糖酵解柠檬酸循环PPT课件
合集下载
课件:柠檬酸循环

• C-C 间脱氢形成双键时,氧化所释放出来的 能量较低,只能使FAD还原不能使NAD+还原.
• 丙二酸是竞争性抑制剂,能阻断柠檬酸循 环。
COOH CHOH CH2 COOH malate
苹果酸
H2O fumarase COOH CH CH
延CO胡O索H 酸
fumarate
7: 延胡索酸酶
水合,形成 L-苹果酸
CoASH
4C
oxaloacetate
6C
citrate
cis-aconitate
4C
malate
6C
isocitrate
NAD+
NADH
CO2
H2O
4C
fumarate FADH
FAD
5C
ketoglutarate (oxo-glutarate)
CoASH
CoASH
NAD+
4C
succinate
GTP
• 异柠檬酸脱氢酶
– NADH,ATP 抑制, – NAD+ ,ADP, Ca2+激活
• a-酮戊二酸脱氢酶
– NADH 和琥珀酰CoA 抑制 – AMP Ca2+激活
主要调节因子 ADP/ATP , NAD+/NADH
Ca2+
TCA 循环的双重作用
• 有化合物进入循环氧化分解也有一些离开 循环做其他的用途
6C
isocitrate
NAD+
NADH
CO2
H2O
4C
fumarate FADH
FAD
5C
ketoglutarate (oxo-glutarate)
• 丙二酸是竞争性抑制剂,能阻断柠檬酸循 环。
COOH CHOH CH2 COOH malate
苹果酸
H2O fumarase COOH CH CH
延CO胡O索H 酸
fumarate
7: 延胡索酸酶
水合,形成 L-苹果酸
CoASH
4C
oxaloacetate
6C
citrate
cis-aconitate
4C
malate
6C
isocitrate
NAD+
NADH
CO2
H2O
4C
fumarate FADH
FAD
5C
ketoglutarate (oxo-glutarate)
CoASH
CoASH
NAD+
4C
succinate
GTP
• 异柠檬酸脱氢酶
– NADH,ATP 抑制, – NAD+ ,ADP, Ca2+激活
• a-酮戊二酸脱氢酶
– NADH 和琥珀酰CoA 抑制 – AMP Ca2+激活
主要调节因子 ADP/ATP , NAD+/NADH
Ca2+
TCA 循环的双重作用
• 有化合物进入循环氧化分解也有一些离开 循环做其他的用途
6C
isocitrate
NAD+
NADH
CO2
H2O
4C
fumarate FADH
FAD
5C
ketoglutarate (oxo-glutarate)
生物化学 第九章 柠檬酸循环(共62张PPT)

2 FADH2 3
2 NADH 5
• Total
25 A底T物P磷酸化
丙酮酸只有4个氢,
但彻底氧化所放出的氢?
加水加氢
糖酵解+三羧酸循环的效率
糖酵解
1G → 2ATP+2NADH+2H++2丙酮酸
→ 7ATP
三羧酸循环 2丙酮酸 → 25ATP
———————————————————————
32ATP
7、延胡索酸水合生成 L-苹果酸 Hydration of Fumarate to Produce Malate
8、 L-苹果酸脱氢形成草酰乙酸
Oxidation of Malate to Oxaloacetate
2F丙1(速21(FEEN氟与 糖或N(22不G四糖E5→变(+、 、 琥 、αoo313AAl异丙C酮率2乙E的生能、酵构2u-rr:--1)DD酮mm) 丙 二 )产产珀琥o:2柠酮)酸 受 酰 有 物 使 柠 解 抑为Pn二的aa戊物物酮酸氢珀v7为檬酸是是Att脱细辅氧细丙檬+制是辅eii氢硫Aoo二M控控酸硫酰三r辅酸糖糖Tnn氢胞酶氧胞酮酸剂s好 酶硫辛PP酸制制脱辛i-羧ooo酶类类C酶能化和酸循:A、氧,辛酸n::氢酸酸ffo(、、:复量(组脱环AP生需oIIA酸上ss/脱NN循αTfD胞脂脂转底oo合状织氢的a物M酮P脱的SAA2Hcc羧氢e环质类类化、物2ugDD体态中酶化ii2体r磷戊tt氢-琥2oα2crr酶酶S的HH也、、aa为aN,c的、的复学b+-内酸延二酶H珀tt酮ci(i效A(、、een有c蛋蛋琥形e结调生糖合计最酶胡酸y酰D戊vvto组率线乙乙l)y白白珀成合ii-控物发体量Hx主、素aa-lC二-分iC粒酰酰d质质C酸氟:合酵活cco要C酸oa酸iio)A体--ss彻彻A柠atCC成为性A的i--o2tAA)ooo底底檬n需氧降+产AAcc)S、oo分分酸求所低能unn胰c解解ii,调抑的途ttcaa岛i的的不tt节制n径ee素a共共能,!t2这e2同同往2N种N2A途途N下AF现DADA径径反HD象HD!!应HH巴2,斯55称5德3致(L死. 性合成
第十一章柠檬酸循环。ppt

(二)异柠檬酸的形成 柠檬酸在顺乌头酸酶(aconitase)的催化下, 柠檬酸在顺乌头酸酶(aconitase)的催化下,异 (aconitase)的催化下 构化转变成异柠檬酸(isocitrate) 顺乌头酸(cis (isocitrate)。 (cis构化转变成异柠檬酸(isocitrate)。顺乌头酸(cisaconitate)是这一转变反应的中间物 是这一转变反应的中间物。 aconitate)是这一转变反应的中间物。该步反应是可 逆的, Go'= kJ/mol,反应有利于柠檬酸。 逆的,△Go'=﹢6.3 kJ/mol,反应有利于柠檬酸。 因此,在平衡时,异柠檬酸大约只占10% 10%。 因此,在平衡时,异柠檬酸大约只占10%。
第二节
柠 檬 酸 循 环
柠檬酸循环( cycle)是乙酰基二碳单位进 柠檬酸循环(citric acid cycle)是乙酰基二碳单位进 一步氧化降解产生CO2和还原型辅酶的代谢途径。 CO2和还原型辅酶的代谢途径 一步氧化降解产生CO2和还原型辅酶的代谢途径。由于该反应顺 序是乙酰CoA与草酰乙酸缩合成柠檬酸开始的 乙酰CoA 缩合成柠檬酸开始的, 序是乙酰CoA与草酰乙酸缩合成柠檬酸开始的,且草酰乙酸经多 步反应之后重新生成,构成了一个循环反应途径,因此, 步反应之后重新生成,构成了一个循环反应途径,因此,该循环 反应称为柠檬酸循环。 反应称为柠约4600kD, 外形 大肠杆菌丙酮酸脱氢酶复合物分子量约4600kD, 4600 呈球形,其直径约为30nm。该复合物的核心由24 E2构成 30nm 24个 构成, 呈球形,其直径约为30nm。该复合物的核心由24个E2构成, 24个E1和12个E3环绕E2排列 环绕E2排列。 24个E1和12个E3环绕E2排列。真核生物丙酮酸脱氢酶复合 物比大肠杆菌的酶更为复杂。 物比大肠杆菌的酶更为复杂。
生物化学课件-柠檬酸循环

抑制該酶複合物的活性 反應物(NAD+、FAD、GDP、ADP、丙酮酸)啟動該
酶複合物的活性 Ca2+、胰島素啟動
Chapter23 檸檬酸迴圈
三、TCA迴圈
(二)檸檬酸迴圈概貌(98頁)
是乙醯CoA與草醯 乙酸結合進入迴圈經 一系列反應再回到草 醯乙酸的過程。在這 個過程中乙醯CoA被 氧化成H2O和CO2並 產生大量的能。其反 應途徑可表示如圖
Chapter23 檸檬酸迴圈
Tricarboxylic acid cycle
Chapter23 檸檬酸迴圈
三羧酸迴圈(tricarboxylic acid cycle, TCA 迴圈or Krebs迴圈)
一、TCA迴圈的發現
●德國科學家Hans Krebs 1937年提出,1953年獲得諾 貝爾獎,並被稱為ATP迴圈 (檸檬酸迴圈)之父。
NAD+
CoASH
+ NADH + H
(1)
CO 2
草醯乙酸
CH3CO~SCoA 乙醯 CoA
(4)(7)(8)(10)
• 產能步驟 • 2NAD(P)H • 1FADH2
OC COOH
(10) C COOH
H
H2
L-蘋果酸HOC COOH NADH+H+
(2)
C COOH
H2O
H 2O (9) H2
NADH CO 2
+
+ H
NAD+
CH2COOH COCOOH
CH 2
(5) 草醯琥珀酸
COCOOH CO2
(6)
CoASH α-酮戊二酸
(7) 琥珀醯CoA合成酶 (8) 琥珀酸脫氫酶 (9) 延胡索酸酶 (10)L-蘋果酸脫氫酶
酶複合物的活性 Ca2+、胰島素啟動
Chapter23 檸檬酸迴圈
三、TCA迴圈
(二)檸檬酸迴圈概貌(98頁)
是乙醯CoA與草醯 乙酸結合進入迴圈經 一系列反應再回到草 醯乙酸的過程。在這 個過程中乙醯CoA被 氧化成H2O和CO2並 產生大量的能。其反 應途徑可表示如圖
Chapter23 檸檬酸迴圈
Tricarboxylic acid cycle
Chapter23 檸檬酸迴圈
三羧酸迴圈(tricarboxylic acid cycle, TCA 迴圈or Krebs迴圈)
一、TCA迴圈的發現
●德國科學家Hans Krebs 1937年提出,1953年獲得諾 貝爾獎,並被稱為ATP迴圈 (檸檬酸迴圈)之父。
NAD+
CoASH
+ NADH + H
(1)
CO 2
草醯乙酸
CH3CO~SCoA 乙醯 CoA
(4)(7)(8)(10)
• 產能步驟 • 2NAD(P)H • 1FADH2
OC COOH
(10) C COOH
H
H2
L-蘋果酸HOC COOH NADH+H+
(2)
C COOH
H2O
H 2O (9) H2
NADH CO 2
+
+ H
NAD+
CH2COOH COCOOH
CH 2
(5) 草醯琥珀酸
COCOOH CO2
(6)
CoASH α-酮戊二酸
(7) 琥珀醯CoA合成酶 (8) 琥珀酸脫氫酶 (9) 延胡索酸酶 (10)L-蘋果酸脫氫酶
三羧酸循环--me55页PPT

丙酮酸脱氢酶复合物三种酶的功能:
(1)丙酮酸脱氢酶(也称丙酮酸脱羧酶): 辅基TPP,E1。 功用: Py 脱羧 2c单位
(2)二氢硫辛酸转乙酰基酶:辅基硫辛酰胺(或称硫辛酸),E2。 功用:氧化2C单位,并将2C单位先转到硫辛酰胺上, 再转到CoA上。
(3)二氢硫辛酸脱氢酶:辅基FAD,E3。
功用:Red型硫辛酰胺→OX型硫辛酰胺
③ 催化丙酮酸转变为乙酰CoA的反应步骤
E1
(1)Py+TPP
羟乙基-TPP+CO2
OX
(2)羟乙基-TPP E2
乙酰基-硫辛酰胺
(3)乙酰基-硫辛酰胺+CoA E2
乙酰CoA+硫辛酰胺
(4)Red型硫辛酰胺
E3
OX型硫辛酰胺
(5) F SA H 2 D N SH A D F S A S D NA H D H
三羧酸循环酶主要有8个,你记住了吗?
依次为柠檬酸合成酶、顺乌头酸酶、异柠檬酸 脱氢酶、α-酮戊二酸脱氢酶、琥珀酰CoA合成 酶、琥珀酸脱氢酶、延胡索酸酶、苹果酸脱氢 酶。难记不?不难记。你根本不用记得那样准 确。
听歌:一只黑猫两只虎,两个柠檬一只狐,一 个苹果一桶醋(α酮戊二酸脱氢酶)有人说黑 猫是什么,那不就是乌头嘛。
9. 三羧酸循环的调节
①三羧酸循环中的三个调控酶
(1)OAA+乙酰CoA→Cit,Cit合酶,限速酶,受琥 珀酰CoA、NADH、ATP和脂酰CoA的抑制。
(2)异Cit→α-KGA,异Cit脱氢酶,ADP是别构激活 剂,ATP和NADH是抑制剂。
(3)α-KGA→琥珀酰CoA,α-KGA脱氢酶被反应 产物琥珀酰CoA和NADH抑制,也被高的能荷 抑制。
糖酵解和柠檬酸循环演示文稿

控制糖酵解的出口
果糖1,6-二磷酸:别构激活
起活化作用,与磷酸果糖激酶协调,加速酵解。
前馈激活(feed-forward activation)
ATP:别构抑制
丙氨酸:别构抑制
丙酮酸转氨生成,表示生物合成过剩。
当前第43页\共有87页\编于星期四\21点
2 柠檬酸循环
2.1 柠檬酸循环途径
2.1.1 基本概念
三个不可逆反应: 己糖激酶 磷酸果糖激酶 丙酮酸激酶
当前第37页\共有87页\编于星期四\21点
⑴ 己糖激酶的调控
G 己糖激酶 G-6-P
-
己糖激酶同功酶中除葡萄糖激酶外,都受到葡萄糖-6-磷酸的抑制。
当前第38页\共有87页\编于星期四\21点
⑵ 磷酸果糖激酶的调控
磷酸果糖激酶(PFK-1 ,Phosphofructokinase-1 )糖酵 解中最关键的限速酶。
糖酵解和柠檬酸循环演示文稿
当前第1页\共有87页\编于星期四\21点
(优选)糖酵解和柠檬酸循环
当前第2页\共有87页\编于星期四\21点
1 糖酵解
1.1糖酵解(glycolysis) : 在无氧条件下,葡萄糖进行分解,形成2分子
丙酮酸并提供能量(ATP)的过程。
它是各种生物细胞中葡萄糖分解产生能量的 共同代谢途径。
1
2 3
An aldehyde
4
5
6
当前第17页\共有87页\编于星期四\21点
⑸ 磷酸二羟丙酮异构成3-磷酸甘油醛
1
1
2
2
3
3
当前第18页\共有87页\编于星期四\21点
准备阶段共消耗2分子ATP,产生2分子3-磷酸甘油醛.
果糖1,6-二磷酸:别构激活
起活化作用,与磷酸果糖激酶协调,加速酵解。
前馈激活(feed-forward activation)
ATP:别构抑制
丙氨酸:别构抑制
丙酮酸转氨生成,表示生物合成过剩。
当前第43页\共有87页\编于星期四\21点
2 柠檬酸循环
2.1 柠檬酸循环途径
2.1.1 基本概念
三个不可逆反应: 己糖激酶 磷酸果糖激酶 丙酮酸激酶
当前第37页\共有87页\编于星期四\21点
⑴ 己糖激酶的调控
G 己糖激酶 G-6-P
-
己糖激酶同功酶中除葡萄糖激酶外,都受到葡萄糖-6-磷酸的抑制。
当前第38页\共有87页\编于星期四\21点
⑵ 磷酸果糖激酶的调控
磷酸果糖激酶(PFK-1 ,Phosphofructokinase-1 )糖酵 解中最关键的限速酶。
糖酵解和柠檬酸循环演示文稿
当前第1页\共有87页\编于星期四\21点
(优选)糖酵解和柠檬酸循环
当前第2页\共有87页\编于星期四\21点
1 糖酵解
1.1糖酵解(glycolysis) : 在无氧条件下,葡萄糖进行分解,形成2分子
丙酮酸并提供能量(ATP)的过程。
它是各种生物细胞中葡萄糖分解产生能量的 共同代谢途径。
1
2 3
An aldehyde
4
5
6
当前第17页\共有87页\编于星期四\21点
⑸ 磷酸二羟丙酮异构成3-磷酸甘油醛
1
1
2
2
3
3
当前第18页\共有87页\编于星期四\21点
准备阶段共消耗2分子ATP,产生2分子3-磷酸甘油醛.
糖酵解和柠檬酸循环

05
糖酵解与柠檬酸循环异常 与疾病关系
糖酵解异常导致疾病
糖尿病
糖酵解是体内葡萄糖分解代谢的主要途径,糖尿病患 者体内糖酵解过程受损,导致血糖升高。
乳酸酸中毒
糖酵解过程中产生的乳酸不能及时被清除,导致乳酸 堆积,引发乳酸酸中毒。
恶性肿瘤
肿瘤细胞通过异常糖酵解获取能量,促进肿瘤生长和 扩散。
柠檬酸循环异常导致疾病
生态学和环境科学领域
糖酵解和柠檬酸循环在生态系统的物质循环和能量流动中发挥着重要作用。研究这些代谢途径有助于深 入了解生物与环境之间的相互作用,为生态保护和环境治理提供科学依据。
THANKS
感谢观看
柠檬酸循环不仅是能量代谢的中心环 节,还参与多种物质的代谢和转化过 程,如脂肪、蛋白质等,对维持细胞 正常生理功能同样具有重要作用。
参与物质代谢和转化过程
糖酵解
糖酵解过程中产生的中间产物可以进一 步转化为其他物质,如氨基酸、脂肪酸 等,参与细胞的物质代谢和转化过程。
VS
柠檬酸循环
柠檬酸循环中的中间产物也可以进一步转 化为其他物质,如酮体、胆固醇等,参与 细胞的物质代谢和转化过程。同时,柠檬 酸循环还与糖异生、脂肪代谢等过程密切 相关,共同维持细胞内物质代谢的平衡。
丙酮酸进入柠檬酸循环
糖酵解产生的丙酮酸,在有氧条件下,可进入线粒体,经过氧化脱羧生成乙酰CoA,进而进入柠檬酸循环。
3-磷酸甘油醛脱氢产生NADH
糖酵解过程中,3-磷酸甘油醛脱氢产生的NADH+H+经线粒体穿梭作用进入柠檬酸循环,参与氧化磷酸化过程。
柠檬酸循环对糖酵解的调节作用
ATP/ADP比值调节
医学领域
糖酵解和柠檬酸循环的异常与多种疾病的发生发展密切相关,如糖尿病、肥胖症、心血管疾病等。深入研究这些代谢 途径有望为疾病的预防、诊断和治疗提供新的思路和方法。
柠檬酸循环与糖异生PPT课件
O
F CH COO-
CH2C OF
HO C COOH2C COO-
氟乙酸
氟柠檬酸
异柠檬酸脱氢酶的催化反应
来自草酰乙酸
异柠檬酸
异柠檬酸脱氢酶 α-酮戊二酸
第一次脱羧
Mn2+:使新形成的羰基极化, 促使了邻近碳碳键的断裂。
β脱羧
α
β
β羟酸氧化 为β酮酸
异柠檬酸
草酰琥珀酸
α-酮戊二酸
异柠檬酸脱氢酶的调节
别构调节酶活:激活——CoA、AMP(低能荷)、 NAD+、Ca2+ (竞争性)抑制——乙酰-CoA、 ATP(高能荷) NADH+H+、
丙酮酸脱氢酶复合物
共价修饰调节酶活(E1):E2分子上结合着激酶和磷酸酶。磷酸 化E1使之失活, 去磷酸化使之活化。
Pi
Ca2+ + E1磷酸酶
H2O
丙酮酸脱氢酶E1 (脱磷酸,活性) ATP
四、血糖及其调节
• 正常人空腹时血糖浓度是相当恒定的 ,维持在 4.5-5.5mmol/L (70-110mg/100ml)之间
• (一) 血糖的来源与去路
食物中的糖 ↘ ↗氧化分解供能 肝糖原的分解 → 血糖 →合成糖原 肝中糖异生作用↗ ↘转化为非糖物质
↘随尿排出
(二)激素对血糖的调节
• 血糖升高作用:胰高血糖素,肾上腺素,糖皮质激素
• 3-磷酸甘油醛 (脱氢)
↓
NADH + H+ 胞浆(不能自由进入线粒体内膜)
↓
↓
↓
磷酸甘油穿梭
苹果酸穿梭
(苹果酸-天冬氨酸穿梭)
↓
• FADH2 (骨骼肌,脑组织)
细胞呼吸——柠檬酸循环课件

TCA背景知识
2、细胞呼吸(cell respiration) 要经历三个阶段:糖酵解阶 段、柠檬酸循环阶段、氧化 磷酸化阶段。 3、糖酵解的产物丙酮酸进入 TCA之前有一准备过程,即 形成乙酰CoA。
TCA准备阶段
丙酮酸在丙酮酸脱氢酶系催化下氧化脱羧形 成乙酰辅酶A。
IRREVERSIBLE
• In 1937, Krebs found that citrate could be formed in muscle suspensions if oxaloacetate and either pyruvate or acetate were added. Now, he get a cycle:
• In 1932, Krebs was studying the rates of oxidation of small organic acids by kidney and liver tissue. Only a few of substances were active in these experiments---notably succinate, fumarate, acetate, malate, and citrate.
Brief history of TCA
• The first major investigation into the intermediary metabolism of oxidation was that of Thunberg, who examined systematically the oxidizability of organic substances in isolated animal tissues. Between 1906 and 1920 he tested the oxidation of over 60 organic substances, chiefly in muscle tissue. He discovered the rapid oxidation of the salts of a number of acids, such as lactate(乳酸盐), succinate(琥珀酸盐), fumarate(延胡索酸盐), malate(苹果酸盐), citrate(柠檬酸盐), and glutamate(谷氨酸盐).
糖酵解ppt课件

糖酵解途径涉及10个酶催化反应,途径中的酶都位于细 胞质中,一分子葡萄糖通过该途径被转换成两分子丙酮酸。 糖酵解的10个反应可以分为己糖和丙糖两个阶段。己糖阶段 是从葡萄糖到果糖-1,6-二磷酸,丙糖阶段是从果糖-1,6-二磷 酸裂解转换为两个三碳的磷酸丙糖开始到糖酵解的产物丙酮 酸为止。
ห้องสมุดไป่ตู้ 葡萄糖酵解过程
磷酸化的葡萄糖被限制在细胞内,因为磷酸化的糖含有带负电荷的磷 酰基,可防止糖分子再次通过质膜。这是细胞的一种保糖机制。在糖代谢 的整个过程中,直至净合成能量之前,中间代谢物都是磷酸化的。
己糖激酶以六碳糖为底物,专一性不强。除了葡萄糖可作为己糖激酶 的底物外,它也可催化甘露糖、果糖等己糖的磷酸化。己糖激酶对D-葡萄 糖的Km=0.1mmol/L,而肝葡萄糖激酶的Km=10 mmol/L,平时细胞内葡萄 糖浓度为5 mmol/L,此时己糖激酶的酶促反应已达最大速度,而葡萄糖 激酶并不活跃。只有在进食后,肝细胞内葡萄糖浓度高时葡萄糖激酶才起 作用,所以葡萄糖激酶是个诱导酶。
-NAD+ CH2O-P
-SH
Pi NADH+ +H+
见教材下册P66,糖酵解过程中,中间产物磷酸化的 重要意义是:
(1) (2) (3)
糖酵解包括10步酶催化反应
1、己糖激酶催化葡萄糖磷酸化形成葡萄糖-6-磷酸,消 耗一分子ATP.
糖酵解的第一步反应是葡萄糖的C-6被磷酸化形成葡萄糖-6-磷酸,这 一磷酰基团转移反应是由己糖激酶催化的,消耗一分子ATP,该反应是不 可逆反应。
2、葡萄糖-6-磷酸异构酶催化葡萄糖-6-磷酸转化为果糖-6-磷酸
葡萄糖-6-磷酸异构酶催化葡萄糖-6-磷酸转化为果糖-6-磷酸,这是一个醛 糖-酮糖同分异构化反应,反应是可逆的。
ห้องสมุดไป่ตู้ 葡萄糖酵解过程
磷酸化的葡萄糖被限制在细胞内,因为磷酸化的糖含有带负电荷的磷 酰基,可防止糖分子再次通过质膜。这是细胞的一种保糖机制。在糖代谢 的整个过程中,直至净合成能量之前,中间代谢物都是磷酸化的。
己糖激酶以六碳糖为底物,专一性不强。除了葡萄糖可作为己糖激酶 的底物外,它也可催化甘露糖、果糖等己糖的磷酸化。己糖激酶对D-葡萄 糖的Km=0.1mmol/L,而肝葡萄糖激酶的Km=10 mmol/L,平时细胞内葡萄 糖浓度为5 mmol/L,此时己糖激酶的酶促反应已达最大速度,而葡萄糖 激酶并不活跃。只有在进食后,肝细胞内葡萄糖浓度高时葡萄糖激酶才起 作用,所以葡萄糖激酶是个诱导酶。
-NAD+ CH2O-P
-SH
Pi NADH+ +H+
见教材下册P66,糖酵解过程中,中间产物磷酸化的 重要意义是:
(1) (2) (3)
糖酵解包括10步酶催化反应
1、己糖激酶催化葡萄糖磷酸化形成葡萄糖-6-磷酸,消 耗一分子ATP.
糖酵解的第一步反应是葡萄糖的C-6被磷酸化形成葡萄糖-6-磷酸,这 一磷酰基团转移反应是由己糖激酶催化的,消耗一分子ATP,该反应是不 可逆反应。
2、葡萄糖-6-磷酸异构酶催化葡萄糖-6-磷酸转化为果糖-6-磷酸
葡萄糖-6-磷酸异构酶催化葡萄糖-6-磷酸转化为果糖-6-磷酸,这是一个醛 糖-酮糖同分异构化反应,反应是可逆的。
柠檬酸循环ppt

第二十章 柠檬酸循环
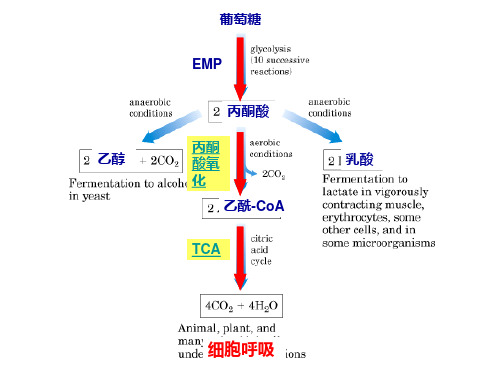
大多数动物、植物和微生物,葡萄糖通过糖酵解产 大多数动物、植物和微生物, 生的丙酮酸,在有氧条件下, 生的丙酮酸,在有氧条件下,氧化脱羧形成乙酰辅酶 乙酰辅酶A经过一系列氧化、脱羧,最终生成H A。乙酰辅酶A经过一系列氧化、脱羧,最终生成H2O 并释放出大量能量的过程称为三羧酸循环 和CO2,并释放出大量能量的过程称为三羧酸循环 cycle)又柠檬酸循环 又柠檬酸循环, (tricarboxylic acid cycle)又柠檬酸循环,简写 为TCA循环,因为它是由H.A.Krebs正式提出,所以又 TCA循环,因为它是由H.A.Krebs正式提出, 循环 H.A.Krebs正式提出 Krebs循环 循环。 称Krebs循环。
丙酮酸脱氢酶复合体催化反应 的图解
高能键
羟乙基TPP 羟乙基TPP
丙酮酸脱氢酶复合体的调控: 丙酮酸脱氢酶复合体的调控: 乙酰1、产物控制:NADH 、乙酰-CoA 产物控制: 2、丙酮酸脱氢酶组分的磷酸化(失活)和去 丙酮酸脱氢酶组分的磷酸化(失活) 磷酸化(激活) 磷酸化(激活) 由E2上结合的激酶和磷酸酶起作用
2、柠檬酸异构化生成异柠檬酸
柠檬酸
顺乌头酸酶
异柠檬酸
乌头酸酶
3、异柠檬酸氧化形成α酮戊二酸 异柠檬酸氧化形成α
异柠檬酸脱氢酶
氧化脱羧
NAD为辅酶, 线粒体) NAD为辅酶,需Mg2+(线粒体) 为辅酶 异柠檬酸脱氢酶(别构酶 异柠檬酸脱氢酶 别构酶) 别构酶 NADP为辅酶(细胞溶胶、线粒体) NADP为辅酶(细胞溶胶、线粒体) 为辅酶
丙酮酸的有氧氧化包括两个阶段: 丙酮酸的有氧氧化包括两个阶段: 阶段:丙酮酸的氧化脱羧( 第一 阶段:丙酮酸的氧化脱羧(丙酮酸 → 乙酰辅酶A 简写为乙酰CoA 乙酰辅酶A,简写为乙酰CoA) 第二阶段:柠檬酸循环(乙酰CoA 第二阶段:柠檬酸循环(乙酰CoA → H2O 释放出能量) 和CO2,释放出能量)
大多数动物、植物和微生物,葡萄糖通过糖酵解产 大多数动物、植物和微生物, 生的丙酮酸,在有氧条件下, 生的丙酮酸,在有氧条件下,氧化脱羧形成乙酰辅酶 乙酰辅酶A经过一系列氧化、脱羧,最终生成H A。乙酰辅酶A经过一系列氧化、脱羧,最终生成H2O 并释放出大量能量的过程称为三羧酸循环 和CO2,并释放出大量能量的过程称为三羧酸循环 cycle)又柠檬酸循环 又柠檬酸循环, (tricarboxylic acid cycle)又柠檬酸循环,简写 为TCA循环,因为它是由H.A.Krebs正式提出,所以又 TCA循环,因为它是由H.A.Krebs正式提出, 循环 H.A.Krebs正式提出 Krebs循环 循环。 称Krebs循环。
丙酮酸脱氢酶复合体催化反应 的图解
高能键
羟乙基TPP 羟乙基TPP
丙酮酸脱氢酶复合体的调控: 丙酮酸脱氢酶复合体的调控: 乙酰1、产物控制:NADH 、乙酰-CoA 产物控制: 2、丙酮酸脱氢酶组分的磷酸化(失活)和去 丙酮酸脱氢酶组分的磷酸化(失活) 磷酸化(激活) 磷酸化(激活) 由E2上结合的激酶和磷酸酶起作用
2、柠檬酸异构化生成异柠檬酸
柠檬酸
顺乌头酸酶
异柠檬酸
乌头酸酶
3、异柠檬酸氧化形成α酮戊二酸 异柠檬酸氧化形成α
异柠檬酸脱氢酶
氧化脱羧
NAD为辅酶, 线粒体) NAD为辅酶,需Mg2+(线粒体) 为辅酶 异柠檬酸脱氢酶(别构酶 异柠檬酸脱氢酶 别构酶) 别构酶 NADP为辅酶(细胞溶胶、线粒体) NADP为辅酶(细胞溶胶、线粒体) 为辅酶
丙酮酸的有氧氧化包括两个阶段: 丙酮酸的有氧氧化包括两个阶段: 阶段:丙酮酸的氧化脱羧( 第一 阶段:丙酮酸的氧化脱羧(丙酮酸 → 乙酰辅酶A 简写为乙酰CoA 乙酰辅酶A,简写为乙酰CoA) 第二阶段:柠檬酸循环(乙酰CoA 第二阶段:柠檬酸循环(乙酰CoA → H2O 释放出能量) 和CO2,释放出能量)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
1905 年 ArhurHarden 和 WilliamYoung 把 酵 母 汁 加 入 葡 萄 糖 中 , 发现发酵过程中无机磷酸盐逐渐消失,只有不断补充无机磷酸 盐才能使发酵速度不降低,因此推测发酵与无机磷将糖磷酸化 有关。他们还发现当将酵母汁透析或加热到50℃后,就会失去 发酵能力,当加热失活的酵母汁与透析失活的酵母汁混合后又 恢复了发酵能力。由此证明,发酵活性取决于两类物质,一类 是热不稳定的,不可透析的组称为酿酶,一类是热稳定,可透 析的组分称为辅酶,还有金属离子。
.
7
后五步反应为产生产生ATP的贮能阶段, 6.甘油醛氧化;7.底物水平磷酸化 8.变位反应;9.烯醇化 10.再次底物水平磷酸化
磷酸三碳糖变成丙酮酸,每分子的三碳糖 产生2分子的ATP。
.
8
1. 葡萄糖磷酸化形成6-磷酸葡萄糖
已糖激酶
已糖激酶(分子量52000)以六碳糖为底物,其专
一性不强,不仅可以作用于葡萄糖,还可以作用于
糖类代谢
第一节:糖的来源及生理作用
• 一.糖的来源
绿色植物和光合微生物的光合作用 和动物体内糖异生
叶绿体
.
1
二. 糖的作用
1. 糖是有机体重要的能源和碳源。糖分解产生 能量,可以供给有机体生命活动的需要,
2. 糖代谢的中间产物又可以转变成其他的含碳 化合物如氨基酸、脂肪酸、核苷等。
3. 糖的磷酸衍生物可以形成重要的生物活性物 质.如NAD、FAD、DNA、RNA、ATP等。
烯醇化酶
.
17
10. 磷酸烯醇式丙酮酸将磷酰基转移给 ADP形成ATP和丙酮酸
丙酮酸激酶
现已得到丙酮酸激酶,分子量是250000,是由66000的亚基
组成的四聚体。丙酮酸激酶是一个别构酶,酵解途径中的重
要调节酶。长链脂肪酸,乙酰COA,ATP和丙氨酸能抑制该
酶活性。1,6—二磷酸果糖活化此酶。
.
18
磷酸甘油醛脱氢酶
碘乙酸可强烈抑制磷酸甘油醛脱氢酶的活性,因为与酶的半胱氨酸残 基上的-SH反应。
砷酸盐(AsO43-)可以与H3PO4竞争同高能硫酯中间物结合,形成不 稳定的化合物1-砷-3磷酸甘油酸,它可以进一步分解产生3磷酸甘
油酸,但没有磷酸化作用。因此砷酸使这一步的氧化作用和磷酸化作
用解偶联。这是砷酸中毒的反应之一. 。
1940年,酵解的全过程才被全面了解。Gustar Embden和Otto
Meyerhof等人发现肌肉中也存在着与酵母发酵十分类似的不
需氧的分解葡萄糖并产生能量的过程,他们称此为酵解过程;
因此有时称酵解为Embden—Meyerhof途径( EMP )。
.
5
糖类代谢
二.糖酵解的概述
糖酵解途径的概念
糖酵解:葡萄糖在酶促反应下生成丙酮酸并 伴随着ATP生成的过程。
.
22
酵解过程ATP的合成
葡萄糖酵解的总反应式为:
葡萄糖+2Pi+2ADP+2NAD+ 2丙酮酸+2ATP+2NADH+2H++2H2O
无氧情况下酵解共产生2分子的ATP,2分子NADH将H交给2分子 丙酮酸生成2分子乳酸。在有氧情况下,2分子NADH经呼吸链氧 化成H2O可产生5分子ATP,因此一分子葡萄糖酵解共产生7分子 ATP。 在某些组织,如骨骼肌、脑组织NADH进入线粒体要通过甘油磷 酸穿羧系统。细胞质中磷酸二羟丙酮被催化成3-磷酸甘油进入线 粒体重新氧化成磷酸二羟丙酮,但在线粒体中的3-磷酸甘油脱氢 酶的辅基是FAD,为此只产生1.5分子ATP。在这些组织中一分子 葡萄糖酵解共产生5分子ATP。(改错,P87)
14
7. 1,3-二磷酸甘油酸将磷酰基转 给ADP生成磷酸甘油酸和ATP
磷酸甘油酸激酶
底物水平磷酸化:ATP(GTP)的形成直接与一个代 谢中间物上的磷酸基因转移相偶联的作用
.
15
8.3-磷酸甘油酸变位形成2-磷酸甘油酸
磷酸甘油酸变位酶
.
16
9. 2-磷酸甘油酸脱水形成磷酸烯醇式丙 酮酸(PEP)
乳酸脱氢酶
.
20
3.转化为乙醇(酵母菌或其它微生物中)
丙酮酸脱羧酶
丙酮酸+H+
乙醛+CO2
醇脱氢酶
乙醛+NADH+H+
乙醇+NAD+
.
21
酵解与发酵概念的区别
发酵(fermentation): 厌氧有机体(如酵母
或其他微生物)把酵解生成的NADH中的氢交给丙 酮酸脱羧生成的乙醛,使之形成乙醇。这个过程称 为酒精发酵。若将氢交给丙酮酸生成的乳酸,则是 乳酸发酵。
D-果糖和D-甘露糖。已糖激酶像其他激酶一样,
需为Mg2+或其他二价金属如Mn2+-磷酸果 糖
OH
磷酸葡萄糖异构酶
.
10
3. 6-磷酸果糖磷酸化形成1,6- 二磷酸果糖( F-1,6-2P)
磷酸果糖激酶
这一步反应是酵解中的关键反应。磷酸果糖激酶的活性决
定了酵解的速度。磷酸果糖激酶是分子量为3400的四聚体,
它是一个别构酶,ATP对此酶有抑制效应,在有柠檬酸、
脂肪酸时加强抑制效应。然而AMP、ADP或无机磷可消
除抑制,增加酶的活性 .
11
4.F-1,6-2P裂解成3-磷酸甘油醛 和磷酸二羟丙酮(DHAP)
F-1,6-2P醛缩酶
.
12
5.磷酸三碳糖的同分异构化
磷酸丙糖异构酶
96%
4%
.
13
6. 3-磷酸甘油醛氧化成1,3-二磷酸 甘油酸
.
19
四. 丙酮酸的去路
1.变为乙酰CoA,进入三羧酸循环(在有氧条件下)
O
丙 酮 酸 O脱 氢 酶 系
C H 3C C O O H +HS-+N C A D o + A C H 3CS C o+ AC O 2+ N A D H
丙 酮 酸 辅 酶 A
乙酰辅酶A
2.生成乳酸(在无氧或暂时缺氧条件下)
• 糖酵解是指葡萄糖在酶促反应下生成 丙酮酸并伴随着ATP生成的过程。酵 解过程相关的酶都在细胞质中。
• 它是动物、植物、微生物细胞中葡萄 糖分解产生能量的共同代谢途径。
.
6
三.糖酵解过程
前五步为准备阶段: 1.葡萄糖磷酸化;2.磷酸已糖异构化; 3.再次磷酸化;4.果糖一1,6-二磷酸裂解; 5.磷酸丙糖异构化 此阶段中,葡萄糖通过磷酸化分解成三碳糖,每 分解一个已糖分子消耗2分子的ATP。
4. 糖蛋白、糖脂与细胞的免疫反应,识别作用 有关。
.
2
糖类代谢
二.糖的中间代谢
• 糖酵解途径 ( EMP) • 柠檬酸循环 (TCA) • 磷酸戊糖途径 (HMP) • 糖异生作用 • 植物体内乙醛酸循环 • 糖原的合成与分解
.
3
第二十二章:糖 酵 解
.
4
一. 糖酵解的研究历史
1897年,Hans Buchner和Eduard Buchner兄弟发现,酵母汁 可以把蔗糖变成酒精,证明了发酵可以在细胞以外进行