月如 钟卓浩,韦丹蕾 李鋆,尹博皓 谢兴丽,陈贵宝 何文军,何仕辉 孙奇轩,陆影 时旭,谢小雨 李天竹,谢志远 刘壮 程源江,周舒利 李竹君 刘天妍,程毓 吴思妤,邹思唯
KYXL2016017 物理与材料科学学院 强磁性纳米粒子表面处理与功能化 KYXL2016018 物理与材料科学学院 基于量子级联激光器的双光谱气体检测技术研究
2009年度南京师范大学国家自然科学基金资助项目清单

2009年度南京师范大学国家自然科学基金资助项目清单学院姓名申请项目名称资助类别金额(万元)数科院(6项)纪春岗L-函数,二次域以及其它数论问题面上项目27魏加群有限维猜测及其相关问题与理论研究面上项目24陈二才动力系统中的拓扑压与维数理论面上项目26曹海涛二维光正交码及相关设计的研究面上项目25王丽大型稀疏非对称线性方程组的预处理及高效算法研究面上项目26赫海龙辛几何中的开“格罗莫夫-威腾”不变量青年基金16物科院(4项)童培庆准周期与非周期系统的若干动力学和相关问题的研究面上项目34马青玉基于声偶极振动矢量的磁感应磁声衍射层析成像技术研究面上项目36肖振军B介子物理高阶QCD修正计算和唯象研究面上项目36张宁异型(复合)铁磁∕铁电复合材料中的电致变磁导效应与微振发电面上项目42化科院(7项)黄晓华稀土作用于辣根细胞膜的若干生物无机化学行为研究面上项目35孙培培高活性纳米金属的制备及其催化有机反应的研究面上项目35魏少华酞菁/二氧化硅靶向给药体系的构建及其光动力活性研究面上项目35杨维本不同结构吸附树脂对典型抗生素类有机物的吸附行为与机理面上项目36韩敏金属纳米晶的可控合成、组装及多功能异质纳米结构的构筑青年基金18李卉卉环境中有机污染物分子与人类肿瘤相关DNA作用机制的研究青年基金19张卉纳米界面上蛋白质构象及其电化学性能青年基金20生科院(11项)陈双林黏菌代表种的DNA标识区段及遗传多样性的研究面上项目25戴亦军不同共代谢基质控制嗜麦芽寡养单胞菌的吡虫啉代谢途径的机理研究面上项目35邵蔚蓝一株嗜冷菌及镶嵌其细胞壁的大型颗粒的研究面上项目35蒋国芳黄脊雷篦蝗的种群遗传结构和系统地理格局研究面上项目31马飞脊椎动物miRNA基因的起源与进化分析面上项目32杨州微囊藻表型转化的野外原位研究面上项目32戴传超广谱植物内生菌优化花生连作土壤微生物区系的研究面上项目30陈龙镉激活神经细胞mTOR通路诱导凋亡及雷帕霉素靶向调控抗凋亡分子机理面上项目31陈华群内皮细胞肌球蛋白轻链激酶调节小鼠肾脏发育及其机制面上项目31裴建军嗜热厌氧乙醇杆菌中还原力感应蛋白介导乙醇代谢调控的分子机理青年基金20陈玉清抗菌肽CM4对癌细胞与正常细胞的选择性与机理青年基金20地科院(17项)汤国安基于DEM的黄土高原地貌形态空间格局研究重点项目150汪涛基于知识网络视角的高科技园区产业空间集聚研究——以苏南为例面上项目30黄震方基于文化视角的区域旅游发展机制与模式研究——以长江三角洲地区为例面上项目30方斌我国东部地区耕地利用的综合优化模式研究——以浙江省浦江县为例面上项目30李云梅面向湖泊水色遥感的多源数据融合与生成研究面上项目35刘学军融合地形结构的DEM质量分析研究面上项目35张雪英面向GIS的文本空间关系解析机制研究面上项目35赵媛中国石油资源流动空间格局演化规律与形成机制研究面上项目36吴江滢辽宁暖和洞年纹层状石笋全新世东亚季风气候记录与驱动机制研究面上项目49张茂恒中更新世以来大九湖盆地泥炭沉积的气候地层学研究面上项目50刘金娥互花米草对苏北盐沼沉积物有机碳库功能格局影响机理面上项目49齐相贞不同繁殖方式的外来入侵植物的扩散机制研究青年基金18周年兴旅游地景观格局演变机制及优化研究:以武陵源为例青年基金18刘会玉植物入侵对不同尺度栖息地时空变异的响应及其预测青年基金20杨昕基于DEM的黄土高原流域边界剖面谱研究青年基金18吴明光基于庞加莱对偶的三维自由拓扑模型青年基金18张卡基于非下采样Contourlet变换的多源影像自动匹配方法研究青年基金18电自院(1项)田恩刚网络控制系统的随机建模、分析与综合面上项目19动院(2项)杨宏旻烟气中单质汞的等离子放电协同光催化氧化特性研究面上项目30王延华生态土壤污水处理系统温室气体的量化和产生机制研究—太湖流域为例面上项目20请项目主持人按照国家自然科学基金项目及经费管理办法,填报项目计划书。
金属离子对林可霉素生物合成的影响

金属离子对林可霉素生物合成的影响
汤海云;郭元昕;李啸;储炬
【期刊名称】《中国抗生素杂志》
【年(卷),期】2007(32)12
【摘要】采用优化设计方法研究金属离子在林可链霉菌(Streptomyces lincolnensis)林可霉素生物合成中的交互作用,并应用均匀设计法,以最终发酵液的生物效价为目标函数建立模型,通过软件Uniform Design Version 3.00分析得出优化配比:FeSO4·7H2O 0.45g/L,CuSO4·5H2O lg/L,MnCl2 lg/L;按优化方案添加金属离子,使发酵液的最终生物效价较原配方提高20%,明显提高了林可霉素的发酵水平.
【总页数】5页(P723-726,761)
【作者】汤海云;郭元昕;李啸;储炬
【作者单位】华东理工大学生物反应器工程国家重点实验室,上海,200237;华东理工大学生物反应器工程国家重点实验室,上海,200237;华东理工大学生物反应器工程国家重点实验室,上海,200237;华东理工大学生物反应器工程国家重点实验室,上海,200237
【正文语种】中文
【中图分类】R978.1
【相关文献】
1.金属离子对褐黄孢链霉菌生长和纳他霉素生物合成的影响 [J], 相启森;岳田利;袁亚宏;高振鹏;王慧;王爽
2.金属离子对红霉素生物合成的影响 [J], 李啸;陈长华;李友元
3.金属离子对螺旋霉素生物合成的影响 [J], 毛全贵;赵胜利;韩德全;张永红;汉国旗;刘守强
4.常见金属离子对过氧化氢酶生物合成的影响 [J], 冯伟;冯雅琪;徐晶雪
5.金属离子对丹参酮Ⅱ_A和原儿茶醛生物合成的影响 [J], 郭肖红;高文远;陈海霞;黄璐琦
因版权原因,仅展示原文概要,查看原文内容请购买。
博士学位论文-中国科学院青岛生物能源与过程研究所机构知识库

In this research, two chaperones (protein disulphide isomerase, pdi1 and HSP70 family chaperones, bip1) were overexpressed and a polyketide synthase (PKS65172) was knocked out respectively in QM6a, the results showed that both methods could improve the cellulase production of QM6a to a certain extent. The differences of secondary metabolites between PKS65172 gene knockout strain and wild-type QM6a were also analyzed, there are many important mechanisms are worthy of being studied.
感谢中国科学院青岛生物能源与过程研究所提供的科研学习平台,感谢中丹 学院提供我赴丹麦科技大学联合培养的机会,感谢研究所各位领导给予的帮助!
古细菌Pyrococcusfuriosus嗜热_淀粉酶基因在大肠杆菌中的表达

古细菌Pyrococcus f uriosus 嗜热α2淀粉酶基因在大肠杆菌中的表达3沈 微 王正祥 刘吉泉 诸葛健(江南大学生物工程学院和教育部工业生物技术重点实验室,无锡,214036)摘 要 用PCR 方法从嗜热古细菌Pyrococcus f uriosus 的基因组DNA 中扩增出胞外α2淀粉酶完整结构基因,插入表达载体p ET28a 中构建成质粒p ET 2amy (sig +),以质粒p ET 2amy (sig +)为底物扩增出不含信号序列的α2淀粉酶成熟肽基因片断,获得质粒p ET 2amy (sig 2),将重组质粒分别转化大肠杆菌BL21(DE3)。
在T7启动子和lac 操纵子控制下,通过IPTG 的诱导在重组大肠杆菌细胞内分别表达出含信号肽和不含信号肽的融合蛋白。
其中不含信号肽的融合蛋白具有与P.f urio 2sus 产生的胞外α2淀粉酶相似的酶学性质:最适p H 为5.0,最适温度约为95℃,在121℃下热处理1h 酶活仍能保持50%以上;含信号肽序列的基因的表达产物不能测到酶活。
关键词 Pyrococcus f uriosus ,耐热α2淀粉酶,融合表达 第一作者:博士研究生。
3教育部骨干教师资助计划(No.1696) 收稿时间:2002-08-24 耐高温α2淀粉酶是应用最广泛的工业酶制剂之一,开发耐热性能好的α2淀粉酶有着重要的意义。
近年来,人们从嗜热古细菌中分离了多种高耐热的α2淀粉酶,其中来源于Pyrococcus f uriosus 的胞外α2淀粉酶以其优异的耐热性能而受到关注[1]。
P.f uriosus 是一种从海洋温泉附近的高温海水中分离到的高嗜热古细菌,其最适生长温度达100℃[2],产生的胞外α2淀粉酶不但具有很高的热稳定性,活性不依赖于金属离子,而且其最适p H 在4.5~5.5之间[3,4],与目前广泛应用的来源于黑曲霉的糖化酶的最适p H 接近,液化液可直接用于糖化。
古生菌的生理生态特点

古生菌的生理生态特点董怡萱(食品科学与工程2班生命科学学院黑龙江大学哈尔滨 150080)摘要:很多古菌是生存在极端环境中的。
一些生存在极高的温度(经常100℃以上)下,比如间歇泉或者海底黑烟囱中。
还有的生存在很冷的环境或者高盐、强酸或强碱性的水中。
然而也有些古菌是嗜中性的,能够在沼泽、废水和土壤中被发现。
很多产甲烷的古菌生存在动物的消化道中,如反刍动物、白蚁或者人类。
古菌通常对其它生物无害,且未知有致病古菌。
关键词:古生菌、微生物、极端环境Ancient lives the fungus the physiological ecology characteristicDong Yixuan(The 2th Food science and engineering, College of Life Science, Heilongjiang University,Harbin, 150080)Abstract:Many Archaea are the survival in the extreme environment. Some survivals in extremely high temperature (frequently 100℃above), for instance geyser or in seabed black chimney. Also some survivals in very cold environment or Gao Yan, strong acid or in alkalinity water. However also some Archaea are neutrophil, can in the bog, the waste water and the soil was discovered. Very prolificacy methane Archaea survival in animal's digestive tract, like ruminant, termite or humanity. The Archaea is usually harmless to other living thing, and unknown has the pathogenesis Archaea.Key words:Ancient lives the fungus, the microorganism, the extreme environment1977年,Carl Woese以16S和18S rRNA的寡核苷酸序列比较为依据,提出的独立于真细菌和真核生物之外的生命的第三种形式。
材料类英文期刊汇总

材料类英文期刊汇总英文材料期刊简介一键发布日志|编辑|删除nn/tp保存于2021年06月21日18:41分类:个人日记太阳能材料和太阳能电池,荷兰go?tgurhfissn:0927-0248,1979年创刊,全年20期,elsevierscience出版社出版,sci、ei收录期刊,sci2021年影响因子1.188,2021年ei收录302篇。
刊载光电、光热和光化太阳能转换的材料科学问题,以及太阳能电池的理论、技术、应用、相关经济问题的研究论文,涉及有关的固体物理学、冶金学、陶瓷学、光学和电化学等。
* l@g&C*-JSOLIDSTATIONICS固体离子,荷兰f*'f/,issn:0167-2738,1980年创刊,全年40期,elsevierscience出版社出版,sci、ei收录期刊,sci2021年影响因子1.599,2021年ei收录372篇。
刊载与固体中的扩散及反应性有关的物理、化学、材料科学问题的理论与实验研究论文和评论,涉及固体中缺陷的物理和化学问题、固体中或固体间的反应(掺杂、腐蚀、氧化、烧结)及离子迁移的理论、机理及量测等。
*|Vhcc固态科学,法国o\dz`issn:1293-2558,1964年创刊,全年12期,elsevierscience出版社出版,sci、ei收录期刊,sci2021年影响因子1.327,2021年ei收录185篇。
1999年前刊名为europeanjournalofsolidstateandinorganicchemistry,刊载固体化学和无机化学领域的研究论文和简讯。
文章用英文发表。
VXPCW\表面科学表面科学,荷兰zr]uv(*lissn:0039-6028,1964年创刊,全年78期,elsevierscience出版社出版,sci、ei收录期刊,sci2021年影响因子2.063,2021年ei收录990篇。
(包括《表面科学快报》)刊载界面物理和界面化学的基础理论和实验研究论文。
《额尔敦-乌日勒的活性成分分析及其对小胶质细胞基因调控作用的研究》范文

《额尔敦-乌日勒的活性成分分析及其对小胶质细胞基因调控作用的研究》篇一摘要额尔敦-乌日勒(Erchim-Urir)作为一种具有深厚药用历史的天然产物,其活性成分的深入研究对于理解其生物活性和药理作用具有重要意义。
本文通过对额尔敦-乌日勒的活性成分进行详细分析,并探讨其对于小胶质细胞的基因调控作用,为进一步开发其药用价值提供理论依据。
一、引言额尔敦-乌日勒是一种具有重要药用价值的天然植物,被广泛用于传统医学中。
近年来,随着对其活性成分的深入研究,发现其具有多种生物活性和药理作用。
小胶质细胞作为神经系统中的重要组成部分,其基因表达和功能调控与许多神经系统疾病密切相关。
因此,研究额尔敦-乌日勒对小胶质细胞基因调控的作用,有助于揭示其在神经系统疾病治疗中的潜在应用价值。
二、额尔敦-乌日勒的活性成分分析1. 化学成分额尔敦-乌日勒的主要化学成分为多种生物碱、黄酮类化合物、多糖等。
其中,生物碱是其主要的活性成分,具有抗炎、抗氧化、抗肿瘤等多种生物活性。
2. 提取与分离采用现代分离技术,如高效液相色谱、薄层扫描等,对额尔敦-乌日勒进行提取与分离,得到纯度较高的活性成分。
3. 结构鉴定通过光谱分析、质谱分析等手段,对分离得到的活性成分进行结构鉴定,为后续研究提供基础数据。
三、额尔敦-乌日勒对小胶质细胞的基因调控作用1. 细胞培养与处理采用小胶质细胞株进行体外培养,将额尔顿-乌日勒的活性成分加入细胞培养体系中,观察其对小胶质细胞的生长和基因表达的影响。
2. 基因表达分析通过实时荧光定量PCR、基因芯片等技术,分析额尔顿-乌日勒对小胶质细胞基因表达的影响,探讨其可能的基因调控机制。
3. 信号通路分析利用生物信息学方法,分析额尔顿-乌日勒对小胶质细胞信号通路的影响,进一步揭示其基因调控作用的分子机制。
四、结果与讨论1. 活性成分分析结果通过化学分析和结构鉴定,明确了额尔顿-乌日勒的主要活性成分为多种生物碱、黄酮类化合物等。
古菌的奥秘

氨氧化古菌的研究
中温泉古菌
中温泉古菌的研究
同位素示踪法
中温泉古菌的研究
泉古菌amoA基因
科学家 Venter Treusch Klnneke
氨氧化作用
氨氧化古菌的研究
与氨氧化细菌 相比,中温泉 古菌并不包含 所有氨氧化基 因,命名为氨 氧化古菌
古菌的应用前景 能源开发
环境保护 科研价值
古菌的应用前景
细胞结构
代谢方式
古菌的简介
球形 杆状 螺旋形 耳垂形 盘状 不规则形状
古菌的简介 生存环境
极端环境 沼泽、土壤中
动物的消化道中
古菌的简介
广布性 多样性
适应性
古菌的简介
极热 极寒 高盐分 强酸性 极度缺氧
古菌的简介
计10纲, 约99属。 每个属的种数不一 但普遍不多 合计约有195种
古菌的简介
产甲烷古菌
无机或有机化合物→甲烷、二氧化碳
厌氧转化
可再生清洁能源
古菌的应用前景
沼气生产
古菌的应用前景
获取极端环境下仍具有活性的酶
PCR技术中的 热稳定DNA聚合酶
嗜热古菌
虽然研究了很久, 但我们对古菌仍然知之甚少
相信不远的将来, 古菌研究的突破性进展, 将会为我们的生活带来新变化。。。
古菌的奥秘
成员:张信凯、崔馨匀、 黄剑秋、 韩林慧、 唐蓉、朱天赐
目录
古菌的简介 极端环境下的古菌——海洋古菌 非极端环境下的古菌——氨氧化古菌 古菌研究的应用 结语
古菌的简介
什么是古菌?
古菌又称古细菌 、古生菌或太古 生物、古核生物 ,构成生物分类 的一个域。
古菌的简介
与细菌、真核生物 的区别
Argonaute蛋白结构及其在植物中的研究进展
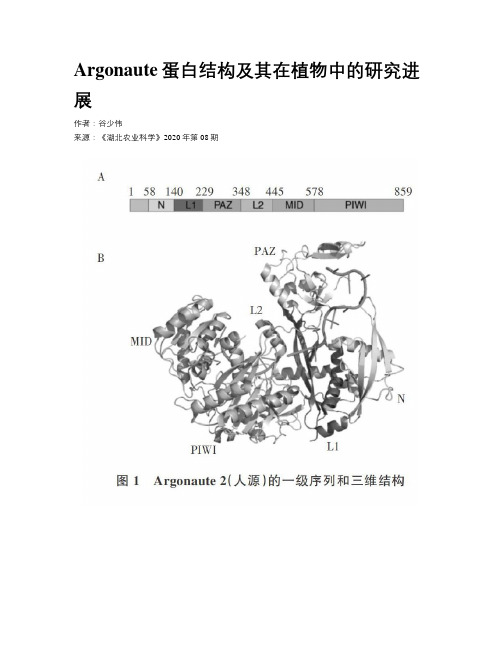
Argonaute蛋白结构及其在植物中的研究进展作者:谷少伟来源:《湖北农业科学》2020年第08期摘要:在真核细胞中,许多小的非编码RNA与Argonaute蛋白结合形成RNA诱导沉默复合体,并通过识别互补的靶标RNA来调控基因表达,此过程叫RNA干扰。
在植物的RNA干扰途径中,Argonaute-sRNA复合物可以通过多种不同的机制发挥作用,进而参与病原体防御、植物发育调控等关键的生物过程。
综述了Argonaute蛋白的分类和结构特征、在植物体中参与RNA干扰的机制及生物学功能等研究进展。
关键词:Argonaute蛋白;结构;RNA干扰;功能中图分类号:Q51文献标识码:A文章编号:0439-8114( 2020) 08-0011-06D01:10.1408 8/j .cnki.issn0439-8114.2020.08.002开放科学(资源服务)标识码(OSID):小的非编码RNA是原核生物和真核生物中基因表达控制的重要参与者。
在真核生物中,几类小的非编码RNA通过特异性识别互补的靶标RNA来调节基因表达,并保护细胞免受外源性和内源性有害遗传物质的影响。
Argonaute蛋白是RNA诱导沉默复合体( RISC)的核心蛋白,是表观遗传调控的主要参与者,在RNAi中通过剪切靶向mRNA、翻译抑制、DNA甲基化等方式发挥作用。
本研究综述了Argonaute蛋白的分类和结构特征、在植物体中参与RNA 干扰的机制及生物学功能等研究进展,旨在为后期研究提供基础。
1 Argonaute蛋白的分类和结构特征Argonaute蛋白在拟南芥突变体的研究中被首次提及[1],后来发现该蛋白是真核生物中RNA干扰( RNA interference,RNAi)途径的关键参与者。
最近对原核生物基因组分析的结果显示,Argonaute蛋白同时也存在于古细菌(约30%)和细菌(约10%)中[2.3]。
1.1 Argonaute蛋白的分类Argonaute蛋白在各种生物中广泛存在,而且在不同的物种之间存在较高同源性,这表明它们在进化过程中具有古老的起源和高度保守性[2]。
古细菌Halobacterium salinarum耐热因子的研究

94科技资讯 SCIENCE & TECHNOLOGY INFORMATION①作者简介:张栩(1984—),女,硕士,讲师,研究方向为生物化学与分子生物学。
DOI:10.16661/ki.1672-3791.2010-5042-2957古细菌Halobacterium salinarum耐热因子的研究①张栩 崔珊珊(长春职业技术学院 吉林长春 130033)摘 要:该研究使用的Halobacterium salinarum是极端嗜盐菌的模型生物之一。
通过对菌热处理后,添加培养滤液,进行37℃振荡培养,测量OD600值观察菌的生长情况,以确定该菌是否含有耐热因子及探讨了该菌培养滤液中所含成分对该菌生长繁殖的影响。
结果,80 ℃热处理后,如果不添加培养滤液,该菌不生长;添加培养滤液时,从第十天左右开始,OD600≈0.2~0.3,并且通过显微镜的观察,确认了菌的存在。
另外,将培养滤液用活性炭进行吸附,再用乙醇进行溶出后的效果也进行了研究。
结果显示:将100%的乙醇溶出液分别加入热处理菌后进行培养,结果70 ℃的热处理菌有生长延迟的倾向。
可以推断出:培养滤液中有可能存在某种阻碍该菌生长的成分。
根据该研究,明确证实了培养滤液中既含有Halobacterium salinarum的生长促进因子,又含有阻碍因子。
通过分析这些成分,期待可以将休眠机制解释清楚并对该菌以外的极端嗜盐菌的耐热性的研究提供研究基础。
关键词:极端嗜盐菌 Halobacterium salinarum 耐热因子 培养滤液中图分类号:Q939 文献标识码:A 文章编号:1672-3791(2021)01(a)-0094-03Study on Heat Resistance Factor of Halobacterium SalinarumZHANG Xu CUI Shanshan(Changchun Polytechnic, Changchun, Jilin Province, 130000 China )Abstract: Halobacterium salinarum is a model organism of extreme halophiles. In this study, f irstly, we collected the culture filtrate of the halophile culture solution which continued to shake culture at 37 ℃ after heat shock at different temperature. Then we continue to culture the heat-activated halophile by using the medium with the culture f iltrate, and observed the growth by measuring the OD600 value to determine whether the halophile contains heat-resistant factors. Simultaneously exploring the effects of the components contained in the culture f iltrate on the growth and reproduction of the halophile. Results: After heat treatment at 80 ℃, only when the culture filtrate was added, the halophile began to grow from about the tenth day, OD600≈0.2~0.3, and the presence of the halophile was confirmed by microscopic observation. Further, the effect of culture filtrate adsorbed by activated carbon and dissolved by ethanol was also studied. As a result, 100% ethanol solution was added to the heat-treated halophile for culture, which at 70 ℃ tend to have a delayed growth. It can be inferred that there may be some components in the culture filtrate that hinder the growth of the halophile. According to the present study, it was clearly confirmed that the culture filtrate contained both the growth promoting factor and the inhibitory factor of halophile. By analyzing these components, it is expected that the dormancy mechanism can be explained clearly and the research basis for the study of the heat resistance of other extreme halophiles is provided.Key Words: Extreme halophiles; Halobacterium salinarum; Heat resistance factor; Culture filtrate最早记载极端嗜盐菌存在的是《本草纲目》中记载的“引清水注入,久则色赤。
《唐古特白刺水孔蛋白NIP家族基因调控植物响应环境胁迫的分子机理研究》范文

《唐古特白刺水孔蛋白NIP家族基因调控植物响应环境胁迫的分子机理研究》篇一一、引言植物作为地球上最重要的生物之一,其生存环境经常受到各种环境胁迫的影响,如干旱、盐碱、寒冷等。
为了适应这些环境变化,植物进化出了一套复杂的响应机制。
近年来,随着分子生物学和基因工程技术的飞速发展,植物对环境胁迫的响应机制逐渐被揭示。
其中,唐古特白刺水孔蛋白NIP家族基因在植物响应环境胁迫的过程中发挥了重要作用。
本文旨在探讨唐古特白刺水孔蛋白NIP家族基因调控植物响应环境胁迫的分子机理。
二、唐古特白刺水孔蛋白NIP家族基因概述唐古特白刺水孔蛋白NIP家族是一类重要的植物膜蛋白家族,具有调节植物细胞内水分和溶质运输的功能。
该家族基因在植物中广泛分布,对植物的生长和发育具有重要影响。
在环境胁迫条件下,NIP家族基因的表达水平会发生显著变化,从而影响植物的适应性。
三、NIP家族基因的调控机制1. 基因表达调控:NIP家族基因的表达受多种内外因素的调控,包括光照、温度、水分等环境因素以及激素、转录因子等生物因素。
这些因素通过调控NIP家族基因的转录和翻译过程,影响其表达水平。
2. 蛋白相互作用:NIP家族蛋白与其他蛋白之间的相互作用也是调控其功能的重要方式。
例如,NIP家族蛋白与AQP(水通道蛋白)等其他膜蛋白之间的相互作用,可以影响细胞膜的通透性和水分运输效率。
3. 信号转导:在环境胁迫条件下,植物会通过一系列的信号转导途径来响应。
NIP家族基因作为信号转导途径的重要节点,可以接收并传递环境胁迫信号,从而调控相关基因的表达和植物对环境胁迫的适应性。
四、唐古特白刺水孔蛋白NIP家族基因在响应环境胁迫中的分子机理1. 干旱胁迫:在干旱条件下,NIP家族基因的表达水平会显著提高,从而促进植物细胞对水分的吸收和运输。
此外,NIP家族蛋白还可以与其他膜蛋白相互作用,提高细胞膜的通透性,进一步促进水分的运输。
2. 盐碱胁迫:盐碱胁迫会导致植物体内离子平衡失调,影响植物的正常生长。
微生物吸附金属离子研究与发展

微生物吸附金属离子研究与发展尹敬群;田君【摘要】本文详细论述了微生物吸附金属离子研究及发展过程,介绍了微生物吸附机制、吸附过程以及微生物吸附剂类型,对其在重金属废水处理工业中的应用前景进行了展望。
【期刊名称】《生物化工》【年(卷),期】2016(000)001【总页数】4页(P65-68)【关键词】微生物吸附;金属离子;微生物吸附剂;废水【作者】尹敬群;田君【作者单位】江西省科学院应用化学研究所【正文语种】中文【中图分类】X703微生物吸附(biosorption)是指用微生物细胞经过一系列生物化学作用而吸附水溶液中的金属或非金属物质,该概念最早是由Ruchhoft C C于1949年提出来的[1],他利用活性污泥去除水中的放射性元素钚(239Pu),并认为钚的去除是由于微生物的繁殖形成了“有巨大表面积的胶状基质能吸收放射性材料”,这类具有较大面积的凝胶网使微生物具有吸附能力。
大量研究表明,一些微生物如细菌、真菌和藻类和一些水生动植物在水体净化中起着独特的作用[2],它们对一些重金属离子有很强的吸附和富集能力。
如细菌、放线菌、霉菌、酵母菌、藻类等都能有效地从水溶液中富集微量的重金属离子[3]。
生物吸附重金属离子兴起于20世纪80年代,1982年Teszos的研究结果表明少根根酶(Rhizopusar hizus)对钚和铀有很高的吸附量;1984年Hosea等人发现普通小球藻(Chlorella vulgaris)对金有很高的亲和力;1986年Norbeng等人发现动胶菌对Cu2+具有较高的选择性吸附能力[4]。
20世纪90年代到来,国内外有关这个领域的研究迅猛发展。
首先是生物吸附材料不断涌现,Holan Z R等人用褐藻吸附Co2+[5];Huang C用曲霉吸附Zn2+;Volesky B用酵母菌吸附Cd2+[6];李清彪等人研究了用黄孢原毛平革菌(Phanerochaete chrysosporium)对Pb2+的吸附[7];刘瑞霞等人研究了用滕黄微球菌(Micrococcus luteus)吸附Cu2+[8]。
微生物应用ppt

传统微生物培养缺陷
通用培养基对大多数极端环境下难培 养微生物(99%以上)没有选择性。
培养条件很难与自然条件一致。
对微生物领域认识的局限。
极端环境微生物培养方法归纳
模拟自然环境分离获得纯培养。 采用荧光标记已探测菌基因,追踪分离。 极端营养研究,实现微生物分离培养。
Isolating “Uncultivable” Microorganisms in Pure Culture in a Simulated Natural Environment
微生物应用技术
曹理想 中山大学生物化学系
主要内容:
前言 微生物多样性与微生物资源开发技术 发酵培养与下游技术 育种技术 检测技术 药物筛选技术 细菌生物膜与细菌信号传导 燃料电池与生物传感器 生物转化技术 疫苗
前言
微生物的特性: 1、体积小、面积大 2、吸收多、转化快 3、生长旺、繁殖快 4、易变异、适应强 5、种类多、分布广
物的工程菌 扩大微生物来源 应用定向生物合成和突变生物合成的原
理寻找新的代谢物 对已知化合物进行化学改造
11 厌氧菌研究技术
烛罐培养技术 钢丝绒技术、厌氧转管法和预还原技术 瓶皿法及厌氧手套箱
厌氧菌的种类
根据对氧气的耐受性,分为三类:专性厌氧菌、微需 氧菌与兼性厌氧菌。
专性厌氧菌指不能利用分子氧作为电子终末受体,只能 在低氧分压、低氧化还原电位下才能生长的细菌,分 为三类:
术 8、微生物酶反应生物合成和化学合成反应相
结合
纯培养技术与科赫法则的不足
不了解病毒 两种以上微生物在致病方面与干酪发酵
过程的协同作用 对海洋微生物、瘤胃微生物、肠道微生
物的研究需掌握微生物类群的生态关系
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
EF8G( H XYZ[Y\]^ EFI- JFI _‘ab EFI acdedfg * hijklmnodp!’/c
p!qb 5 rsBtuvwxyz<{TU| KL3 M * K@3 M } N83 M HYZ~ !" K(%O8@&$&$$?G P8@@8G$O99 bc QQ # EF8G( H J‘cz $ $ 5 %&’( Z )*+, | ( YJ‘t - cz $ $ ./ 5 01 N| (2 : 34 EH EF8G( HQQ 5 +R + J‘ 5 6 EF8G( HQQ g 789 "} :; TU <= |L >?@ 5 345 EF8G( HQQ* uvwxyz * AB : 34 E
u v w x y z m ! 30 i ,5 ,,,0 " jN 2 q 5 r ,5 . 7Z ]^J stu myz v * KLN:3* K@N:3* N8N:3 m[wxa;iyz m ]^J * u mwx 2 s E b 5 w x 6 { & . "Z* D 30 & w x * | 3,, "Z5 /D&w;D"M!uvwxyz>}~ !" 2q5
#$%&’()
!"#$$% ! !"#$ %"&’"( )"(*&+ " ,-.& /0123 450$63 7$83 * !""#$ #% ! +, " - ./0 1 ./2
./0
CB
CD
!"# &’()* +,, $%&’()*+,-./0!
! " # $ %&’ ()*
+,,.2+ "
! DEFGHIGJK45LMNOP * QRS$!"#$TUV * QRS$#$$W * QR 12
!
!- #
)H$IJ
%&’()*K
KL
3M
Z9L8@< @?7’() 8 c9@<9@L $&@G%8@% : ‘ # % e fR 7&: ‘ + ,5 .3 +5 ,B ,5 .W A 32+ .3 0+B ++ WWW 2 W0B +3 ,,. . 3.+ K@3 M N83 M
uvwxyz # ; $ h D % h & ‘ # 9 # + } # 3* & ‘J ’ 9_ +5 EF8G( H c:34 EJ‘
EF8G( H XYZ [Y\ ]^ EFI- JFI _‘ ab EFI acdedfg 5 hijklmnod p!’/cp!qb S H ;g I 5 CDEF \ G \ u \ J * K L EF8G( H 5 M D 6 EF8G( HQ } EF8G( HQQ* (NO g PQRSTEF \ * URVW cJK EF \ 5 l !" b * EF8G( HQ XY>Z’ * l ~ ! " } [ d p ! b * EF8G( HQQ X Y > Z ’ 5 EF8G( H J K \ k l m Y 2 L > c ] ^ J _ HQS4Q ‘ab g} c ‘gb C /* 2 D 5 EF8G( H cijD def | g L > cp" hi 5 7 j ’A k6l * m no EF8G( H c|^ pqrRs * tR c uZ S v w bm 65 $&:9 EF8G( HQ5 EF8G( H c I\ x>AB : 3y4 E 5 CDz qJK {| 65 $&:9 EF8G( HQ c: 34 EJ‘ } M XY ~ !" ~ f\ # 9fK $ c % 5 YZ~ !" K(%O8@&$&$$?G P8@@8G$O99 bc EF8G( HQQ c z qJ K &’( ^ ) * hcSJK \ H 65 $&:9 EF8G( HQ Q RVW c q * \ * 0 +, klY ~ !" ~ f\ # 9 fK $ c % 5 mn’ uvwxyz )* - x| 3M 3M KL * K@ } N83 M H K(%O8@&$&$$?G P8@@8G$O99 EF8G( HQQ J‘cz $$* ./xgJ‘:34Ec / A 0 .0 6 9 1 * 2 Y 3 TUg 789 ";: 3 4 EJ‘o]^J:; c45 5
‘o]^J:; 45 <=|L>?@ 5
&*=*9*13*)
+ #O%8@9 F* H8)?>9 K* K&)9>8k8 K* N)&?$O E f* Q%8U8 K* l8@8U8 V5 ;$+-0(6$2.<1= #???$ >@ - /,0 3 mO8@L n c* IU8:(k V* Z8$>G V I5 >3 ;&-.(<$+#+?1= #??%$ #%? ! +3 " - B.3. B W Q%8U8 K5 /<+-3 @&.#3 ,-&A3 7-$3= #??"$ @% - .0.2 h)8@> \* I:’()% V* N8o(@8X( N* a&?:7( f f5 @"-#($- ,-$A2 B(23= #??A$ !! ! 3W " - 03W2 0 H8)?>9 K* H8U8GO9 l* l&$O9 l* K?)&U8 I* l&L8 n* K&)9>8k8 K* Q78@8>8 a* l8@8U8 V5 >3 ;&-.(<$+#+?1= #??@$ #@" ! 3B " - /3,2 / J8X9(G f h* H&G%&7G>8 m5 H&G%&7G>U m* f&)<8@ V E5 K8%%O(kG J I5 7-$(8-(= #??#$ !B! - .. 2 . n8@L p* V%(9%o a I5 7.<"-."<(= #??B$ > ! 3 " - +B+ l8%8U8@8L9 l* K9U8L8k8 K* K8%G?GO978 K* QGO9>8k8 K* l8@8U8 V* Q>(O8)8 K* K8%G?o8>9 a* K&)9>8k8 l5 @&."<(= #??"$ >A% - B,/ A Z89 Z H* n&>&%8 H* H?8@L Z p* l97 E* l97 V H5 7.<"-."<(= !"""$ @ ! . " - .A2 +, l97 E* V8@<:() V f* ^&:<78@ V* n&>&%8 H* N:8)> I f* l97 V H5 ;$+.(-03 C(..3= #??@$ !" ! B " - 3,2 ++ +3 c:8$> N c* N&k8@ f I5 D8+<?3 50(63= #??A$ >> - 0.,0 l8@8U8 V* #&’8%8>( K* Z9? n n5 >3 ;$+#3 50(63= #??C$ !%# ! 0+ " B323A +B l8GO9k8L9 a* f(8@%(?) J* H8)?>9 K* l8%8U8@8L9 l* l8@8U8 V* K&)9>8k8 l5 /<+.3 E8?3= #??C$ ? ! +, " - .02 +W J?<(X a* Z97 N5 >3 /0123 50(63= !""#$ #"B - WWW/
8 " . 77&: F Z ‘ + KLN:3 %9%)8%(< %& ,5 .W 77&: F Z ‘ + =)&%(9@g ’ " 0 77&:F Z ‘ + K@N:3 %9%)8%(< %& ,5 0+ 77&:F Z ‘ + =)&%(9@g $ " W 77&:F Z ‘ + N8N:3 %9%)8%(< %& ,5 B. 77&:F Z ‘ + =)&%(9@
CA D C. D C Ʒ
:;$<=
4 6 TS4+33+ 7 8 4 K 9 D : : ; < = N8:&)97(%)U V$9(@$( N&)=5 6 W3,, # uvwxyz >= H(%%9$O 6 T@9X()G8: +/E ?@ 4 A 9 = K9::9=&)( 6 K9::94Y Z8’& B C ] D " E = K9::9=&)( 6 T:%)84 JK t FL [)((4+0 4 ABFG * HI DEy 0,,,= J6 \O8)78$98 6 V(=O8<(]4^305 V ’ ] MN K9::94Y Z8’& BC ] D " ED " * OPQ _ +2 K! R $7= V R STM 6 D UC.V * W X f c V R Y Z [ M 6 3, 77&: R Z ‘ + a)9G R HN:G =H b .5 ,* B 77&:R Z ‘ + \9]^ 5 #- ! +5 35 + 67>? !"#$%&’()*+(,-