分子生物学第六章
第六章:常用分子生物学技术——RNA干扰技术

识别并切割(qiēgē)与siRNA反义链互补的靶mRNA
(5)dsRNA的再生成
第十ānrǎo)
第十七页,共四十页。
RNA干扰(gānrǎo)
(RNA interference, RNAi)
siRNA(小干扰RNA):21-23nt,由dsRNA裂 解而成的小片段,可诱导mRNA降解。siRNA主要
药学 分子生物 (yào xué)
学
第一页,共四十页。
生物芯片 (biochip) (shēnɡ wù xīn piàn)
以生物、电子、机械和信息技术为基础,在 固相支持物表面建立集成、连续、微型分析系 统(xìtǒng),形成芯片,实现对生物大分子的准确、 快速、高通量、自动化检测。
第二页,共四十页。
第三十一页,共四十页。
REGγ基因敲除鼠(下图)出现脊椎(jǐ zhuī)弯曲等早衰现象
第三十二页,共四十页。
Gab1基因(jīyīn)敲除导致小鼠缺血性血管新生、侧枝循环建立出现 缺陷(图示上半部分),主要是由于血管内皮细胞管状结构形成的 信号调控通路出现障碍而引起的(图示下半部分)。
第三十三页,共四十页。
2、 RNA干扰技术缺点
“脱靶效应”(off-target effects)
第二十四页,共四十页。
RNA干扰技术(jìshù)的应用
1、基因功能研究(功能失活策略) “基因敲减(knockdown)”
2、基因治疗(基因(jīyīn)失活性治疗) (1)病毒感染性疾病
通过RNAi抑制 RNA病毒(bìngdú)复制 (2)基因过表达引起的疾病(如肿瘤)
特定功能基因的部分片段被同源的外源DNA片 段替代,从而使靶基因失活。
现代分子生物学第六章作业

现代分子生物学第六章作业09级一班芮世杭2220093170110271,列举两种研究基因表达模式的方法并简述其原理。
(1)基因表达序列分析技术(SAGE)是一种以DNA序列测定为基础定量分析全基因组表达模式的技术能够直接读出任何一种细胞类型或组织的基因表达信息在转录组水平上,任何长度超过9—10个碱基的核苷酸片段都可能代表一种特异性核苷酸的转录产物,因此,用特定限制性核酸内切酶分离转录产物中具有基因特异性的9—10个碱基的核苷酸序列并制成标签。
将这些序列标签连接,克隆,测序后,根据其占总标签数的比例即可分析其对应编码基因的表达频率。
(2)原位杂交技术(ISH)是用标记的核酸探针,经放射自显影或非放射检测体系,在组织,细胞,间期核及染色体上对核酸进行定位和相对定量研究的一种手段,分为RNA和染色体原位杂交两大类。
RNA原位杂交用放射性或非放射性标记的特异性探针与被固定的组织切片反应。
若细胞中存在与探针互补的mRNA分子,两者杂交产生双链RNA,课通过反射性标记或经酶促免疫显色,对该基因的表达产物做出定性定量分析。
(3)基因芯片技术(FISH)对寡核苷酸探针做特殊的修饰和标记,用原位杂交与靶染色体或DNA上特定的序列结合,再通过与荧光素分子相耦联的单克隆抗体来确定该DNA序列在染色体上的位置。
2,简述基因芯片技术对分子生物学研究的意义。
解某些基因对特定生长发育阶段的重要性;基因芯片还可用于进行基因诊断,可建立正常人特定组织、器官的基因芯片,给出标准杂交信号图。
用可疑病人的cDNA做探针与之杂交,检查哪些基因的表达受抑制或激活,另可研究表达基因的生物学特性。
3,比较酵母双杂交技术和免疫共沉淀技术在研究蛋白质相互作用方面的优缺点?(1)酵母双杂交技术称Two-hybrid system也叫interaction trap(相互作用陷井),是90年代初发展起来的分离基因的新方法,可用于分离能与已知靶蛋白质(target protein)相互作用的基因。
第六章 基因调控1:原核生物基因的表达调控 分子生物学习题

第六章基因表达调控1:原核生物基因表达调控名词解释:操纵子基因表达持家基因正调控和负调控安慰诱导物衰减子(弱化子)魔斑结构基因和调节基因本底水平表达填空1 操纵子的基因表达调节系统属于水平的调节,乳糖操纵子模型由和1961年提出的。
色氨酸操纵子包括和两方面的调控。
2、能够诱导操纵子但不是代谢底物的化合物称为诱导物。
能够诱导乳糖操纵子的化合物就是其中一例。
这种化合物同蛋白质结合,并使之与分离。
乳糖操纵子的体内功能性诱导物是。
3、色氨酸是一种调节分子,被视为。
它与一种蛋白质结合形成。
通过控制起作用。
色氨酸操纵子受另一种系统------ 的调控,它涉及到第一个结构基因被转录前的转录。
4、大肠杆菌乳糖操纵子调节基因编码的与结合,对Lac结合,对Lac表达实施负调控;与复合物结合于上游部分,对Lac表达实施正调控。
5、操纵子中没有基因产物的是和选择题1、下面哪些真正是乳糖操纵子的诱导物?()A.乳糖B.蜜二糖C.O- 硝基苯酚-β-半乳糖苷(ONPG)D.异丙基-β-半乳糖苷E. 异乳糖2、色氨酸操纵子的调控作用是受两个相互独立的系统控制的,其中一个需要前导肽的翻译,下面哪一种调控这个系统?()A.色氨酸B.色氨酰-tRNA TrpC.色氨酰-tRNAD.cAMPE.以上都不正确3、阻遏蛋白(阻抑蛋白)识别操纵子中的()A 启动基因B 结构基因C 操纵基因D 内含子E 外显子4、乳糖、阿拉伯糖、色氨酸等小分子物质在基因表达调控中作用的共同特点是A 与启动子结合B 与DNA结合影响模板活性C 与RNA聚合酶结合影响其活性D 与蛋白质结合影响该蛋白质结合DNAE 与操纵基因结合5.下面那项不属于原核生物操纵元的结构A:启动子B:终止子C:操纵子D:内含子6、下列有关操纵子的论述哪个是错误的()A 操纵子是由启动基因、操纵基因与其所控制的一组功能上相关的结构基因组成的基因表达调控单位B 操纵子不包括调节基因C 代谢底物往往是该途径的可诱导酶的诱导物,代谢终产物往往是可阻遏酶的辅阻遏物D 真核细胞的酶合成也存在诱导和阻遏现象,因此也是由操纵子进行调控的7、操纵子调节系统属于哪一种水平的调节?A 复制水平的调控B 转录水平调控C 转录后加工的调控D 翻译水平的调控8、对调节基因下述哪些论述哪些是对的()A 是编码阻遏蛋白的结构基因B 各种操纵子的调节基因都与启动基因相毗邻C 调节基因是操纵子的组成部分D 调节基因的表达另有转移的调控区9、以下有关阻遏蛋白的哪些是对的()A 阻遏蛋白是调节基因表的的产物B 可诱导操纵子的阻遏蛋白具有直接与操纵子基因结合的活性,与诱导物相互作用后丧失活性C 可阻遏操纵子的阻遏蛋白没有直接与操纵子基因结合的活性,与辅阻遏物结合后才有此活性D 阻遏蛋白可与RNA聚合酶竞争同一结合部位10、关于启动基因的下述论点哪些是错误的()A 启动基因是RNA聚合酶识别并最县结合的一段DNA序列B 启动基因是最先被RNA聚合酶转录的DNA 序列C 启动基因是DNA上富含A-T碱基对的部分D 启动基因是引发DNA复制的特殊序列11、下列有关降解物基因活化蛋白(CAP)的哪些论点是正确的()A CAP-cAMP可专一地与启动基因结合,促进结构基因的转录B CAP可单独与启动子相互作用,促进转录C CAP-cAMP可与调节基因结合,控制阻遏蛋白的合成D CAP-cAMP可与RNA聚合酶竞争地结合于启动基因,从而阻碍结构基因的转录12、与乳糖操纵子操纵基因结合的物质是()A RNA聚合酶B DNA聚合酶C 阻遏蛋白D 反密码子是非题1、葡萄糖和乳糖并存时,细菌优先利用乳糖并启动乳糖操纵子()2、小分子物质如ITPG诱导乳糖操纵子表达时起负调控作用与操纵基因相结合阻抑结构基因的表达()3、色氨酸操纵子中含有衰减子区,其调控作用主要受Trp浓度高低影响()4、色氨酸操纵子(trpoperon)中含有衰减子序列()5、cAMP在laz 操纵子中起正调控作用,其浓度受环境中的葡萄糖影响,与其浓度成正比()6、大肠杆菌乳糖操纵子真正的诱导物不是乳糖,而是它的异构体别乳糖()7、操纵基因又称操纵子,如同启动基因又称启动子一样()8、可诱导操纵子是负责调节糖分解代谢的,可阻遏操纵子是负责调节氨基酸代谢的()问答题:1、试述乳糖操纵子的结构及负控诱导的调控机理2、色氨酸操纵子的结构特点?其弱化子在调控中如何起作用?3、简述色氨酸操纵子中启动子调控作用特点。
第六章1分子生物学改造

一、定点突变
利用分子生物学技术,在体外通过碱基取代、插入 或缺失可以使基因DNA序列中任何一个特定的碱基 发生改变。这种体外特异性改变某个碱基的技术, 称谓定点突变(site directed mutagenesis)。
定点突变具有简单易行、重复性高等优点,现已发 展成为基因操作的一种技术。这种技术不仅适用于 基因结构与功能的研究,还可通过改变基因的密码 子来改造天然蛋白质。
(%) 100 100
96 106
0 95 0
0
(℃) 41.9 41.9 46.7 48.3 52.9 57.6 58.9 65.9
wtα:野生型T4溶菌酶;pwt:假野生型酶;A-F:六种设计的半胱氨酸 变体;Tm:熔点温度
将Asn和Gln转换成其他氨基酸
当蛋白质暴露于高温时:
天冬酰胺(Asn)
而将2个Asn分别换为苏氨酸(Thr)和异亮氨酸(Ile),其半衰 期则延长。
酵母菌磷酸丙糖异构酶及其变体的热稳定性
氨基酸位点
酶
14
78
半衰期(min)
野生型 Asn
Asn
13
变体A Asn
Thr
17
变体B Asn
Ile
16
变体C Thr
Ile
25
变体D Asp
Asn
天冬氨酸(Asp) + NH3
谷氨酰胺(Gln)
谷氨酸(Glu) + NH3
导致肽链折叠的局部的改变,可能影响其活性。
如酵母的丙糖磷酸异构酶是由两个相同的亚基组成的二聚体, 每个亚基都含有两个Asn残基,均位于两个亚基相互接触的 表面上,可能与该酶的热稳定性有关。
分子生物学第5章、第6章

•DNA分子内或分子间发生遗传信息的重新组合,称为 遗传重组,或基因重排。→ 重组DNA •真核生物基因组间重组多发生在减数分裂时同源染 色体之间的交换;细菌及噬菌体的基因组为单倍体, 来自不同亲代两组DNA之间可通过多种形式进行遗传 重组。 •DNA重组对生物进化起着关键的作用。 •重组分类:同源重组(homologous recombination) 、 位点特异性重组(site-specific recombination)、 转座重组(transposition recombination)和 异常重组(illegitimate recombination)。
1. 互变异构体:碱基发生烯醇式-酮式互变异构或者氨 基-亚氨基互变异构时,使碱基错配。 2. 脱氨基作用:碱基上氨基自发脱落,或在诱变剂的 作用下脱去氨基,则C→U、A →I、G →X,引起子 链错误。 3. DNA聚合酶“打滑”:DNA复制时发生碱基的环出现 象,引起一个或数个碱基的插入或缺失,易发生于 几个相同碱基串联的部位。 4. 活性氧(O3)引起的诱变:①氧化碱基与C、A配对, 造成GC → TA颠换,这种损伤可以积累;②H2O2造成 的DNA氧化损伤,此类损伤一般能被修复。
核苷酸切除修复
错配修复
错配修复对 DNA复制忠实 性的贡献力达 102-103,DNA 子链中的错配 几乎完全都被 修正,充分反 映了母链的重 要性。
大肠杆菌甲基化引 导的错配修复
重组修复
易错修复和SOS反应
•SOS反应:当DNA损伤广泛难以继续复制时,由此而
诱发出一系列复杂的反应。
•这种修复特异性低,对碱基的识别、选择能力差。
5.3.4 基因突变的后果
基因突变的后果主要是生物功能的丧失。 某一基因突变后使其所表达的蛋白质或酶失活, 有时还会引起多种酶的缺乏。 有些突变可产生功能获得性显性表现型。 典型的人体细胞突变每个基因每代发生率为107~10-5,但并非所有的突变都会导致疾病。
第六章DNA和RNA的提取

β-巯基乙醇是抗氧化剂,有效地防止酚氧化成醌,避免褐变,使酚容易去除
(二)基因组DNA的提取- CTAB法
组份
CTAB提取缓冲液的改进配方
Tris-HCl EDTA NaCl (pH8.0) (pH8.0) 100 mM 20 mM 1.4M CTAB 3%(W/ V) PVP40 5%(W/ V) β-巯基乙醇 2%(V/V) 使用前加入
主要方法: (1)浓盐法 利用RNP和DNP在电解溶液中溶解度不同,将二者分离。 1)用1M 氯化钠提取,得到的DNP粘液 2) 与含有少量异戊醇的氯仿一起摇荡,使乳化 3) 离心除去蛋白质,此时蛋白质凝胶停留在水相及氯仿相 中间,而DNA位于上层水相中 4) 用2倍体积95%乙醇可将DNA 钠盐沉淀出来.
质粒DNA的提取
使用处于对数期的新鲜菌体 (老化菌体导致开环质粒增加)
质粒DNA-碱裂解法
碱裂解法原理
染色体DNA比质粒DNA分子大得多,且染色体DNA为线状分 子,而质粒DNA为共价闭合环状分子; 当用碱处理DNA溶液时,线状染色体DNA容易发生变性,共 价闭环的质粒DNA在回到中性pH时即恢复其天然构象; 变性染色体DNA片段与变性蛋白质和细胞碎片结合形成沉 淀,而复性的超螺旋质粒DNA分子则以溶解状态存在液相 中,从而可通过离心将两者分开。
磁珠
磁性微粒挂上不同基团可吸附不同的目的 物,从而达到分离目的。
(三)基因组DNA-其它方法
浓盐法:
利用RNP和DNP在盐溶液中溶解度不同,将二者分离
有机溶剂抽提法:
有机溶剂作为蛋白变性剂,同时抑制核酸酶的降解作用
密度梯度离心法:
利用不同内容物密度不同的原理分离各种内容物
分子生物学第六章:DNA损伤与修复

48
4.直接插入嘌呤
DNA链上嘌呤的脱落造成无嘌呤位点,能被
DNA嘌呤插入酶(insertase)识别结合,并在K+
存在下催化游离的嘌呤碱基或脱氧核苷与DNA无
嘌呤部位形成糖苷键。且催化插入的碱基有高
度专一性、与另一条链上的碱基严格配对,使
DNA完全恢复。
49
三、碱基切除修复(Base
Excision Repair,BER)
35
第二节
错配修复
DNA修复
DNA的修复主要类型:
直接修复
切除修复 重组修复 跨损伤修复 (SOS修复)
36
一、错配修复
在DNA复制过程中, DNA聚合酶能够利用
其3ˊ一5ˊ外切核酸酶活性去除错配核苷酸,但
是这种校正作用并不十分可靠, 某些错配核苷酸
可能逃避检测, 出现于新合成的DNA链中。 错
胞嘧啶
O6-乙基鸟嘌呤 胸腺嘧啶
25
(一)烷化剂对DNA的损伤 2.碱基脱落 烷化鸟嘌呤的糖苷键不稳定,容易脱 落形成DNA上的无碱基位点,复制时可以 插入任何核苷酸,造成序列的改变。
26
(一)烷化剂对DNA的损伤
3.断链
DNA链的磷酸二酯键上的氧也容易被 烷基化,结果形成不稳定的磷酸三酯键, 易在糖与磷酸间发生水解,使DNA链断裂。
不识别任何特殊的碱基损失,而是识 别双螺旋形状的改变;修复时切除含有损 伤碱基的那一段 DNA。
54
55
56
核苷酸切除修复 (大肠杆菌)
紫外线诱导uvrA、 uvrB、uvrC和uvrD 四种基因表达
UvrA:识别损伤 部位 UvrB:解旋双链
57
UvrC:
5ˊ末端内切
分子生物学研究方法

优点
(1)相互作用的蛋白质都是经翻译后修饰的,处于天 然状态; (2)蛋白的相互作用是在自然状态下进行的,可以避 免人为的影响; (3)可以分离得到天然状态的相互作ቤተ መጻሕፍቲ ባይዱ蛋白复合物。
缺点
(1)可能检测不到低亲和力和瞬间的蛋白质-蛋白 质相互作用; (2)两种蛋白质的结合可能不是直接结合,而可能 有第三者在中间起桥梁作用; (3)必须在实验前预测目的蛋白是什么,以选择最 后检测的抗体,所以,若预测不正确,实验就得不到 结果,方法本身具有冒险性。 (4)灵敏度没有亲和色谱高。
➢通过报告基因的功能筛选相互作用的蛋白。
Bait and Hunter Plasmids
Yeast two-hybrid transcription
Transfection
举例
3. GST融合蛋白沉降技术
▪ 研究体外蛋白质相
互作用技术
▪ 利 用 GST 对 谷 胱 甘
肽偶联的琼脂糖球 珠的亲和性,从混 合蛋白质样品中纯 化得到相互作用蛋 白。
操作步骤
(1)转染后24-48 h 可收获细胞,加入适量细胞裂解缓冲液 (含蛋白酶抑制剂),冰上裂解30min, 细胞裂解液于4°C, 最 大转速离心30 min后取上清; (2)取少量裂解液以备Western blot分析,剩余裂解液加1μg 相应的抗体加入到细胞裂解液,4°C缓慢摇晃孵育过夜; (3)取10μl protein A 琼脂糖珠,用适量裂解缓冲液洗3 次,每 次3,000 rpm离心3 min; (4)将预处理过的10μl protein A 琼脂糖珠加入到和抗体孵育 过夜的细胞裂解液中4°C缓慢摇晃孵育2-4h,使抗体与protein A琼脂糖珠偶连; (5)免疫沉淀反应后,在4°C 以3,000 rpm 速度离心3 min, 将琼脂糖珠离心至管底;将上清小心吸去,琼脂糖珠用1ml裂解 缓冲液洗3-4次;最后加入15μl的2×SDS 上样缓冲液,沸水煮 5分钟; (6)SDS-PAGE, Western blotting或质谱仪分析
医学分子生物学第六章_信号转导
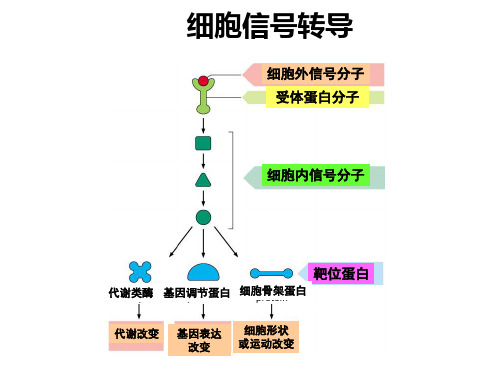
调节蛋白质功能 水平,调节细胞分化和增
和表达水平
殖
受体的结构特点
• 结合结构域-----识别外源信号分子并与之结 合
• 效应结构域-----转换配体信号,使之成为细 胞内分子可识别的信号
3、信号转导分子和分子开关
• 信号转导分子(signaling molecule):细 胞内执行信号转导的成分的一些蛋白质分 子和小分子活性物质。
• 信号转导分子组织在支架蛋白上的意义:
① 保证相关信号转导分子容于一个隔离而稳定的信号转导 通路内,避免与其他不需要的信号转导通路发生交叉反 应,以维持信号转导通路的特异性;
② 增加调控复杂性和多样性。
信号转导通路中的一些环节是由多种分子聚集形成的 信号转导复合物(signaling complex)来完成信号 传递的。
激酶
磷酸基团的受体
蛋白丝氨酸/苏氨酸激酶 蛋白酪氨酸激酶 蛋白组/赖/精氨酸激酶 蛋白半胱氨酸激酶 蛋白天冬氨酸/谷氨酸激酶
丝氨酸/苏氨酸羟基 酪氨酸的酚羟基 咪唑环,胍基,ε-氨基 巯基 酰基
蛋白磷酸酶衰减或终止蛋白激酶诱导的效应
• 蛋白质磷酸酶(phosphatidase)使磷酸化的 蛋白分子发生去磷酸化,与蛋白激酶共同 构成了蛋白质活性的调控系统。
及信息传递,是指一个细胞发出的信息通过介 质传递到另一个细胞并与靶细胞相应的受体相 互作用,然后通过信号转导产生胞内一系列生 理生化反应,最终表现为细胞整体的生物学效 应的过程。
T淋巴细胞
(一)细胞通讯的方式
靶细胞
细胞间隙连接
细胞表面分子接触通讯 可溶型信号分子
化学信号介导通讯
❖分泌化学信号
根据体内化学信号分子作用距离,可以将 其分为三类:
分子生物学教学资料第6章rna剪接

Primary transcript定义:Exons (外显子):编码序列Introns (内含子) : 间隔序列RNA splicing(RNA剪接): 从前mRNA中除去内含子的过程Alternative splicing (可变剪接): 有些前mRNA存在不止一种的剪接方式,从而产生不同的mRNA。
通过这种方式一个基因可以产生多个多肽产物。
通过可变剪接,从一个基因得到的不同产物数量可以从2种到数百甚至数千种。
Why RNA splicing is important?1.RNA剪接的化学基础2. 剪接体Spliceosome:执行RNA的剪接的大复合体,有5种snRNA(核内小RNA: U1,U2,U4,U5,U6),主要执行功能是RNA非蛋白质。
snRNA的三个功能:Recognizing:识别5’剪接位点和分支位点Bringing:将这两个位点集结到一起U2 取代BBP3. 剪接过程可变剪接Alternative splicing and regulation通过可变剪接一个基因可以得到多个产物。
RNA剪接的5种模式①正常剪接②外显子遗漏③外显子延伸④内含子保留⑤可变剪接可变剪接:组成型:同一个基因总是产生多种不同产物调控型:不同的时间、条件下或不同的细胞、组织中,产生不同mRNA剪接调控蛋白结合到特殊序列上:外显子/内含子剪接增强子enhancer(ESE or ISE)-增强附近剪接位点的剪接(剪接->未剪接)外显子/内含子剪接减弱子silencer(ESS or ISS)–减弱附近剪接位点的剪接(未剪接->剪接) (在不同条件下引导剪接体到不同的剪接位点发挥作用;在发育的某个阶段或在某种类型的细胞中,一种特定的SR蛋白的存在与否或者活性高低,就可以决定某一个特定的剪接位点是否得到利用)特殊内含子剪切体:AT-AC型剪接体催化的剪接反应:U1->U11,U2->U12自剪接内含子Self-splicing introns and mechanisms自剪接:前体RNA中的内含子自身折叠成一种特殊的构象,然后催化自身释放的化学过程。
分子生物学 第六章

摆动性
• 反密码子与密码子之间的配对并不完全遵照 碱基互补规律,称为摆动配对。
二、tRNA
(一)结构特点 1.二级结构:三叶草结构
四环: 二氢尿嘧啶环 反密码子环 额外环 胸腺嘧啶假尿嘧啶胞嘧啶环 一臂: 氨基酸接受臂
2.三级结构——“倒L型”
(二)起始tRNA
密码子 氨基酸 表示方法
(二)延伸
1.进位 • 氨酰-tRNA 按照mRNA 分子的编码 信息进入并 结合到核糖 体A位。
(二)延伸
2.成肽
• 转肽酶催化 肽酰-tRNA 上的肽酰基 转移到A位 氨酰-tRNA 上的氨基酸 α-氨基上。
(二)延伸
3.转位
• 转位酶催化核 糖体沿mRNA 的3‘方向移动 一个密码子的 距离,使 mRNA上的下 一个密码子进 入A位,肽酰tRNA由A位移 入P位。
三、修饰
(一)磷酸化 是指在蛋白激酶的催化作用下,ATP的γ-磷酸 基被转移到蛋白质特定位点上的过程。 通常蛋白质的丝氨酸、苏氨酸和在糖基转移酶的作用下,蛋白质的特定 氨基酸残基被共价连接上寡糖链的过程。 • 糖链与氨基酸的连接主要有O型连接和N型 连接两种方式。
终止密码子: 琥珀石(UAG) 赭石(UAA) 卵白石(UGA)
起始密码子: AUG(甲硫氨酸)
2.特性
(1)完整性:有始有终 (2)方向性:5’到3’ (3)连续性:不中断、无重叠 (4)简并性:多对一 (5)统一性:万物统一 (6)摆动性::3’位可变 (7)偏爱性:使用频率各异
简并性
• 一种氨基酸具有 两个或两个以上 的密码子为其编 码,这一特性称 为遗传密码的简 并性。
一、mRNA (一)结构特点
原核 生物
真核 生物
细胞分子生物学 第六章 内质网和蛋白质合成

一、化学组成
1、水和无机离子等一些小分子。离子主要是Na+、K+、Ca2+、Mg2+、
Cl-等。 2、脂类、糖类、氨基酸、核苷酸及其衍生物等中等分子。
3、蛋白质、多糖和RNA等大分子。
4、含有糖原等一些处于贮存状态的重要化合物。
二、细胞质溶质的基本属性
1、酸度稳定
主要内容
第一节 细胞质溶质
第二节 内质网 第三节 核糖体 第四节 蛋白质合成的命运
1 、胞质溶质:是指细胞质中可溶性 的液体。含有大量的各种可溶性蛋白 和代谢物质。在细胞的物质代谢、维 持细胞内环境稳定性等方面具有非常 重要的作用。 2.内膜系统:细胞内的许多膜性细胞 器在结构和功能上有一定的连续性, 构成了膜体系,故称为内膜系统,主 要包括内质网、高尔基体、溶酶体等, 但不包括半自主性的线粒体和叶绿体。
SRP为GTP结合蛋白,可结合GTP。当SRP与信号序列和核糖体结合
后,翻译过程暂时停止。 糙面内质网上有信号识别颗粒的受体,SRP引导核糖体与内质网膜 上的的SRP受体结合。内质网膜上存在转运体,转运体可形成通道。当 SRP与SRP受体结合后,新生肽链随即从核糖体的大单位进入转运体的中
央通道。
肽链信号序列进入内质网腔后,随即被膜上与转运体结合的信号肽 酶切除。切除信号肽的新生肽链则继续穿过转运体通道进入内质网腔。
动有条不紊地进行。细胞骨架对细胞质溶质中的各种小泡运输也具有 导向作用。
三、细胞质溶质的功能
1、维持一些代谢反应途径
细胞质溶质中含有很多酶,大部分中间代谢是在细胞质溶质中进 行的。例如:糖酵解的起始步骤
2、保持细胞内环境的稳定
缓冲pH值,使酸度保持在 pH7.2 ,为各种中间代谢反应的进行
分子生物学(杨洋)第六章 rna剪接-rna splicing

Step 1: The OH of the conserved A at the branch site attacks the phosphoryl group of the conserved G in the 5’ splice site. As a result, the 5’ exon is released and the 5’-end of the intron forms a three-way junction structure.
Trans-splicing:原本位于两个不同RNA分子 上的外显子可以连接到同一条mRNA中
标准剪接:套索结构
反式剪接:Y型分支结构
Figure 13-5
Not a lariat
Topic 2 THE SPLICESOME MACHINERY (剪接体)
2.1 RNA splicing is carried out by a large complex called spliceosome (RNA剪接是由一个称为剪接体的大复合体 执行的) The above described splicing of introns from pre-mRNA are mediated by the spliceosome. The spliceosome comprises about 150 proteins and 5 snRNAs(5种RNA,核内小RNA). Many functions of the spliceosome are carried out by its RNA components(剪接体的多数 功能是由其RNA而不是蛋白质执行的)
分支点
RNA-RNA interactions between different snRNPs, and between snRNPs and pre-mRNA
分子生物学期末复习

分子生物学期末考试复习题型:名词解释(英文)、选择、判断、简答、设计性问答(最后一节)第六章DNA和RNA结构1、DNA 构建模块(DNAbuilding blocks): 碱基(Base)、核背酸(Nucleoside)。
核廿酸是DNA基本的构建模块。
2、DNA的全称:脫氧核糖核酸3、DNA 的4 种碱基:卩剽令(Purines) : Adenine (A)、Giiaiuiie (G)喘唳(Pyriinidine) : Cytosine (C)、Thvinine (T) 碱基具有形成异构体的能力是DNA合成时出错的普遍来源。
4、DNA的结构特点:一条DNA分子是由2条反向平行的多核背酸链相互旋转形成的双螺旋结构。
以磷酸二酯键为基础构成规则的不断重复的糖磷酸骨架组成的多核苛酸链。
双螺旋的两条链具有互补的序列,方向相反。
5、决定DNA双链稳定性的因素:①氢键贡献于双螺旋的热动力学稳定性;②双螺旋堆积时碱基间的相互作用(兀-兀共轨)对双螺旋的稳定性起重要作用。
6、DNA双螺旋有大沟和小沟(Mmoi and Major grooves),这是由碱基对的空间几何结构所决定的。
人沟(Major groove)富含丰富的化学信息。
7、双螺旋的多重构彖:A> (RNA双螺旋与其类似,右手螺旋)E型(最接近生理状态,右手螺旋)Z型(左手螺旋)8、变性(Denaturation):当DNA溶液温度高于生理温度(接近100 °C)或者pH较高时,互补的两条链就会分开, 这一过程称为变性。
杂交(Hybridization):两条不同来源的单链DNA或RNA通过碱基互补配对形成双链杂交分子的过程。
复性(Annealing/renature):当变性的DNA热溶液缓慢降温,DNA的互补链又可重新聚合,形成规则双螺旋,称为复性。
熔点(Tm (melting pomt)):吸收值增加到最犬值一半时的温度。
9、DNA超螺旋结构的解除是靠拓扑异构酶实现的。
分子生物学-课件-分子生物第六章可编辑全文

• 使双链DNA解链度达到50%所需的温度称为解链温度 (Tm)、变性温 度、熔点
• DNA的解链温度一般在82-95℃ ,与DNA的分子大小和碱基组成、 溶液的pH值和离子强度(+)等有关
二、复性
• DNA复性:缓慢降温可以使热变性DNA重新 形成互补双链结构
且RNA的完整性和纯度都很高
3、氯化锂-尿素法
• 利用高浓度尿素变性蛋白质同时抑制RNA 酶,氯化锂选择性沉淀RNA
• 缺点:存在DNA污染,氯化锂沉淀RNA会 丢失一些小分子RNA,如5sRNA等
• 优点:快速、简便、产量高,尤其适用于大 量样品少量组织细胞的RNA提取
4·热酚法
• 将异硫氰酸胍、巯基乙醇和SDS等联合使用,可以快速裂 解细胞,解离核蛋白复合物,释放RNA,并有效抑制 RNase的活性
• 在某些理化因素的作用下,维 系核酸二级结构的氢键和碱基 堆积力受到破坏,DNA双螺旋 结构松散,变成单链的过程称 为核酸的变性
• 核酸双螺旋区的氢键断裂,变 成单链,但并不涉及共价键的 断裂
DNA解链曲线
DNA变性的本质是氢键的断裂
核酸的变性因素
• 变性方法
–热变性、酸碱变性、化学变性剂(乙醇、 尿素和甲酰胺 )
(3)煮沸裂解法
• 以溶菌酶、Triton裂解细菌,然后以沸水浴加热,不 仅促进细菌裂解,还可以使蛋白质和染色体DNA、质 粒DNA变性
• 当降低温度时,闭环质粒DNA复性留在上清液中,而 染色体DNA保持与细胞膜碎片结合、沉淀,可以通过 离心除去
• 在离心上清液中加入有机溶剂 (例如异丙醇)便得到质 粒DNA粗品沉淀
分子生物学6

W
M
隐性
W
M
顺式显性
分子生物学6
W
M
顺式及反式显性
W
M显性负突变体源自分子生物学6三、原核生物的基因转录及表达调控
2. 基因表达调控 (2)操纵子的发现 n 结论
通过阻遏基因(阻遏物)与操纵区, lacZ、lacY、lacA这3个结构基因是一起 被调控的,它们组成了一个操纵子(the lac operon) n 后来的研究证实了Jacob与Monod的预 言,并使操纵子的慨念进一步完善
分子生物学6
三、原核生物的基因转录及表达调控
2. 基因表达调控 (4)乳糖操纵子的正调控
n 乳糖操纵子的启动子实际上是一种弱启动子, 其 -35 box与原核启动子相应的共同序列相差 甚大,因此才需要CAP-cAMP进行正调控; 如果是强启动子(与共同序列十分相近), 则不需要正调控,但这种情况会使得即使葡 萄糖存在,乳糖操纵子也一样运作。
分子生物学6
negative control
分子生物学6
三、原核生物的基因转录及表达调控 2. 基因表达调控
(2)操纵子的发现 n 操纵子概念的提出者
n François Jacob n Jacques Monod
分子生物学6
François Jacob, Jacques Monod, et André Lwoff, Prix Nobel 1965
分子生物学6
三、原核生物的基因转录及表达调控
2. 基因表达调控 (4)乳糖操纵子的正调控
n 阻遏物的调控是负调控,乳糖操纵子的调控还需正调 控因子
n 代谢物阻遏效应(catabolite repression) n cAMP在细胞中的浓度与葡萄糖的含量成反比,它与
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
I gene active repressor
38 kd / monomer tetramer 152 kd
(来源:不详)
binding on Operator
I+
mut. iC (constitutive mut.) iS (super--reperessible)
Lac or IPTG
ic
p o
Zபைடு நூலகம்
Y
A
iC gene产物repressor
丧失与O位点结合的能力
ic
tetramer
Non binding repressor
ic
tetramer
tetramer
等位基因间的显隐关系 Lactose
I+/IC
I+
tetramer 变构
I+ > iC
(Source:Molecular Biology(2002),Robert F.Weaver,Page177)
c) Model
●
Signal molecular be needed forboth types Add signal mol. Operon on (inducible operon)
Inducer
Add signal mol. Operon off (repressibleoperon)
I+ > iS
operon off
e.g. cAMP control (universal controllingsystem)
(Source:Molecular Biology(2002),Robert F.Weaver,Page173)
E.coli Glucose Lactose
Why?
Lac operon open but no transcripts
Neg.
Pos.
●
Repressorbinding on O site
阻止转录启动
(来源:分子生物学(2007),郑用琏,第239页)
• Expressorbinding front p site
激活转录启动
●
Operon off
意味转录效率极低
●
Negative control 是广泛保险的机制 (自然选择使Prok. 获得选择优势) Positive control 是灵活,严格,经济的调控机制
I+
OC
Z
Y
A
OC失去与repressor 特异结合的能力
mRNA
lactose
OC > O+
mRNA
cis-dominant
I+
O+
等位基因间的显隐关系
cis-dominant
The ability of a site (cis-factor) to control adjacent gene irrespective of the presence in the cell of other alleles of the site.
(Source:Cohn. Journal of Molecular Biology, Vol. 34: 366.1968)
O gene (operator) cis-action factor
mRNAstartpoint
unwinding
Obstruction? Competition?
RNA polymerasebinding
Inactive expressor
active expressor
binding on front Psite
激活RNApolymerase启动
(来源:不详)
w.t. (I+O+P+) 诱导型
iSmut. 超阻突变 (super-repression) expressorcan not be activated by inducer
w.t. (I+ O+ P+) 诱导型 add inducer operon on no inducer operon off OC mut. (I+ OC P+) constitutive mut. (组成型) OC失去与repressor特异结合的能力
wild-type operator(O+)
6.2. post-transcriptional level control
6.3.Gene expression control in translation level 6.4. 染色质重建对基因表达的控制 6.5.ProgrammedCell Death (PCD) and
development
紧密连锁地排列在一起,受同一操纵子控制 Lac operon
● ● ●
I p o Z Y A
各结构基因按一定比例协调翻译 ( Z : Y :A= 5 : 2 : 1 ) 具有极性突变效应 P& O基因(cis)紧密连锁或 彼此重叠 I 基因(trans) 位点不固定
b) operon control type Negative & Positive
O+ — mut. OC occur frequently at left site of axis (?)
---调控机理
repressortetramer与operator发生特异结合 operon off Inducer(lactose) 与repressor特异结合 tetramer 变构 特异结合力下降1000X operonon 作用于 O 位点上的repressor 变构 脱离O位 作用于游离的repressor 变构 失去结合于O位的能力
调控机理上
调控层次上
基因表达的调控涉及
RNA转录的开/关
数量 选择性加工
蛋白质翻译速率
数量
加工、降解和分泌…
转录水平上的调控是最为经济,
灵活,又是最为重要,复杂的调控
●在复杂的基因组内,确定需要基因转录的起始位点
●精细调节基因表达的水平,以保证生物体对环境的适应 ●
cis factor& trans factor间严格而又灵活的互作 的进行式转录(不中断,准确终止)
unwinding
repressor& RNA polymerase 对重叠位点竞争
RNA polymerasebinding
Repressorbinding
O gene (operator) cis-action factor
Obstruction
O gene (operator) cis-action factor
cis acting factor 对与其紧密连锁基因的控制 效应不受其等位基因的影响。
•Positive—inducible operon (多为分解酶类)
I inactive expressor (apoinducer诱导蛋白) inducer
operon off
active expressor
第6章 基因表达调控
(Controlling of the Gene Expression)
(Source:Pique,Michaeland Peter E.Wright,Dept.(cover photo,science 245 (11Aug 1989).))
6.1. transcriptionallevel control
• I gene
repressor
expressor (apoinducer)
无辅基诱导物
Negative
Positive
i- or 不加入I基因产物 I+ or 加入I基因产物 i- or 不加入I基因产物 I+ or 加入I基因产物
operonon operonoff operonoff operon on
lactose analog
is inducer of Lac operon
bindingbetween labeled lacO-DNA with 32p and added increasing amountsof lac repressor.
IPTG;异丙基 -β -D-硫 代半乳糖苷, 自己不被分解 的强诱导物 (安慰诱导物)
iS
p o
iS gene 产物repessor 不能与inducer结合
mutrepressor
lactose
IS/I+
tetramer
iS > I+ iS > iC
等位基因间的显隐关系
I+
(Source:Molecular Biology(2002),Robert F.Weaver,Page178)
Co-repressor
Operon control model
negative positive
Ind.
active inactive
Rep. inactive active
(来源:分子生物学(2007),郑用琏,第218页)
• Negative—inducible operon
I
例 (分解酶类lactoseoperon)
基因表达调控的简介
阐明的科学问题
真核与原核生物如何调控数以千、万计
的基因以最为经济、有效的时空模式进
行转录,从而实现对环境的适应、细胞
的分化, 组织的特化 和个体的发育。
原核生物与真核生物基因
表达调控机制具有惊人的相似性
共同的起源与共同的分子基础
核酸分子间的互作 核酸与蛋白质分子间的互作 蛋白质分子间的互作 transcriptionallevel post—transcriptionallevel translationallevel post—translationallevel