第六章 线粒体和叶绿体
细胞生物学:第6章 线粒体和叶绿体

透射电镜
1. 外膜:包围在线粒体最外面的一层单位膜
结构特点: 光滑而有弹性;
具直径2~3nm的小孔
含有孔蛋白,所以外膜的通透性 非常高,可允许所有相对分子量小 外膜 于5x103以下的小分子通过,使 得膜间隙中的环境几乎与胞质溶 胶相似 ;
脂质与蛋白质比值为1:1,比内
膜高(0.3:1),即:外膜中的蛋
是电子传递释放能量驱动的一种质子泵
复合物Ⅳ:细胞色素C氧化酶 组成: 含有四个氧化还原中 心(细胞色素a和a3以及两个铜 离子)
作用:催化电子从细胞色素c 传给O2;每传递一对电子要从 基质中摄取4H+,其中2 H+转移 到膜间隙,2 H+ 用于水的形成
线 粒 体 内 膜 中 的 运 输 系 统 丙酮酸
3. 膜间隙:内外膜之间的腔隙 宽约6-8nm,含许多可溶性酶、底物及辅助因子;
由于外膜的通透性非常高,使得膜间隙中的环境几 乎与胞质溶胶相似 ;
4. 基质或内室:内膜所包围的脊外空间 含有大量的酶:参与三羧酸循环、脂肪酸氧化、氨 基酸降解等有关的酶都存在于基质之中 ;
作用:催化琥珀酸(FADH2)中的2个低能电子 通过FAD和铁硫蛋白传给辅酶Q。电子传递时不伴 随质子的跨膜转移
只是电子传递体而非质子泵
复合物Ⅲ:细胞色素bc1复合物
或称CoQ-细胞色素c还原酶、细胞色素还原酶
组成:包括1个细胞色素c1、1个细胞色素b和1个 铁硫蛋白 作用:催化电子从辅酶Q传递给 细胞色素c。每传递一对电子会伴 随4H+泵出(通过Q循环的途径实 现)
占细胞质比例的50%以上
线粒体的融合与分裂 可能是细胞应对生命活动需求进行合理分布的手 段之一
线粒体经常包围 着含脂肪酸的油滴, 从中获取大量的用 于氧化反应的原料
第六章线粒体和叶绿体-细胞生物学

线粒体形状的可塑性
• 多分布在细胞功能旺盛的区域,可向这些区域迁 移,微管是其导轨、马达蛋白提供动力。
线粒体通常位于高度需要ATP的细胞部位
(二)超微结构与化学组成
基质 内膜 外膜 膜间隙
An TEM image of mitochondrion
Models of mitochondrial membrane structures
c 2 血红素 O2 2 Cu
FADH2呼吸链——复合物Ⅱ、Ⅲ、Ⅳ组成,催化琥珀酸的 脱氢氧化
氧化磷酸化的作用机理
• (一)质子动力势(P91) • Mitchell(1961)提出“化学渗透假说”。认为: • 电子沿呼吸链传递时,所释放的能量将质子从内膜
基质侧(M侧)泵至膜间隙(C侧),形成质子动力势 ( △P)。
(三)、质子转移与质子驱动力的形成: NADH呼吸链
内外膜间隙
细胞色素 c
线粒体 内膜
基质
ቤተ መጻሕፍቲ ባይዱ泛醌
电子传递 路线
NADH 脱氢 酶复合体
细胞色素 bc1 复合体
细胞色素氧 化酶复合体
> 40 多肽链
> 11多肽链
NADH Flavin Q 2 血红素
7 铁硫中心
1 铁硫中心
> 13多肽链
1.外膜: 含有许多大型膜通道蛋白 “porin” 允许分子量小于 < 5000的物质自由通过 标志酶为单胺氧化酶
2.内膜:
内膜具有嵴,能扩大表面积(5~10倍),
分两种:①板层状、②管状;嵴上有基粒。 是建立 H+ 电化学梯度的场所 含有双重磷脂分子心磷脂 (cardiolipin).心 磷脂含有四个脂肪酸, 使内膜对离子具有高 度的不通透性 内膜上的蛋白质包括:
第6章线粒体和叶绿体

2.光合磷酸化 (1)CF0-CF1ATP 合酶 (2)光合磷酸化的类型 (3)光合磷酸化的作用机制 (4)ATP 合成机制
叶绿体类囊体膜中进行的电子传递和H+跨膜转移
(三)光合碳同化 1.卡尔文循环
(1)羧化阶段 (2)还原阶段 (3)RuBP再生阶段
2. C4 途径
Ø存在于一些热带或亚热带起源的植物(C4 植 物)中,如甘蔗、玉米、 高粱等。
γ亚基的一个结构域构成穿过F1的中央轴,另一结构域与3 个β亚基中的一个结合。
ε亚基协助γ亚基附着到ATP合酶的基部结构F0上。 γ与ε亚基具有很强的亲和力,结合形成“转子”(rotor),
旋转于α3β3的中央,调节3个β亚基催化位点的开放和关 闭。
δ亚基为F1和F0相连接所必需的。
ATP 合酶的基部结构被称作F0,是偶联 因子0 (coupling factor 0)的简称。与 亲水性的F1 相比,F0 是一个疏水性的 蛋白复合体,嵌合于线粒体内膜,由a、 b、c 3种亚基按照ab2c10~12 的比例组 成跨膜的质子通道。
(二)叶绿体的分化和去分化
Ø 叶绿体是原质体的一种分化方式;
Ø 叶绿体的分化表现为形态上和在生化与分子生 物学上;
Ø 在特定情况下,叶绿体的分化是可逆的,叶绿 体可去分化再次形成原质体。
(三)叶绿体的分裂 Ø 质体和叶绿体通过分裂而实现增殖。
Ø 叶绿体的分裂与线粒体分裂具有相同的细 胞动力学机制。
控与细胞类型相关,随着细胞分化而变化。
(二)线粒体的融合与分裂
动、植物细胞中均可观察到频繁的线粒体融 合与分裂现象,这被认为是线粒体形态调控 的基本方式,也是线粒体数目调控的基础。
(三)线粒体融合与分裂的分 子及细胞生物学基础
第六章-线粒体和叶绿体20160323

定功能。
Contact Sites of Mitochondria
②外膜融合。
主要表现为两个线粒体外膜脂质分子的混合,通过“融合装置”的介导完
成该过程。
③内膜融合。
这是关键性的一步,可能伴随着融合复合物内部分子的重排。当外膜融 合之后,两个线粒体内膜之间的障碍被消除,此时,内膜上并不产生新 的“融合装置”,也不需要再形成一个新的锚合环使两个内膜并列,而是 由同一个“融合装置”通过自身结构的变化将外膜的融合与内膜的融合耦 联起来,立即启动内膜融合。如果没有这种耦联,内膜脂质的混合会明 显缺乏效率。
1.线粒体的融合与分裂 线粒体融合与分裂是线粒体形态调控的基本 方式,也是线粒体数目调控的基础。
洋葱表皮细胞内线粒体在1min时间内相继发生融合与分裂的偶 联图
• 频繁的融合与分裂可能帮助线粒体共享遗传信息
1.线粒体的融合与分裂 线粒体融合与分裂是线粒体形态调控的基本 方式,也是线粒体数目调控的基础。
利用脱氧胆酸(一种离子型去污剂)处理线粒体内膜、分离
出呼吸链的4种复合物,即复合物Ⅰ、Ⅱ、Ⅲ和Ⅳ,线粒体内膜 上的电子传递实际上是由这些膜蛋白复合物分段催化完成的。 辅酶Q和细胞色素C不属于任何一种复合物。 辅酶Q溶于内膜,细胞色素C位于线粒体内膜的C侧,属于膜 的外周蛋白。
线粒体内膜电子传递复合物的排列及电 子和质子传递示意图
线粒体的主要功能是氧化磷酸化,合成细胞内95%的
ATP,为细胞的生命活动提供能量。
细胞质 线粒体 糖、脂肪 丙酮酸和脂肪酸 乙酰coA(经三羧酸循 环)
氢通过电子传递链到达氧生成水,同时ADP磷酸化生成ATP
真核细胞中碳水化合物代谢
氧化磷酸化:指当电子从NADH或FADH2经呼吸链传递给氧形 成水时,同时伴有ADP磷酸化形成ATP的过程。
第六章 线粒体与叶绿体

电子传递链
• 在三羧酸循环中,乙酰CoA氧化释放的大 部分能量都储存在辅酶(NADH和FADH2)分 子中。细胞利用线粒体内膜中一系列的电 子载体(呼吸链),伴随着逐步电子传递,将 NADH或FADH2进行氧化,逐步收集释放 的自由能最后用于ATP的合成,将能量储 存在ATP的高能磷酸键。
电子传递链
概念:在线粒体的内膜上存在有关氧化磷酸化的
脂蛋白复合物,它们是传递电子的酶体系,由一
系列能可逆的接受和释放电子或H+的化学物质所
组成,在内膜上相互关联地有序排列,称为电子
传递链或呼吸链。 呼吸链电子载体主要有:黄素蛋白、细胞色素、
铁硫蛋白、辅酶Q等。
• 黄素蛋白:含FMN或FAD的蛋白质,每个 FMN或FAD可接受2个电子和2个质子。呼吸 链上具有FMN为辅基的NADH脱氢酶,以 FAD为辅基的琥珀酸脱氢酶。 • 细胞色素:以铁卟啉为辅基的色蛋白,通过 Fe3+、Fe2+形式变化传递电子。呼吸链中 有5类,即细胞色素a、a3、b、c、c1,其 中aa3含有铜原子。
复合物Ⅲ
CoQ-细胞色素c还原酶,由10条多肽链组成,以二聚体形式存 在,每个单体包含两个细胞色素b、一个细胞色素c1和一个铁 硫蛋白。作用是催化电子从辅酶Q传给细胞色素c,每转移1对 电子,同时将4个质子由线粒体基质泵至膜间隙。所以复合物
Ⅲ既是电子传递体又是质子移位体。
复合物Ⅳ 细胞色素氧化酶,由6-13条多肽链组成,以二聚体形式存在, 每个单体含有细胞色素a和细胞色素a3及2个铜原子。作用是催 化电子从细胞色素c传给氧,每转移1对电子,在基质侧消耗2
基质
真核细胞中碳水化合物代谢
Hale Waihona Puke 一)氧化磷酸化的分子结构基础• 如何证明氧化(放能)和磷酸化(贮 能)是同时进行并密切偶联在一起的, 但却是由两个不同的结构系统实现的?
细胞生物学第六章 线粒体和叶绿体

F1和γ旋转的实验证明
氧化磷酸化抑制剂
1.电子传递抑制剂: 抑制NADH→CoQ的电子传递。阿米妥、鱼藤酮。
抑制复合物III。抗霉素A 。
抑制复合物IV。如:CO、CN、H2S。
电子传递抑制剂可用来研究呼吸链各组分的排列顺序,当
呼吸链某一特定部位被抑制后,底物一侧均为还原状态,
■ 呼吸链含有三个氧化磷酸化偶联位点 ● 氧化磷酸化偶联位点 根据对呼吸链中不同复合物间氧化还原电位的 研究,发现复合物Ⅰ、Ⅲ、Ⅳ每传递一对电子, 释放的自由能都足够合成一分子ATP,因此将 复合物Ⅰ、Ⅲ、Ⅳ看成是呼吸链中电子传递与 氧化磷酸化偶联的三个位点。 如果以FADH2作为电子供体,则只有两个ATP 合成偶联位点。
● 外膜
单位膜结构。外膜含有孔蛋白, 通透性高, 膜间隙中的环境与胞质溶胶相似。外膜含 有一些特殊的酶类,如单胺氧化酶,这种酶 能够终止胺神经递质,如降肾上腺素和多巴 胺的作用。 线粒体外膜的功能:建立膜间隙;对那些 将在线粒体基质中进行彻底氧化的物质先 行初步分解。
● 内膜
内膜富含心磷脂,通透性差,一般不允许 离子和大多数带电的小分子通过。 线粒体内膜通常要向基质折褶形成嵴, 其 上有ATP合酶(F0-F1 复合体)。 内膜的酶类可以粗略地分为三类∶运输酶 类、合成酶类、电子传递和ATP合成酶类。 内膜是线粒体进行电子传递和氧化磷酸化 的主要部位。在电子传递和氧化磷酸化过 程中,线粒体将氧化过程中释放出来的能 量转变成ATP。
■ 线粒体各部分的特性和功能 ● 蛋白分布:
● 功能
由于线粒体各部分结构的化学组成和性质的不同,它们的 功能各异。
● 标志酶
通过细胞化学分析, 线粒体各部位有特征 性的酶, 称为标志酶。 外膜: 单胺氧化酶 内膜: 细胞色素氧化酶 膜间隙: 腺苷酸激酶 基质: 苹果酸脱氢酶
第六章 线粒体和叶绿体
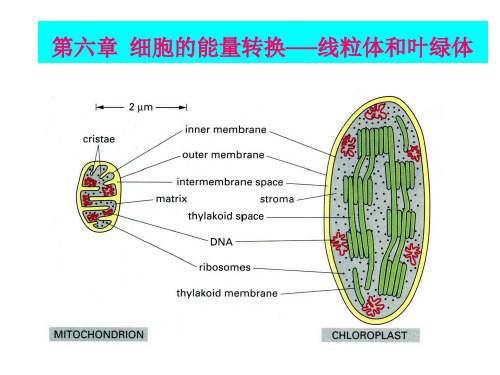
1) 内膜结构
单层膜,比外膜厚; 通透性较外膜小; 向内折叠成嵴,表面积极大增加,代谢效率 极大提高;
嵴的形状与数量和细胞种类与生理状况有关
2) 线粒体分类(根据嵴的形态结构)
(a)板层形 (b)管形 (a),(b)为基本形 (c)羽冠形 (d)网膜形 (e)绒毛形 (f)平行形 (g)同心圆形
返回
第二节 叶绿体与光合作用
●一.叶绿体(Chloroplast)的形态结构
二.叶绿体的结构与化学组成
●三.叶绿体的功能—光合作用
(photosynthesis)
一、叶绿体(Chloroplast)的形态结构
●叶绿体与线粒体形态结构比较
叶绿体内膜并不向内折叠成嵴;内膜不含电
子传递链;除了膜间隙、基质外,还有类囊体; 捕光系统、电子传递链和ATP合成酶都位于类囊体 膜上。 ●叶绿体超微结构
1964年由Paul Boyer最先提出来的。此学
说认为电子沿电子传递链传递使线粒体 内膜蛋白质组分发生了构象变化,形成 一种高能形式。这种高能形式通过ATP 的合成而恢复其原来的构象。未能找到 有力的证据。
(3)化学渗透学说 1961 由英国生物化学家Peter Mitchell最先提出来
的。此学说认为电子传递释放出的自由能和ATP合成
+
个H 泵出内膜。
c 内膜对H+不能自由通过,泵出
膜外侧的H+不能自由返回膜内侧,
因而造成H+浓度的跨膜浓度梯度。
此梯度包含的能量可驱使ADP和Pi 生成ATP。
d.氢通过ATP合成酶上 的特殊的途径,返回基 质。H+浓度梯度所释放
的自由能,偶联ADP和
第六章 线粒体和叶绿体
ctDNA复制的时间:G1期,复制仍受核控制
二、线粒体和叶绿体的蛋白质合成
1、线粒体和叶绿体合成蛋白质的种类十分有限 2、线粒体或叶绿体蛋白质合成体系对核基因组有依赖性
3、参加叶绿体组成的蛋白质来源有三种情况
■由ctDNA编码,在叶绿体核糖体上合成 ■由核DNA编码,在细胞质核糖体上合成 ■由核DNA编码,在叶绿体核糖体上合成 4、线粒体蛋白的来源
第二节 叶绿体与光合作用(photosynthesis) 一、叶绿体(Chloroplast)的超微结构 1、叶绿体膜(chloroplast envelope): (1)组成: 外膜与内膜、膜间隙 (2)主要成分:蛋白质和脂质 (3)叶绿体膜中的酶:ATP酶、腺苷酸激酶以及参与糖脂合 成与代谢有关的酶。 (4)功能:控制代谢物进出叶绿体的功能 外膜通透性大 内膜对物质通过的选择性强 内外膜共同构成细胞质和叶绿体基质间的功能性屏障
2、类囊体(thylakoid): (1)定义:在叶绿体基质中由许多由单位膜封闭形成的扁平 小囊,类囊体一般沿叶绿体长轴平行排列 (2)基粒(grana):在某些部位,许多圆饼状的类囊体叠置 成垛称为基粒,组成基粒的类囊体称基粒类囊体 (3)垛叠成基粒的意义:大大增加了膜片层的总面积, 更有效地捕获光能,加速光反应。 (4) 基粒经基质类囊体(贯穿在两个或两个以上基粒之间, 没有发生垛叠的类囊体称为基质类囊体)相连 使类囊体彼此相通,使膜系统的膜囊与基质隔开,有利于光和 磷酸化过程中H+梯度的形成
组成:F1头部和嵌于膜内的F0基部 F1头部:3α、3β、1γ、1δ、1ε 3α和3β交替排列形成一个橘瓣状 结构 ,β亚基具有催化ATP合成 功能
6-线粒体和叶绿体PPT课件

-
44
-
45
2)类囊体膜的化学组成
主要成分:蛋白质和脂质(比例约60:40)。 • 1.脂质
主要是磷脂和糖脂及色素、醌化合物等 不饱和的亚麻酸约87%,流动性大 • 2.蛋白质 (1)外在蛋白:CF1 、与光反应有关的酶 (2)内在蛋白:20余种多肽
-
46
3.叶绿体基质(stroma)
• 1)核酮糖-1,5-二磷酸羧化酶RuBPase • 2)环状DNA • 3)RNA:rRNA、tRNA、mRNA • 4)核糖体(蛋白质合成体系) • 5)脂滴(Lipiddroplet)或称嗜锇滴 • 6)植物铁蛋白、淀粉粒等
形成“转子”; 2)嵌入膜中的F0(基
部),组成“定子”;
-
25
F1:3:3:1:1:1
具3个ATP合成的催 化位点
F0: 1a:2b:12c
环形结构,具质 子通道
-
26
❖ F1因子具有ATP酶活性
-
27
b)ATP合成酶性质:是一种具有双向作用的装置
-
28
c)ATP合成酶的作用机制
Step 1: Proton gradient is built up as a result of NADH (produced from oxidation reactions) feeding electrons into electron transport system.
• 3由)1Fa、0偶1联b、因1子0-1:2c是亚内基膜组上成的,疏多水个蛋c亚白基复形合成体多,(形1成2)跨聚膜体质,子成通一道。 环 合状于结c亚构基,环和状转结子构结的合外。侧a,、并b亚通基过及δ亚F1的基δ和亚头基部形相成结“合定。子”,结
第六章 线粒体和叶绿体

质子驱动力
第二节 叶绿体与光合作用
叶绿体的结构 透镜形,长轴约为4~10μm,短轴为2~5μm。比 线粒体大2 ~ 4倍。 构成:外被、层膜系统和基质 叶绿体外被 (外膜,间隙和内膜) 外膜:透性很高; 内膜:透性很低,对物质选择性比外膜强。细胞质和 叶绿体基质间的功能屏障。内膜有特殊载体——转运 体(translocator),可以转运代谢物。 膜间隙:约10—20 nm;一起形成叶绿体外被 (chloroplast envelope)。
线粒体的融合与分裂
线粒体的融合是与分裂 协同进行的,过程高度 保守,需要在多种蛋白 质的精确调控下完成。 两者一般保持动态平衡, 这种平衡对维持线粒体 正常的形态、分布和功 能十分重要。在植物细 胞中,可以观察到线粒 体融合与分裂的偶联现 象,即颗粒状和短线状 的线粒体融合后随即发 生分裂。
线粒体融合与分裂的的分子及细胞生物学基础
第六章 线粒体和叶绿体
本章讲述线粒体和叶绿体的基本结构与功能,同
时对这两种细胞器在细胞中的行为及调控机制进行
初步的讨论。
线粒体发现历史
1890年,德国生物学家Altmann首先在光学 显微镜下观察到动物细胞内存在一种颗粒 状结构,取名为生命小体。 1897年,Benda将之命名为线粒体。 在植物细胞中,Meves于1904年首次发现了 线粒体,从而确认线粒体是普遍存在于真 核细胞内的重要细胞器。
第三节 线粒体和叶绿体的半自主性 及其起源
线粒体和叶绿体的遗传信息系统被称为真核细胞的第二遗传 信息系统,或核外基因及其表达体系。因为研究发现,线粒体 和叶绿体中除有DNA外,还有RNA(mRNA、tRNA、 rRNA)、核糖体、氨基酸活化酶等。说明这两种细胞器都 具有独立进行转录和转译的功能。也就是说,线粒体和叶绿 体都具有自身转录RNA和翻译蛋白质的体系。但迄今为止, 人们发现叶绿体仅能合成13种蛋白质,线粒体能够合成的蛋 白质也只有60多种,而参与组成线粒体和叶绿体的蛋白质却 分别有上千种。这说明,线粒体和叶绿体中自身编码合成的 蛋白质并不多,它们中的叶绿体的自主 程度是有限的,它们对核遗传系统有很大的依赖性。因此,线 粒体和叶绿体的生长和增殖是受核基因组及自身的基因组两 套遗传信息系统控制的,所以它们都被称为半自主性细胞器。
细胞生物学第六章线粒体与叶绿体
ATP合酶的“结合变构模 型
三、线粒体与疾病
1、线粒体病:属母系遗传,如克山病、Leber遗 传性视神经病、肌阵挛性癫痫 2、线粒体与衰老 数目与体积的变化 mtDNA的损伤、缺失 线粒体是细胞内自由基的主要来源 3、线粒体与细胞凋亡
第二节 叶绿体与光合作用
一、叶绿体的形态、大小和数目 不同植物种类之间有很大差异。 一般形态为香蕉形,或者称为透镜形。宽2-4um, 长5-10um。叶肉细胞含50-200个叶绿体。 二、叶绿体的结构和化学组成 (一)叶绿体被膜 双层单位膜,6-8nm,之间为10-20nm的膜间 隙。外膜有孔蛋白,通透性大。
THE ESTABLISHMENT OF A PROTONMOTIVE FORCE
33
(四)ATP形成机制——氧化磷酸化
氧化磷酸化:在呼吸链上与电子传递相耦联的由 ADP被磷酸化生成ATP的酶促过程。 呼吸链上有三个部位是氧化还原放能与ADP磷酸 化生成ATP偶联的部位。这三个部位有较大的自 由能变化,足以使ADP与无机磷结合形成ATP。 部位Ⅰ在NADH至CoQ之间。部位Ⅱ在细胞色素 b和细胞色素c之间。部位Ⅲ在细胞色素a和氧之 间。
第二节 叶绿体与光合作用
内膜通透性低。 内膜上有特殊的转运载体,如磷酸交换载体、二 羧酸交换载体,都属于被动运输。 内膜上有与糖脂、磷脂合成有关的酶类。因此, 叶绿体被膜不仅是叶绿体脂合成的场所,也是整 个植物细胞脂合成的主要场所。
(二)类囊体 1、类囊体的结构 叶绿体基质中由单位膜包围形成的扁平小 囊称为类囊体。 基粒、基粒类囊体、基质类囊体。 一个叶绿体的全部类囊体是一个完整连续 的封闭膜囊。含有丰富的具半乳糖的糖脂。 流动性很大。类囊体膜上的成分有:PSI、 PSII、细胞色素bf、CF0-CF1ATP酶。其分 布不对称。
细胞生物学 名词解释 第六章 线粒体和叶绿体

光合磷酸化数据
2H2O光解生成1分子O2和4H+,PSI和PSII分别吸收4个光子(共8个光子)传递4e-,造成12H+(水光解产生4H+,PQ释放4H+,PSI消耗基质中4H+)=3ATP,生成2NADPH
电子传递链
electron transport chain
膜上一系列由电子载体组成的电子传递途径。这些电子载体接受高能电子,并在传递过程中逐步降低电子的能量,最终将释放的能量用于合成ATP或以其他能量形式储存。
捕光复合体Ⅱ
light harvesting complex Ⅱ,LHCⅡ
位于光系统Ⅰ之外的色素蛋白复合,含有大量天线色素为光系统Ⅱ(PSⅡ)收集光子。
ATP合酶
ATP synthase
位于线粒体内膜或叶绿体的类囊体膜上,通过氧化磷酸化或光合磷酸化催化ADP和无机磷合成ATP的酶,由F1头部和嵌入膜内的F0基部组成,也常见于细菌膜上。
F1头部组成
α3β3γεδ,α3β3形成橘瓣,β亚基的结合位点具有催化ATP合成或水解的活性,γε形成转子,调节3个β亚基催化位点的开放和关闭;δ亚基为F1和Fo相连接所必须的。
电子从PS I开始经过Fd后传给Cytb6f,再经PC回到PS I,由PSI单独完成,电子传递呈现闭合的回路,只产生ATP,不产生NADPH和O2。
作用:当植物缺乏NADP+时,启动循环光合磷酸化,以调节ATP与NADPH的比例,适应碳同化对ATP与NADPH的比例需求(3:2)
第六章 线粒体与叶绿体

●ATP水解酶的活性
●ATP合成酶的活性
故 F0-F1 颗 粒 又 称 为 ATP 合 酶 (ATP synthase )。
Structure of ATP synthase
◆结构
● Head section: 即 F1, 由 5 种多肽组成九聚 体 , 含有 3 个催化 ATP 合成的位点 , 每个β亚 基一个。 ● Stalk section: 由 F1 的γ亚基和ε亚基构 成,γ亚基穿过头部作为头部旋转的轴。 ● Membrane section: 即 F0, 由 3 种不同的亚 基组成的十五聚体(1a∶2b∶12c)。
一、线粒体和叶绿体的DNA
●mtDNA /ctDNA形状、数量、大小
●mtDNA和ctDNA均以半保留方式进行自我复制
●mtDNA复制的时间主要在细胞周期的S期及G2期,
DNA 先复制,随后线粒体分裂。 ctDNA 复制的时
间在G1期。 复制仍受核控制
mtDNA /ctDNA形状、数量、大小
◆双链环状(除绿藻mtDNA,草履虫mtDNA) ◆mtDNA 大小在动物中变化不大,但在植物中变化 较大高等植物,120kbp~200kbp; ◆ 人 mtDNA : 16,569bp , 37 个 基 因 ( 编 码 12S,16S rRNA ; 22 种 tRNA ; 13 种多肽: NADH 脱氢酶 7 个
性等特点. 线粒体的形状多种多样,但以线状和粒状最常见.
线粒体在不同类型细胞中,其数目相差很大,但在同一类型细胞中的数目
是相对比较 稳定的.
●线粒体的超微结构 ◆外膜(outer membrane):含孔蛋白(porin), 通透性较高。标志酶是
单胺氧化酶
◆内膜(inner membrane):高度不通透性,向内折 叠形成嵴。 含有与能量转换相关的蛋白.标志酶是细胞色素氧化酶.
细胞生物学第六章 细胞的能量转换-线粒体和叶绿体

线粒体的形态结构
线粒体的形态与分布
动态细胞器 外形:线状、粒状、哑铃状、环状、圆柱状等 大小:直径0.5-1.0μm,长为2-3μm,可达1020μm 数目:动物细胞植物细胞 人成熟红细胞中无线粒体 代谢旺盛的细胞 分布:细胞功能旺盛区域;定位与迁移与微管相关
线粒体可塑性
心肌
状如蘑菇,属F型质子泵。 分为球形的F1(头部)和嵌入膜中的F0(基部)。 F1由5种多肽组成α3β3γδε复合体,具有三个ATP合成 的催化位点(每个β亚基具有一个)。 F0由三种多肽组成ab2c12复合体,嵌入内膜,12个c亚基组 成一个环形结构,具有质子通道。
F1 particle is the catalytic subunit; The F0 particle attaches to F1 and is embedded in the inner membrane. F1: 5 subunits in the ratio 3:3:1:1:1 F0: 1a:2b:12c
2、复合物II:琥珀酸脱氢酶
组成:至少由4条肽链,含有一个FAD,2个铁硫蛋白。 作用:催化琥珀酸的低能量电子转至辅酶Q,但不转移质子。
琥珀酸→FAD→Fe-S→Q。 琥珀酸+Q→延胡索酸+QH2
3、复合物III:细胞色素c还原酶。 组成:至少11条不同肽链,以二聚体形式存在,每个单体包 含两个细胞色素b(b562、b566)、一个铁硫蛋白和一个 细胞色素c1 。 作用:催化电子从辅酶Q传给细胞色素c,每转移一对电子, 同时将4个质子由线粒体基质泵至膜间隙。 2还原态cyt c1 + QH2 + 2 H+M→2氧化态cyt c1 + Q+ 4H+C
细胞生物学第六章 线粒体与叶绿体

线粒体的增殖
分裂(隔膜分离、收缩分离)
出芽
叶绿体的发育和增殖
前质体→质体→
光照:叶绿体 黑暗 白色体 黑暗:白色体 光照 叶绿体
有色体:含非光合作用色素、DNA
增殖:分裂增殖
五、线粒体和叶绿体的起源
内共生起源学说 线粒体和叶绿体分别起源于原始真核细胞内共 生的行有氧呼吸的细菌和行光能自养的蓝细菌。
1. 叶绿体的形态结构
叶绿体膜:双层膜(外膜和内膜) 外膜通透性大,MW<10000的物质可通过; 内膜选择性强,具有特殊载体,仅允许 O2、CO2和H2O分子自由通透。
类囊体:由单位膜封闭形成的扁平小囊 基粒类囊体:叠垛 基质类囊体:连通相邻的基粒类囊体 类囊体腔:彼此相通 类囊体膜:含叶绿素,绿色植物进行光合作用 的场所
该病为母系遗传,家系内女性患者的子女均可能患病,男 性患者的子女均不患病。
二、质体
质体 植物细胞,双层膜; 颗粒状、杆菌状、线条状; 含遗传信息和蛋白质合成系统; 分裂增殖
类型
1. 叶绿体:含类囊体、叶绿素,能进行光合作用。
2. 白色体:含未发育的类囊体,不含叶绿素和其他色 素,不能进行光合作用;呈白色,存在于根和芽的 幼嫩细胞中。
2. 线粒体的超微结构
外膜:单位膜,平整光滑,有弹性,上有排列 整齐的筒状圆柱体,成分为孔蛋白,孔径 2~3nm,MW<1000的小分子物质可自由通过。
内膜:内突为嵴,嵴上有大量基粒(ATP合成 酶)(负染方法可见),标志酶细胞色素氧化 酶。氧化磷酸化的关键场所。
嵴:板层状,管状,其形态、数量、排列与细胞 种类及生理状态有关。
通过以上复合物I、III、IV时都有一个大的氧还电位变化, 认为是质子泵的部位。
细胞生物学-6第六章线粒体和叶绿体

第二节 叶绿体与光合作用
●叶绿体(Chloroplast)的形态结构 ●叶绿体的功能—光合作用(photosynthesis)
一、叶绿体(Chloroplast)的形态结构
●叶绿体与线粒体形态结构比较 叶绿体内膜并不向内折叠成嵴;内膜不含电
子传递链;除了膜间隙、基质外,还有类囊体; 捕光系统、电子传递链和ATP合成酶都位于类囊体 膜上。 ●叶绿体超微结构
●线粒体和叶绿体是半自主性细胞器 (Semiautonomous organelle) ●线粒体和叶绿体的增殖与起源 (proliferation and genesis)
第一节 线粒体与氧化磷酸化
●线粒体的形态结构 ●线粒体的化学组成及酶的定位 ●氧化磷酸化 ●线粒体与疾病
一、线粒体的形态结构
一、线粒体和叶绿体的DNA
●mtDNA /ctDNA形状、数量、大小
●mtDNA和ctDNA均以半保留方式进行自我复制
●mtDNA复制的时间主要在细胞周期的S期及G2期, DNA先复制,随后线粒体分裂。ctDNA复制的时 间在G1期。 复制仍受核控制
mtDNA /ctDNA形状、数量、大小
◆双链环状(除绿藻mtDNA,草履虫mtDNA) ◆mtDNA大小在动物中变化不大,但在植物中变化较大高等
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
线粒体和叶绿体第一节线粒体与氧化磷酸化一、线粒体形态、大小、数目和分布二、线粒体的超微结构本世纪50年代后,在电镜下观察研究线粒体的结构问题。
是由双层单位膜套叠成的所谓“囊中之囊”,在空间结构上人为地划分为四大部分,即外膜、内膜、外室、内室。
(一)外膜指包围在线粒体最外面的一层膜,看上去平整光滑而具有弹性,膜厚约6nm。
对各种小分子物质(分子量在10000 doldon以内,如电解质、水、蔗糖等)的通透性较高,有人认为外膜上具有小孔(ф2~3nm)。
(二)内膜也是一单位膜,约厚6~8nm。
内膜不同于外膜。
首先是在结构上,内膜不是平滑的,而是由许多向线粒体腔内的突起(褶叠或小管),被称为“线粒体嵴”,是线粒体最富有标志性的结构,它的存在大大扩大了内膜的表面积,增加了内膜的代谢效率。
(三)外室(outer space)(膜间隙)指内、外膜之间的窄小空隙,宽约6~8 nm,又称膜间隙。
(四)内室指由内膜包围的空间,其内充满蛋白质性质的物质,称线粒体基质。
三、线粒体的化学组成及定位(一)蛋白质外膜含量(60%)低于内膜含量(80%),主要为酶类(约120余种)。
外膜:单胺氧化酶(标记酶)、NADH—细胞色素C还原酶、脂肪酸辅酶A连接酶等等;内膜:呼吸链酶系(细胞色素氧化酶为标记酶)、ATP合成酶、琥珀酸脱H酶等等;外室:腺苷酸激酶(标记酶)、核苷二磷酸激酶;内室:三羧酸循环酶系(其中苹果酸脱H酶是标记酶)、脂肪酸氧化酶、蛋白质合成酶系等等(二)脂类外膜中含量(40%)高于内膜中的含量(20%)。
其中内膜不含胆固醇,而含心磷脂较多。
(三)核酸基质中有DNA,称mt—DNA四、线粒体的功能——生物氧化亦称细胞呼吸(cellular respiration),指各类有机物质在细胞内进行氧化分解,最终产生CO2和H2O,同时释放能量(ATP)的过程。
包括TCA环、电子传递和氧化磷酸化三个步骤,分别是在线粒体的不同部位进行的。
(一)生物氧化的分区和定位(二)电子传递和氧化磷酸化的结构基础虽然电子传递和氧化磷酸化偶连在一起,但它们又是通过不同的结构完成的。
1968年,E.Racker等的亚线粒体小泡重建实验说明了这一问题(图示)。
由此可见,电子传递是在线粒体内膜上,氧化磷酸化由基粒承担。
1 电子传递链(呼吸链)呼吸链是由存在于线粒体内膜上的众多酶系和其它分子组成的电子传递链。
(1)复合物I NADH—Q还原酶,催化NADH的2个电子→辅酶Q(2)复合物Ⅱ琥珀酸—Q还原酶,催化电子从琥珀酸通过FAD和铁硫蛋白传至辅酶Q(3)复合物Ⅲ细胞色素还原酶,催化电子从辅酶Q传至CytC(4)复合物Ⅳ细胞色素氧化酶,将电子从CytC→氧。
2 基粒(F1—FO复合物)的超微结构F1—FO复合物,又称内膜亚单位、呼吸集合体、ATP酶复合物、ATP合成酶等。
这一结构最初是在1962年,由Fernadezmoran经负染色在电镜下观察到的,后来D.Green 将其称为线粒体基粒,后改称基粒,实际上是一种ATP酶复合体,分子量约在448000。
它是由多条多肽链构成的复合结构,可分为三部分,即头、柄、膜三部。
在ATP形成过程中共同发挥作用。
3 氧化磷酸化的偶联机制(1)化学偶联假说(Chemieal coupling hypothesis)(2)构象偶联假说(Conformational coupling hypothesis)(3)化学渗透学说(Chemiosmotic coupling hypothesis)亦称电化学偶联学说,是1961年英国生化学家P.Mitchell提出的。
对电子传递和氧化磷酸化问题作了较为另人信服的解释,故普遍为人接受,米切尔因此而获1978年诺贝尔化学奖。
这一假说的中心思想是:在电子传递过程中所释放的能量转化成了跨膜的氢离子浓度梯度的势能,这种势能驱动氧化磷酸化反应,合成ATP。
(1)NADH提供一对电子,经电子传递链,最后为O2所接受。
(2)电子传递链中的载氢体和电子传递体相间排列,每当电子由载氢体传向电子传递体时,载氢体的H+便释放到内膜外。
一对电子在呼吸链三次穿膜运动,向外室排放三对H+ 。
(3)内膜对H+具有不可透性,故随电子传递过程的不断进行,H+ 在外室中积累,造成膜两侧的质子浓度差。
(4)外室中H+有顺浓度梯度返回基质的倾向,当H+通过F1—FO复合物时,ATP酶利用这一势能合成ATP。
(5)F1—FO复合物需2个质子合成一个ATP。
第二节叶绿体与光合作用叶绿体是植物细胞特有的双层膜围成的细胞器,它对生物界的存在和进化有着重大贡献(三个最初:一是人类、动物、多数微生物的食物的最初来源;二是人类社会利用的古生物燃料——煤、石油、天然气的最初来源;三是地球上氧气的最初来源),主要功能在于:吸收光能,合成碳水化合物,同时产生分子氧,总称为光合作用一、叶绿体的形状、大小、数目、分布二、超微结构近年来,先后有许多学者采用超薄切片、负染色和冰冻蚀刻等先进技术,研究叶绿体的形态和组成,揭示叶绿体囊状膜系统的超微结构。
1 叶绿体膜是两层光滑的单位膜(内、外膜)6-8nm,也称外被,是一个有选择的屏障,控制着叶绿体代谢物质的进入和排出。
2 基质指叶绿体膜包围的,无结构,呈流动状态的物质。
即叶绿体内膜与类囊体之间无定形物质,在基质中存在:(1)叶绿体DNA 环状,每一叶绿体内可含有几十个拷贝;(2)70S核糖体;(3)mRNA、tRNA;(4)酶类;(5)RUBP羧化酶;(6)各种离子。
3 类囊体类囊体在基质中有两种形式存在,一种是较小的扁囊,多个5—30(10—100个)相互叠置成一摞,形成的结构称基粒。
每一叶绿体中约含有40—80个基粒。
组成基粒的类囊体称基粒类囊体或基粒片层。
另一种是较大的扁囊,贯穿于基粒之间,称基粒间类囊体或基质类囊体或基质片层。
它们顺着叶绿体的纵轴彼此平行排列。
其存在意义在于,使膜片层的总面积大大超出叶绿体的面积。
可见基粒中有PSI和PSII的机能单位,并分布在膜内表面,是PSII核心颗粒和捕光复合物结合成的。
而基质中多有PSI的机能单位,多布于膜外侧。
除上述内在蛋白外,还有组成电子传递链的众多载体,包括○1PQ(质体醌)、○2PC (质体兰素,、○3细胞素(Cytb—559,Cytf—553,Cytb6—563等)、○4铁硫蛋白(铁氧还蛋白ferrdoxin,Fd)、○5黄素蛋白。
故将类囊体称为光合膜。
三、化学组成四、叶绿体的功能——光合作用绿色植物细胞,吸收光能,还原CO2,并利用水提供氢合成碳水化合物,同时放出分子氧的过程,称为光合作用。
总过程分为两个阶段:光反应和暗反应。
(一)光反应叶绿素等色素分子捕获光能,将光能转化为ATP和NADPH的化学能,并放出氧的过程,是在类囊体膜上进行的,为能量转换过程。
光反应包括三个基本反应:原初反应、电子传递反应、光合磷酸化。
(1)原初反应:指聚光色素分子吸收光量子传到反应中心进行光化学反应的物理过程。
包括光能的吸收、传递与转换。
(2)电子传递反应:包括三个阶段:NADP+的还原反应;PSII与PSI之间的传递;放氧反应。
(3)光合磷酸化反应:在有光存在下,当电子沿电子传递链传递时,形成ATP的过程称为光合磷酸化。
当电子从还原势高处(Q)向还原势低的PSI传递时,能量下降,利用这一能量将ADP 磷酸化形成ATP,这一过程称非循环式光合磷酸化。
当NADPH NADP+比值大时(缺少NADP+时),铁氧还蛋白(Fd)则将电子通过cytb6、cytf、pc传给P700+,利用这一能量使ADP磷酸化形成ATP,称循环式光合磷酸化(电子通路是闭合的)。
(4)光合磷酸化机制在一对电子的传递过程中,膜外消耗了三个质子,膜内则增加了四个质子,随着过程的不断进行,膜内外便建立了质子梯度,有向膜外穿出的趋势,当每3对H+通过CF1-FO复合物时,在CF1的催化下,合成一个ATP。
(二)暗反应利用光反应产生的ATP和NADPH还原CO2形成碳水化合物,将活跃化学能变为稳定化学能,是在叶绿体基质中进行的。
为物质代谢过程。
在高等植物固定CO2有三条途径:卡尔文循环(C3途径)、C4途径(Hatch-slack途径)和景天科酸代谢。
卡尔文循环是最基本、最普通的,只有这一途径具备合成淀粉之能力,又称C3途径。
第三节线粒体和叶绿体是半自主性细胞器一、线粒体与叶绿体的DNA(一)线粒体DNA (mt-DNA)(二)叶绿体DNA(ct-DNA)二、线粒体和叶绿体的蛋白质合成(一)线粒体的蛋白质合成线粒体基质中除有DNA外,还有各种RNA、核糖体、氨基酸活化酶等,说明它能合成自我繁殖所需的某些成分,但数量不多,只占线粒体全部蛋白质的10%,约有13种(20个分子)左右。
有人估算:×10(每周10个核苷酸)=14705对核苷酸,能编码4902个氨基酸(除以3),假设一个蛋白质分子由150个氨基酸组成,则能编码30个左右蛋白质分子,如果除去编码mRNA、rRNA、tRNA的信息量(占总信息量的30%),余下的信息量只能编码约20个左右的蛋白质分子。
综上所述,线粒体有自身的DNA,有一整套蛋白质合成系统,能够复制和再生,使其一代代传下去,所以具有一定的自主性。
(二)叶绿体蛋白质的合成叶绿体中的蛋白质(酶)一部分是在叶绿体中由它自己的DNA编码,经过mRNA转录和翻译形成的,有一部分则是由核基因编码,在细胞质中形成后转入叶绿体的。
还有一部分是由核基因编码,在叶绿体的核糖体上合成。
三、对细胞核和细胞质的依赖性大无论是线粒体还是叶绿体,它们的自主性是有限的,下面以线粒体为例说明之。
核质蛋白质合成系统通过合成某些酶类来调节线粒体的蛋白质合成系统。
在有氯霉素存在的条件下培养链孢霉细胞,这时线粒体的蛋白质合成受抑制,但线粒体的三羧酸循环酶类、电子传递链中的NADH脱氢酶、CytC、以及DNA-Poly-merase、RNA-Polymerase、核糖体蛋白质、各种氨基酸活化酶等有关线粒体DNA复制和基因表达的酶类依然存在。
而这些酶是由核基因编码,在细胞质中合成,然后转移到线粒体的。
这说明细胞质的蛋白质合成系统(或者说核——质蛋白质合成系统)通过合成某些酶类来调节线粒体的蛋白质合成系统。
又:在有放线菌酮存在下培养链孢霉细胞,由于细胞质蛋白质合成系统受抑制,结果培养一段时间后,线粒体的合成活性也显著下降。
这足以说明线粒体对细胞核和其他细胞质部分有很大依赖性。
实际上也是这样,线粒体DNA所编码的蛋白质只有它自身全部蛋白质的10%,绝大部分是由核DNA编码的。
从上述看出,线粒体的生长增殖是受核基因组和线粒体基因组两套遗传系统的共同控制,故称线粒体为半自主性细胞器。