糖的合成代谢
生物化学 糖代谢

6 ATP
第三阶段:三羧酸循环
2*异柠檬酸→2*α -酮戊二酸 2*α -酮戊二酸 →2*琥珀酰CoA
辅酶
NAD+ NAD+ FAD
ATP
2*3 2*3
2*琥珀酰CoA →2*琥珀酸
2*琥珀酸→2*延胡索酸
2*1
2*2
2*苹果酸→2*草酰乙酸
NAD+
2*3
24ATP
总ATP数: 第一阶段——6或8 第二阶段——6 第三阶段——24 36 或 38ATP
活性受NADP+/NADPH比值的调节,NADPH能强烈
抑制6-磷酸葡萄糖脱氢酶。磷酸戊糖途径的流
量取决于机体对NADPH的需求。
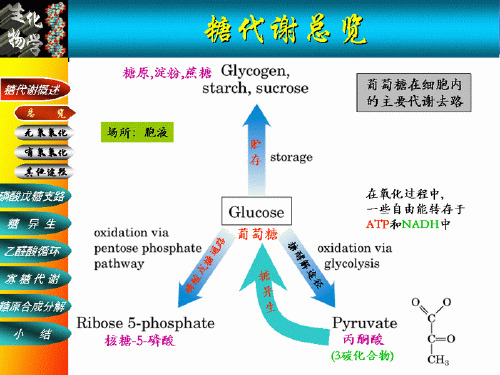
• 概念:有氧,葡萄糖(糖原) → CO2 + H2O • 反应部位:细胞液、线粒体 cytoplasm mitochondria
+ ATP
有氧氧化的概况
有氧氧化的反应过程
• 第一阶段:葡萄糖→ →丙酮酸(胞液) • 第二阶段:丙酮酸→ →乙酰CoA (线粒体) • 第三阶段:乙酰CoA → →CO2 + H2O + ATP (三羧酸循环)(线粒体)
植物和某些藻类能够利用太阳能,将二氧化碳和水合成
糖类化合物,即光合作用。光合作用将太阳能转变成化 学能(主要是糖类化合物),是自然界规模最大的一种 能量转换过程。
一、多糖和低聚糖的酶促降解
1.概述 多糖和低聚糖只有分解成小分子后才 能被吸收利用,生产中常称为糖化。 2. 淀粉
3.淀粉水解 淀粉 糊精
7.无氧发酵 (Fermentation)
⑴乙醇发酵
COOH C CH3
CO2
糖的合成代谢

糖的合成代谢糖的合成代谢是生物体内繁重且至关重要的生物化学过程之一。
在有氧条件下,合成代谢主要涉及两个方面的过程:糖异生和糖原合成。
这两个过程基本上体现了糖的生物合成和降解的动态平衡。
糖异生是指机体从非糖高碳化合物中,在无氧或低氧情况下产生糖的过程。
在这个过程中,生物体通过解释质、谷氨酸、丙酮酸等物质,生成新的葡萄糖,以供进行能量代谢。
糖异生过程涉及的酶和复杂的调节机制,为机体提供了在紧急情况下保持能量平衡的手段。
糖原合成是通过糖原的合成酶将多个葡萄糖分子的简单单元沟成一个大分子的过程。
这个过程主要发生在肝脏和骨骼肌中,以便在高强度的长时间运动或长时间饥饿的情况下提供充足的营养支持。
糖的代谢主要存在于肝脏、骨骼肌和脂肪组织中。
在肝脏中有一个中枢机构,称为肝酸酯化酶,它能够协调糖异生和糖原合成的过程。
在糖异生过程中,肝酸酯化酶将解释质转化为聚糖,并导致糖原的合成。
而当需要糖分进行能量代谢时,肝酸酯化酶会在葡萄糖水平下降时释放糖原。
当血糖水平过低时,胰岛素的释放也会减慢,从而促进肝脏释放糖原并协助糖异生。
而在血糖过高的情况下,胰岛素将促进肝脏中糖原的合成和葡萄糖的上传。
糖的合成代谢对生物体的能量平衡至关重要。
当机体还有足够的营养储备时,合成代谢将持续进行,并促进能量储存。
而当机体处于饥饿状态时,糖异生和糖原合成的过程将被激活,以获得额外的能量支持。
总结来说,糖的合成代谢是生物体通过从非糖高碳化合物中合成糖或将多个葡萄糖分子的简单单元合成为一个大分子的生物化学过程。
这个过程涉及复杂的酶和调节机制,对于生物体的能量平衡至关重要。
第十一章 糖类代谢--王镜岩《生物化学》第三版笔记(完美打印版)

反应放能,在生理条件下不可逆(K大于300)。由己糖激酶或葡萄糖激酶催化,需要Mg2+或Mn2+。己糖激酶可作用于D-葡萄糖、果糖和甘露糖,是糖酵解过程中的第一个调节酶,受6-磷酸葡萄糖的别构抑制。有三种同工酶。葡萄糖激酶存在于肝脏中,只作用于葡萄糖,不受6-磷酸葡萄糖的别构抑制肌肉的己糖激酶Km=0.1mM,肝脏的葡萄糖激酶Km=10mM,平时细胞中的葡萄糖浓度时5mM,只有进后葡萄糖激酶才活跃,合成糖原,降低血糖浓度,葡萄糖激酶是诱导酶,胰岛素可诱导它的合成。6-磷酸葡萄糖也可由糖原合成,由糖原磷酸化酶催化,生成1-磷酸葡萄糖,在磷酸葡萄糖变位酶的催化下生成6-磷酸葡萄糖。此途径少消耗1个ATP。6-磷酸葡萄糖由葡萄糖6-磷酸酶催化水解,此酶存在于肝脏和肾脏中,肌肉中没有。
三、能量变化
C6H12O6+2Pi+2ADP+2NAD+=2C3H4O3+2ATP+2NADH+2H++2H2O
有氧时2个NADH经呼吸链可产生6个ATP,共产生8个ATP;无氧时生成乳酸,只有2个ATP。在骨骼肌和脑组织中,NADH进入线粒体要经过甘油磷酸穿梭系统,在细胞质中由3-磷酸甘油脱氢酶催化,将磷酸二羟丙酮还原生成3-磷酸甘油,进入线粒体后再氧化生成磷酸二羟丙酮,返回细胞质。因为其辅酶是FAD,所以生成FADH2,只产生2个ATP。这样其还原当量(2H++2e)被带入线粒体,生成FADH2,进入呼吸链,结果共生成6个ATP。
二、糖的消化和吸收
(一)消化
淀粉是动物的主要糖类来源,直链淀粉由300-400个葡萄糖构成,支链淀粉由上千个葡萄糖构成,每24-30个残基中有一个分支。糖类只有消化成单糖以后才能被吸收。
糖的生物合成与代谢途径

糖的生物合成与代谢途径糖是生命中不可或缺的重要物质,它是生物体的主要能量来源之一,也是构成生物体的重要组成部分。
糖的生物合成与代谢是一系列复杂而精细的过程,它们通过一定的途径在细胞内进行。
在本文中,我们将探讨糖的生物合成与代谢的主要途径和相关机制。
第一节糖的生物合成糖的生物合成是细胞利用光能或化学能将无机物合成糖类化合物的过程。
主要的合成途径有光合作用和糖异生两种形式。
光合作用是指细胞通过叶绿体内的光化学反应,将二氧化碳和水转化为葡萄糖等有机物的过程。
在光照条件下,叶绿体中的叶绿素可以吸收太阳能,光合色素体可将太阳能转化为化学能,进而促使光合作用的进行。
光合作用分为光反应和暗反应两个阶段。
光反应发生在光合体系中,通过光合色素体捕捉光能,产生氧化还原电位,将光能转化为高能物质膜内的质子激励。
暗反应指的是光合作用中的还原和碳固定反应,主要在叶绿体基质内进行。
通过一系列酶的作用,将光反应所得的ATP和NADPH利用碳源还原为葡萄糖或其他有机物。
糖异生是指细胞在无光照条件下,通过有机物合成糖的过程。
糖异生主要发生在细胞质基质内,包括糖异生途径的两个重要过程:糖酵解和有机酸循环。
糖酵解是指将葡萄糖分解为丁醛酸,再将丁醛酸氧化为甲酸,最终合成糖的过程。
有机酸循环是指细胞质基质内的一系列反应,将葡萄糖分解为丙酮酸、柠檬酸等有机酸,最终通过一系列酶的作用合成糖。
第二节糖的代谢途径糖的代谢指的是细胞对糖化合物进行分解和利用的过程。
糖的代谢途径包括糖酵解、糖异生和糖氧化三个主要途径。
糖酵解是指细胞内部一系列酶的作用,将葡萄糖分解为丙酮酸或乙酸,产生ATP和还原能力分子NADH的过程。
糖酵解包括糖原糖酵解和异物糖酵解两种形式。
糖原糖酵解是指细胞内糖原被酵解,通过一系列的反应将糖原分解为葡萄糖,再进一步分解为丙酮酸,转化为乙酸最终释放能量。
异物糖酵解是指细胞利用外源性的碳水化合物,如蔗糖、木糖等进行糖酵解的过程。
糖异生是指细胞利用非糖类有机物合成糖的过程。
糖的合成与代谢

糖的合成与代谢糖是一种重要的碳水化合物,在生物体内扮演着多种角色。
糖的合成与代谢过程是维持生物体正常功能的关键部分。
本文将围绕糖的合成与代谢展开讨论。
一、糖的合成糖的合成主要通过光合作用进行。
在光合作用中,光能被转化为化学能,用于合成葡萄糖等有机物质。
光合作用发生在光合细胞中的叶绿体内,其中最重要的反应是光合系统I和光合系统II的光反应和暗反应。
光合作用的光反应阶段发生在叶绿体的基质内膜上,通过光能将水分解为氧气、电子和质子。
电子随后被传递给光合色素,并在其中形成高能化合物。
这些高能化合物经过一系列的反应,最终使ADP和磷酸根结合生成ATP,这是光合作用产生的化学能。
同时,质子也积累在基质内膜上,形成质子梯度。
光合作用的暗反应阶段是在基质内膜上进行的。
此阶段中,ATP和NADPH被利用来合成糖类物质。
暗反应主要是通过卡尔文循环进行的,其中CO2被固定为糖酮磷酸。
随后,糖酮磷酸经过一系列反应,最终合成葡萄糖。
二、糖的代谢糖的代谢主要包括糖酵解和细胞呼吸两个过程。
1. 糖酵解糖酵解是在无氧环境下进行的代谢过程,产生乳酸或乙醇和二氧化碳。
糖酵解可以分为三个阶段:糖的准备阶段、糖的裂解阶段和乙酸的产生阶段。
糖的准备阶段是将葡萄糖转化为两个分子的三碳糖类物质。
糖的裂解阶段是将三碳糖类物质分解为两个分子的丙酮酸。
乙酸的产生阶段是将两个分子的丙酮酸经过一系列反应,最终转化为乙酸。
2. 细胞呼吸细胞呼吸是在有氧条件下进行的代谢过程,将葡萄糖完全氧化为CO2和H2O,同时产生大量ATP。
细胞呼吸包括三个阶段:糖的燃烧、三羧酸循环和电子运输链。
糖的燃烧阶段是将葡萄糖和氧气进行直接反应,生成CO2、H2O和ATP。
三羧酸循环是将糖类物质逐步分解为二氧化碳,并释放能量。
电子运输链是将通过糖的裂解和三羧酸循环生成的高能电子转移到氧气上,生成水和额外的ATP。
细胞呼吸是产生ATP和维持有氧呼吸的重要过程,是生物体内能量供应的主要途径。
生物化学第八章糖代谢

§2 糖的分解代谢
主要有以下途径: (一)糖的无氧酵解 (二)糖的有氧氧化 (三)乙醛酸循环 (四)戊糖磷酸途径
途径具体过程
提示
反应实质 个酶作用 进程变化 学习途径时要重点注意噢!
温馨提示
加油!!!
• 酵解过程要学好
• 首条途径很重要 • 总结经验找规律 • 后边学习基础牢
• 举一反三相比较 • 触类旁通有参照 • 事半功倍学的巧 • 一路轻松兴趣高
甘油酸-3-磷酸
磷酸甘油8反酸应变图位酶
甘油酸-2-磷酸
9、2-磷酸甘油酸脱水烯醇化
甘油酸-2-磷酸
烯醇化9反酶应图
磷酸烯醇式丙酮酸
9、2-磷酸甘油酸的脱水生成磷酸烯醇式丙 酮酸
烯醇化酶(enolase) 这一步反应也可看作分子内氧化还原反应,分子 内能量重新分布,又一次产生了高能磷酯键。
反应可以被氟离子抑制,取代天然情况下酶分 子上镁离子的位置,使酶失活。
细胞核
内质网 溶酶体
细胞膜
动物细胞
植物细胞
细胞壁 叶绿体
有色体 白色体 液体 晶体
葡萄糖的主要代谢途径
糖异生
葡萄糖
6-磷酸葡萄糖 (有氧或无氧)
(无氧) 丙酮酸
糖酵解
(有氧)
乳酸 乙醇
乙酰 CoA
磷酸戊糖 途径
三羧酸 循环
第八章:糖代谢
§1 多糖和底聚糖的酶促降解 §2 糖的分解代谢 §3 糖的合成代谢
⑹氧化脱氢,产生 NADH+H+ (磷酸化,使用无机磷酸)
甘油醛-3-磷酸
无机磷酸
甘油醛-3-磷酸 脱氢酶
1,3-二磷酸甘油酸
产生 的 NADH+H+ 的氢,条件不同, H的去向不同,走进的途径不同。
糖代谢的概况 (一)分解代谢:主要途径:1 糖酵解(糖的

不活跃的磷酸化的丙酮酸激酶
H2 O
和甘油醛-3-磷酸总是处于平衡状态,但由于甘油醛-3-磷酸在
酵解途径中不断被消耗,因此,反应得以向生成甘油醛-3-磷酸
反向202进1/5/行14 ,实际最后生成两分子甘油醛-3-磷酸。
(六)甘油醛-3-磷酸氧化成1,3-二磷酸甘油酸
生成1分子 NADH+H+
形成1个高能磷 酸键
3-甘油醛磷酸 脱氢酶
O=C—O—As—O–
–
水解
1-砷酸-3-磷酸甘油酸
O=C—OH
+ 3-磷酸甘油酸
–O—As—O–
–
在有砷酸盐存在的情况下,酵解过程可以照样进行下去,但不能形成高能磷酸 键,即20砷21/酸5/14盐起着解偶联作用,解除了氧化和磷酸化的偶联作用。
(七)1,3-二磷酸甘油酸转移高能磷酸键基团 形成ATP
2021/5/14
三、糖酵解的意义
1、糖酵解是存在一切生物体内糖分解代谢的普遍 途径。
2、通过糖酵解使葡萄糖降解生成ATP,为生命活 动提供部分能量,尤其对厌氧生物是获得能量 的主要方式。
3、糖酵解途径的许多中间产物可作为合成其他物 质的原料(提供碳骨架),如磷酸二羟丙酮 甘油。
4、是糖有氧分解的准备阶段。 5、由非糖物质转变为糖的异生途径基本为之逆过
• 1940年被阐明。(研究历史) Embden,Meyerhof,Parnas等人贡献最多, 故糖酵解过程一也叫Embdem-MeyerhofParnas途径,简称EMP途径。
• 在细胞质中进行
2021/5/14
糖酵解的研究历史:
• 应追溯到4000年前的制酒工业。(发酵过程)
• 1854-1864年,Louis Paster的观点占统治地位:认
动物生物化学 第六章 糖的代谢

2. 糖原的 合成
(UDP-葡萄 糖焦磷酸化 酶、糖原合 成酶、糖原 分支酶)
糖原合成酶催化的反应
糖原的合成与分解总反应示意图
3. 糖原代谢的调节
• 葡萄糖分解代谢总反应式 • C6H6O6 + 6 H2O + 10 NAD+ + 2 FAD + 4 ADP +
4Pi 6 CO2 + 10 NADH + 10 H+ + 2 FADH2 + 4 ATP • 按照一个NADH能够产生3个ATP,1个FADH2能够产 生2个ATP计算,1分子葡萄糖在分解代谢过程中共产 生38个ATP: • 4 ATP +(10 3)ATP + (2 2)ATP = 38 ATP
Байду номын сангаас
CH2OH CO
HO C H
CHO
H C OH + H C OH
H C OH H C OH
CH2O P
转醛酶
CH2O P
7-磷酸景天庚酮糖 3-磷酸甘油醛
CHO
H C OH +
H C OH CH2O P
4-磷酸赤藓糖
CH2OH CO HO C H HO C H H C OH CH2O P
6-磷酸果糖
H
O
H
OH H HO
H OH
H2O
H C OH
HO C H
O 内酯酶
H C OH
H C OH
G-6-P
6-磷酸葡萄 糖酸内酯
CH2O P 6-磷酸葡萄糖酸
COOH H C OH
NADP+
+ NADPH + H
糖的合成代谢

固氮酶的厌氧微环境:固氮酶的两个蛋白组分对氧是 极端敏感的,而且一旦接触氧就很快导致不可逆失活。 大多数的固氮菌都是好氧菌,它们需要利用氧气进行 呼吸和产生能量。 固氮菌发展出多种机制来解决其既需要氧又须防止氧对 固氮酶损伤的矛盾:呼吸保护、构象保护、蓝细菌异形胞、 豆血红蛋白、不同时间进行固氮作用与光合作用,等。
乙醇酸、草酸、甘氨酸
乳酸 谷氨酸、天冬氨酸 亮氨酸
甘油酸途径
氧化 脱氨基 降解
甘油醛-3-磷酸
丙酮酸 α-酮戊二酸、草酰乙酸 丙酮酸
3. 多糖的合成
微生物中的多糖可分为同型多糖和杂多糖。同型多糖是由相 同单糖分子聚合而成,如糖原、纤维素、甲壳素等。杂多糖 是由不同单糖分子聚合而成,如肽聚糖、脂多糖等。 微生物多糖合成的特点: ①不需要模板,而是由转移酶的特异性来决定亚单位在多聚 链上的次序。 ②合成的开始阶段需要引物,引物通常由小片断多糖充当。 ③多糖合成时,由糖核苷酸作为糖基载体,将单糖分子转移 到受体分子上,使多糖链逐步加长。
2)厌氧乙酰-辅酶A途径
自养微生物的CO2还原途径:1分子CO2被还原成甲醇水平,另一分 子CO2被还原成CO,二者合成产生乙酰-CoA,经丙酮酸合成酶催化由 乙酰-CoA接受第3个CO2分子生成丙酮酸,用于合成各种有机物。
3)还原性TCA循环途径
CO2通过琥珀酰-CoA的还原性羧化生成-酮戊二酸而被固定
(2)化能自养型微生物的生物氧化和产能
化能自养型微生物中,其ATP是通过氧化还原态无机物 产生的,其NAD(P)H2是通过消耗ATP将无机氢(H++e)逆 呼吸链传递产生的。
化能自养型微生物均为细菌,且绝大多数为好氧菌 能量代谢特点: 脱氢酶或氧化还原酶催化无机底物脱氢或脱电子; 无机底物脱下的氢直接进入呼吸链,通过氧化磷酸化产能; (少数菌在无机硫化物存在时,能部分通过底物磷酸化产能) 氢或电子可从多处进入呼吸链,所以,呼吸链多样; 由于从中间进入呼吸链,因此产能效率低,菌体生长缓慢,细 胞产率低。
第七章 糖代谢

甘油醛-3-磷酸脱氢酶的作用机理: 甘油醛-3-磷酸脱氢酶由4个相同亚基组成,每个亚 基牢固地结合一个分子的NAD+,并能独立参与催化作 用。亚基中第149位的半胱氨酸残基的-SH是活性基团, NAD+的吡啶环与活性-SH基很近,共同组成酶的活性 部位。
甘油酸-1,3-二磷酸将磷酰基转给ADP形成了磷酸甘油酸和 ATP,催化这个反应的酶是磷酸甘油酸激酶 。
第七章 糖代谢
生物化学
教 学 内 容
多糖和低聚糖的酶促降解 糖的分解代谢 糖的合成代谢
糖类的重要的生物学功能
供给能量:糖的主要功能是供给能量,人体所需
能量的70%以上是由糖氧化分解供应的。1克葡萄糖在 体内完全氧化分解,可释放能量16.7千焦。 供给碳源:糖分解过程中形成的中间产物可以提 供合成脂类和蛋白质等物质所需要的碳架。 构成组织细胞的重要组成成分:如核糖和脱氧 核糖是细胞中核酸的成分;糖与脂类形成的糖脂是组 成神经组织与细胞膜的重要成分;糖与蛋白质结合的 糖蛋白,具有多种复杂的功能。
第一节 多糖和低聚糖的酶促降解
教 学 内 容
淀粉的酶促降解 糖原的降解 纤维素的酶促降解 双糖的酶水解
一、淀粉的酶促降解
淀粉的种类:有直链淀粉和支链淀粉两类。
淀粉的水解:
α-淀粉酶:又称α-1,4-葡萄糖水解酶。其作用方式是从淀 粉分子的内部,随机水解分子内的α-1,4-糖苷键,若底物 是直链淀粉,生成葡萄糖、麦芽糖、麦芽三糖等混合物。 如果底物是支链淀粉,则水解产物中有葡萄糖、麦芽糖和 α-糊精等混合物。 β-淀粉酶:又称β-1,4-麦芽糖苷酶。此酶具有外切酶的特 性,能专一地从直链淀粉或支链淀粉外层的非还原性末端, 依次切下两个葡萄糖单位(即麦芽糖)。 α-1,6糖苷酶:支链淀粉分子中的α-1.6糖苷键需要由α-1,6 糖苷酶作用,如植物中的R酶和动物小肠中的α-糊精酶, 其作用方式是从支链淀粉的外部开始,将α-1,6糖苷键水解 掉,其产物是由α-1,4糖苷键组成的直链片段。 麦芽糖酶:麦芽糖酶可催化麦芽糖水解成葡萄糖。
第七章 糖代谢
K2=3250
在植物光合组织中蔗糖磷酸合酶的活性较高,而非光合组 织中蔗糖合酶的活性较高。这是目前认为可能在光合组织中合 成蔗糖的主要途径。
(二)淀粉的合成:
存在于植物体内,尤其是谷类、豆类、薯类 作物的籽粒和贮藏组织都含丰富的淀粉。
淀粉合成中的糖基供体有ADPG、UDPG, 主要是ADPG。
合成分两阶段进行,先合成直链淀粉,然后 分支形成支链淀粉。
二、反应过程 反应可分为两个阶段: 第一阶段:氧化阶段,生成NADPH+H+和 CO2;由6-磷酸葡萄糖直接脱氢脱羧生 成磷酸戊糖; 第二阶段:非氧化阶段,一系列基团转 移反应;磷酸戊糖分子再经重排最终又 生成6-磷酸葡萄糖。
第一阶段:氧化阶段
1、脱氢反应:6-磷酸葡萄糖脱氢酶以NADP+ 为辅酶,催化6-磷酸葡萄糖脱氢生成6-磷酸 葡萄糖酸δ内酯,不可逆。
生物合成的供氢体
脂肪酸、胆固醇和类固醇化合物 的生物合成,均需要大量的NADPH。
0 R-CH2-C-R’
=
OH R-CH2-CH-R’ NADP+
R-CH2-CH2-R’
NADPH + H+
H R-C=C-R’
3、磷酸戊糖途径与疾病
神经精神病
(neuropsychiatric disorder)
第六节 糖的合成代谢
一、光合作用 二、糖异生途径 三、蔗糖和多糖的生物合成
一、光合作用
• 光合作用是糖合成代谢的主要途径。 • 绿色植物、光合细菌或藻类等将光能转变成化学 能的过程,即利用光能,由CO2和H2O合成糖类化 合物并释放出氧气的过程,称为光合作用。 • 光合作用的总反应式可表示如下: 光能 • n CO2 + n H2O (CH2O)n + n O2 叶绿体 糖类化合物
生物化学糖代谢小结

糖代谢知识要点(一)糖酵解途径:糖酵解途径中,葡萄糖在一系列酶的催化下,经10 步反应降解为2 分子丙酮酸,同时产生2 分子NADH+H和2 分子ATP。
主要步骤为:(1)葡萄糖磷酸化形成二磷酸果糖;(2)二磷酸果糖分解成为磷酸甘油醛和磷酸二羟丙酮,二者可以互变;(3)磷酸甘油醛脱去2H 及磷酸变成丙酮酸,脱去的2H 被NAD所接受,形成NADH+H。
(二)丙酮酸的去路:(1)有氧条件下,丙酮酸进入线粒体氧化脱羧转变为乙酰辅酶A,同时产生1 分子NADH+H。
乙酰辅酶A 进入三羧酸循环,最后氧化为CO和HO。
(2)在厌氧条件下,可生成乳酸和乙醇。
同时NAD得到再生,使酵解过程持续进行。
(三)三羧酸循环:在线粒体基质中,丙酮酸氧化脱羧生成的乙酰辅酶A,再与草酰乙酸缩合成柠檬酸,进入三羧酸循环。
柠檬酸经脱水加水转变成异柠檬酸,异柠檬酸经连续两次脱羧和脱羧生成琥珀酰CoA;琥珀酰CoA 发生底物水平磷酸化产生1 分子GTP 和琥珀酸;琥珀酸再脱氢,加水及再脱氢作用依次变成延胡索酸,苹果酸及循环开始的草酰乙酸。
三羧酸循环每循环一次放出2 分子CO,产生3 分子NADH+H和一分子FADH。
(四)磷酸戊糖途径:在胞质中,在磷酸戊糖途径中磷酸葡萄糖经氧化阶段和非氧化阶段被氧化分解为CO,同时产生NADPH + H。
其主要过程是G-6-P 脱氧生成6-磷酸葡萄糖酸,再脱氢,脱羧生成核酮糖-5-磷酸。
6 分子核酮糖-5-磷酸经转酮反应和转醛反应生成5 分子6-磷酸葡萄糖。
中间产物甘油醛-3-磷酸,果糖-6-磷酸与糖酵解相衔接;核糖-5-磷酸是合成核酸的原料,4-磷酸赤藓糖参与芳香族氨基酸的合成;NADPH+H提供各种合成代谢所需要的还原力。
(五)糖异生作用:非糖物质如丙酮酸,草酰乙酸和乳酸等在一系列酶的作用下合成糖的过程,称为糖异生作用。
糖异生作用不是糖酵解的逆反应,因为要克服糖酵解的三个不可逆反应,且反应过程是在线粒体和细胞液中进行的。
糖的合成代谢

糖的合成代谢介绍糖是生命体内最重要的能量来源之一。
在细胞内的代谢过程中,糖的合成和分解密切相关,这个过程被称为糖的合成代谢。
糖的合成代谢不仅仅与能量供应有关,还与细胞功能、生长发育、信号传导等多个方面密切相关。
本文将深入探讨糖的合成代谢的相关内容。
糖的来源在细胞内,糖可以从外部摄入进入体内,也可以通过内源性途径合成。
外源性糖主要来自食物,例如葡萄糖、果糖等。
内源性糖的合成则涉及到细胞的代谢过程。
糖的合成途径糖异生途径糖异生是指细胞通过非糖类物质合成糖的过程。
这个过程发生在多个组织和器官中,如肝脏、肾脏等。
糖异生途径有多个关键步骤,其中最主要的是糖异生途径的前期和后期。
前期主要包括糖异生途径的启动和前体物质的合成,后期主要是糖异生途径的继续以及糖的合成。
糖原代谢糖原是多个葡萄糖分子通过α-(1→4)键和α-(1→6)键连接而成的聚糖,是动物体内主要的糖贮存形式。
糖原的合成和分解是糖原代谢的关键步骤。
在低血糖状态下,肝脏中的糖原分解为葡萄糖,释放到血液中供给全身各个组织。
同时,在高血糖状态下,胰岛β细胞分泌胰岛素,促进葡萄糖的合成和储存为糖原。
糖合成的调控糖的合成代谢在细胞内受到多个因素的调控,以维持机体内糖代谢的平衡。
以下是糖合成的调控机制的一些重要方面:激素调控激素在糖合成代谢中发挥着重要的作用。
胰岛素是降低血糖的主要激素,它促进糖的合成和储存,同时抑制糖异生和糖原分解。
而对于提高血糖的肾上腺素、胰高血糖素等激素,则抑制糖的合成和储存,促进糖异生和糖原分解。
细胞内信号传导调控细胞内信号传导通路对糖的合成代谢也有重要作用。
例如,糖酵解的速率可以通过磷酸化酶的调节而受到精细调控,而糖异生的调节则受到乙酰辅酶A羧化酶的磷酸化控制。
反馈抑制在糖的合成代谢中,存在很多反馈抑制机制。
一旦合成产生足够的糖,会触发反馈抑制机制,抑制糖的进一步合成。
例如,高浓度的葡萄糖和ATP可以抑制磷酸果糖激酶活性,从而抑制糖异生途径。
糖原的分解合成代谢

腺苷环化酶
腺苷环化酶(有活性)
(无活性) ATP
cAMP
PKA
(无活性)
PKA
(有活性)
磷酸化酶b激酶 磷酸化酶b激酶-P
Pi
磷蛋白磷酸酶-1
–
糖原合酶
糖原合酶-P
磷酸化酶b 磷酸化酶a-P
Pi
糖原的分解合成代谢
磷蛋白磷酸酶-1
Pi
–
PKA(有活性)
磷蛋白磷酸酶-1
–
磷蛋白磷酸酶抑制剂-P
磷蛋白磷酸酶抑制剂
葡萄糖-6-磷酸酶(肝,
肾)
葡萄糖-6-磷酸
葡萄糖
葡萄糖-6-磷酸酶只存在于肝、肾中,而不存在
于肌中。所以只有肝和肾可补充血糖;而肌糖原不
能分解成葡萄糖,只能进行糖酵解或有氧氧化。
糖原的分解合成代谢
第15页
肌糖原分解
➢ 肌糖原分解前三步反应与肝糖原分解过程相同, 不过生成6-磷酸葡萄糖之后,因为肌肉组织中 不存在葡萄糖-6-磷酸酶,所以生成6-磷酸葡萄 糖不能转变成葡萄糖释放入血,提供血糖,而 只能进入酵解路径深入代谢。
合成部位: 组织定位:主要在肝脏、肌肉 细胞定位:胞浆
糖原的分解合成代谢
第4页
糖原合成路径:
1.葡萄糖磷酸化生成葡萄糖-6-磷酸
ATP
ADP
葡萄糖 己糖激酶;
葡萄糖-6-磷酸
葡萄糖激酶(肝)
糖原的分解合成代谢
第5页
2.葡萄糖-6-磷酸转变成葡萄糖-1-磷酸
磷酸葡萄糖变位酶
葡萄糖-6-磷酸
葡萄糖-1-磷酸
➢ 当糖原合成路径活跃时,分解路径则被抑制, 才能有效地合成糖原;反之亦然。
糖原的分解合成代谢
第三节糖的合成代谢

第四节 糖的合成代谢
Biosynthesis of carbohydrates
1. 光合作用(photosynthesis)
光反应 (photo reaction) 光合作用分成两个部分
暗反应 (dark reaction)
基质 类囊体
非还非原还端原7糖端残7糖基残被基切下
Glycogen-branching enzyme
非还原端11个糖基
非还原端7糖残基被切下
7糖残基的还原端被转移到同一或另一相 邻糖链距分枝点或核心第4个糖残基上
Glycogen-branching enzyme
△G0’ = -15.5 kJ/mol
非还原端11个糖基
O2 2H+ + 2e
Actived chlorophyl H2O
NADP
+
ADP
ATP
(CH2O)
NADPH•H+ CO2
三碳循环 — Calvin循环
CO2固定的方式:
四碳循环 — Hatch-Stack 途径
①
逆HMP途径
The initial CO2 fixation is catalyzed by ribulose 1,5bisphosphate carboxylase/oxygenase
• The enzyme is present on the lumen side of the ER membrane of hepatocytes (肝细胞) and renal (肾的) cells.
• The enzyme is not present in muscle or brain cells, where gluconeogenesis does not occur.
糖 代谢

(1)低血糖是指血糖浓度<3.33mmol/L
空腹血糖浓度低于3.33~3.89mmol/L时称为低血糖(hypoglycemia) 。血 糖水平过低会影响脑细胞功能,出现 头晕、倦怠无力、心悸等症状, 严重时出现昏迷,称为低血糖休克。
低血糖的病因有: ① 胰性(胰岛β-细胞功能亢进、胰岛α-细胞功能低 下等);② 肝性(肝癌、糖原积累病等);③ 内分泌异常(垂体功能 低下、肾上腺皮质功能低下等);④ 肿瘤(胃癌等);⑤ 饥饿或不能 进食;
无氧代谢不能将葡萄糖完全分解为二氧化碳,部分能量仍积累在其 代谢产物中; 有氧代谢通过呼吸链将葡萄糖完全氧化为二氧化碳和水 ,可将葡萄糖的能量全部释放出来为生物体利用;
有氧氧化是糖分解代谢的主要途径。
重要概念
糖酵解(glycolysis):一分子葡萄糖裂解为两分子丙酮酸的过程。 乳酸发酵(lactic acid fermentation):在缺氧条件下,葡萄糖经酵解生 成的丙酮酸还原为乳酸(2-羟基丙酸,lactate) 。 乙醇发酵(ethanol fermentation):在某些植物、脊椎动物组织和微生 物,酵解产生的丙酮酸转变为乙醇和CO2,即乙醇发酵。(丙酮酸脱羧产生 乙醛,乙醛在醇脱氢酶催化下被NADH还原成乙醇) 有氧氧化(aerobic oxidation):在有条件下,需氧生物和哺乳动物组织 内的丙酮酸彻底氧化分解为CO2和H2O,即糖的有氧氧化 。
糖代谢异常与临床疾病
(一)先天性酶缺陷导致糖原累积症
糖原累积症(glycogen storage disease)是一类遗传性代谢病,其 特点为体内某些器官组织中有大量糖原堆积。
引起糖原累积症的原因是患者先天性缺乏与糖原代谢有关的酶类。
糖原积累症分型
糖的化学分解与合成代谢

(二)糖的吸收
1. 吸收部位
小肠上段
2. 吸收形式
单糖
3. 吸收机制
刷状缘 肠 腔
Na+
G
小肠粘膜细胞
ATP ADP+Pi Na+泵
细胞内膜 门静脉
K+
Na+依赖型葡萄糖转运体
(Na+-dependent glucose transporter, SGLT)
4. 吸收途径 小肠肠腔 SGLT 肠粘膜上皮细胞
消化部位: 主要在小肠,少量在口腔
消化过程
口腔 胃 肠腔
淀粉
唾液中的α-淀粉酶 胰液中的α-淀粉酶
肠粘膜 上皮细胞 刷状缘
麦芽糖+麦芽三糖 α-临界糊精+异麦芽糖 (40%) (25%) (30%) (5%)
α-葡萄糖苷酶
α-临界糊精酶
葡萄糖
食物中含有的大量纤维素,因人体 内无-糖苷酶而不能对其分解利用,但却 具有刺激肠蠕动等作用,也是维持健康 所必需。
1,6-双磷酸果糖
CH2 O P
C O 磷酸二羟丙酮
C H 2O H
+
CHO
C H O H 3-磷酸甘油醛 CH2 O P
磷酸烯醇式丙酮酸
ADPቤተ መጻሕፍቲ ባይዱ
ATP
丙酮酸
Glu
ATP
ADP
G-6-P
⑸ 磷酸丙糖的同分异构化
F-6-P
ATP ADP
F-1,6-2P
CH2 O P
CHO
磷酸二 3-磷酸 羟丙酮 甘油醛
2-磷酸甘油酸
磷酸烯醇式丙酮酸
ADP
ATP
丙酮酸
己糖异构酶
6-磷酸果糖 (fructose-6-phosphate, F-6-P)
第七章 糖代谢—糖的合成

糖异生的生理意义
(一)维持血糖浓度恒定 (二)补充肝糖原,回收乳酸能量(乳酸循环) 补充肝糖原,回收乳酸能量(乳酸循环) (三)协助氨基酸代谢 (四)调节酸碱平衡(乳酸异生为糖) 调节酸碱平衡(乳酸异生为糖)
五、糖代谢的紊乱
糖尿病:体内胰岛素缺乏,血糖分解缓慢, 糖尿病:体内胰岛素缺乏,血糖分解缓慢,导致血糖过高由尿 排出。患者的肝糖原合成和糖的氧化能力均降低,脂代谢增加 排出。患者的肝糖原合成和糖的氧化能力均降低, 产生过多的酮体(乙酰乙酸、 羟丁酸 丙酮), 羟丁酸、 ),并发酮血症 ,产生过多的酮体(乙酰乙酸、β-羟丁酸、丙酮),并发酮血症 和酮尿症。 和酮尿症。
G-1-P
UTP (a) ADP ATP
G-6-P
Pi
G
糖原的合成与分解代谢
第六节 糖的异生
* 概念
糖异生(gluconeogenesis)是指从非糖化 糖异生(gluconeogenesis)是指从非糖化 (gluconeogenesis) 合物转变为葡萄糖或糖原的过程。 合物转变为葡萄糖或糖原的过程。
合成具有α 糖苷键的有分支的糖原, 3. 合成具有α-1,6糖苷键的有分支的糖原,反应由 分支酶催化 催化: 分支酶催化:
糖原 n+1
UDP (c)
Pi
(d)
UDPG UDPG
PPi (b)
(a) 磷酸葡萄糖变 位酶 UDPG焦磷酸 (b) UDPG焦磷酸 化酶 (c) 糖原合成酶 分支酶) (分支酶) (d) 磷酸化酶 脱支酶) (脱支酶)
2、UDPG在糖原合成酶催化下合成新糖原 UDPG在糖原合成酶催化下合成新糖原 新的葡萄糖残基加在糖原引物的非还原末端的葡萄糖 残基的第四碳的羟基上, 糖苷键。 残基的第四碳的羟基上,形成α-1,4糖苷键。UDP被 , 糖苷键 被
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
光反应:捕获光能并转变成化学能,提供ATP和NADPH 。 (位于真核生物叶绿体类囊体膜,原核生物的内膜系统)
暗反应:还原或固定CO2并合成细胞物质(位于真核生物 的叶绿体基质中,原核生物的羧酶体)
1)循环光合磷酸化
循环光合磷酸化是指电子从菌绿素分 子逐出后循环一周仍返回菌绿素。其 反应中心的吸收光波为“P870”。菌 绿素受日光照射后成为激发态,氧化 还原电位由+0.5变为-0.7,由它逐 出的电子通过类似呼吸链的传递,经 Bph(脱镁菌绿素)、辅酶Q、Cyt b/c1、FeS、Cyt c2的循环传递,最
作为氢受体的氨基酸主要有甘氨酸、脯氨酸、羟脯氨 酸、鸟氨酸、精氨酸和色氨酸等。
(2)化能自养型微生物的生物氧化和产能
化能自养型微生物中,其ATP是通过氧化还原态无机物 产生的,其NAD(P)H2是通过消耗ATP将无机氢(H++e)逆 呼吸链传递产生的。
化能自养型微生物均为细菌,且绝大多数为好氧菌
能量代谢特点: 脱氢酶或氧化还原酶催化无机底物脱氢或脱电子;
嗜盐菌 细胞膜
红色部分(红膜) 主要含细胞色素和黄素蛋白等用于氧化磷酸化的呼吸链载体
紫色部分(紫膜) 在膜上呈斑片状(直径约0.5 mm)独立分布,其总面积约占 细胞膜的一半,主要由细菌视紫红质组成。
实验发现,在波长为550-600 nm的光照下,嗜盐菌ATP的合成速率 最高,而这一波长范围恰好与细菌视紫红质的吸收光谱相一致。
氧化为NO3-获得能量,用同位素18O分析实验证明,在 NO2-氧化为NO3-过程中,氧来自水分子而非空气。
NO2- + H2O ——→ H2O NO2- ——→ NO3- + 2H+ + 2e
无机底物氧化的自由能
反应
氧化亚铁硫杆菌: 2Fe2+ + 2H+ + ½O2 → 2Fe3+ + H2O
第六章 微生物的代谢
二、糖的合成代谢 1. 糖合成的能量来源
包括:化能异养型、化能自养和光能营养微生物的生 物氧化和产能
(1)化能异养型微生物的生物氧化和产能
糖的分解代谢所产生的能量都可以用于糖的生物合 成,本节第一部分已经介绍过。 此外,某些化能异养微生 物(如Closterdium sporogenes 生孢梭菌)能利用一些氨基 酸同时当作碳源、氮源和能源。
终重新由菌绿素 接受,其间建立 质子动势并产生 1分子ATP。
当外源氢供体(H2S、H2、Fe2+等)提 供电子,沿呼吸链链逆向传递,由 NAD(P)+接受电子,产生可用于还原 CO2的NAD(P)H+H+。
循环光合磷酸化特点:
①电子传递途径属循环式的; ②产ATP和NAD(P)H+H+分别进行; ③NAD(P)H+H+中的[H]是来自H2S等无机氢供体; ④无O2产生。
硝化细菌: NO2- + ½O2 → NO3-
氢细菌:
H2 + ½O2 → H2O
亚硝化细菌: NH4+ + 1½O2 → NO2- + H2O + 2H+
硫细菌:
S0 + ½O2 + H2O → H2SO4 S2O32- + 2O2 + H2O → 2SO42- + 2H+
⊿G0’ kcal/mole
氨基酸发酵产能——Stickland反应 Stickland反应产能机制是通过部分氨基酸(如丙氨酸)
的氧化与另一些氨基酸(如甘氨酸)的还原相偶联的发酵过 程而产生能量的。一种氨基酸作氢供体和以另一种氨基酸作 氢受体。产能效率很低,每分子氨基酸仅产1个ATP。
作为氢供体的氨基酸主要有丙氨酸、亮氨酸、异亮氨 酸、缬氨酸、苯丙氨酸、丝氨酸、组氨酸和色氨酸等。
非循环光合磷酸化特点:
①电子的传递途径属非循环式的; ②在有氧条件下进行; ③有两个光合系统,其中色素系统I(PS I)含叶绿素a,
可以吸收利用红光,反应中心的吸收光波为“P700”, 色素系统II(PSII)含叶绿素b,可以吸收利用蓝光, 反应中心的吸收光波为“P680” ; ④反应中同时有ATP(产自PSII)、NAD(P)H+H+ (产自 PS I)和O2 产生; ⑤NAD(P)H+H+中的[H]是来自H2O分子光解后的H+和电 子。
-11.2 -17.4 -56.6 -65.0 -118.5 -223.7
区别:
无氧呼吸
无机物氧化
(化能异养型)
(化能自养型)
同
都涉及无机物
电子供体
(底物)
有机物
还原型无机物
电子受体 氧化型无机物(主)
O2
异 过程
举例
电子传递链
有机物 脱氢
无机物
反硝化细菌
无机物
电子传递链
脱氢
O
或氧化
硝化细菌
(3)光能营养微生物的生物氧化和产能
无机底物脱下的氢直接进入呼吸链,通过氧化磷酸化产能; (少数菌在无机硫化物存在时,能部分通过底物磷酸化产能) 氢或电子可从多处进入呼吸链,所以,呼吸链多样;
由于从中间进入呼吸链,因此产能效率低,菌体生长缓慢,细 胞产率低。
亚硝酸的氧化 Nitorbacter(硝化杆菌属)以亚硝酸作为能源,将NO2-
这种不产氧的循环式的光合磷酸化,只存在于原核生 物(光合细菌)中。
2)非循环光合磷酸化
各种绿色植物、藻类和蓝细菌的光合作用属非循环光合磷酸化
该光合磷酸化过程中 ,有氧气放出,其来 源是H2O的光解,整 个过程中,电子须经 过PSII和PS I两个系 统接力传递,传递体 包括PSII系统中的 Phea(褐藻素)、Q (醌)、Cyt bf、Pc (质体蓝素),在 Cyt bf和Pc间产生1 个ATP;还包括PS I 系统中的FeS(非血 红素铁硫蛋白)、Fd (铁氧还蛋白),最 终由NADP+接受电子 ,产生可用于还原 CO2的NADPH+H+。
3)嗜盐菌紫膜的光合作用
一种只有嗜盐菌才有的,无叶绿素或细菌叶绿素参与的 独特的光合作用。
嗜盐菌是一类必须在高盐(3.5~5.0mol/L NaCl)环境中才 能生长的古细菌。
嗜盐菌可通过两条途径获取能量: 有氧条件下的氧化磷酸化途径; 无氧条件下的紫膜光合磷酸化途径。
嗜盐菌在无氧条件下,利用光能所造成的紫膜蛋白上视黄醛 辅基构像的变化,将质子不断驱至膜外,从而在膜的两侧建 立一个质子动势,推动ATP酶合成ATP。