姊妹染色单体区分染色法
姐妹染色单体交换试验遗传学终点

姐妹染色单体交换试验是一种遗传学实验方法,用于研究染色体重组和遗传连锁。
通过染色体的染色单体交换,可以观察到不同等位基因之间的连锁程度,从而推断它们在染色体上的位置关系。
这一实验方法提供了对基因连锁和重组的直接证据,对于理解遗传变异和进化过程有着重要的意义。
1. 姐妹染色单体交换试验的基本原理姐妹染色单体交换试验是利用配子形成过程中的染色体分离和重新组合现象。
在该实验中,首先需要选择一对具有明显遗传特征的等位基因,在雌性生殖细胞的减数分裂过程中,这对基因所在的染色体会发生染色单体交换。
然后通过后续的配子形成和受精过程,可以得到一系列基因型不同的后代,通过统计分析后代的表型比例,可以确定基因之间的连锁程度以及它们在染色体上的位置关系。
2. 实验步骤及注意事项姐妹染色单体交换试验的具体步骤包括:(1) 选择实验材料:选择具有明显遗传特征的实验材料,保证基因型的稳定和清晰的表型特征。
(2) 实验前培养:对实验材料进行适当的培养和繁殖,以确保实验所用生物材料的数量和质量。
(3) 染色体减数分裂:观察生物材料的配子形成过程,确定染色单体交换发生的时间和位置。
(4) 配子形成和受精:观察配子形成和受精过程,收集各种基因型的后代生物,并进行表型统计和分析。
(5) 数据统计和分析:根据后代的表型比例,推断基因之间的连锁程度和位置关系。
在进行姐妹染色单体交换试验时,需要注意以下事项:(1) 实验材料的选择和保存:选择适宜的实验材料和种裙,确保实验所用生物材料的纯度和稳定性。
(2) 实验环境的控制:保持实验环境的稳定性和一致性,避免环境因素对实验结果的干扰。
(3) 数据的采集和统计:对后代生物的表型进行准确统计和记录,确保实验数据的可靠性和准确性。
(4) 结果的解释和推断:根据实验数据进行合理的结果解释和推断,提出对基因连锁和重组的相关假设和理论。
3. 实验在遗传学研究中的应用和意义姐妹染色单体交换试验作为一种重要的遗传学实验方法,在基因连锁和重组、遗传图谱绘制、遗传进化和种裙遗传结构等方面有着广泛的应用和重要的意义。
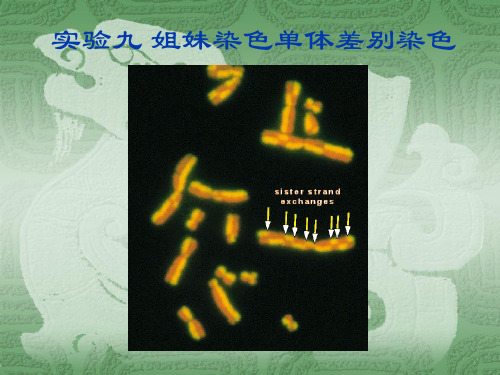
实验九 姐妹染色单体差别染色
SCE平均交换频率(次/细胞)= 交换总次数 / 细胞数
作业
完成此次实验报告(包含实验原理、实验药品、
实验步骤)
染色体识别要求: 1、 B组一对
2、 C组的8号、9号、10号任选取1对
3、 D组一对 4、 E组17号、18号任选取1对 5、 性染色体一对
实验九 姐妹染色单体差别染色
实验用品
1、器材:水浴锅、紫外灯、温度计、擦镜纸、 显微镜、滴管 2、试剂: 2×SSC缓冲液、Giemsa染液、 PH6.8 PBS、 3、材料:人外周血淋巴细胞染色体标本片
实验步骤
1、姐妹染色单体差别染色
A、将标本片放入培养皿中,加2×SSC数滴于 玻片上,盖一张擦镜纸,以湿润擦镜纸为宜,并在 培养皿中加入2×SSC使之浸在玻片底面。
B、把培养皿置于预温75℃水浴锅平板上,距 30W紫外灯管5cm,照射30分钟,其间滴加数次 2×SSC,勿使擦镜纸干燥。
C、用Giemsa染液染色10分钟,自来水冲洗、 晾干即为SCE标本。
2、SCE观察计数 镜下(油镜)观察中期分裂相时,注意区分各细胞周期 分裂相的染色特点:①染色体的两个单体均为深染的细胞 记为第一周期的细胞;②染色体的两个单体染色一深一浅 的细胞为第二周期细胞;③部分染色体的两个单体染色都 为浅色的细胞为第三周期的细胞。
ቤተ መጻሕፍቲ ባይዱ
第二周期细胞
选择染色体分散良好、长度适中、轮廓清晰、数目完整、 分化良好的的第二周期的分裂相进行计数。人体姐妹染色单体 交换(SCE)的计数方法:
①凡是在染色单体端部出现互换,记为1次交换; ②在染色单体中间出现互换,记为2次交换; ③如果在着丝粒处发生交换,需判明是否为扭转,不是扭转的 为着丝粒部位交换,记为1次交换。每一份标本至少需要计数 20~50个细胞的染色体,计算SCE平均交换频率。
高三复习-姐妹染色单体和染色单体的区别

姐妹染色单体和染色单体的区别
染色单体又称染色分体,它是染色体的一部分。
染色单体是在有丝分裂的细胞间期就形成的,这样被分成的二条染色体就叫染色单体。
姐妹染色单体是组成的染色单体的成分,在有丝分裂和减数第二次分裂的后期,每对姐妹染色单体都随着着丝点的分裂而彼此分开。
姐妹染色单体其原理是,当细胞接触到5-溴脱氧尿嘧啶核苷(Brdu)时,Brdu可作为核苷酸前体物,专一替代胸腺嘧啶掺入到新合成的DNA链中。
只要通过两个细胞复制周期,就可使姐妹染色单体中的一条单体的DNA双链中,有一股链是掺入有Brdu的,而另一条单体的DNA双链中,两股链全掺入有Brdu.这样的细胞经过分化染色处理,即可见到同一条染色体的两条姐妹染色单体有明显的差异,有一条单体为深色,另一条为浅色。
染色单体是复制时产生的染色体拷贝。
此名字通常用来形容处于随后的细胞分裂期它们分开之前的染色体。
从有丝分裂前期到中期(在有丝分裂后期,着丝点断裂,此时不存在染色单体),染色体沿其长轴发生纵裂。
这样被分成的二条染色体各称为染色单体。
开始成为一对的染色单体两者并不分开,逐渐它们具有独立的基质,并在其中各自形成二条染色丝。
而且染色单体往往出现互相关联的螺旋。
这些螺旋的圈数在中期以前逐渐减少,并且着丝粒也开始分裂。
染色体、染色单体、姐妹染色体、同源染色体、染色体组概念的区分

染色体、染色单体、姐妹染色体、同源染色体、染色体组概念的区分。
染色体:在生物的细胞核中,有一种易被碱性染料染上颜色的物质,叫做染色质。
染色体只是染色质的另外一种形态。
它们的组成成分是一样的,但是由于构型不一样,所以还是有一定的差别。
染色体在细胞的有丝分裂间期由染色质螺旋化形成。
染色单体:有丝分裂前中期其实就是一条染色体复制,产生两条染色体,但着丝点未分裂,那一条染色体上就有2条染色单体所以说着两条染色单体式由复制形成的,应该是相同的同源染色体:形态、结构、遗传组成基本相同和在减数第一次分裂前期中彼此联会(配对),并且能够形成四分体,然后分裂到不同的生殖细胞的一对染色体,一个来自母方,另一个来自父方。
可见是在联会后才会有同源染色体的概念,所以是减数分裂前中期出现。
姐妹染色单体:是减数分裂时期同源染色体上的两条不同染色体上的染色单体出现在减数分裂前中期呃,特点,部分片段可以互换,进行交叉互换,也是基因重组的一种方式判断数量的话,首先你要搞清楚是什么细胞的什么时期,是体细胞还是生殖细胞,是有丝分裂还是减数分裂,前中后期,然后根据不同时期不同特点去数PS:数染色体数量要计数着丝点,那样不容易错数染色体就数着丝点,一个着丝点就是一条染色体姐妹染色单体数X形态的染色体,再乘2DNA,没复制的染色体,一条就一个DNA;复制后的染色体,一条两个DNA同源染色体:减数分裂可以配对的就一定是同源染色体组:二倍体的配子含的就是一个染色体组笔记:染色体:在生物的细胞核中,有一种易被碱性染料染上颜色的物质,叫做染色质。
染色体只是染色质的另外一种形态。
它们的组成成分是一样的,但是由于构型不一样,所以还是有一定的差别。
染色体在细胞的有丝分裂间期由染色质螺旋化形成。
染色单体:有丝分裂前中期其实就是一条染色体复制,产生两条染色体,但着丝点未分裂,那一条染色体上就有2条染色单体所以说着两条染色单体式由复制形成的,应该是相同的同源染色体:形态、结构、遗传组成基本相同和在减数第一次分裂前期中彼此联会(配对),并且能够形成四分体,然后分裂到不同的生殖细胞的一对染色体,一个来自母方,另一个来自父方。
(实验九)姊妹染色单体分染技术

〖材料和方法〗
1、蚕豆发芽处理。 2、BrdU、青霉素中黑暗培养。 3、秋水仙素溶液预处理,卡诺氏 液固定,HCL溶液离解,Schiff 试剂染色。 4、压片观察
〖作 业〗
1、 制所观察到的色差染色单体图 象。 2、 计算SCE的频率。 3、 思考:本实验的材料掺入 BrdU时为什么要在全黑的条件下 培养?
遗传学实验
实验九 姊妹染色单体分染技术 (3学时)
一、教学基本要求
1、 了解姊妹染色单体差别染色技术 的原理 2、 掌握制作SCE标本的方法。
二、教学内容
〖目的和要求〗 通过这一实验,要求学生了解姊 妹染色单体差别染色技术的原理和 制作SCE标本的方法,并通过S CE标本的观察,掌握SCE计数 方法。
Hale Waihona Puke 〖实验原理〗在DNA复制过程中,核苷的类似物5-溴脱氧 尿嘧啶核苷(BrdU)能代替胸腺嘧啶进入到新合成 的DNA链中,植物根尖在含有适当BrdU的培养液 中生长二个分裂周期后,中期染色体的两条染色单体 在化学组成上有了差别,一条染色单体的两条DNA 链均为新合成的,因此T位完全由BrdU代替,另一 条染色单体的一条DNA链是新合成的,所以只有一 条DNA中含有BrdU,这样的细胞经染色后,一条 为深染,一条为淡染,所以称之为姊妹染色单体的差 别染色。本实验运用这一原理,利用蚕豆根尖作为材 料,经过适当处理制片,可观察到姊妹染色单体分染 现象。
生物染色体染色单体姐妹染色体同源染色体染色体组概念的区分

『高中生物』染色体、染色单体、姐妹染色体、同源染色体、染色体组概念的区分.
染色体:在生物的细胞核中,有一种易被碱性染料染上颜色的物质,叫做染色质.染色体只是染色质的另外一种形态.它们的组成成分是一样的,但是由于构型不一样,所以还是有一定的差别.染色体在细胞的有丝分裂间期由染色质螺旋化形成.
染色体由蛋白质和DNA组成,在分裂期内绝对存在不会消失,在前期和中期每条染色体有两条染色单体组成.
判断染色体个数:有一个着丝点就有一个染色体
染色单体:有丝分裂前中期,其实就是一条染色体复制,产生两条染色体,但着丝点未分裂,那一条染色体上就有2条染色单体,所以说着两条染色单体式由复制形成的,应该是相同的.染色单体是染色体在间期复制的结果,染色单体的存在要么是染色体的两倍,要么是0.在有丝分裂和减数第二次分裂后期减为零.染色单体出现个数为偶数
姐妹染色单体:是减数分裂时期同源染色体上的两条不同染色体上的染色单体.出现在减数分裂前中期.特点,部分片段可以互换,进行交叉互换,也是基因重组的一种方式.
姐妹染色单体由同一条染色体复制而来,如果不考虑件数第一次分裂前期的交叉互换,那么姐妹染色单体完全相同.
P.S.说得简单些,你看到一个像有两条中间黏在一起的油条,那么这就是经过复制的染色体,黏在一起的部分叫着丝点.每一条“油条”就叫染色单体,又叫姐妹染色单体.
而没经过复制的染色体中只有一条,也只有一个着丝点
也可以说当看到一个有两条“油条”的染色体就有染色单体.
而要知道染色体数目,就要看着丝点的数量.。
姐妹染色单体课件

目 录
• 姐妹染色单体概述 • 姐妹染色单体的结构与组成 • 姐妹染色单体的复制与分离 • 姐妹染色单体在细胞分裂中的作用 • 姐妹染色单体异常与疾病的关系 • 姐妹染色单体的研究方法与技术应用
01
姐妹染色单体概述
定义与特点
定义
姐妹染色单体是指染色体复制过 程中产生的染色单体,它们在复 制过程中由同一条染色体复制而 来。
延长阶段
在DNA聚合酶的作用下,以解开 的单链模板为模板,按照碱基互 补配对原则,合成新的DNA链。
终止阶段
复制终止后,姐妹染色单体形成 。
分离过程
着丝粒分裂
有丝分裂过程中,着丝粒 分裂,姐妹染色单体开始 分离。
纺锤丝牵引
纺锤丝的牵引作用将姐妹 染色单体向两极移动。
分离完成
姐妹染色单体最终在赤道 板位置完全分开。
防止染色体断裂
姐妹染色单体在分裂过程中,能够防 止染色体的断裂和丢失,确保染色体 的稳定性。
对基因组重组的影响
促进基因重组
姐妹染色单体在减数分裂过程中,能够促进同源染色体的交叉互换,产生基因重组,增加生物多样性 。
维持基因组稳定性
姐妹染色单体在减数分裂过程中,能够确保基因组的稳定性,避免因重组错误导致的基因组变异。
遗传重组
姐妹染色单体的X形结构为遗传重组提供了基础。在减数分裂过程中, 同源染色体之间的交叉互换导致姐妹染色单体之间的交换,形成新的遗 传组合,为生物多样性的产生提供可能。
03
姐妹染色单体的复制与分离
复制过程
起始阶段
姐妹染色单体复制的起始阶段需 要依赖DNA复制酶和相关蛋白质 的参与,通过解旋酶的作用将双 螺旋DNA解开,形成单链模板。
染色单体和姐妹染色单体的区别

染色单体和姐妹染色单体的区别一、姐妹染色单体的概念:两条染色单体,其中之一是另一条染色单体的基因组DNA。
大多数天然产物中有几个不同的分子,每个都含有一定量的染色体和染色单体。
在基因工程中用于构建基因组DNA的人工染色体和染色单体称为“姐妹染色单体”。
常见的染色单体有两种形式:一种是可独立复制的完整的染色单体,叫做独立染色单体;另一种是能够与配子结合的染色单体片段,叫做重组染色单体。
这些染色单体可以是不同来源或不同类型的染色体片段重组形成的。
两个染色单体也可以是由不同的天然单体在化学结构上相连,例如玉米胚芽鞘的四分体的基因组DNA中含有一个染色体片段( X染色体)和一个染色单体片段( Y染色体)。
而对于真菌来说,四分体时期就只含有一条染色体,即四分体DNA中含有X、 Y各一条,它们是重组的姐妹染色单体。
二、辨别染色单体和重组染色单体1.在分类学上的意义。
凡属独立染色单体者,通常在分类上不占优势,在系统演化上也无特殊的作用。
但有少数植物的多数种类(如马齿苋属),天然分离的独立染色单体往往在生态习性和抗病性等方面具有某些特征,故可将独立染色单体分别置于种下的不同等级,作为单独种类处理,以简化分类,更有利于探索进化规律。
鉴别实验可采用木栓层的薄片制备,浸泡,观察分层情况,细胞质被溶解后所呈现的半透明度,红色、蓝色和紫色不同浓度的染色液以及显微摄影等手段。
如红色染液是外周木栓层细胞质的原生质,呈均匀的浅红色,蓝色染液是内皮层细胞质的原生质,呈均匀的蓝绿色,紫色染液是管胞的原生质,呈均匀的淡紫色。
其他染液不会被这三种染液所染色。
经过综合分析,在观察和测定的基础上,确定红色染液是维管束鞘细胞壁的细胞液,蓝色染液是维管束鞘细胞质,紫色染液是管胞原生质。
依此分出独立染色单体,从而获得许多科、属间形态、结构、生理和生态等方面的不同,有助于阐明各种植物的起源和进化。
如蓖麻中含有四分体的染色单体,可依据染色单体而区分这一类植物,从而认识到蓖麻的演化关系。
人淋巴细胞姐妹染色体区分染色实验原理及步骤

实验七人淋巴细胞姐妹染色体区分染色
一、试验原理:
姐妹染色体在正常情况下化学组成及结构都是一致的,因此不能用单纯染色方法将之区别开来。
1973年Latt发现让细胞在含Brdu的培养基中培养了2个细胞周期,再以Hoechst33258染色后,两条姐妹染色单体就出现了色差,这就是姐妹染色但提到区分染色(Sister Chromatid Diffeientiation SCD)。
当细胞在含Brdu的培养基中复制DNA时,Brdu会当作胸苷的类似物而掺入到DNA分子中,经过一个复制期,两条染色单体的DNA分子中都有一条是含Brdu的新链和不含Brdu的老链,称为BT-染色单体,BT-染色单体再进行一次复制,形成两条DNA分子。
其中一条DNA分子仍属于BT-型,另外一条则是两条单链都含有Brdu,到中期时为BB-染色单体,如图所示:。
实验四、姊妹染色单体分染技术(3学时)(综合性、设计性...

组成上有了差别,一条染色单体的两条DNA链均为
新合成的,因此T位完全由Brdu代替,另一条染色单
体的一条DNA链是新合成的,所以只有一条DNA
中含有Brdu,这样的细胞经染色后,一条为深染,一 条为淡染,所以称之为姊妹染色单体的差别染色。本
实验运用这一原理,利用大蒜根尖作为材料,经过适
当处理制片,可观察到姊妹染色单体分染现象。
五、实验说明
本实验需要综合应用到遗传学、动物学、植物学等相关
知识。 1. BrdU的参入是姊妹染色单体区分的重要基础。由于 植物种子萌发过程中自身合成的胸苷与外来的BrdU发 生竞争性矛盾,所以若处理不当往往会造成参入不进,
导致实验失败。因此一要注意选择饱满。发芽势强的蚕
豆种子;二是要注意抓住插入的时机,一定要在大多数 侧根长0.5-1.5cm时开始参入,因为此时侧根和幼芽的
(3)SO2含量: Schiff试剂中的SO2含量也影响孚尔根反
应的颜色表现。SO2含量低时呈红色,含量高时则偏向
蓝色。 (4)染色质中DNA的含量: 供试材料的染色质中DNA含 量的不同是影响显色反应强度的根本因素。不同生物和 不同的组织与细胞中DNA含量不同,显色强度也各异,
并且二者之间是正相关的。由于上述因素和实践了解,
使处理均匀分散,然后覆上两层吸水纸,以解剖针的木
柄端(或用铅笔上的橡皮头)敲击,使细胞进一步分离 铺展。
8、镜检观察: 选择染色体分散的离散细胞进行观察,在高 倍镜下即能看清姊妹染色单体间呈现的浓淡差 别。部分染色体上出现自发的姊妹染色单体互 换,凡在染色单体端部出现的互换计为一个 SCE,在中部出现的互换计为两个SCE。
水解是本实验成败的关键之一。温度应保持在600.5℃ 之间(用两支温度计相互校正)。如果温度过低或时间过 短,酸解不够,醛基暴露不充分,染色会过浅;若温度 过高或时间过长,会造成酸解过度,使DNA完全解聚, 糖与醛基之间的键被破坏,游离的核酸分子会扩散到细 胞质中,从而造成染色浅或不均一的现象。 水解的时间很重要,因为核酸的水解有两个过程。第 一,嘌呤碱很快被除掉,脱氧核糖中潜在的醛基显露出 来。第二,组蛋白和核酸越来越多地被除掉。在短时间 的水解作用以后,第一个过程占优势,这时候用Schiff试 剂染色,染色体的染色作用最强。随水解作用的继续进 行,第二个过程逐渐变成优势,因此水解液中的Schiff反 应增强,而染色体中的Schiff反应减弱。最后,第二个过 程超过第一个过程时,染色体也随之停止反应。
姊妹染色单体区分染色法

姊妹染色单体区分染色法(Sister chromatid differentiation,SCD)是70年代中期发展起来的染色体处理技术。
Latt(1973)在培养的细胞中加入5-溴脱氧尿嘧啶核苷(BrdU),当用Hoechst33258 荧光染料染色时,发现了姊妹染色单体的色差反映和它们之间互换的现象。
1974年KO Renberg和Froeed-Lender改进了这一技术,建立了较简易的BrdU-Giemsa技术。
这种技术用于研究细胞周期、染色体半保留复制、染色体的分子结构和畸变,以及DNA的复制、损伤与修复等一系列重要理论问题,还可以用于分析姊妹染色单体互换(Sister chromatid exchange, SCE)频率。
由于SCE能灵敏地检测染色体的变化,表现出剂量-效应关系。
因此,目前已把SCE列为检测致突变物、致癌物地常规指标之一。
一、原理5-溴脱氧尿嘧啶核苷(5-Bromodeoxy-urdine, BrdU)在DNA的复制过程中,掺入新合成的链并占有胸腺嘧啶(thymidine, T)的位置。
根据DNA的半保留复制规律,哺乳动物或人的细胞在BrdU的培养液中经历了两个周期后,它的两条姊妹染色单体的DNA双链在化学组成上有了差别。
当染色体的DNA链的两条多核苷酸链都被BrdU所替换,Giemsa染色显示浅色,如果染色体的DNA链中仅有一条多核苷酸链被BrdU所替换,Giemsa染色显示深色。
应用姊妹染色单体区分染色法(SCD)研究来自一个染色体的两条单体之间在同一个位点发生同源片段的交换,称为姊妹染色单体互换。
二、用品和试剂45℃水浴,紫外线灯(20W)。
余同外周血染色体制备。
试剂:BrdU溶液:用无菌青霉素瓶,在普通条件下称取BrdU 2mg,然后在无菌室内加入无菌生理盐水4ml,用黑纸避光,4℃冰箱保存,新鲜配置。
1×SSC溶液:0.15mol/L NaCl,0.015mol/L 柠檬酸钠。
染色体和染色单体姐妹染色单体和非姐妹染色单体同源染色体PPT精选文档
四、细胞分裂方式、所处时期及细胞名称的识别与判断 1.前期:
同源染色体 联会,出现 四分体
减数第 一次分 裂前期
初级精 (卵)母 细胞
有同源 染色体
有丝分 裂前期
体细胞
无同源 染色体
无同源 染色体 染色体 为奇数
减数 第二 次分 裂前 期
次级精 (卵) 母细胞
或第一 极体
9
2.中期:
有同 源染 色体
有性生殖细胞
染色体复制一次, 细胞连续分裂两次
1个卵或4个精子
染色体数目减一半
2N
N
同源染色体在第一次分裂发5 生
联会、交叉互换、分离等行为
三、细胞分裂中染色体及核DNA变化规律 (设体细胞的染色体数为2N)
有丝分裂
染色体行为变化: 决定
染色体 复制
出现染 色体和 纺锤体
染色体数目变化规律:
一个核内DNA
1、实际产生卵细胞的种类: 1
(含N对同源染色体)
2、可能产生卵细胞的种类: 2n
1个含有n对同源染色体的生物体 产生的精子或卵细胞种类 :2n
13
练习
1(1)A图表示 动物 细胞,判断的依据是无细胞壁有中心;体
(2)图A细胞内染色体、染色单体和DNA体的是图 A、B 。
一个四分体一对同源染色体两条染色体四条染色单体含四个dna分子8条脱氧核苷酸链染色体和染色单体姐妹染色单体和非姐妹染色单体同源染色体非同源染色体联会和四分体赤道板与细胞板细胞周期基本概念55区别区别有丝分裂有丝分裂减数分裂减数分裂形成细胞类型形成细胞类型染色体复制次数染色体复制次数细胞分裂次数细胞分裂次数形成子细胞数目形成子细胞数目染色体数目变化染色体数目变化有无同源染色体有无同源染色体的行为的行为体细胞有性生殖细胞染色体复制一次细胞分裂一次染色体复制一次细胞连续分裂两次亲子细胞染色体数目相等2n2n染色体数目减一半2n同源染色体在第一次分裂发生联会交叉互换分离等行为无同源染色体的联会交叉互换分离等行为二有丝分裂和减数分裂的比较66染色体复制出现染纺锤体着丝点排在赤着丝点分裂染色体平均移向两极染色体又变成染色质2n2n2n4n4n2n2a4a4a4a4a4a2a染色体数目变化规律
人类姐妹染色单体色差染色标本的制备和观察

原理:
当细胞在加有Brdu(5-溴脱氧尿核苷)培养基中 进行分裂时,Brdu能取代胸腺嘧啶核苷酸掺入到 新复制的DNA核苷酸链中。经过两个生长周期后, 这种细胞的同一染色体的两条姐妹染色单体一条 是由单股含Brdu的DNA链所组成,另一条是由双 股都含有Brdu的DNA链所组成,而这种双股都含 有Brdu的DNA链具有螺旋化程度较低的特性,在 热盐溶液中受光的照射后更易于水解,从而影响 了与Giemsa染料的亲和力,因此染成较浅的颜色。 而单股含Brdu的DNA链所组成的染色单体则着色 深。这样根据两条姐妹染色单体所显示的深浅不 同的颜色,可以区别中期染色体的姐妹染色单体。 即在普通显微镜下可见到两条姐妹染色单体显出 深浅不同的颜色。
SCE的计数 (1)凡是在染色单体端部出现互换记为1 次交换。 (2)在染色单体中间出现互换记为2次交 换。 (3)如果互换发生在着丝粒部位记为1次 交换。
姐妹染色单体交换(sister chromatid exchange,SCE)
在机体内外有害因素的作用下,染色体 的两条姐妹染色单体的片段会发生等位 点交换,称为姐妹染色单体交换。 由于SCE频率是DNA损伤的灵敏指标,而 且观察方便,故该技术已成为检测致突 变物对人染色体诱变效应大小的常用手 段。
标本片放在2×SSC溶液中,并置于60℃ 水浴。 紫外灯照射30分钟(灯与玻片的距离 5cm)。 1:10 Giemsa染液染色8分钟。 蒸馏水冲洗后干燥、镜检。
分裂周期分析: (1)染色体的两个单体均为深染的细胞记 为第一次分裂周期的细胞。 (2)染色体的两个单体一深一浅的细胞记 为第二次分裂周期的细胞。 (3)染色体的两个单体染色都为浅色的细 胞为第三次分裂周期的细胞。
实验十七姊妹染色单体色差方法

三、实验材料
• 人的外周血。
四、实验器具和药品
• 1.用具:2mL和1mL灭菌注射器,吸管,移 液管,离心管,量筒,烧杯,无菌青霉素 瓶,试剂瓶,培养瓶,酒精灯,载玻片, 天平,紫外灯(15W),离心机,恒温培 养箱,显微镜。
• 2.药品配制:RPMI1640,肝素,植物凝血 素(PHA),秋水仙素,青、链霉素,吉 姆萨染液等。
• 1974年Korenberg和Freedlender改进了这一方法,单 独用Giemsa染色即获得姐妹染色单体差别染色 (sister-chromatid differential staining简称SCD) (图24-1)。这是因为双股都含有BrdU核苷酸的DNA链 组成的染色单体,螺旋化程度较低,在热盐溶液中受 光的照射后更易于水解,从而影响了与Giemsa染料的 亲和力,因此染成较浅的颜色。这样根据两条姐妹染 色单体所显示的深浅不同的颜色,可以区别中期染色 体的姐妹染色单体。
实验十七 姊妹染色单 体色差方法
一、实验目的
• 1.子解姐妹染色单体差别染色技术的原理 和制作SCE标本的方法。
• 2.通过SCE标本的观察,掌握SCE计数方 法。
二、实验原理
• 在DNA复制过程中,核苷的类似物5-溴脱氧尿嘧啶核 苷(5-Bromodeoxy uridine简称BrdUrd)或5-碘尿嘧啶 核苷(5-Iodo-2′-deoxy uridine简称IdUrd)可以代替核 苷酸掺入新合成的DNA链,并占有胸腺嘧啶 (Thymidine,T)的位置。哺乳类细胞在含有适当浓度 的Brdurd的培养液中经历二个分裂周期之后,其中期 染色体的两个单体的DNA双链在化学组成上就有了差 别:即一条染色单体的两股DNA中的T位完全由 BrdUrd代替,而另一条染色单体的两股DNA中的一股 含BrdUrd,另一股则不含BrdUrd,这样的细胞经过制片 和苯并咪唑荧光染料(Hoechst-33258)染色后,在荧 光显微镜下可观察到两条姐妹染色单体显示强弱不同 的荧光。两股DNA链都含有BrdUrd的单体荧光较强, 其中只有一股有BrdUrd的单体荧光较弱。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
姊妹染色单体区分染色法(Sister chromatid differentiation,SCD)是70年代中期发展起来的染色体处理技术。
Latt(1973)在培养的细胞中加入5-溴脱氧尿嘧啶核苷(BrdU),当用Hoechst33258 荧光染料染色时,发现了姊妹染色单体的色差反映和它们之间互换的现象。
1974年KO Renberg和Froeed-Lender改进了这一技术,建立了较简易的BrdU-Giemsa技术。
这种技术用于研究细胞周期、染色体半保留复制、染色体的分子结构和畸变,以及DNA的复制、损伤与修复等一系列重要理论问题,还可以用于分析姊妹染色单体互换(Sister chromatid exchange, SCE)频率。
由于SCE能灵敏地检测染色体的变化,表现出剂量-效应关系。
因此,目前已把SCE列为检测致突变物、致癌物地常规指标之一。
一、原理
5-溴脱氧尿嘧啶核苷(5-Bromodeoxy-urdine, BrdU)在DNA的复制过程中,掺入新合成的链并占有胸腺嘧啶(thymidine, T)的位置。
根据DNA的半保留复制规律,哺乳动物或人的细胞在BrdU的培养液中经历了两个周期后,它的两条姊妹染色单体的DNA双链在化学组成上有了差别。
当染色体的DNA链的两条多核苷酸链都被BrdU所替换,Giemsa染色显示浅色,如果染色体的DNA链中仅有一条多核苷酸链被BrdU所替换,Giemsa染色显示深色。
应用姊妹染色单体区分染色法(SCD)研究来自一个染色体的两条单体之间在同一个位点发生同源片段的交换,称为姊妹染色单体互换。
二、用品和试剂
45℃水浴,紫外线灯(20W)。
余同外周血染色体制备。
试剂:BrdU溶液:用无菌青霉素瓶,在普通条件下称取BrdU 2mg,然后在无菌室内加入无菌生理盐水4ml,用黑纸避光,4℃冰箱保存,新鲜配置。
1×SSC溶液:0.15mol/L NaCl,0.015mol/L 柠檬酸钠。
三、操作步骤
l.细胞培养:常规培养人外周血淋巴细胞,24h后,加入BrdU使其终浓度为20μg/ml。
2.继续避光培养48小时,终止培养前2—3小时加秋水仙碱。
3.培养结束收获细胞,常规制备染色体,操作同实验一。
4.染色体制片在37℃恒温箱内老化24小时或室温放置1—2天。
5.将染色体制片的玻片正面向上平铺在恒温(45℃)水浴锅上,在玻片上滴加已预热至45℃的1×SSC溶液。
6.将紫外灯放在恒温水浴上,灯与标本垂直,其外加盖报纸数张以阻挡紫外线。
照射距离为6cm,时间15min。
7.照射完毕后以蒸馏水洗去1×SSC。
8.1∶10 Giemsa染色5分钟。
9.自来水细流冲洗去多余染料,干燥,镜检。
10.计数SCE。
选择染色体分散较好,数目为46的中期分裂相20个进行观察计数,
在染色单体端部出现的互换计为一次SCE,
在染色单体中间出现的互换计为两次SCE。
凡在着丝粒部位发生一次互换,判断不是两条染色单体在着丝粒部发生的扭转,计为一次SCE,但另列入“着丝粒区互换(CME)”一项。
Sister Chromosome Exchange (SCE)
【实验目的】
熟悉制备SCE标本的原理和基本程序。
【实验原理】
姊妹染色单体交换(Sister chromatialexchange,SCE)是染色体同源座位上复制产物间的相互交换,是同一染色体的两条单体之间发生的一类特殊的同源重组,主要在DNA合成期形成,可能与DNA双链的断裂与复制有关,SCE的发生的频率可反映细胞在S期的受损程度。
如果一个个体的SCE率明显增高,可表明染色体受到环境中的一定因素的影响,或是受到遗传缺陷的内在制约因素所致。
在细胞分裂时,每条染色体均有两条染色单体组成,每条染色单体有一条双链DNA组成。
5-溴脱氧尿嘧啶核苷(5-bromodeoxy- uridine,BrdU)是脱氧胸腺嘧啶核苷的类似物,在DNA链的复制过程中,可替代胸腺嘧啶。
当细胞生存环境中存在BrdU时,BrdU取代脱氧胸苷掺入到复制的DNA中。
经两个复制周期后,两条姊妹染色单体中一条DNA的双链均有BrdU掺入,而另一条DNA双链中仅有一条链有BrdU掺入。
利用特殊的分化染色技术对染色体标本进行处理,可使双链均含有BrdU掺入的单体浅染,而只有一条链掺入BrdU 的单体深染。
当姊妹染色体间存在同源片段交换时,可根据每条单体夹杂着深浅不一的着色片段加以区分。
由于姐妹染色单体的DNA序列相同,SCE并不改变遗传物质组成,但SCE是由于染色体发生断裂和重接而产生的,因此,SCE显示方法通常用来检测染色体断裂频率,用来研究药物和环境因素的致畸效应。
【实验用品】
1. 超净工作台、恒温培养箱、恒温水浴箱、冰箱、离心机、显微镜、采血器材、酒精灯、培养瓶、刻度离心管、胶塞、乳头吸管、试管架、载玻片、托盘天平、干燥烤箱、染色缸、电吹风、30W紫外灯。
2. 试剂:RPMI 1640培养液、小牛血清、秋水仙素(100μg/mL)、5-溴尿嘧啶(BrdU,200μg/mL)、低渗液(0.075 mol/L KCL溶液)、2×SSC、Giemsa原液、甲醇、冰乙酸。
【实验步骤】
1. 按半微量法采取外周静脉血,接种培养。
2. 37℃培养24小时后,加BrdU溶液(终浓度为8μg/mL)混匀。
3. 用黑纸包裹培养瓶,继续培养48小时。
4. 收获前加入秋水仙素(终浓度为0.2μg/mL),继续培养1小时。
5. 常规法制片。
将玻片标本室温放置2~3天。
6. 分化染色前一天,将标本置37℃过夜。
7. 在平皿中平行放置两根牙签,将玻片放在牙签上,加适量2×SSC溶液(以不超过标本为度)。
在标本上盖一张比玻片稍大的擦镜纸,使纸边浸入2×SSC溶液中,并使2×SSC溶液渗至玻片标本上,保持标本湿润。
8. 将平皿置55℃水浴面上,用30W紫外灯垂直照射标本30分钟,照射距离10cm。
9. 照射后轻轻取掉擦镜纸,立即用蒸馏水冲洗标本。
10. Giemsa染色5~10分钟;蒸馏水冲洗标本,气干。
11. 镜检,计数。
每个末端交换记为1次SCE,中部交换记为2次SCE。
【实验结果与分析】
观察30个细胞的分裂相,记录每个细胞的SCE总数,计算SCE频率。
中国人正常SCE频率为5.7±0.4。