分子生物学:第七章翻译1
分子生物学 第七章 蛋白质的生物合成

7.1 遗传密码 7.2 tRNA 7.3 氨酰-tRNA合成酶 7.4 核糖体 7.5 与蛋白质合成有关的因子 7.6 蛋白质合成的生物学机制 7.7 蛋白质转运机制
比DNA复制和转录更为复杂的过程
氨基酸活化与转运----这个过程是在氨基酸活化酶和镁 离子作用下把氨基酸激活成为活化氨基酸。 起始----核糖体与mRNA结合并与氨酰基tRNA生成起 始复合物。 延伸----由于核糖体沿mRNA5’端向3’端移动,开始了 从N端向C端的多肽合成,这是蛋白质合成过程中速度 最快的阶段。 终止以及肽链的释放,核糖体从mRNA上解离,准备 新一轮合成反应。
(3)校正tRNA 校正tRNA分为无义突变及错义突变校正。 无义突变:在蛋白质的结构基因中,一个核 苷酸的改变可能使代表某个氨基酸的密码 子变成终止密码子(UAG、UGA、 UAA),使蛋白质合成提前终止,合成无 功能的或无意义的多肽。
错义突变是由于结构基因中某个核苷酸
的变化使一种氨基酸的密码变成另一种 氨基酸的密码。 错义突变的校正tRNA通过反密码子区的 改变把正确的氨基酸加到肽链上,合成 正常的蛋白质。(摆动原理现象)
结合的AA-tRNA分开。
7.1.3 遗传密码的特点
⑴连续性
⑵简并性与偏爱性
⑶通用性与专一性
(特殊性)
⑷终止密码
⑸密码子与反密码子
简并性: 由一种以上密码子编码同一个氨基酸的现象。 18种氨基酸都有一个以上的密码子 。
同义密码子。
只有精氨酸是个
例外,因为在真 核生物中CG双联
7.2 tRNA
7.2.1 tRNA的结构 由于链内的碱基互 补配对,tRNA呈现
分子生物学中英文对照

acetyl CoA / 乙酰辅酶A 一种小分子的水溶性代谢产物,由与辅酶A 相连的乙酰基组成,产生于丙酮酸、脂肪酸及氨基酸的氧化过程;其乙酰基在柠檬酸循环中被转移到柠檬酸。
actin / 肌动蛋白,肌纤蛋白富含于真核细胞中的结构蛋白,与许多其他蛋白相互作用。
其球形单体( G2肌动蛋白) 聚合形成肌动蛋白纤丝( F2肌动蛋白) 。
在肌肉细胞收缩时F2肌动蛋白与肌球蛋白相互作用。
activation energy / 活化能(克服障碍以) 启动化学反应所需的能量投入。
降低活化能,可增加酶的反应速率。
active site / 活性中心,活性部位酶分子上与底物结合及进行催化反应的区域。
active transport / 主动转运离子或小分子逆浓度梯度或电化学梯度的耗能跨膜运动。
由ATP 耦联水解或另一分子顺其电化学梯度的转运提供能量。
adenylyl cyclase / 酰苷酸环化酶催化由ATP 生成环化腺苷酸(cAMP) 的膜附着酶。
特定配体与细胞表面的相应受体结合引发该酶的激活并使胞内的cAMP 升高。
allele / 等位基因位于同源染色体上对应部位的基因的两种或多种可能形式之一。
allosteric transition / 变构转换小分子与蛋白质上特定调节部位相结合所引起的蛋白质之三级及(或) 四级结构的改变,其活性随之发生变化。
多亚单位酶的变构调节很普遍。
alpha(α) helix /α螺旋常见的蛋白质二级结构,其氨基酸线性序列叠为右旋螺旋,借助主链上的羧基与酰胺基间的氢键维持稳定。
aminoacyl2tRNA / 氨酰转移核糖核酸用于蛋白合成的氨基酸的激活形式,含有借高能酯键与tRNA 分子上3’2羟基相结合的氨基酸。
amphipathic / 两亲的,兼性的指既有亲水性部分又有疏水性部分的分子或结构。
anaphase / ( 细胞分裂) 后期姐妹染色体(或有丝分裂期的成对同源物) 裂开并分别(分离) 朝纺锤体两极移动的有丝分裂期。
分子生物学(杨洋)第七章 翻译(1)

How the eukaryotic ribosome recognize the start codon AUG?
What is the function of initiator tRNA---Met-tRNAMet in the translation initiation of eukaryotic cell?
2. a charged tRNA must be placed into the P site of ribosome
3. the ribosome must be precisely positioned over the start codon
start codon
01.11.202 0
One-base shift will result in the synthesis of completely unrelated polypeptide
01.11.202 0
How to answer the scientific question(解决科学问题)
Establish a good model
Design the elegant experiment
Analyze the experimental results
Answer the scientific question
a
20
研究论文(原始研究工作)的学习
01.11.202 0
设计实验
实验数据分析
得出结论
提出科学问题
分析科学问题
解决科学问题
a
22
01.11.202
tRNAiMet functions in directing th0 e scanning ribosome to the start site of translation. Science, Vol
分子生物学2-7章作业及答案

第二章一、名词解释1、DNA的一级结构:四种脱氧核苷酸按照一定的排列顺序以3’,5’磷酸二酯键相连形成的直线或环状多聚体,即四种脱氧核苷酸的连接及排列顺序。
2、DNA的二级结构:DNA两条多核苷酸链反向平行盘绕而成的双螺旋结构.3、DNA的三级结构:DNA双螺旋进一步扭曲盘绕所形成的特定空间结构。
4、DNA超螺旋:DNA双螺旋进一步扭曲盘绕所形成的特定空间结构,是DNA结构的主要形式,可分为正超螺旋与负超螺旋两大类。
按DNA双螺旋的相反方向缠绕而成的超螺旋成为负超螺旋,反之,则称为正超螺旋。
所有天然的超螺旋DNA均为负超螺旋。
5、DNA拓扑异构体:核苷酸数目相同,但连接数不同的核酸,称拓扑异构体6、DNA的变性与复性:变性(双链→单链)在某些理化因素作用下,氢键断裂,DNA双链解开成两条单链的过程。
复性(单链→双链)变性DNA在适当条件下,分开的两条单链分子按照碱基互补配对原则重新恢复天然的双螺旋构想的现象。
7、DNA的熔链温度(Tm值):DNA加热变性时,紫外吸收达到最大值的一半时的温度,即DNA分子内50%的双链结构被解开成单链。
Tm值计算公式:Tm=69.3+0.41(G+C)%;<18bp的寡核苷酸的Tm计算:Tm=4(G+C)+2(A+T)。
8、DNA退火:热变性的DNA经缓慢冷却后即可复性,称为退火9、基因:编码一种功能蛋白或RNA分子所必需的全部DNA序列。
10、基因组:生物的单倍体细胞中的所有DNA,包括核DNA和线粒体、叶绿体等细胞器DNA11、C值:生物单倍体基因组中的全部DNA量称为C值12、C值矛盾:C值的大小与生物的复杂度和进化的地位并不一致,称为C值矛盾或C值悖论13、基因家族:一组功能相似、且核苷酸序列具有同源性的基因。
可能由某一共同祖先基因经重复和突变产生。
14、假基因:假基因是原始的、有活性的基因经突变而形成的、稳定的无活性的拷贝。
表示方法:Ψα1表示与α1相似的假基因15、转座:遗传可移动因子介导的物质的重排现象。
分子生物学7第七章 蛋白质翻译ppt课件

d、按paracodon在tRNA上的位置(氨基 酸序列) 将AARS分为两类
type I 包括 Val,Arg,Gln,Glu,Ile,Leu,Met,Trp,Tyr paracodon 大多位于反密码子臂
type II paracodon 大多位于氨基酸接受臂 个别还同时在附加臂上有相应碱基
二、 mRNA
★ 蛋白质翻译是基因表达的第二步 ★ tRNA在翻译过程中起“译员”的作用 ★ 参与翻译的RNA 除tRNA外,还有rRNA和mRNA
★ tRNA 既是密码子的受体,也是氨基酸的受体
★ tRNA 接受AA要通过氨酰tRNA合成酶及其自身的 paracodon 的作用才能实现
★ tRNA 通过其自身的 anticodon而识别 codon ★ 密码子有自身的特性
c、 paracodon 的特征
--- 同一种AARS 所识别的一组同功受体具有相同的 副密码子
tRNA
Ala (GGC)
tRNA
Ala (UGC)
具有G3 :U70 paracodon
--- paracodon 是为AARS(特定氨基酸)所识别的若干 碱基(并非均为一对核苷酸),且位置不固定。
--- 尽管副密码子不是单独与AA发生相互作用,但是副密码 子可能与AA的侧链基团有某种相应性。
三联体 前两个重要 摇摆性 通用性 有一定的使用频率
★ 多种翻译因子组成翻译起始复合物,完成翻译的起始、 延伸和终止,并且保证其准确性
第一节 基本元件
一、 tRNA
最小的 RNA,4S,70 ~ 80个NT 1、 tRNA的高级结构
1964 Holly. R. 鉴定出 tRNAphe 的二级结构为三叶草 形(77个NT)
分子生物学名词解释

分子生物学名词解释1.翻译(translation):以mRNA为模板,氨酰-tRNA为原料直接供体,在多种蛋白质因子和酶的参与下,在核糖体上将mRNA分子上的核苷酸顺序表达为有特定氨基酸顺序的蛋白质的过程。
2.密码子(codon):mRNA中碱基顺序与蛋白质中氨基酸顺序的对应关系是通过密码实现的, mRNA中每三个相邻的碱基决定一个氨基酸,这三个相邻的碱基称为一个密码子。
3.密码的简并性(degeneracy):—个氨基酸具有两个以上密码子的现象。
4.同义密码子(synonym codon):为同—种氨基酸编码的各个密码子,称为同义密码了。
5.变偶假说(wobble hypothesis):指反密码子的前两个碱基(3’-端)按照标准与密码子的前两个碱基(5’-端)配对,而反密码子中的第三个碱基那么有某种程度的变动,使其有可能与几种不同的碱基配对。
6.移码突变(frame-shift mutation):在mRNA中,假设插入或删去一个核苷酸,就会使读码发错误,称为移码,由于移码而造成的突变、称移码突变。
7.同功受体(isoacceptor):转运同一种氨基酸的几种tRNA称为同功受体。
8.反密码子(anticodon):指tRNA反密码子环中的三个核苷酸的序列,在蛋白质合成过程中通过碱基配对,识别并结合到mRNA的特殊密码上。
9.多核糖体(polysome):mRNA同时与假设干个核糖体结合形成的念珠状结构,称为多核糖体。
1.中心法那么(central dogma):生物体遗传信息流动途径。
最初由Crick(1958)提出,经后人的不断补充和修改,现包括反转录和RNA复制等内容。
2.半保存复制(简称复制)〔semiconservative replication):亲代双链DNA 以每条链为模板,按碱基配对原那么各合成一条互补链,这样一条亲代DNA双螺旋,形成两条完全相同的子代DNA螺旋,子代DNA分子中都有一条合成的“新〞链和一条来自亲代的旧链,称为半保存复制。
分子生物学复习7-9

第七章基因的表达与调控(上)——原核基因表达调控模式(一)基本概念1.基因表达:细胞在生命过程中,把蕴藏在DNA中的遗传信息经过转录和翻译,转变成为蛋白质或功能RNA分子的过程称为基因表达。
2.基因表达调控:围绕基因表达过程中发生的各种各样的调节方式都统称为基因表达调控。
rRNA或tRNA的基因经转录和转录后加工产生成熟的rRNA或tRNA,也是rRNA或tRNA 的基因表达,因为rRNA或tRNA就具有在蛋白质翻译方面的功能。
3.组成型表达:指不大受环境变动而变化的一类基因表达。
如DNA聚合酶,RNA聚合酶等代谢过程中十分必需的酶或蛋白质的表达。
管家基因:某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因。
管家基因无论表达水平高低,较少受到环境因素的影响。
在基因表达研究中,常作为对照基因适应型表达:指环境的变化容易使其表达水平变动的一类基因表达。
应环境条件变化基因表达水平增高或从无到有的现象称为诱导,这类基因被称为可诱导的基因;相反,随环境条件变化而基因表达水平降低或变为不表达的现象称为阻遏,相应的基因被称为可阻遏的基因。
4.结构基因:编码蛋白质或功能性RNA的任何基因。
所编码的蛋白质主要是组成细胞和组织基本成分的结构蛋白、具有催化活性的酶和调节蛋白等。
原核生物的结构基因一般成簇排列,真核生物独立存在。
结构基因簇由单一启动子共同调控。
调节基因:参与其他基因表达调控的RNA或蛋白质的编码基因。
①调节基因编码的调节物质通过与DNA上的特定位点结合控制转录是调控的关键。
②调节物与DNA特定位点的相互作用能以正调控的方式(启动或增强基因表达活性调节靶基因,也能以负调控的方式(关闭或降低基因表达活性)调节靶基因。
操纵子:由操纵基因以及相邻的若干结构基因所组成的功能单位,其中结构基因的转录受操纵基因的控制。
(二)原核基因调控的分类和主要特点一、原核生物的基因调控特点:(1)基因调控主要发生在转录水平上,形式主要是操纵子调控.(2)有时也从DNA水平对基因表达进行调控,实质是基因重排。
分子生物学 第7章 RNA复制与逆转录ppt课件

精选ppt课件2021
19
tRNA
R U5 PB
gag
R′ U′5
R U5 PB
gag
R′ U′5
PB
gag
负链 DNA
An
pol
env
U3 R
以 tRNA 为引物开始合成 cDNA 负链
pol
env
U3 R An
RNase H 降解模板 R 和 U5
An
pol
env
U3 R
负链 DNA3′端与模板 R 区配对(第一次跳跃)
精选ppt课件2021
11
7.2.1 RNA复制酶类
由病毒自己编码的RNA复制酶主要是依赖RNA的RNA 聚合酶(RNA-dependent RNA polymerase,RdRP)。
RNA复制酶功能与特性: ⑴ 以“–RNA”为模板合成“+RNA”活性高,而以“+RNA” 为模板合成“–RNA”活性低; ⑵ 合成RNA链方向为5′→3′; ⑶ 能将互补的双链RNA分开; ⑷ 缺少核酸酶活性,校正功能低,差错率高; ⑸ 多数分布在细胞质中,少数在细胞核中; ⑹ 酶活性具有可调控性,复制时,RdRP抑制外壳蛋白表达, 而成熟期,外壳蛋白抑制RdRP活性。
这类病毒包括滤泡性口腔炎病毒、流感病毒、
副流感病毒、莴苣坏死黄化病毒、麻疹病毒、狂犬病毒
等。
脱壳
病毒
RdRP
加工
亲代 –RNA
+RNA ① RdRP
子代
mRNA ② 翻译
–RNA
GpppN GpppN
帽子 mRNA
③ 翻译
子代病毒颗粒 组装
RNA 复制酶 病毒外壳蛋白 致病蛋白
分子生物学第7章RNA加工和核糖核蛋白
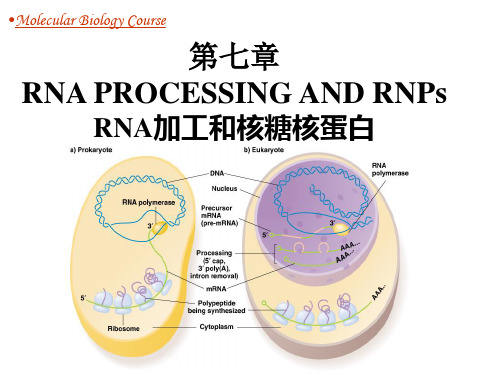
每个转录单位转录 成单个的RNA前体 分子,经剪切后变 成为成熟的RNA的 分子。
rRNA前体的加工 是由RNase Ⅲ负责 的。
2. 大肠杆菌rRNA转录后加工
➢ 6500nt初始转录物折叠形成一 些茎环结构
➢ 初始转录物与蛋白质结合形成 核糖核蛋白复合体
➢ 进行甲基化修饰 S-adenosylmethionine (SAM, S-腺苷甲硫氨酸)
RNA加工的类型
RNA加工的类型 ➢内切核酸酶和外切核酸酶切除核苷酸 ➢在初始转录物或剪切产物的5’端或3’端加上 核苷酸 ➢对某些核苷酸的碱基或糖苷进行修饰 ➢RNA编辑
第1节 rRNA加工与核糖体
一、原核生物rRNA 加工 二、真核生物 rRNA 加工
一、原核生物rRNA 加工
1. rRNA 基因的结构 ➢ E.coli 有7种不同的rRNA操纵子分散在整个基因组 中 ➢ 每个操纵子包含一个拷贝的5s rRNA、16s rRNA、 23s rRNA序列。 ➢ 有一到4个编码tRNA的序列。
(4) 最后修正:通过外切酶等将20S中和已退火的32S 中残余的ITS切除掉。
47S前 rRNA经历了一系列剪切,先切去外部转录间隔区,再切去内部 转录间隔区,释放32S和20S两个前RNA,最后释放18S、5.8S、28S rRNA。
47S pre-rRNA
ETS:external transcribed spacers ITS:internal transcribed spacers
rplJ(L10) rplJ(L10) rpoB(β亚基) rpoC (β’亚基)
转录
前体多顺反子mRNA
RNase Ⅲ 加工
成熟的mRNA
T7噬菌体早期区转录单条前体RNA,经 RNase Ⅲ剪切成5条成熟的mRNA
分子生物学名词英汉对照

ρ依赖型终止
A form of transcription termination in prokaryotes that depends on the protein ρ as well as on sequences in the DNA/RNA.
原核生物中的一种转录终止方式,依赖于ρ蛋白和DNA/RNA序列引起转录终止。
─10 box
─10框
Common promoter element inE. coli. Named for its location approximately 10 bases upstream of the transcription start site.
大肠杆菌中常见的启动子元件。由于它位于转录起始为点上游约10个碱基处而得名。
蛋白质的一种二级结构,相对平坦,在两条平行的或反向平行的肽段之间形成氢键。
σsubunห้องสมุดไป่ตู้t
σ亚基
Component of prokaryotic RNA polymerase holoenzyme. Required for recognition of promoters.
原核生物RNA聚合酶全酶的组成成分。在启动子识别中需要。
氨酰-tRNA合成酶
A protein that matches tRNAs with the correct amino acid. Name origin: enzymesynthesizes theacylbond that joinsaminoacids totRNAs.
原核生物中DNA聚合酶I的一种功能。允许以DNA合成方向去除核苷酸。常在去除RNA引物中使用。
分子生物学(杨洋)第七章 翻译-1

Why?
10
Outline
2020/4/4
Topic 1-4: Four components of translation machinery
mRNA
tRNA
attachment of amino acids to tRNA (aminoacyl-tRNA synthetases)
This part concerned with one of the greatest challenges in understanding the gene- how the gene is expressed
3
2020/4/4
Ch 12: Mechanisms of transcription
mRNA:5-AUGAGUAACGCG-3 translation
Protein: Met-Ser-Asn-Ala
9
2020/4/4
Translation extremely costs
In rapid growing bacterial cells, protein synthesis consumes
因
组
的
保
基
持
RNA processing
因 组
的
Translation 表 达
6
2020/4/4
What is translation?
--it is the story about decoding the genetic information contained in messenger RNA (mRNA) into proteins
分子生物学第七章 翻译-3

Model: poly(U)-ribosome-poly(Phe) (14C-labeled) UUU--Phe poly(U)—poly(Phe)
放射性标记(14C-labeled)
Release of peptidyl-puromycin from ribosome
Detection of the 14C-labeled released peptidyl-puromycin can reflect the peptidyl transferase activity
50S subunit of ribosome alone can have peptidyl transferase activity
[14C]
[14C]
How can we distinguish the released 14C-poly(Phe)-puro from the 14C-poly(Phe)-ribosome?
We should establish a system which can detect peptidyl transferase
activity easily and in vitro(体外)
Puromycin can be used to detect the peptidyl transferase reaction in vitro
First question:
Which part of ribosome is responsible for peptidyl transferase reaction? (50S subunit or 30S subunit?)
Localization of peptidyl transferase
分子生物学第七章原核生物基因表达调控

(三)、阻遏物 lac I 基因产物及功能
Lac 操纵子阻遏物 mRNA 是由弱启动子控制下组 成型合成的,该阻遏蛋白具有4个相同的亚基,每个亚 基均含347个氨基酸残基。
lacI 基因为组成型,通过启动子的上升突变体可获 得较多的阻遏蛋白;
阻遏物 2022/10/18
β-半乳糖苷酶 透过酶 转乙酰3酶2
2022/10/18
16
调节机理:
细胞中某一氨基酸或嘧啶的浓度发生改变
氨酰 – tRNA的浓度变化
核糖体在转录产物RNA上的结合位置不 同,使得RNA形成特定的二级结构 由RNA的二级结构判断基因能否继续转录
2022/10/18
17
3、降解物对基因活性的调节P252
葡萄糖效应或降解物抑制作用:细菌培养基中在 葡萄糖存在的情况下,即使加入乳糖、半乳糖等 诱导物,与其对应的操纵子也不会启动,这种现 象称为葡萄糖效应或降解物抑制作用。
这是通过阻止乳糖操纵子表达来完成的,这种 效应称为降解物抑制(catabolite repression)。
2022/10/18
35
(五)、cAMP与代谢物激活蛋白
葡萄糖
葡萄糖-6-磷酸
甘油 某些代谢产物抑制活性
腺苷酸环化酶
ATP
cAMP
编码
cAMP-CAP
Crp基因
代谢物激活蛋白 CAP
葡萄糖对其它糖的代谢抑制,是通过对 cAMP的抑制完成的。
2022/10/18
22
一、酶的诱导 ——
lac 体系受调控的证据
两种含硫的乳糖类似物:
异丙基巯基半乳糖苷
(IPTG)
巯甲基半乳糖苷(TMG)
E. coli 在不含乳糖的培养基生 长时,β-半乳糖苷酶含量极低;
分子生物学课件第七章 蛋白质的生物合成-翻译

2020/10/28
38
2020/10/28
39
原核生物翻译过程中核蛋白体结构模式:
P位:肽酰位 (peptidyl site)
A位:氨基酰位 (aminoacyl site)
E位:排出位 (exit site)
2020/10/28
40
蛋白质生物合成体系
mRNA、tRNA、rRNA n 氨基酸
大多数简并性表现在密码子的第三个核苷酸上,即 第一、二个核苷酸确定后,第三个核苷酸可变。
色氨酸
意义: 简并密码子越多,生物遗传的稳定性越大,
氨基酸出现频率越高
2020/10/28
18
(3)摆动性(wobble)
转运氨基酸的tRNA的反密码需要通过碱基互补与 mRNA上的遗传密码反向配对结合,但反密码与密码间 不严格遵守常见的碱基配对规律,称为摆动配对。
氨基酰-tRNA + AMP +PPi
2020/10/28
31
(2)氨基酰tRNA合成酶(amino acyl-tRNA synthetase,AARS)
存在于胞液中,催化一个特定的aa结合到相应的 tRNA分子上。
每种氨基酰tRNA合成酶对相应氨基酸以及携带氨 基酸的数种tRNA具有高度特异性,保证tRNA能够 携带正确的氨基酸对号入座。
UUU,UUG,UGU,GUU, GGG,GGU,GUG,UGG。 • U和G随机加入到三联体中,这样按比例各个位置 上进入U和G的概率不同,如氨基酸测定结果:
2020/10/28
7
• 如UUU:UGG=(555):(511)
= 25 : 1
• 同理UUU:UUG =5 :1,
• 根据检测结果推测:
分子生物学-13-4-第七章原核基因表达调控-Arb

因此只有在没有葡萄糖的时候,同时又有半乳糖的时候,启动子1才是开放的为什么gal 操纵子需要两个转录起始位点?(涉及半乳糖在细胞代谢中的双重功能)半乳糖两个作用: 可以作为唯一碳源供细胞生长; 与之相关的物质--尿苷二磷酸半乳糖(UDPgal )是大肠杆菌细胞壁合成的前体。
而启动子也有两个: galP1起始的转录——无内源葡萄糖、有外源半乳糖时进行,以保证碳源的供应。
galP2起始的转录——有内源葡萄糖、无外源半乳糖时进行,以保证细胞壁的合成需要。
生理功能(可以理解为生物学意义?)无论从必要性和经济性考虑,都要有一个不依赖于cAMP-CAP 的启动子(s2) 进行本底水平的组成型合成,以及一个依赖于cAMP-CAP 的启动子(s1),进行高水平的调节,这样既可以满足细胞最基本的需要(细胞壁),又可以满足在没有葡萄糖而有半乳糖时,细胞能够利用半乳糖进行生长。
进一步解释:gal P2是不依赖于cAMP-CRP 的,相反: cAMP-CRP 对gal P2还起到一种抑制作用,这是因为其与结合位点的结合,会影响到RNA 聚合酶对gal P2的利用。
因此教材上(page257)认为:只有S2活性完全被抑制时,(S1)的调控作用才是有效的。
7.4.2 阿拉伯糖操纵子(arabinose operon)araB 基因、araA 基因和araD, 形成一个基因簇,简写为araBAD三个基因的表达受到ara 操纵子中araC 基因产物AraC 蛋白的调控。
C 蛋白有三个结合位点O2、O1和 I 。
I BADCRPO2O1C结构基因调节基因P BADaraC 基因是araBAD 的调节基因L核酮糖激酶L阿拉伯糖异构酶L核酮糖-5-磷酸-4-差相异构酶结合到ara I 的时候,由于araBAD的启动子本身与ara I有部分重叠,另外还可以引起上游序列回折弯曲,使得AraC同时与O2结合,从而使CRP 聚合酶也不能结合到启动子上,araBAD基因不转录。
分子生物学-2 chapter 7

7 : Bases
2. The histone N-terminal tails
The histone tails emerge from the core of the nucleosome at
Higher-order chromatin structure
Regulation of chromatin structure
Nucleosome assembly
Chromosomes: DNA
Shape: circular or linear problem Number in an organism is characteristic Copy: haploid, diploid, polyploid
Critical DNA elements in Chromosome duplication & segregation
Origins of replication Centromeres Telomeres
Figure 7-6 Centromeres, origin of replication and telomere are required for eukaryotic chrom. maintenance
DNA core DNA: 147 bp
linker DNA: 20-60 bp
Proteins Five abundant histones
Histones Structural fold N-terminal “tail”
Histones are small, positively charged (basic) proteins
Other proteins associated with the chromosome are referred to as non-histone proteins, including numerous DNA binding proteins that regulate the transcription, replication, repair and recombination of DNA.
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 1965年Khorana利用已知的核苷酸重复序列,以此多聚核苷酸作模 板,在体外进行蛋白质合成。
1、组成重复的RNA只能是UCU-CUC,翻译的多肽为丝氨酸、亮氨 酸排列,此处不能确定其密码子
2、怎么读都产生多聚苯、多聚丝、多聚亮。1与2只有一个密码 子相同,都有丝氨酸、亮氨酸,不能确定
3、怎么读都为四个密码子的循环,但合成的肽连中氨基酸有三 种。亮氨酸、苏氨酸、酪氨酸 3与2对照,彼此共有密码子CUU和亮氨酸,因此,亮氨酸码 子为CUU 2中已知CUU是亮氨酸,2与1对照丝氨酸一定为UCU,因为1、 2中都有丝氨酸。苯丙氨酸则为UUC
• 1961年Crick 和Brenner证明遗传密码是三联体密码子。
• 原黄素(二氨基吖啶)可以使DNA增加或减少一个碱基。
(-)(-)(-)
一次删去5’-GUA ↓ ↓ ↓ UAC GGA U………3’
读码框不变
1961年Nirenberg 的无细胞蛋白合成系统:
(1)去模板:用DNAase处理E.coli抽提物,使DNA降解,除去原有的细菌 模板。这样的抽提物中含有:
• TψC常由5bp的茎和7nt和环组成。此臂负责和核糖体上的rRNA 识别 结合;
• 反密码子臂(anticodon arm)常由5bp的茎区和7nt的环区组成,它负 责对密码子的识别与配对。
• D环 (D arm)的茎区长度常为4bp,含有二氢尿嘧啶(dihydrouracil)。 负责和氨基酰tRNA聚合酶结合;
二人获得1968年诺贝尔化学奖
遗传密码特点:
• 连续性:密码子之间是连续的,中间没有停顿。如果插入或删除一 个碱基将会发生错误,称为移码,由移码产生的突变,称为移码突 变。
• 不重叠性:一般情况下,密码子之间是相连的,之间没有重叠 (non-overlapping)。但有例外。密码子的重叠与重叠基因是不同 的概念。
• 起始tRNA:能特异性识别mRNA模板上起始密码子的tRNA;
• 延伸tRNA:其他tRNA统称为延伸tRNA。
• 同工tRNA:将携带相同氨基酸的不同tRNA称为同工tRNA。 在一个同工tRNA组内,所有tRNA均专一于相同的氨基酰tRNA合成酶。同工tRNA既要有不同的反密码子以识别该氨基 酸的各种同义密码,又要有某种结构上的共同性,能被AAtRNA合成酶识别。
• 1964年Nirenberg又采用三联体结合实验,发现
• 游离AA-tRNA,因分子较小,可以通过硝酸纤维素滤膜;
• 加入三核苷酸,使核糖体跟三联体结合后,AA-tRNA可以结合上来形 成大的复合体,无法通过滤膜而被滞留。所以可以将结合核糖体的 AA-tRNA与未结合的分开。
• 每次加入一种已知顺序的三联体(如ACA),再加入AA-tRNA,看哪 种AA-tRNA被留在滤膜上。
• 简并性:4种碱基可以组合成64种密码子,而生物体内有20种氨基酸, 故存在多个密码子(以2-4个居多)编码一种氨基酸的情况,称为密码 子的简并性(degeneracy)。编码同一种氨基酸的密码子称为同义密 码子。
• 摆动性:在密码子与反密码子的配对中,密码子的前两位严格遵守碱基 配对原则,第三对碱基有一定的自由度,可以“摆动”, 称为摆动性 (wobble)。
• 终止密码子: 通用UAG,UAA,UGA,不编码氨基酸。
第二节 TRNA
tRNA二级结构
• tRNA长度范围一般为70-80个碱基, 其中有22个恒定核苷酸。
• 5’端和3’端配对(常为7bp)形成茎区, 称为受体臂(acceptor arm)或称氨 基酸臂 。在3’端永远是4个碱基 (XCCA)的单链区,在其末端有2’OH或3’-OH,是被氨基酰化位点。此 臂负责携带特异的氨基酸。
第七章 翻译1
第一节 遗传密码
遗传密码的破译
• 1954年Gamow 首先对遗传密码进行探讨,指出遗传密码子应该是 三联体密码。
• 若一种碱基对应与一种氨基酸,那么只可能产生4种氨基酸; • 若2个碱基编码一种氨基酸的话,4种碱基共有42=16种不同的排列组
合; • 3个碱基编码一种氨基酸,排列组合可产生43=64种不同形式; • 若是四联密码,就会产生44=256种排列组合。
• 额外环(extra arm)可变性大,从4 nt到21 nt不等,其功能是在tRNA 的L型三维结构中负责连接两个区域(D环-反密码子环和TψC-受体 臂)。
tRNA三级结构
• D环和TψC环形成了“L” 的转角。
• 由在二级结构中未配对碱基间 形成的三级氢键氢键维持结构 稳定。
• 碱基平面之间的堆积力也是结 构稳定因素之一。
• 通用性:不同生物的密码子具有相对通用性,这也从一个侧面证明了生 物来源于共同祖先的假说。线粒体和纤毛虫的密码子有特殊性。
• 偏好性:不同生物对同义密码子有一定的偏好。那些不被经常利用的同 义密码称为稀有密码。
• 起始密码子: 通用AUG(同时编码Met或fMet),少数细菌中使用 GUG(编码Val)。
反密码子
• tRNA上有三个特定的碱基组成反密码子,与密码子互补配对,方 向相反。
• 反密码子的5’碱基在与密码子配对时具有摆动性。
ห้องสมุดไป่ตู้
副密码子
• tRNA上存在一些特殊部位,能够 保证tRNA与特定的氨基酸结合, 这些部位称为鉴别元件或副密码 子。不同的tRNA的鉴别元件差异 很大。
• tRNA分类
第三节 核糖体
核糖体
• 由蛋白质和rRNA组成。核糖体是 翻译的场所,发挥着重要作用。
• 核糖体可游离存在,真核中,也 可同内质网结合,形成粗糙的内 质网。原核中,与mRNA形成多 聚核糖体
核糖体的功能位点
• P位点(肽酰tRNA 位点,peptidyl-tRNA site)与携带新生多肽链的 tRNA结合。
核糖体、ATP、氨基酸,mRNA(因半衰期短,很快被降解掉)
(2)加入尿苷二磷酸,人工合成poly(U):合成了多聚苯丙氨酸 这一结果不仅证实了无细胞系统的成功,同时还表明UUU是苯丙氨酸的 密码子。
(3)分别加入polyA,polyC和polyG,相应地获得了多聚赖氨酸,多聚脯氨 酸和多聚甘氨酸。