一、 成果简介 - 南京农业大学研究生院
研究生学术不端行为的德育思考

用。 二 战后,《 军人权利法案 》的颁布 以及前苏联卫 星上天促使 美 国重视教育投 资, 美 国联邦政 府开始与大学建立 紧密的科研 合 作关 系。 大学在政 府巨额 财政支持下开始与外部世界接 触 , 大学 的工具 主义成为学 者新 的信念 。 在 巨大 利益 的诱 惑之下,
2 o 1 3 年 第 3 1 期
研究生学术不端行为的德 育思考
刘兆磊
摘要 : 学术 不端相伴学术行为而生。 研 究生学术不端现象被频繁披 露, 在一定范围内对大众心理和学术尊严产生了不可低估 的负面 影响。 研 究生学术不端行为受多种因素影响, 社会 环境 的恶化 、 研 究生培养及学术评价 制度的缺 陷、 导师的指导不力、自身学习动力不足
招收大 量研究 生 , 努力 向研 究型或教 学科 研型大学 转变 , 这 在
人格缺陷提 出追问。 虽有放大 的嫌疑 , 但不可否认 的是 , 高校研
究生德育工作面临严峻 的挑 战。
一
、
学 术 不 端 的 概 念界 定 和 问题 的 提 出
客观 上为 “ 拜 金”现象在 大学 的出现 提供 了契机 , 学术 不端 现 象开始引起 国内关注。 特 别是一些 知名学 者的不端行为被频 频
美 国学术界 出现 了一ห้องสมุดไป่ตู้ 因编造 或篡改实验数 据来争取研究项 目
2 0 0 7 年 中国科协发布 《 科 技工作者 科学道德规 范 ( 试行 ) 》, 对
学术 不端定义为 “ 在科学研究和学术活动中的各种造假、 抄袭、
剽窃和其他违背 科学共 同体惯 例的行为。 ”2 0 0 9 年 教育部针对 高校学术不 端行为, 将 其列为七种表现形式 : 抄袭 、 剽窃、 侵 吞
南京农业大学研究生学位论文答辩及-南京农业大学园艺学院

南京农业大学研究生学位论文答辩及学位申请工作暂行规定(二OO四年一月六日校学位评定委员会七届六次全会审议通过)根据《中华人民共和国学位条例》和《中华人民共和国学位条例暂行实施办法》,结合我校学位工作具体情况,特制定研究生学位论文答辩及学位申请工作暂行规定如下。
一、总则硕士研究生学位论文答辩及学位申请工作包括答辩资格的申请、论文评阅、答辩及学位授予等环节。
硕士学位论文的答辩及学位申请工作由各学院组织实施。
博士研究生学位论文答辩及学位申请工作包括预答辩资格的申请、预答辩、答辩资格的申请、论文评阅、答辩、学位授予及学位公示等环节。
凡向我校申请博士学位者,一律实行学位论文预答辩制度和论文“双盲”评阅制度。
博士学位论文的预答辩工作由各学院组织实施;博士学位论文的答辩及学位申请工作由研究生院学位办组织实施。
二、硕士学位论文答辩及学位申请(一)答辩申请资格硕士研究生须按计划修满本专业培养方案规定的全部课程,成绩合格,达到所规定的总学分,并参加一定时数的教学实习和社会实践,经研究生管理部门审核认可;完成了硕士学位论文的研究和撰写工作,经导师和学科点点长同意签字,方可申请硕士学位论文答辩。
(二)答辩的组织与表格领取硕士学位论文答辩由所在学院组织。
硕士学位论文答辩申请人凭学位论文答辩稿一份,向所在学院提出申请,资格审查通过后,领取《学位论文答辩申请表》(一份)、《毕业研究生登记表》(一式二份)、《硕士学位申请书》(一式三份)、《知识产权认定书》(一式三份)、《学位论文修改鉴定意见存查表》(一份)、《论文发表情况表》(一份)。
由答辩秘书认真填写《学位论文答辩申请表》,经所在学院分管领导签署意见并加盖公章后,送交所在学院研究生管理部门办理答辩手续。
(三)论文评阅人与答辩委员会硕士学位论文评阅人与答辩委员会成员由本学科点点长与导师协商提名,报学院分管领导审批同意并签名,方可正式聘请,并由各学院组织实施。
被江苏省学位委员会办公室抽检的学位论文评阅按江苏省教育评估院制订的办法(江苏省教育厅文件苏教研[2002]1号)实行。
高校与科研院所联合培养研究生的教育中心模式研究——以南京农业大学与江苏省农科院的合作为例

高校与科研院所联合培养研究生的教育中心模式研究——以南京农业大学与江苏省农科院的合作为例作者:董维春,刘晓光,宋力沁,詹群来源:《研究生教育研究》 2014年第2期董维春,刘晓光,宋力沁,詹群(南京农业大学公共管理学院,南京210095)摘要:高校与科研院所联合培养研究生是发挥双方资源优势,促进人才培养和科学研究深度融合的有效举措。
本文根据对南京农业大学江苏省农科院研究生教育中心的调查,分析其建设成效和存在的主要问题,进而提出相应的对策建议,以期为高校与科研院所联合培养研究生提供组织和制度借鉴。
关健词:高校;科研院所;联合培养研究生;研究生教育中心中图分类号: G643文献标识码: A收稿日期:2013-07-05作者简介:董维春(1964—),男,江苏兴化人,南京农业大学校长助理,发展规划与学科建设处处长,教授.刘晓光(1980—),男,河南项城人,南京农业大学公共管理学院副教授,管理学博士宋力沁(1975—),女,江西南昌人,南京农业大学公共管理学院硕士生詹群(1989—),女,湖北黄冈人,南京农业大学公共管理学院硕士生基金项目:2011年度江苏省高校哲学社会科学重点项目“科研院所与高等学校联合培养研究生的体制机制研究”(编号:2011ZDIXM030)研究生教育作为高等教育的最高层次,是培养拔尖创新人才的主要阵地。
为满足社会经济发展对高层次人才的需要,我国研究生招生规模近年来逐步扩大,学位结构日趋多样,全日制专业学位等应用型学位与研究生教育也迅速发展。
以高校为主体的传统培养模式,在新形势下面临教育资源短缺、培养目标不适应社会需求、课程与教学手段落后等诸多问题,研究生培养质量受到来自各方的质疑。
与此同时,中国科学院、各省市各行业科学研究院等机构经过多轮体制改革调整,资源优化程度逐步提高,在内外部环境的综合作用下,建立在科学研究基本职能基础上的人才培养工作日渐增多。
通过组织和管理创新有效发挥高校与科研机构的优势,提高研究生培养质量和社会适应能力,探索新形势下研究生培养模式的任务日趋紧迫。
研究生专业实践实习项目计划及时间安排

研究生专业实践实习项目计划及时间安排研究生专业实践实习项目计划及时间安排:全日制硕士专业学位研究生实践要求及计划表考核表南京农业大学全日制硕士专业学位研究生实习实践要求及管理规定为保证我校全日制专业学位研究生实习实践工作的顺利进行,切实有效地对研究生实习实践进行全过程管理、服务和考核,确保实践教学质量,根据《教育部关于做好全日制硕士专业学位研究生培养工作的若干意见》文件精神,结合我校实际情况制订本规定。
一、基本要求应用性和实践性是专业学位研究生培养的突出特点,科学有效的专业实践是专业学位研究生培养的重要环节,是专业学位研究生教育质量的重要保证。
通过专业实践,使专业学位研究生具有较强的解决实际问题能力,具备良好的综合素质和职业素养。
相关学院和指导教师应高度重视专业学位研究生的专业实践工作,在学校研究生院总体布臵和协调下,整体规划,统筹协调,主动与企事业单位等建立多种形式的实践基地或联合培养基地,加强专业学位研究生培养与用人单位实际需求的紧密联系,积极探索人才培养的供需互动机制。
二、组织管理全日制硕士专业学位研究生的实践环节由各学院落实和实施。
各学院须制定相关实施细则和方案,对专业实践的项目、模式、组织、管理和考核等问题作出规定,并由专门人员负责管理和联络。
三、实践模式专业实践的组织工作应贯彻和体现集中实践与分段实践相结合、校内实践和企业实践相结合、专业实践与论文工作相结合的原则。
专业实践可采取以下几种方式灵活进行。
1、由校内导师结合自身承担的应用型课题,安排学生的专业实践环节。
2、充分发挥校外导师的指导作用,利用企业的资源,由校外导师负责安排相应的专业实践环节。
3、依托省级、校级、学院等的企业研究生工作站、专业实践基地、研究生联合培养基地等,在校内外导师的共同指导下,由学院统一组织和选派学生去现场进行专业实践。
4、依托校内实验中心、工程中心和研究中心等单位,在导师的指导下进行以专业课程实践和科技技能训练为主的专业实践。
由博士学位论文盲审数据诱发的几点思考——以南京农业大学为例

由博士学位论文盲审数据诱发的几点思考李剑红,崔海燕,刘妍(南京农业大学研究生院,江苏南京210095)[摘要]通过列举近三年南京农业大学博士学位论文的盲审情况,分析专家在评审过程中存在的问题,由此引发对博士学位论文盲审的几点思考。
提出保证博士学位论文质量需要评审专家认真评审,学校严把培养环节质量关,关注和重视对博士学位论文完成过程的考核,做好对存在问题论文的处理工作,以期更好地完善博士学位论文盲审制度,让盲审工作在提高博士学位论文质量乃至提高博士研究生培养质量方面发挥更大的作用。
[关键词]学位论文;盲审;培养质量[基金项目]2015年度南京农业大学“研究生收费制度改革与培养质量研究———以南京农业大学为例”[作者简介]李剑红(1974—),女,江苏南京人,农业硕士,南京农业大学研究生院学位办科员,中级一级,主要从事研究生教育管理研究。
[中图分类号]G643[文献标识码]A [文章编号]1674-9324(2021)52-0027-04[收稿日期]2021-04-16———以南京农业大学为例国民教育体系中最高层次的博士研究生教育是衡量国家高等教育发达程度和科学文化发展水平的重要指标,提高博士研究生教育培养质量已成为我国教育改革发展的核心任务。
而博士学位论文质量是反映高校博士培养质量的重要标志之一,既能反映博士研究生掌握和综合运用知识的水平,也能体现其创新能力,同时能客观衡量培养院校的综合实力。
学位论文评审是衡量博士研究生学业水平高低,判断博士研究生是否符合答辩条件、达到学位要求的主要方法之一。
博士学位论文评审工作正是保证博士研究生水准的重要环节,是检测论文质量是否合格的重要途径。
博士学位论文是攻读博士学位的研究生在导师指导下,选择相关领域研究方向,进行继承、开拓、创新的研究工作后,最终形成的科研成果,较其他形式论文具有更高的学术价值,其科学性、创新性对学科领域的发展具有非常重要的推动作用,是衡量研究生培养质量的一项重要依据。
南京农业大学关于学术型研究生攻读学位期间发表学术论文要求的规定

南京农业大学关于学术型研究生攻读学位期间发表学术论文要求的规定(二〇一二年六月二十六日校学位委员会十届三次全会审议通过)为了提高我校研究生学位论文研究成果的水平,建立健全研究生学位质量保障体系,根据《中华人民共和国学位条例》、《中华人民共和国学位条例暂行实施办法》和《南京农业大学关于研究生学位论文答辩及学位申请工作的规定》,结合我校实际情况,特修订学术型研究生攻读学位期间发表学术论文要求的规定如下:一、总体要求本规定适用于我校2011级及以后学术型博士研究生和硕士研究生。
2011级以前博士研究生和硕士研究生攻读学位期间发表学术论文的要求按照相应时期学校及各学位分委员会的相关规定。
(一)博士研究生1.自然科学类学科须发表SCI、EI收录的学术论文1篇。
直博生、硕博连读生要求至少发表影响因子1.0以上的SCI或EI收录的学术论文1篇,或SCI、EI收录的学术论文2篇。
2.人文社会科学类学科须发表一类人文社会科学类核心期刊或SCI、EI、SSCI论文1篇,或三类及以上人文社会科学类核心期刊论文2篇(其中二类及以上刊物1篇),或三类及以上人文社会科学类核心期刊论文3篇。
国家重点学科的直博生、硕博连读生至少要求发表3篇人文社会科学类核心期刊论文(其中二类核心期刊论文至少2篇),或一类核心期刊或SCI、EI、SSCI收录的学术论文1篇。
(二)硕士研究生1.自然科学类学科须发表自然科学类核心期刊论文1篇;或研究生署名前三位的SCI、EI收录的学术论文1篇。
2.人文社会科学类学科须公开发表学术论文1篇。
二、具体要求1.提交的论文均须与学位论文内容有关,研究生为第一作者、导师须是作者之一,或导师为第一作者、研究生为第二作者。
作者第一署名单位以及论文第一通讯单位必须为南京农业大学。
文献综述(SCI期刊特约文献综述除外)、中文期刊增刊和会议论文集(包括EI 收录的会议论文集)不计。
2.对SCI源刊收录学术论文的文献类型的认可,由分委员会讨论确定。
南京农业大学研究生培养方案-观赏园艺
南京农业大学硕士研究生培养方案一、学科简介园林植物与观赏园艺学科分别于1983年和1989年经国家教委批准、在农业院校中建立第一个本科专业和硕士点,2003年设置观赏园艺博士点,现为校级重点学科。
学科现有师资16人,其中教授和兼职教授8人,副教授6人。
先后承担国家自然科学基金、教育部新世纪优秀人才支持计划、国家科技支撑计划、863计划等国家及省部级课题50余项。
在菊花种质资源搜集、保存和新品种选育及花卉采后生理与保鲜技术等研究领域居国内领先水平。
培育菊花新品种100余个,申请国家发明专利5项、菊花新品种保护权8件,国际登录梅花新品种12个,“菊花种质资源收集、保存、研究与利用”成果获江苏省科技进步二等奖。
主编了《中国菊花》、《观赏园艺概论》等专著和教材,在国内外核心刊物发表研究论文160余篇。
部分毕业生已成为教学、科研和农业推广部门的骨干人才。
二、培养目标1.培养德、智、体全面发展的园林植物与观赏园艺高级专门人才。
拥护党的基本路线,热爱祖国,遵纪守法,学风严谨,品德优良,艰苦奋斗,勇于创新。
2.掌握园林植物与观赏园艺科坚实的基础理论和系统的专门知识,掌握一门外国语,具有从事教学、科研和其他实际工作的能力。
3.身心健康。
三、研究方向1、观赏植物种质资源2、观赏植物遗传育种与分子生物学3、观赏植物生理生化4、观赏植物栽培七、开题报告研究生学位论文的开题报告应在学科内公开进行。
研究生的开题报告原则上在中期考核前完成,其中硕士生的开题报告审核通过后至少半年方可申请答辩。
八、中期考核:硕士生在第四学期初进行中期考核,根据其课程学习的学分是否满足要求及是否具有培养前途,决定是否进入学位论文阶段。
具体按研究生院有关规定执行。
九、学位论文:学位论文工作是研究生培养的重要组成部分,是对研究生进行科学研究或承担专门技术工作的全面训练,是培养研究生创新能力、综合运用所学知识发现问题、分析问题和解决问题能力的主要环节。
黏细菌捕食生物学研究进展及其在农业领域的应用潜力

㊀南京农业大学学报㊀2021ꎬ44(2):208-216http://nauxb.njau.edu.cn㊀JournalofNanjingAgriculturalUniversityDOI:10.7685/jnau.202010034收稿日期:2020-10-29基金项目:国家自然科学基金项目(32070027ꎬ32000101ꎬ31700054)ꎻ江苏省现代农业面上项目(BE2020340)ꎻ中国博士后科学基金项目(2020M671513)作者简介:李周坤ꎬ副研究员ꎬE ̄mail:zkl@njau.edu.cnꎮ∗通信作者:崔中利ꎬ教授ꎬ研究方向为环境微生物学ꎬE ̄mail:czl@njau.edu.cnꎮ李周坤ꎬ叶现丰ꎬ杨帆ꎬ等.黏细菌捕食生物学研究进展及其在农业领域的应用潜力[J].南京农业大学学报ꎬ2021ꎬ44(2):208-216.LIZhoukunꎬYEXianfengꎬYANGFanꎬetal.Thepredationbiologyofmyxobacteriaanditsapplicationinagriculturalfield[J].JournalofNanjingAgri ̄culturalUniversityꎬ2021ꎬ44(2):208-216.特约综述黏细菌捕食生物学研究进展及其在农业领域的应用潜力李周坤1ꎬ叶现丰1ꎬ杨帆1ꎬ黄彦1ꎬ范加勤2ꎬ王辉4ꎬ崔中利1ꎬ3∗(1.南京农业大学生命科学学院/农业农村部农业环境微生物重点实验室ꎬ江苏南京210095ꎻ2.南京农业大学植物保护学院ꎬ江苏南京210095ꎻ3.南京农业大学作物免疫学重点实验室ꎬ江苏南京210095ꎻ4.中国科学院南京土壤研究所土壤环境与污染修复重点实验室ꎬ江苏南京210008)摘要:黏细菌(myxobacteria)是一类具有多细胞群体行为特征的捕食性微生物类群ꎬ能够以活的微生物细胞或者其他生物大分子作为食物获取营养ꎬ同时能够形成抗逆性强的子实体和黏孢子ꎬ从而使黏细菌具有良好的环境适应性ꎮ黏细菌对于植物病原真菌和细菌的捕食特性使其在植物病害防治方面具有重要的应用潜力ꎬ被视为是新的生防微生物类型ꎮ本文综述黏细菌对于微生物的捕食机制以及捕食行为的生态学功能ꎬ概述捕食性黏细菌作为一种新型的生防微生物在病害防治方面的应用潜力ꎮ在此基础上ꎬ也讨论目前黏细菌捕食生物学研究存在的问题ꎬ旨在为黏细菌捕食作用的深入研究及其在农业生产上的实际应用提供参考ꎮ关键词:黏细菌ꎻ捕食性微生物ꎻ生防微生物ꎻ植物病害防治中图分类号:Q939.96㊀㊀㊀㊀文献标志码:A㊀㊀㊀㊀文章编号:1000-2030(2021)02-0208-09ThepredationbiologyofmyxobacteriaanditsapplicationinagriculturalfieldLIZhoukun1ꎬYEXianfeng1ꎬYANGFan1ꎬHUANGYan1ꎬFANJiaqin2ꎬWANGHui4ꎬCUIZhongli1ꎬ3∗(1.CollegeofLifeScience/KeyLaboratoryofAgriculturalEnvironmentalMicrobiologyꎬMinistryofAgricultureandRuralAffairsꎬNanjingAgriculturalUniversityꎬNanjing210095ꎬChinaꎻ2.CollegeofPlantProtectionꎬNanjingAgriculturalUniversityꎬNanjing210095ꎬChinaꎻ3.KeyLaboratoryofPlantImmunityꎬNanjingAgriculturalUniversityꎬNanjing210095ꎬChinaꎻ4.KeyLaboratoryofSoilEnvironmentandPollutionRemediationꎬInstituteofSoilScienceꎬChineseAcademyofSciencesꎬNanjing210008ꎬChina)Abstract:MyxobacteriaareGram ̄negativebacteriaubiquitousdetectedinsoilenvironment.Theyarehighlysocialmicrobeswithacomplexmulticellularpopulationbehaviorwhichareassociatedwithfeedingonabroadrangeofsoilbacteriaandfungitoachievenutri ̄tion.Theformationoffruitingbodyandmyxosporesfromvegetativecellsdifferentiationthatrespondstostarvationmaymakemyxobac ̄teriaexcellentadaptationinnaturalenvironment.Myxobacteriaareversatilepredatorsthatpreyonmicrobesasexcellentcandidatesforbiologicalcontrolagents(BCAs).Hereꎬwediscussedthemechanismsandecologicalfunctionsoftheubiquitousandaccomplishedgeneralistpredatorꎬandthebiocontrolpotentialofplantdiseaseusingmyxobacteriawasalsosummarized.Onthisbasisꎬthefuturechal ̄lengesfortheinvestigationofpredatorymyxobacteriaincontrollingplantpathogenswereprovidedꎬwhichmightprovideinsightsforthepracticalapplicationofmyxobacteriainagriculturalfield.Keywords:myxobacteriaꎻpredatorymicrobesꎻbiologicalcontrolagentsꎻbiocontrolofplantdisease植物病害是制约农作物优质高产的重要因素之一ꎬ据统计全球主要农作物的病害损失约占作物总产量的20%~40%ꎬ每年直接经济损失高达数十亿美元[1-2]ꎬ其中70%~80%的病害是病原真菌所致[3-4]ꎮ至2050年全球粮食产量需要增加70%以满足日益增长的人口需求ꎬ令人担忧的是自2000年以来ꎬ新的真菌类型或者真菌类植物病原菌(fungal ̄likeplantpathogen)呈现逐年增加的趋势[5]ꎬ粮食生产安全问题越来越受到关注ꎬ已成为最重要的国际问题之一[6]ꎮ902㊀第2期李周坤ꎬ等:黏细菌捕食生物学研究进展及其在农业领域的应用潜力目前ꎬ农业生产上的植物病害主要以化学防治为主ꎬ而过度依赖和滥用化学农药产生了有害生物抗药性㊁农药残留超标㊁环境污染等一系列问题ꎬ严重影响我国农业的绿色可持续发展[7]ꎮ为了避免过度依赖化学农药的农业病害防治现状ꎬ近年来利用微生物的抗菌㊁植物免疫调节以及根际或叶际微生物组调控作用ꎬ阻止病原菌入侵植物已成为有效策略[8-10]ꎮ生防微生物的抗菌方式具有多样性ꎬ目前ꎬ受到关注比较多的生防微生物主要来自于假单胞菌(Pseudomonas)㊁芽胞杆菌(Bacillus)㊁伯克氏菌(Burkholderia)㊁溶杆菌(Lysobacter)㊁木霉(Trichoderma)和腐霉(Pythium)等属[11-12]ꎬ其中以枯草芽胞杆菌㊁哈茨木霉和寡雄腐霉等为代表的生防菌已被开发成商业化微生物菌剂ꎬ应用于农业生产的病害控制ꎬ其主要作用机制包括拮抗作用㊁竞争作用㊁诱导植物系统抗性等ꎬ且孢子形成特性使其在菌剂长效保存和土壤生存方面具有一定的优势ꎮ生防微生物进入环境中受到环境因子的多变性[13]㊁植物与微生物互作过程中的免疫识别[14]以及植物根际调控[15]等因素的影响ꎬ生防微生物在开放环境中难以定殖且防治效果不稳定ꎬ导致生物菌剂的实际应用受到一定程度的限制ꎮ在自然生态系统中ꎬ存在着大量不同类型的生物体ꎬ这些生物个体之间为争夺养分和空间形成了复杂的生态网络结构ꎮ捕食是生物体之间广泛存在的一种相互作用模式ꎬ是构建生态系统群落结构和维持生物多样性的关键过程[16]ꎮ目前ꎬ捕食性微生物包括吸血球菌(Vampirococcus)㊁蛭弧菌(Bdellovibrio)㊁噬菌弧菌(Bacteriovorax)㊁Micavibrio㊁Daptobacter㊁拟杆菌(Bacteroidetes)和黏细菌(myxobacteria)等[17-18]ꎮ其中ꎬ蛭弧菌能够直接入侵细胞周质空间实现对革兰氏阴性细菌的捕食ꎬ被广泛应用于养殖产业中改善水质和治疗水生动物细菌性疾病等方面[19]ꎮ此外ꎬ研究人员发现Micavibrioaeruginosavorus可通过黏附到细菌的细胞壁上ꎬ掠夺它们的养分来维持自身的生存和繁殖ꎬ研究成果为治疗多种传染性疾病提供了依据ꎬ从而减缓了微生物耐药性问题[20]ꎮ利用微生物的捕食作用能实现对病原微生物的控制ꎬ为农业病害防治提供策略ꎮ黏细菌是一类具有多细胞群体行为特征的革兰氏阴性细菌ꎬ可以捕食包括细菌和真菌在内的多种微生物ꎬ存在于土壤㊁树皮㊁朽木㊁动物粪便㊁地衣和昆虫等不同类型的环境中[21]ꎮ黏细菌具有超大的基因组(大约10Mb)ꎬ能形成类似真菌的不同形态的子实体结构ꎬ被称为 高等原核生物 ꎮ该类群大多具有复杂的生活史和生长代谢调控过程以及较强的环境适应能力[22]ꎮ然而ꎬ黏细菌作为一类具有捕食特性的新型生防微生物ꎬ其在农业生产过程中植物病害控制方面的应用并未引起太多的关注ꎮ本文综述黏细菌对于微生物的捕食策略㊁抗菌机制㊁捕食的生态学功能等方面的研究进展ꎬ评估捕食性黏细菌在病害防治方面的应用潜力ꎮ同时ꎬ讨论目前黏细菌对于微生物的捕食研究方面所存在的问题及其应对策略ꎬ为黏细菌应用于农业生产过程的病害控制提供理论依据和策略ꎮ1㊀黏细菌是一类通才型(generalistpredator)的微生物捕食者捕食性微生物分布广泛ꎬ在5个门(Proteobacteria㊁Chloroflexi㊁Cytophagaceae㊁Actinobacteria和Nanoar ̄chaeota)中的15个科均发现了捕食性细菌ꎬ包括寄生于硬蜱属的立克次氏体(Rickettsia)等[23]ꎮ除细菌外ꎬ真菌中也存在具有捕食能力的类群ꎬ如Arthrobotrys等可以捕食线虫和一些微生物[24]ꎮ微生物的捕食作用可能是通才型的(generalistpredator)ꎬ如黏细菌ꎬ对不同类型的细菌和真菌均具有捕食作用[25-26]ꎻ也可能是专性的(obligatepredator)ꎬ如蛭弧菌ꎬ通过侵入革兰氏阴性细菌的周质空间进而实现对猎物细胞的分解[27]ꎮ然而ꎬ黏细菌和蛭弧菌都可以在固体界面上滑行运动实现捕食ꎬ但是所涉及的分子机制是相互独立的[28]ꎮ对于黏细菌的捕食研究开展的较早[29]ꎬ其捕食作用与已报道的几种捕食性微生物的作用方式不同ꎮ相对于以盘基网柄菌(Dictyosteliumdiscoideum)为代表的细胞吞噬作用㊁蛭弧菌(Bdellovibriobac ̄teriovorus)为代表的胞质入侵作用㊁溶杆菌(Lysobacter)为代表的分泌扩散性抗菌物质以及腐生螺旋体属(Saprospira)为代表的通过黏性物质捕捉猎物(Ixotrophy)等方式[30]ꎬ黏细菌利用一种特殊的胞外猎杀机制入侵猎物菌落ꎮ黏细菌的捕食策略类型分为:1)直接攻击模式(frontalattack)ꎮ黏细菌细胞间相互合作形成直接攻击模式ꎬ如黏细菌Myxococcussp.BS对软腐果胶杆菌等病原细菌的捕食作用[26]ꎮ2)狼群围捕攻击模式(wolfpackattack)ꎮ黄色黏球菌(Myxococcusxanthus)DZ2对苜蓿中华根瘤菌(Sinorhizobiummedicae)AK21的捕食过程中ꎬ由于菌株AK21产生大量的胞外半乳葡聚糖抵御捕食ꎬ因此黏细菌DZ2采取一种先围捕后猎杀的方式实现对猎物的捕食[31]ꎮ3)孤立捕食模式(solitarypredation)ꎮ通常认为黏细菌通过类似群体狩猎南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷的策略捕食猎物[30]ꎬ然而研究者也发现黏细菌单个细胞的孤立捕食也能实现对猎物细胞的猎杀[32-34]ꎮ基于自然环境中可获取资源的局限性ꎬ多样化的捕食策略有助于黏细菌类群在环境中的适应能力ꎮ2㊀捕食性黏细菌(predator)与猎物(prey)之间的多重博弈关系目前ꎬ关于黏细菌捕食作用的研究主要集中在细菌捕食方面ꎬ包括黏细菌产生的次级代谢物㊁裂解酶㊁外膜囊泡(OMVs)等[30ꎬ35-36]ꎬ其中次级代谢物的抗菌活性被认为在黏细菌捕食过程中发挥着重要的作用[37-38]ꎬ而外膜囊泡被认为是黏细菌攻击猎物的短距离 运输机 [39]ꎮ此外ꎬ黏细菌分泌的蛋白酶或肽酶㊁溶菌酶等裂解酶可能参与黏细菌的捕食作用[30ꎬ40]ꎬ但到目前为止并无直接的证据ꎮ在黏细菌捕食大肠杆菌的研究中ꎬ转录组学分析发现上千个基因响应捕食过程ꎬ同时推测猎物细胞壁和蛋白质是黏细菌攻击的首要目标[41]ꎮ与对细菌的捕食研究相比ꎬ黏细菌对真菌的捕食研究较少ꎬ仅涉及具有抗真菌活性的几丁质酶㊁β-1ꎬ3-葡聚糖酶等细胞壁裂解酶[42-44]以及抗真菌活性的次级代谢物[45]等ꎮ在细菌与真菌互作关系研究中ꎬ伯克霍尔德菌和沙雷氏菌分别进化出三型分泌系统(T3SS)和六型分泌系统(T6SS)ꎬ将毒性蛋白直接注入真菌细胞内实现对真菌的猎杀[46-47]ꎮ与之不同的是ꎬ黏细菌通过分泌一种新型外膜型β-1ꎬ6-葡聚糖酶分解真菌细胞壁中的β-1ꎬ6-葡聚糖组分ꎬ进而实现对植物病原真菌的捕食(图1)[48]ꎮ目前具有酶活性的外膜蛋白主要包括膜结合蛋白酶或酯酶等[49-50]ꎬ黏细菌来源的外膜型β-1ꎬ6-葡聚糖酶是目前已报道的唯一具有糖苷水解酶活性的外膜蛋白ꎬ具有广谱的抗真菌活性ꎬ是黏细菌捕食真菌的关键因子[48]ꎮ图1㊀黏细菌与猎物细胞之间的捕食与防御策略Fig 1㊀Theattack ̄defensemodelbetweenpredatorymyxobacteriaandprey捕食性黏细菌通过多种模式攻击猎物以获取营养并建立竞争优势ꎬ而猎物群体面对捕食者的攻击进化出相应的防御策略抵御微生物的捕食ꎬ从而实现种群的自我保护(图1)ꎮ猎物抵御黏细菌捕食的防御策略类型主要分为:1)产生抑菌物质ꎮ黏细菌可以分泌抗生素杀死猎物ꎬ而猎物也可以分泌抗菌物质抑制黏细菌的生长ꎮ在黄色黏球菌(M.xanthus)DK1622与天蓝色链霉菌(Streptomycescoelicolor)M45的互作研究中ꎬ链霉菌M45通过气生菌丝和抗菌物质抵御黏细菌的捕食[18]ꎬ在黏细菌与芽胞杆菌的互作研究中也有类似的抑制作用报道[51]ꎮ此外ꎬ真菌也能够产生具有生物活性的次级代谢物ꎬ如青霉菌产生的抗生素能够抑制炭疽杆菌的生长[52]ꎬ具有抑制黏细菌生长的可能ꎮ2)形成外围屏障ꎮ枯草芽胞杆菌(Bacillus012112㊀第2期李周坤ꎬ等:黏细菌捕食生物学研究进展及其在农业领域的应用潜力subtilis)NCIB3610和大肠杆菌与黄色黏球菌(M.xanthus)DK1622的互作研究中ꎬ芽胞杆菌和大肠杆菌作为猎物类群通过产生胞外基质和生物膜抵御黏细菌的入侵[53-54]ꎻ根瘤菌利用分泌的胞外半乳葡聚糖保护细胞免受黏细菌的攻击[31]ꎮ3)修饰捕食因子结构ꎮ黏细菌捕食细菌的一个重要武器是抗生素myxovirescin(TA)ꎬ地衣芽胞杆菌通过分泌糖基转移酶(YjiC)对黏细菌分泌的抗生素TA进行葡萄糖糖基化修饰ꎬ减弱其对自身细胞的毒性ꎬ进而逃脱黏细菌的捕食[55](图1)ꎮ4)共进化ꎮ地衣芽胞杆菌在黏细菌的捕食压力下进化出对抗生素TA的修饰能力[55]ꎻ在黏细菌与大肠杆菌共进化研究中发现ꎬ大肠杆菌通过增加黏液量降低黏细菌的运动速度ꎬ同时通过突变自身的毒力蛋白-外膜蛋白酶(OmpT)以适应捕食的压力ꎬ而黏细菌则通过突变1个未知的eatB基因从而增强对细菌捕食的适应性[56]ꎮ5)改变细胞壁结构组成ꎮ黏细菌通过分泌抗菌蛋白β-1ꎬ6-葡聚糖酶实现对真菌的猎杀ꎬ然而粗糙脉孢菌等微生物细胞壁不含β-1ꎬ6-葡聚糖ꎬ进而避免了黏细菌的捕食[48ꎬ57]ꎮ6)其他类型ꎮ改变猎物细胞表面成分与细胞形态(如丝状细胞)和增加游动速度等也能够保护猎物逃避捕食[58]ꎮ3㊀黏细菌捕食行为在土壤菌群生态调控中的功能微生物群落中的捕食涉及原生生物㊁噬菌体以及具有捕食能力的细菌和真菌等[59]ꎮ黏细菌作为微生物食物网结构中的捕食者ꎬ在土壤微生物食物网中代谢活跃ꎬ在土壤生态系统碳循环中起着关键作用[60]ꎮ土壤中黏细菌占总细菌群落的比例为0.4%~4.5%ꎬ几乎包含了所有黏细菌科或属ꎬ因此ꎬ黏细菌被认为是土壤细菌群落的重要组成部分[61]ꎮ此外ꎬ黏细菌在农田土壤环境中与捕食性细菌存在显著的正相关性ꎬ推测其在农田土壤细菌群落调控方面具有重要的作用[62]ꎮ尽管黏细菌广泛分布ꎬ并且在微生物生态调控中发挥着重要的作用ꎬ但对具体的影响或控制机制的研究并不统一ꎮ研究者在开展土壤微生物与植物互作关系研究中发现ꎬ黏细菌Corallococcussp.EGB能够对植物根际分泌物中麦芽糖和麦芽糖醇具有较强的趋化作用ꎬ使黏细菌向根部定向迁移并定殖ꎮ由于黏细菌EGB对包括尖孢镰刀菌在内的多种植物病原真菌和细菌均表现良好的捕食作用[25ꎬ48]ꎬ在向根部迁移的过程中ꎬ黏细菌通过捕食作用调控土壤微生物群落结构ꎮ其中ꎬBacillus和Pseudomonas等潜在的病害生防菌以及植物促生菌(PGPR)等丰度上升ꎬ尖孢镰刀菌黄瓜专化型(F.oxysporumf.sp.cucumerinumꎬFOC)数量明显下降ꎬ从而抑制病害的发生[63]ꎮ研究结果为利用捕食性黏细菌调控土壤微生物菌落进而实现植物病害的控制提供了新思路ꎬ同时也暗示着捕食性微生物作为土壤食物网的重要组成部分[64]ꎬ在微生物生态系统动态过程调控中起着重要的作用ꎮ4㊀捕食性黏细菌在植物病害控制方面的应用潜力研究发现来源于不同种属的黏细菌菌株对不同类型的植物病原菌均表现良好的抗菌活性[38ꎬ65-66]ꎬ在植物病害生物防治方面表现出潜在的应用价值ꎮ盆栽试验中ꎬ黏细菌对病原细菌㊁真菌和卵菌等造成的植物病害具有良好的生防效果ꎬ表现出较好的土壤定殖能力ꎬ从而保护植物免受病原菌的危害[25ꎬ63ꎬ67-68]ꎮ此外ꎬ在水果采后病害控制方面ꎬ黏细菌Corallococcussp.EGB产生的多种挥发性抗真菌次级代谢物(VOC)ꎬ能够有效抑制青霉菌对橘子的侵染[69]ꎬ延长水果采后货架期(图2)ꎮ基于黏细菌潜在的生防效果ꎬ研究者进一步开展了田间试验ꎮ黏细菌SorangiumcellulosumKYC3262在辣椒炭疽病的防控试验中连续3年表现出稳定的生防效果ꎬ防控效率与化学杀菌剂相当[70]ꎮ黏细菌Corallococcussp.EGB在连续2年的黄瓜和香蕉枯萎病田间防控试验中也表现出良好的生防效果ꎬ并优于化学药剂处理ꎬ显著提高作物产量[63] (图2)ꎮ与目前已报道的生防微生物相比ꎬ黏细菌在植物病害控制方面具有显著的特点:1)能在固体表面滑行运动ꎮ黏细菌利用2种不同类型的运动系统(A运动:AdventurousmotilityꎻS运动:Socialmotility)实现在固体界面的滑行运动[71]ꎬ降低微生物对土壤水分的要求[72]ꎬ而土壤水分含量是生防微生物在土壤中运动的限制因素之一[13]ꎮ2)产生丰富的抗菌物质ꎮ从黏细菌中已经发现了大量具有抗菌活性的次级代谢产物和酶类[43ꎬ73-74]ꎬ是原核生物中仅次于放线菌的第二大次级代谢物来源菌ꎮ3)发育形成抗性黏孢子ꎮ黏细菌是革兰氏阴性细菌ꎬ但可以分化或诱导分化形成具有抗逆性的孢子[75]ꎬ有利于生防菌剂的研发和保存ꎮ4)环境适应性强ꎮ黏细菌广泛分布于土壤㊁水体㊁腐败的树木枯枝落叶㊁草食类动物的粪便等不同的环境ꎬ具有较强的环境适应力[76-77]ꎮ5)能通过多样化的策略捕食真菌和细菌[30ꎬ78]ꎮ黏细菌通才型的南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷图2㊀黏细菌Corallococcussp.EGB在植物真菌病害控制方面的应用性评估Fig 2㊀ApplicationevaluationofofCorallococcussp.EGBinthebiocontrolofplantfungaldisease㊀㊀a.黏细菌利用挥发性抗菌物质(VOC)抑制气传性植物病原菌的生长[69](VOC:抑制灰霉病菌对橘子的侵染)ꎻb.黏细菌通过捕食和土壤微生物调控作用控制土传枯萎病害的发生[63](捕食与调控:抑制黄瓜枯萎病菌对黄瓜的侵染)ꎮa.Biocidaleffectsofvolatileorganiccompounds(VOC)producedbythemyxobacteriaagainstfungalphytopathogens[69]ꎻb.Predationandmicrobialcommunityregulationofmyxobacteriaareinvolvedinthecontrolofsoil ̄borneFusariumwilt[63].捕食特性㊁良好的环境适应性㊁土壤微生物群落调控能力ꎬ使其被视为是一类新型的生防微生物ꎬ可应用于农业生产过程中植物病害的生防控制ꎮ除了在植物病害生物防治方面ꎬ黏细菌在动物病害控制方面也表现一定的应用潜力ꎮ研究人员发现包含不同种属的113株黏细菌对多种动物致病菌均具有高效的捕食作用ꎬ包括肺炎克雷伯菌(Klebsiellapneumoniae)㊁奇异变形杆菌(Proteusmirabilis)㊁白色念珠菌(Candidaalbicans)㊁肠球菌(Enterococcus)㊁葡萄球菌(Staphylococcus)等[79]ꎮ此外ꎬ黏细菌分泌的具有生物活性的次级代谢物在保护动物健康方面也具有应用潜力ꎬ如S.cellulosum来源的Ambruticin等可以有效抑制多种动物致病菌的生长ꎻ来源于Myxococcusstipitatus的Rhizopodin㊁S cellulosum的Epothilone被认为在抗肿瘤方面具有重要的作用[35ꎬ80]ꎮ黏细菌通过捕食作用和次级代谢物的抗菌作用使其在致病性微生物引起的动物病害控制方面也具有一定的应用潜力ꎮ然而ꎬ目前黏细菌在病害防控中的作用还未引起足够的关注和重视ꎮ5㊀捕食性黏细菌在植物病害控制方面所面临的问题目前ꎬ捕食性黏细菌通过多种模式对植物病原菌进行捕食或者抑制ꎬ在病害控制方面具有良好的应用潜力ꎬ然而黏细菌捕食生物学研究中还存在一些瓶颈直接制约黏细菌的实际应用ꎮ例如:1)黏细菌捕食行为的复杂性ꎮ目前对于黏细菌的捕食相关研究多数集中在行为特征的描述等方面ꎬ已确定的黏细菌捕食因子只有抗生素TA和外膜型β-1ꎬ6-葡聚糖水解酶等ꎬ而黏细菌在捕食过程中降解酶与代谢调控㊁互作关键因子㊁感知与猎物响应㊁细胞依赖㊁群体捕食效率等方面的机制未知ꎬ限制了研究者对黏细菌捕食行为的深入了解ꎮ2)黏细菌生长与营养需求的特殊性ꎮ黏细菌特殊的生长发育方式导致黏细菌的分离周期长ꎬ且可培养性黏细菌资源有限ꎬ我国只有山东大学㊁广东省微生物研究所㊁内蒙古大学㊁河北大学等在黏细菌菌种资源收集方面建立了良好的基础ꎮ同时ꎬ黏细菌生长聚集成团和丰富的胞外多糖等特性使野生型黏细菌的遗传操作难以建立ꎬ具有优良特性的野生型黏细菌的作用机制研究存在瓶颈ꎮ此外ꎬ黏细菌生长过程中的自溶特性也导致扩大培养黏细菌受限ꎬ直接限制了黏细菌的实际应用和菌剂的规模化制备ꎮ3)研究材料的单一性ꎮ目前对于黏细菌的基础研究主要是以黄色黏球菌(M.xanthus)DK1622为材料ꎬ然而黏细菌与植物㊁微生物共进化过程中ꎬ不同种属之间的特性差异较大ꎬ如黄色黏球菌DK1622分泌的次级代谢物具有良好的抗菌作用ꎬ而黏细菌EGB主要是通过分泌真菌细胞壁裂解酶实现对真菌的抗性作212312㊀第2期李周坤ꎬ等:黏细菌捕食生物学研究进展及其在农业领域的应用潜力用ꎬ研究材料的单一性直接导致黏细菌捕食研究的进展较为缓慢ꎮ6 研究展望目前ꎬ黏细菌的基础研究主要是以黏细菌为模式生物ꎬ开展发育生物学㊁种群识别㊁进化生物学以及生态学等研究ꎬ包括黏细菌子实体的形成㊁运动性ꎬ多糖的生物合成ꎬ多形态细胞表面受体蛋白TraA(poly ̄morphiccellsurfacereceptor)及其互作蛋白TraB(cohortprotein)依赖的外膜融合参与黏细菌细胞识别的作用以及生物多样性等[81-82]ꎮ此外ꎬ以黏细菌为种质资源库ꎬ分离筛选一系列具有生物活性的次级代谢物ꎬ黏细菌已成为重要的生物活性物质来源菌[83]ꎮ黏细菌早在1941年就被报道具有捕食细菌的能力ꎬ然而黏细菌是如何完成对细菌和真菌的捕食这一关键科学问题至今未知ꎮ因此ꎬ解析黏细菌的捕食机制是未来黏细菌研究的重要方向ꎮ黏细菌在自然环境中分布广泛ꎬ具有较高的丰度ꎬ然而已分离培养的黏细菌资源依然较少ꎮ广东省微生物研究所科研人员利用病原菌作为被捕食菌构建了直接面向生物防治用途的黏细菌筛选模型[38]ꎬ为未来黏细菌分离方法的优化提供了方向ꎮ同时ꎬ2010年启动的地球微生物组计划(EarthMicrobiomeProject)ꎬ也为获取不同黏细菌的基因组信息ꎬ构建基因资源库提供了可能ꎮ此外ꎬ野生型黏细菌的分子生物学研究体系对于黏细菌研究的深入开展至关重要ꎮ研究者前期发现黏细菌细胞分散性㊁胞外多糖(exopolysaccharideꎬEPS)㊁限制-修饰系统(restriction ̄modificationsystemꎬR ̄Msystem)以及分泌系统等在黏细菌转化过程中起着重要的作用[84]ꎮ突破黏细菌胞外多糖的物理屏障ꎬ强化黏细菌生长过程细胞分散性ꎬ建立高效的遗传转化体系对于深入了解黏细菌的抗菌机制具有重要的作用ꎮ因此ꎬ为了促进捕食性黏细菌在农业生产过程中的实际应用ꎬ需要深入了解黏细菌捕食的作用机制和生态学功能ꎬ通过基因组学和培养组学等方法获取具有良好抗菌活性的优良菌株ꎻ利用代谢组学㊁蛋白和转录组学等鉴定参与黏细菌捕食行为的关键因子ꎬ系统解析黏细菌的捕食和代谢调控机制ꎻ结合微生物学㊁生态学㊁植物保护等多学科交叉阐明捕食性黏细菌在自然环境中的生态学功能以及与植物㊁土壤微生物菌群之间的互作关系ꎻ同时ꎬ建立和优化黏细菌规模化培养工艺ꎬ为黏细菌的实际应用提供依据ꎮ参考文献References:[1]㊀SavarySꎬWillocquetLꎬPethybridgeSJꎬetal.Theglobalburdenofpathogensandpestsonmajorfoodcrops[J].NatureEcology&Evolutionꎬ2019ꎬ3(3):430-439.[2]SavarySꎬFickeAꎬAubertotJNꎬetal.Croplossesduetodiseasesandtheirimplicationsforglobalfoodproductionlossesandfoodsecurity[J].FoodSecurityꎬ2012ꎬ4(4):519-537.[3]康振生.我国植物真菌病害的研究现状及发展策略[J].植物保护ꎬ2010ꎬ36(3):9-12.KangZS.CurrentstatusanddevelopmentstrategyforresearchonplantfungaldiseasesinChina[J].PlantProtectionꎬ2010ꎬ36(3):9-12(inChinesewithEnglishabstract).[4]CasadevallA.Fungaldiseasesinthe21stcentury:thenearandfarhorizons[J].Pathogens&Immunityꎬ2018ꎬ3(2):183-196. [5]FisherMCꎬHenkDAꎬBriggsCJꎬetal.Emergingfungalthreatstoanimalꎬplantandecosystemhealth[J].Natureꎬ2012ꎬ484(7393):186-194.[6]KeinanAꎬClarkAG.Recentexplosivehumanpopulationgrowthhasresultedinanexcessofraregeneticvariants[J].Scienceꎬ2012ꎬ336(6082):740-743.[7]王桂荣ꎬ王源超ꎬ杨光富ꎬ等.农业病虫害绿色防控基础的前沿科学问题[J].中国科学基金ꎬ2020ꎬ34(4):374-380.WangGRꎬWangYCꎬYangGFꎬetal.Frontiersinscientificissuesofcontrollingagriculturalpestsanddiseasesbyenvironmental ̄friendlymethods[J].BulletinofNationalNaturalScienceFoundationofChinaꎬ2020ꎬ34(4):374-380(inChinesewithEnglishabstract). [8]ChenTꎬNomuraKꎬWangXLꎬetal.Aplantgeneticnetworkforpreventingdysbiosisinthephyllosphere[J].Natureꎬ2020ꎬ580(7805):653-657.[9]WeiZꎬGuYAꎬFrimanVPꎬetal.Initialsoilmicrobiomecompositionandfunctioningpredeterminefutureplanthealth[J].ScienceAdvancesꎬ2019ꎬ5(9):eaaw0759.[10]TringeSG.Alayereddefenseagainstplantpathogens[J].Scienceꎬ2019ꎬ366(6465):568-569.[11]LegeinMꎬSmetsWꎬVandenheuvelDꎬetal.Modesofactionofmicrobialbiocontrolinthephyllosphere[J].FrontiersinMicrobiologyꎬ2020ꎬ11:1619.[12]RahmanSFSAꎬSinghEꎬPieterseCMJꎬetal.Emergingmicrobialbiocontrolstrategiesforplantpathogens[J].PlantScienceꎬ2018ꎬ267:102-111.[13]BabalolaOO.Beneficialbacteriaofagriculturalimportance[J].BiotechnologyLettersꎬ2010ꎬ32(11):1559-1570.412南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷[14]Trd LꎬBoutrotFꎬClaverieJꎬetal.Perceptionofpathogenicorbeneficialbacteriaandtheirevasionofhostimmunity:patternrecognitionreceptorsinthefrontline[J].FrontiersinPlantScienceꎬ2015ꎬ6:219.[15]ZhalninaKꎬLouieKBꎬHaoZꎬetal.Dynamicrootexudatechemistryandmicrobialsubstratepreferencesdrivepatternsinrhizospheremicrobialcommunityassembly[J].NatureMicrobiologyꎬ2018ꎬ3(4):470-480.[16]ErkenMꎬLutzCꎬMcDougaldD.Theriseofpathogens:predationasafactordrivingtheevolutionofhumanpathogensintheenvironment[J].MicrobialEcologyꎬ2013ꎬ65(4):860-868.[17]GuerreroRꎬPedros ̄AlioCꎬEsteveIꎬetal.Predatoryprokaryotes:predationandprimaryconsumptionevolvedinbacteria[J].ProcNatlAcadSciUSAꎬ1986ꎬ83(7):2138-2142.[18]PérezJꎬMoraleda ̄MuñozAꎬMarcos ̄TorresFJꎬetal.Bacterialpredation:75yearsandcounting![J].EnvironmentalMicrobiologyꎬ2016ꎬ18(3):766-779.[19]陈康勇ꎬ钟为铭ꎬ高志鹏.蛭弧菌在水产养殖中应用研究进展[J].水产科学ꎬ2018ꎬ37(2):283-288.ChenKYꎬZhongWMꎬGaoZP.ResearchprogressonutilizationofBdellovibrioinaquaculture[J].FisheriesScienceꎬ2018ꎬ37(2):283-288(inChinesewithEnglishabstract).[20]WangZꎬKadouriDEꎬWuM.Genomicinsightsintoanobligateepibioticbacterialpredator:MicavibrioaeruginosavorusARL ̄13[J].BMCGenomicsꎬ2011ꎬ12:453.[21]李曙光.黏细菌的环境分布㊁季节演替及其相互作用[D].济南:山东大学ꎬ2014.LiSG.Distributionꎬseasonalsuccessionandintraspeciesinteractionsofmyxobacteria[D].Jinan:ShandongUniversityꎬ2014(inChinesewithEnglishabstract).[22]王春玲ꎬ冯广达ꎬ姚青ꎬ等.黏细菌基因组学研究进展[J].微生物学通报ꎬ2019ꎬ46(9):2394-2403.WangCLꎬFengGDꎬYaoQꎬetal.Researchprogressingenomicsofmyxobacteria[J].MicrobiologyChinaꎬ2019ꎬ46(9):2394-2403(inChinesewithEnglishabstract).[23]JurkevitchE.Predatorybehaviorsinbacteria:diversityandtransitions[J].MicrobeMagazineꎬ2007ꎬ2(2):67-73.[24]BarronGL.Predatoryfungiꎬwooddecayꎬandthecarboncycle[J].Biodiversityꎬ2003ꎬ4(1):3-9.[25]LiZKꎬYeXFꎬChenPLꎬetal.AntifungalpotentialofCorallococcussp.strainEGBagainstplantpathogenicfungi[J].BiologicalControlꎬ2017ꎬ110:10-17.[26]LiZKꎬWangTꎬLuoXꎬetal.BiocontrolpotentialofMyxococcussp.strainBSagainstbacterialsoftrotofCallalilycausedbyPectobacteriumcarotovorum[J].BiologicalControlꎬ2018ꎬ126:36-44.[27]DavidovYꎬHuchonDꎬKovalSFꎬetal.Anewalpha ̄proteobacterialcladeofBdellovibrio ̄likepredators:implicationsforthemitochondrialendo ̄symbiotictheory[J].EnvironmentalMicrobiologyꎬ2006ꎬ8(12):2179-2188.[28]ZhangYꎬGuzzoMꎬDucretAꎬetal.AdynamicresponseregulatorproteinmodulatesG ̄protein ̄dependentpolarityinthebacteriumMyxococcusxanthus[J].PLoSGeneticsꎬ2012ꎬ8(8):e1002872.[29]BeebeJM.Studiesonthemyxobacteria:ⅠꎬdistributioninIowasoilsanddescriptionofanewspeciesꎻⅡꎬMyxobacteriaasbacterialparasitesꎻⅢꎬthemorphologyandcytologyofMyxococcusxanthus[D].Iowa:IowaStateUniversityꎬ1941.[30]BerlemanJEꎬKirbyJR.Decipheringthehuntingstrategyofabacterialwolfpack[J].FEMSMicrobiologyReviewsꎬ2009ꎬ33(5):942-957. [31]PérezJꎬJiménez ̄ZurdoJIꎬMartínez ̄AbarcaFꎬetal.RhizobialgalactoglucandeterminesthepredatorypatternofMyxococcusxanthusandprotectsSinorhizobiummelilotifrompredation[J].EnvironmentalMicrobiologyꎬ2014ꎬ16(7):2341-2350.[32]ZhangWCꎬWangYꎬLuHNꎬetal.DynamicsofsolitarypredationbyMyxococcusxanthusonEscherichiacoliobservedatthesingle ̄celllevel[J].AppliedandEnvironmentalMicrobiologyꎬ2019ꎬ86(3):e02286-19.[33]McBrideMJꎬZusmanDR.BehavioralanalysisofsinglecellsofMyxococcusxanthusinresponsetopreycellsofEscherichiacoli[J].FEMSMicrobiologyLettersꎬ1996ꎬ137(2/3):227-231.[34]ShiloM.Lysisofblue ̄greenalgaebymyxobacter[J].JournalofBacteriologyꎬ1970ꎬ104(1):453-461.[35]KaurRꎬKumariAꎬKaurRꎬetal.Myxobacteria:producersofenormousbioactivesecondarymetabolites[J].InternationalJournalofResearchinPharmaceuticalSciencesꎬ2018ꎬ9(1):309-313.[36]EvansAGLꎬDaveyHMꎬCooksonAꎬetal.PredatoryactivityofMyxococcusxanthusouter ̄membranevesiclesandpropertiesoftheirhydrolasecargo[J].Microbiologyꎬ2012ꎬ158(11):2742-2752.[37]XiaoYꎬWeiXMꎬEbrightRꎬetal.Antibioticproductionbymyxobacteriaplaysaroleinpredation[J].JournalofBacteriologyꎬ2011ꎬ193(18):4626-4633.[38]代京莎ꎬ李安章ꎬ朱红惠.黏细菌在植物病害生物防治中的作用[J].生物技术进展ꎬ2016ꎬ6(4):229-234.DaiJSꎬLiAZꎬZhuHH.Thefunctionofmyxobacteriainbiologicalcontrolofplantdisease[J].CurrentBiotechnologyꎬ2016ꎬ6(4):229-234(inChinesewithEnglishabstract).[39]KeaneRꎬBerlemanJ.ThepredatorylifecycleofMyxococcusxanthus[J].Microbiologyꎬ2016ꎬ162(1):1-11.[40]EnsignJꎬWolfeR.Characterizationofasmallproteolyticenzymewhichlysesbacterialcellwalls[J].Journalofbacteriologyꎬ1966ꎬ91(2):524-534.[41]LivingstonePGꎬMillardADꎬSwainMTꎬetal.TranscriptionalchangeswhenMyxococcusxanthuspreysonEscherichiacolisuggestmyxobacterialpredatorsareconstitutivelytoxicbutregulatetheirfeeding[J].MicrobialGenomicsꎬ2018ꎬ4(2):e000152.512㊀第2期李周坤ꎬ等:黏细菌捕食生物学研究进展及其在农业领域的应用潜力[42]HockingDꎬCookFD.Myxobacteriaexertpartialcontrolofdamping ̄offandrootdiseaseincontainer ̄growntreeseedlings[J].CanadianJournalofMicrobiologyꎬ1972ꎬ18(10):1557-1560.[43]LiZKꎬXiaCYꎬWangYXꎬetal.Identificationofanendo ̄chitinasefromCorallococcussp.EGBandevaluationofitsantifungalproperties[J].InternationalJournalofBiologicalMacromoleculesꎬ2019ꎬ132:1235-1243.[44]ZhouJꎬChenJHꎬLiZKꎬetal.Enzymaticpropertiesofamulti ̄specificβ ̄(1ꎬ3) ̄glucanasefromCorallococcussp.EGBanditspotentialantifungalapplications[J].ProteinExpressionandPurificationꎬ2019ꎬ164:105481.[45]KunzeBꎬSteinmetzHꎬHöfleGꎬetal.Cruentarenꎬanewantifungalsalicylate ̄typemacrolidefromByssovoraxcruenta(Myxobacteria)withinhibitoryeffectonmitochondrialATPaseactivity[J].TheJournalofAntibioticsꎬ2006ꎬ59(10):664-668.[46]SwainDMꎬYadavSKꎬTyagiIꎬetal.Aprophagetail ̄likeproteinisdeployedbyBurkholderiabacteriatofeedonfungi[J].NatureCommunicationsꎬ2017ꎬ8(1):1-9.[47]TrunkKꎬPeltierJꎬLiuYCꎬetal.ThetypeⅥsecretionsystemdeploysantifungaleffectorsagainstmicrobialcompetitors[J].NatureMicrobiologyꎬ2018ꎬ3(8):920-931.[48]LiZKꎬYeXFꎬLiuMXꎬetal.Anoveloutermembraneβ ̄1ꎬ6 ̄glucanaseisdeployedinthepredationoffungibymyxobacteria[J].TheISMEJournalꎬ2019ꎬ13(9):2223-2235.[49]RuttenLꎬMannieJPBAꎬSteadCMꎬetal.Active ̄sitearchitectureandcatalyticmechanismofthelipidAdeacylaseLpxRofSalmonellatyphimurium[J].ProcNatlAcadSciUSAꎬ2009ꎬ106(6):1960-196.[50]FairmanJWꎬNoinajNꎬBuchananSK.Thestructuralbiologyofβ ̄barrelmembraneproteins:asummaryofrecentreports[J].CurrentOpinioninStructuralBiologyꎬ2011ꎬ21(4):523-531.[51]MüllerSꎬStrackSNꎬHoeflerBCꎬetal.BacillaeneandsporulationprotectBacillussubtilisfrompredationbyMyxococcusxanthus[J].AppliedandEnvironmentalMicrobiologyꎬ2014ꎬ80(18):5603-5610.[52]BillsGFꎬGloerJB.Biologicallyactivesecondarymetabolitesfromthefungi[J].MicrobiologySpectrumꎬ2016ꎬ4(6):1-32.[53]MüllerSꎬStrackSNꎬRyanSEꎬetal.PredationbyMyxococcusxanthusinducesBacillussubtilistoformspore ̄filledmegastructures[J].AppliedandEnvironmentalMicrobiologyꎬ2015ꎬ81(1):203-210.[54]DepasWHꎬSyedAKꎬSifuentesMꎬetal.BiofilmformationprotectsEscherichiacoliagainstkillingbyCaenorhabditiselegansandMyxococcusxanthus[J].AppliedandEnvironmentalMicrobiologyꎬ2014ꎬ80(22):7079-7087.[55]WangCꎬLiuXꎬZhangPꎬetal.BacilluslicheniformisescapesfromMyxococcusxanthuspredationbydeactivatingmyxovirescinAthroughenzymaticglucosylation[J].EnvironmentalMicrobiologyꎬ2019ꎬ21(12):4755-4772.[56]NairRRꎬVasseMꎬWielgossSꎬetal.Bacterialpredator ̄preycoevolutionacceleratesgenomeevolutionandselectsonvirulence ̄associatedpreydefences[J].NatureCommunicationsꎬ2019ꎬ10(1):1-10.[57]MaddiAꎬDettmanAꎬFuCꎬetal.WSC ̄1andHAM ̄7areMAK ̄1MAPkinasepathwaysensorsrequiredforcellwallintegrityandhyphalfusioninNeurosporacrassa[J].PLoSOneꎬ2012ꎬ7(8):e42374.[58]JoussetA.Ecologicalandevolutiveimplicationsofbacterialdefencesagainstpredators[J].EnvironmentalMicrobiologyꎬ2012ꎬ14(8):1830-1843. [59]ThakurMPꎬGeisenS.Trophicregulationsofthesoilmicrobiome[J].TrendsinMicrobiologyꎬ2019ꎬ27(9):771-780.[60]LuedersTꎬKindlerRꎬMiltnerAꎬetal.Identificationofbacterialmicropredatorsdistinctivelyactiveinasoilmicrobialfoodweb[J].AppliedandEnvironmentalMicrobiologyꎬ2006ꎬ72(8):5342-5348.[61]ZhouXWꎬLiSGꎬLiWꎬetal.Myxobacterialcommunityisapredominantandhighlydiversebacterialgroupinsoilniches[J].EnvironmentalMicrobiologyReportsꎬ2014ꎬ6(1):45-56.[62]WangWHꎬLuoXꎬYeXFꎬetal.PredatoryMyxococcalesarewidelydistributedinandcloselycorrelatedwiththebacterialcommunitystructureofagriculturalland[J].AppliedSoilEcologyꎬ2020ꎬ146:103365.[63]YeXFꎬLiZKꎬLuoXꎬetal.ApredatorymyxobacteriumcontrolscucumberFusariumwiltbyregulatingthesoilmicrobialcommunity[J].Microbiomeꎬ2020ꎬ8(1):49.[64]Mendes ̄SoaresHꎬVelicerGJ.Decomposingpredation:testingforparametersthatcorrelatewithpredatoryperformancebyasocialbacterium[J].MicrobialEcologyꎬ2013ꎬ65(2):415-423.[65]任兴波ꎬ张子良ꎬ赵璞钰ꎬ等.马铃薯晚疫病菌拮抗黏细菌YR ̄35的分离鉴定及其代谢产物稳定性[J].中国生物防治学报ꎬ2016ꎬ32(3):379-387.RenXBꎬZhangZLꎬZhaoPYꎬetal.IsolationandidentificationofthestrainYR ̄35resistanttophytophthorainfestansanditsmetabolites[J].ChineseJournalofBiologicalControlꎬ2016ꎬ32(3):379-387(inChinesewithEnglishabstract).[66]李百元ꎬ谢小林ꎬ张鲜娇ꎬ等.不同被捕食细菌对新疆盐碱地黏细菌分离的影响[J].微生物学报ꎬ2013ꎬ53(4):379-389.LiBYꎬXieXLꎬZhangXJꎬetal.Influenceofdifferentpreystrainsonisolationofmyxobacteriainsaline ̄alkalinesoilsofXinjiang[J].ActaMicrobiologicaSinicaꎬ2013ꎬ53(4):379-389(inChinesewithEnglishabstract).[67]KimSTꎬYunSC.BiocontrolwithMyxococcussp.KYC1126againstanthracnoseinhotpepper[J].ThePlantPathologyJournalꎬ2011ꎬ27(2):156-163.[68]DahmMꎬBrzezińskaAJꎬWrótniak ̄DrzewieckaWꎬetal.Myxobacteriaasapotentialbiocontrolagenteffectiveagainstpathogenicfungiofeconomicallyimportantforesttrees[J].Dendrobiologyꎬ2015ꎬ74:13-24.[69]YeXFꎬChenYꎬMaSYꎬetal.BiocidaleffectsofvolatileorganiccompoundsproducedbythemyxobacteriumCorrallococcussp.EGBagainst612南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷fungalphytopathogens[J].FoodMicrobiologyꎬ2020ꎬ91:103502.[70]YunSC.Selectionanda3 ̄yearfieldtrialofSorangiumcellulosumKYC3262againstanthracnoseinhotpepper[J].ThePlantPathologyJournalꎬ2014ꎬ30(3):279-287.[71]NanBꎬZusmanDR.Uncoveringthemysteryofglidingmotilityinthemyxobacteria[J].AnnualReviewofGeneticsꎬ2011ꎬ45:21-39. [72]SpormannAM.Glidingmotilityinbacteria:insightsfromstudiesofMyxococcusxanthus[J].MicrobiologyandMolecularBiologyReviewsꎬ1999ꎬ63(3):621-641.[73]刘新利ꎬ李越中.黏细菌次级代谢产物及其在农业上的应用价值[J].中国农业科技导报ꎬ2007ꎬ9(3):44-51.LiuXLꎬLiYZ.Myxobacterialsecondarymetabolitesandtheirpotentialapplicationsinagriculture[J].JournalofAgriculturalScienceandTechnologyꎬ2007ꎬ9(3):44-51(inChinesewithEnglishabstract).[74]KaurRꎬSinghSꎬKaurRꎬetal.Myxococcusxanthus:asourceofantimicrobialsandnaturalbio ̄controlagent[J].ThePharmaInnovationJour ̄nalꎬ2017ꎬ6(11):260-262.[75]DworkinM.Recentadvancesinthesocialanddevelopmentalbiologyofthemyxobacteria[J].MicrobiologyReviewꎬ1996ꎬ60(1):70-102. [76]DawidW.Biologyandglobaldistributionofmyxobacteriainsoils[J].FEMSMicrobiologyReviewsꎬ2000ꎬ24(4):403-427.[77]李曙光ꎬ周秀文ꎬ吴志红ꎬ等.黏细菌的种群生态及其生存策略[J].微生物学通报ꎬ2013ꎬ40(1):172-179.LiSGꎬZhouXWꎬWuZHꎬetal.Populationecologyandsurvivalstrategyofmyxobacteria[J].MicrobiologyChinaꎬ2013ꎬ40(1):172-179(inChinesewithEnglishabstract).[78]Muñoz ̄DoradoJꎬMarcos ̄TorresFꎬGarcia ̄BravoEꎬetal.Myxobacteria:movingꎬkillingꎬfeedingꎬandsurvivingtogether[J].FrontiersinMicrobiologyꎬ2016:781.[79]LivingstonePGꎬMorphewRMꎬWhitworthDE.Myxobacteriaareabletopreybroadlyuponclinically ̄relevantpathogensꎬexhibitingapreyrangewhichcannotbeexplainedbyphylogeny[J].FrontiersinMicrobiologyꎬ2017ꎬ8:1593.[80]刘新利ꎬ李越中.黏细菌资源与埃博霉素研发[J].生物产业技术ꎬ2011(2):26-32.LiuXLꎬLiYZ.Myxobacteriaresourcesanddevelopmentofepothilone[J].Biotechnology&Businessꎬ2011(2):26-32(inChinese). [81]CaoPꎬWallD.Directvisualizationofamolecularhandshakethatgovernskinrecognitionandtissueformationinmyxobacteria[J].NatureCommunicationsꎬ2019ꎬ10(1):3073.[82]YuYTNꎬYuanXꎬVelicerGJ.AdaptiveevolutionofansRNAthatcontrolsMyxococcusdevelopment[J].Scienceꎬ2010ꎬ328(5981):993. [83]ReichenbachH.Myxobacteriaꎬproducersofnovelbioactivesubstances[J].JournalofIndustrialMicrobiologyandBiotechnologyꎬ2001ꎬ27(3):149-156.[84]WangJꎬHuWꎬLuxRꎬetal.NaturaltransformationofMyxococcusxanthus[J].JournalofBacteriologyꎬ2011ꎬ193(9):2122-2132.责任编辑:刘怡辰。
南京农业大学研究生学位论文答辩及-南京农业大学园艺学院
南京农业大学研究生学位论文答辩及学位申请工作暂行规定(二OO四年一月六日校学位评定委员会七届六次全会审议通过)根据《中华人民共和国学位条例》和《中华人民共和国学位条例暂行实施办法》,结合我校学位工作具体情况,特制定研究生学位论文答辩及学位申请工作暂行规定如下。
一、总则硕士研究生学位论文答辩及学位申请工作包括答辩资格的申请、论文评阅、答辩及学位授予等环节。
硕士学位论文的答辩及学位申请工作由各学院组织实施。
博士研究生学位论文答辩及学位申请工作包括预答辩资格的申请、预答辩、答辩资格的申请、论文评阅、答辩、学位授予及学位公示等环节。
凡向我校申请博士学位者,一律实行学位论文预答辩制度和论文“双盲”评阅制度。
博士学位论文的预答辩工作由各学院组织实施;博士学位论文的答辩及学位申请工作由研究生院学位办组织实施。
二、硕士学位论文答辩及学位申请(一)答辩申请资格硕士研究生须按计划修满本专业培养方案规定的全部课程,成绩合格,达到所规定的总学分,并参加一定时数的教学实习和社会实践,经研究生管理部门审核认可;完成了硕士学位论文的研究和撰写工作,经导师和学科点点长同意签字,方可申请硕士学位论文答辩。
(二)答辩的组织与表格领取硕士学位论文答辩由所在学院组织。
硕士学位论文答辩申请人凭学位论文答辩稿一份,向所在学院提出申请,资格审查通过后,领取《学位论文答辩申请表》(一份)、《毕业研究生登记表》(一式二份)、《硕士学位申请书》(一式三份)、《知识产权认定书》(一式三份)、《学位论文修改鉴定意见存查表》(一份)、《论文发表情况表》(一份)。
由答辩秘书认真填写《学位论文答辩申请表》,经所在学院分管领导签署意见并加盖公章后,送交所在学院研究生管理部门办理答辩手续。
(三)论文评阅人与答辩委员会硕士学位论文评阅人与答辩委员会成员由本学科点点长与导师协商提名,报学院分管领导审批同意并签名,方可正式聘请,并由各学院组织实施。
被江苏省学位委员会办公室抽检的学位论文评阅按江苏省教育评估院制订的办法(江苏省教育厅文件苏教研[2002]1号)实行。
研究生教学改革类成果

研究生教学改革类成果Title: Graduate Teaching Reform Achievements研究生教学改革类成果是指在高等教育阶段,对研究生教育教学模式、内容、方法等方面进行的创新与改进,以提高研究生培养质量和综合素质。
近年来,我国在研究生教育领域不断进行教学改革,取得了一系列显著成果。
Graduate teaching reform achievements refer to the innovations and improvements in the educational teaching models, content, and methods for graduate students at the higher education stage, in order to enhance the quality and comprehensive capabilities of graduate education.In recent years, China has continuously carried out teaching reforms in the field of graduate education, achieving a series of significant results.一方面,我国加大了研究生课程建设的力度,推动课程内容与前沿科技、产业发展相结合。
例如,通过引入实战项目、企业导师制等多元化教学方式,提高学生的实践能力和创新能力。
另一方面,我国注重研究生教育教学方法的创新,推广案例教学、讨论式教学、翻转课堂等教学模式,激发学生的学习兴趣和主动性。
On the one hand, China has increased its efforts in graduate curriculum construction, promoting the combination of curriculum content with cutting-edge technology, industrial development, and practical projects.For example, by introducing diversified teaching methods such as practical projects and corporate mentorship, thepractical and innovative capabilities of students are improved.On the other hand, China emphasizes the innovation of graduate teaching methods, promoting teaching models such as case-based teaching, discussion-based teaching, and flipped classrooms, to stimulate students" learning interests and initiative.此外,我国还加强了研究生教育质量管理,建立健全质量保障体系。
博士研究生学术英语写作教学“合作完成任务法”的理论与实践

进行外 语教学 ” 式[ “ 模 、 任务教学法 ” 和 “ 合作学 习 法” …的综合 运用 , 三者 的有益 成分合 理地运 用于博 将
士研究生学术论文英文写作 教学过 程。 ( ) 一 依托课程 内容进行 外语教 学 依 托 课 程 内 容 进 行 外 语 教 学 ( o t tbsd C n n ae e— is ut n , nt co ) 简称 C I是上世 纪七八十年代兴起 于加拿 r i B, 大 、 国, 美 随后被其 它国家广泛应用的外语教学模式[。 5 该 ]
教 学 模 式 将 某 个 学 科 的 内 容 教 学 ( uj tm t r Sbe — a e c t cnet 与外语教学相结合 , ot ) n 是一种双语教学 的路子[7 6] -。
人们根 据各 自的认 识和不 同的教学 目的 , 丰富 和发展 了
CI B 教学模式 , 使之成 为从 以内容 为重点 到以语言 为重
学过程 中 , 笔者边学 习有关 教学 理论 , 边进行 教学实践 ,
只是学 习学科 内容的媒介 。 这种 教学要 求语言教师从一
个全新 的角度审视 自己的教学 , 能够通过某 一学科 内容 的教学带动外语教学 , 他最好 是一个 d ul ep  ̄ obe xe 。
( ) 二 任务教 学法
维普资讯
博 士 研 究 生 学 术 英 语 写 作教 学
ቤተ መጻሕፍቲ ባይዱ
“ 作 完成 任 务 法"的理论 与实践 合
顾 飞 荣 , 桂 珍 , 圣 兵 施 高
( 京农 业大 学 , 苏 南 京 2 09 南 江 10 5)
[ 摘 要] 将依 托课程 内容进行 外语教 学的模 式 、 任务语 言教 学 法和合作 学 习法综合 运用于博 士研 究生学
乙烯利对大豆芽菜生长代谢和酚类物质合成的影响

㊀南京农业大学学报㊀2021ꎬ44(2):359-365http://nauxb.njau.edu.cn㊀JournalofNanjingAgriculturalUniversityDOI:10.7685/jnau.202006028收稿日期:2020-06-22基金项目:国家级大学生创新训练项目(202010307042Z)作者简介:王冕ꎬ硕士研究生ꎮ∗通信作者:杨润强ꎬ副教授ꎬ博士ꎬ研究方向为农产品贮藏与加工ꎬE ̄mail:yangrq@njau.edu.cnꎮ王冕ꎬ丁羽萱ꎬ王尧ꎬ等.乙烯利对大豆芽菜生长代谢和酚类物质合成的影响[J].南京农业大学学报ꎬ2021ꎬ44(2):359-365.WANGMianꎬDINGYuxuanꎬWANGYaoꎬetal.Effectofethephonongrowthandphenoliccompoundssynthesisofsoybeansprouts[J].JournalofNanjingAgriculturalUniversityꎬ2021ꎬ44(2):359-365.乙烯利对大豆芽菜生长代谢和酚类物质合成的影响王冕ꎬ丁羽萱ꎬ王尧ꎬ王沛ꎬ顾振新ꎬ杨润强∗(南京农业大学食品科学技术学院ꎬ江苏南京210095)摘要:[目的]本文旨在研究不同浓度乙烯利对大豆芽菜生长代谢和酚类物质合成的影响ꎮ[方法]以大豆品种 云鹤 为材料ꎬ在大豆发芽期间ꎬ用不同浓度乙烯利(乙烯供体)喷淋大豆芽菜ꎬ研究其生理形态和总酚㊁总黄酮㊁异黄酮和酚酸等功能成分以及抗氧化能力的变化ꎮ[结果]乙烯利处理显著影响大豆芽菜的生长发育ꎬ尤其是0.05mmol L-1乙烯利处理时大豆芽菜的鲜/干重比㊁呼吸速率分别比对照增加18.25%㊁43.57%ꎮ随乙烯利浓度的增加ꎬ大豆芽菜的酚类物质含量和抗氧化能力呈先升高后降低的趋势ꎮ当乙烯利浓度为0.05mmol L-1时ꎬ大豆芽菜的总酚和总黄酮含量分别比对照提高7.26%和30.43%ꎻ大豆芽菜的总异黄酮含量达最大值ꎬ其中丙二酰基黄豆苷含量比对照增加30.11%ꎮ[结论]乙烯利不仅影响大豆芽菜的生长代谢ꎬ而且参与大豆芽菜中酚类物质的合成调控ꎬ对提高营养和功能品质具有重要作用ꎮ关键词:大豆芽菜ꎻ乙烯利ꎻ酚类物质ꎻ生物合成中图分类号:TS213.3㊀㊀㊀㊀文献标志码:A㊀㊀㊀㊀文章编号:1000-2030(2021)02-0359-07EffectofethephonongrowthandphenoliccompoundssynthesisofsoybeansproutsWANGMianꎬDINGYuxuanꎬWANGYaoꎬWANGPeiꎬGUZhenxinꎬYANGRunqiang∗(CollegeofFoodScienceandTechnologyꎬNanjingAgriculturalUniversityꎬNanjing210095ꎬChina)Abstract:[Objectives]Theaimofthisarticlewastoinvestigatetheeffectsofdifferentconcentrationsofethephononthegrowthmeta ̄bolismandphenolicssynthesisofsoybeansprouts.[Methods]Duringsoybeanvariety Yunhe germinationꎬthephysiologicalmorphologyandaccumulationoffunctionalcomponentsincludingtotalphynolicsꎬflavonoidsꎬisoflavonesꎬphenolicacidsꎬantioxidantcapacityꎬetc.weremeasuredunderdifferentconcentrationsofethephon(ethylenedonor).[Results]Itshowedthatethephonsignificantlyaffectedthephysiologicalindexesofsoybeansproutsꎬespeciallytheratiooffreshtodryweightandrespirationratewhichincreasedby18.25%and43.57%under0.05mmol L-1ethephontreatmentcomparedwiththecontrol.Phenolicscontentandantioxidantcapacityincreasedfirstlyandthendecreasedwiththeincreaseofethephonconcentration.Totalphenolicsandflavonoidscontentsofsoybeansproutsincreasedby7.26%and30.43%under0.05mmol L-1ofethephonꎬrespectivelyꎬcomparedwiththecontrol.Totalisoflavonecontentofsoybeansproutsreachedthemaximumunder0.05mmol L-1ofethephonꎬwhilemalonyldaidzincontentincreasedby30.11%comparedwiththecontrol.[Conclusions]Ethephonsignificantlyaffectedthegrowthandmetabolismofsoybeansproutsꎬparticipatingintheregu ̄lationofphenolicssynthesisꎬandplayinganimportantroleinimprovingnutritionandfunctionalquality.Keywords:soybeansproutsꎻethephonꎻphenolicsꎻbiosynthesis酚类物质是植物主要的次生代谢产物ꎬ广泛分布于水果㊁蔬菜㊁谷物㊁豆类中ꎬ其具有抗氧化㊁抗肿瘤㊁降血糖㊁预防心血管疾病等多种保健功能ꎮ目前ꎬ酚类物质在植物体中的代谢和积累已有大量研究ꎬ例如:Tian等[1]发现发芽糙米中游离酚类如阿魏酸㊁香豆酸和芥子酸均明显高于未发芽糙米ꎬ且不溶性酚类含量增加ꎻXu等[2]研究发现燕麦发芽后香豆酸㊁阿魏酸和总酚含量显著增加ꎮ此外ꎬ在籽粒萌发阶段逆境胁迫可有效提高酚类物质含量ꎬ例如生菜在100mmol L-1NaCl处理12d后ꎬ总酚和类黄酮含量分别增加1.3倍和14.5倍[3]ꎻ适度的盐胁迫处理时ꎬ烤烟幼苗中绿原酸㊁类黄酮㊁花青苷等酚类物质含量显著增加[4]ꎻWang等[5]研究发现ꎬ亚麻籽萌发第8天时植物化学成分显著富集ꎬ其中维生素C含量增加22.1倍ꎬ酚类物质含量增加2.67倍ꎬ类黄酮含量增加5.48倍ꎮ乙烯(ETH)参与植物次生代谢过程ꎬ可促进生菜㊁胡萝卜㊁绿豆中总酚酸的积累[6]ꎮ曲文颖等[7]研究063南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷发现ꎬ乙烯利可有效促进蓝莓果实中黄酮类物质㊁酚类物质的积累ꎬ提高超氧化物歧化酶(SOD)活性ꎮ刘红开[8]研究发现ꎬ乙烯处理绿豆芽ꎬ不仅促进绿豆芽下胚轴的横向生长ꎬ改善其外观品质及营养价值ꎬ亦可增强酶促氧化和非酶氧化系统ꎬ提高SOD㊁过氧化物酶(POD)㊁过氧化氢酶(CAT)等抗氧化酶活性ꎮ此外ꎬ发芽过程中葡萄糖-6-磷酸脱氢酶(G6PDH)和苯丙氨酸氨裂解酶(PAL)活性的增加促进酚类物质的合成[8]ꎮ由此可见ꎬ乙烯不仅可有效促进植物芽苗生长发育ꎬ而且作为一种信号分子ꎬ促进酚类物质的积累及其抗氧化能力的提高ꎮ大豆(GlycinemaxL.)籽粒富含酚类物质ꎬ发芽过程中会经历复杂的生理生化变化ꎬ使生理活性成分不断积累ꎬ营养价值进一步提高ꎮ其中黄酮类物质是天然的抗氧化剂ꎬ主要分布于大豆种子和胚轴部位ꎬ并随着发芽过程含量不断增加ꎻ此外ꎬ异黄酮㊁β-葡萄糖苷㊁γ-氨基丁酸(GABA)[9]等功能性成分较发芽前大量累积ꎬ抗氧化能力进一步提高[10]ꎮ研究发现大豆在发芽过程中酚类物质会发生不同程度的变化ꎬ营养成分也会显著提高[11]ꎮ目前ꎬ乙烯对大豆发芽过程中酚类物质的富集过程及其变化情况的研究较少ꎬ相关机制还需深入研究ꎮ因此ꎬ本试验采用不同浓度乙烯利处理大豆芽菜ꎬ分析其总酚㊁总黄酮㊁酚酸㊁异黄酮等物质的含量ꎬ筛选适宜的乙烯利浓度ꎬ旨在为高品质大豆芽菜生产提供理论依据ꎮ1㊀材料与方法1.1㊀试验材料及处理供试大豆品种 云鹤 ꎬ收获后置于-20ħ保存备用ꎮ乙烯利(有效成分含量ȡ90%)购自北京索莱宝科技有限公司ꎮ称取适量颗粒饱满的大豆种子ꎬ先用去离子水洗去浮沉与污渍后ꎬ置于1%(体积分数)次氯酸钠水溶液中浸泡消毒15minꎬ然后再用去离子水冲洗3~5次至pH呈中性ꎬ最后加入去离子水于30ħ下浸泡6hꎮ将浸泡后的大豆籽粒均匀铺在发芽机(梦飞扬科技有限公司)苗盘上ꎬ在30ħ黑暗条件下恒温发芽ꎬ分别用浓度为0.025㊁0.050㊁0.100和0.500mmol L-1的乙烯利培养液处理大豆ꎬ以去离子水为对照ꎬ每24h换培养液1次ꎬ每次2Lꎮ取处理4d时的大豆芽菜进行各项指标分析ꎮ1.2㊀指标测定芽长:随机选取25株大豆芽菜ꎬ游标卡尺测定其芽长并取平均值ꎮ鲜/干重比:随机选取大豆芽菜50株ꎬ称鲜重ꎬ然后经真空冷冻干燥后称干重ꎮ重复3次ꎬ计算鲜/干重比ꎮ呼吸速率:采用小篮子法[12]测定呼吸速率ꎮ酚类物质提取:采用Ma等[13]的方法ꎮ取发芽4d大豆芽菜ꎬ经真空冷冻干燥ꎬ粉碎后过孔径380μm的筛得到豆粉ꎬ提取游离酚和结合酚ꎮ总酚含量:采用Ma等[13]的方法测定大豆芽菜的总酚含量ꎮ总酚含量以没食子酸(GAE)当量计ꎬ单位为mg 100g-1ꎮ结果以干重计ꎮ酚酸含量:采用Chen等[14]的方法测定大豆芽菜的酚酸含量ꎮ结果以干重计ꎮ总黄酮含量:采用Islam等[15]的方法ꎮ称取0.2g豆粉ꎬ测定大豆芽菜的总黄酮含量ꎮ总黄酮含量以芦丁(RE)当量计ꎬ单位为mg 100g-1ꎮ结果以干重计ꎮ异黄酮含量:采用Jiao等[16]的方法测定大豆芽菜的异黄酮含量ꎮ结果以干重计ꎮ2ꎬ2ᶄ-联氮-双-3-乙基苯并噻唑啉-6-磺酸)(ABTS)自由基清除能力的测定:参考Islam等[17]的方法ꎮ将7mmol L-1ABTS与2.45mmol L-1过硫酸钾等体积混合ꎬ室温下避光反应12~16hꎬ用纯甲醇稀释至734nm下吸光度为0.7左右ꎬ即A液ꎮ取100μL适当稀释的样品提取液ꎬ加入3mLA液于室温下避光反应30minꎬ测定734nm下吸光值ꎮ以Trolox绘制标准曲线ꎬ蒸馏水为对照ꎮABTS自由基清除能力以干重计ꎮ1ꎬ1-二苯基-2-三硝基苯肼(DPPH)自由基清除能力的测定:参考He等[18]的方法ꎮ称取11.8mgDPPHꎬ纯甲醇定容至500mLꎬ即60μmol L-1B液ꎮ取200μL适当稀释的样品提取液ꎬ加入3.8mLB液ꎬ室温下避光反应30minꎬ于515nm处测定吸光值ꎮ以Trolox绘制标准曲线ꎬ蒸馏水为对照ꎮDPPH自由基清除能力以干重计ꎮ㊀第2期王冕ꎬ等:乙烯利对大豆芽菜生长代谢和酚类物质合成的影响1.3㊀数据统计与分析采用SPSS23.0软件对数据进行方差分析㊁邓肯氏多重比较和相关性分析ꎬ显著性水平为P<0.05ꎮ试验设3次生物学重复ꎬ数据以 平均值ʃ标准差 表示ꎮ2㊀结果与分析2.1㊀不同乙烯利浓度对大豆芽菜生理代谢的影响由图1可知:不同乙烯利浓度处理下大豆芽菜生长形态存在差异ꎮ对照大豆芽菜芽长最长ꎬ乙烯利处理后大豆芽菜生长受到抑制ꎬ且随乙烯利浓度的增加ꎬ大豆芽菜下胚轴的纵向生长抑制程度增加ꎬ但对横向生长先促进后抑制ꎮ当乙烯利浓度为0.050mmol L-1时ꎬ侧根生长最为旺盛ꎮ此后继续增加乙烯利浓度时ꎬ大豆芽菜整体生长逐渐受到显著抑制ꎮ随着乙烯利浓度增加ꎬ大豆芽菜鲜/干重比呈先增加后降低的趋势ꎬ且当乙烯利浓度为0.050mmol L-1时ꎬ其鲜/干重比达到最大值ꎬ比对照增加18.25%(P<0.05)ꎬ说明适宜的乙烯利浓度可有效促进大豆芽菜的鲜重累积ꎮ当乙烯利浓度为0.050mmol L-1时大豆芽菜的呼吸速率最高ꎬ比对照增加43.57%ꎮ而乙烯利浓度进一步增加时ꎬ大豆芽菜的呼吸速率与对照相比则无显著性差异ꎬ说明一定浓度的乙烯利可有效提高大豆芽菜的呼吸速率和生长代谢水平ꎮ图1㊀乙烯利对大豆芽菜生长发育的影响Fig 1㊀Effectsofethephononthegrowthanddevelopmentofsoybeansprouts不同小写字母表示不同处理之间存在显著差异(P<0.05)ꎮ下同ꎮDifferentlowercaselettersindicatedsignificantdifferenceat0.05levelamongdifferenttreatments.Thesameasfollows.2.2㊀不同乙烯利浓度下大豆芽菜总酚和总黄酮含量变化由图2可知:大豆芽菜中总酚含量随乙烯利浓度的增加呈先增加后降低的趋势ꎬ当乙烯利浓度为0.050mmol L-1时总酚含量达到最大值ꎬ说明适宜的乙烯利可促进大豆芽菜中总酚的合成ꎬ但浓度过高时总酚合成会受到抑制ꎮ大豆芽菜中总黄酮含量与总酚含量变化基本一致ꎮ当乙烯利浓度为0.025和0.050mmol L-1时ꎬ大豆芽菜中总黄酮显著富集ꎬ比对照分别增加30.37%和30.43%ꎬ说明一定浓度的乙烯利可有效促进大豆芽菜中总黄酮的合成水平ꎮ图2㊀乙烯利对大豆芽菜总酚和总黄酮含量的影响Fig 2㊀Effectofethephononphenolicsandflavonoidscontentsofsoybeansprouts2.3㊀不同乙烯利浓度下大豆芽菜异黄酮含量变化由表1可知:大豆芽菜中总异黄酮含量变化趋势与总黄酮基本一致ꎬ乙烯利浓度为0.050mmol L-1时总异黄酮含量达最大值ꎮ大豆芽菜中异黄酮组分ꎬ如:黄豆苷㊁染料木苷㊁黄豆苷元含量均在0.025mmol L-1乙烯利处理时达到最大值ꎮ丙二酰基黄豆苷含量在乙烯利浓度为0.050mmol L-1时达到最大值ꎬ比对照163南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷增加30.11%ꎮ丙二酰基黄豆黄苷和丙二酰基染料木苷含量随乙烯利浓度增加均有不同程度地下降ꎬ说明一定浓度乙烯利处理可提高异黄酮总量ꎬ但对各组分的调控作用存在差异ꎬ主要表现为促进黄豆苷元㊁丙二酰基黄豆苷等组分的合成ꎮ表1㊀乙烯利处理下大豆芽菜中异黄酮各组分含量Table1㊀Theindividualisoflavonecontentsofsoybeansproutswithdifferentethephontreatments组分Component乙烯利浓度/(mmol L-1)Ethephonconcentration00.0250.0500.1000.500黄豆苷含量/(μg g-1)Daidzincontent㊀508.96ʃ2.69c㊀612.64ʃ41.34a㊀570.55ʃ2.45b㊀459.47ʃ17.98d㊀455.99ʃ22.10d黄豆黄苷含量/(μg g-1)Glycitincontent9.53ʃ0.00a 5.04ʃ0.00b7.92ʃ0.00a染料木苷含量/(μg g-1)Genisteincontent284.20ʃ6.08bc350.40ʃ12.26a304.57ʃ5.60b298.79ʃ10.04b262.56ʃ13.85c丙二酰基黄豆苷含量/(μg g-1)Malonyldaidzincontent2929.26ʃ20.52c3548.91ʃ20.18b3811.14ʃ50.87a2723.71ʃ10.64d2954.50ʃ41.86c丙二酰基黄豆黄苷含量/(μg g-1)Malonylglycitincontent309.70ʃ14.26a289.76ʃ0.16ab280.99ʃ1.11b231.81ʃ2.90c251.89ʃ11.64c丙二酰基染料木苷含量/(μg g-1)Malonylgenistincontent1717.24ʃ47.19a1514.54ʃ13.08b1458.52ʃ37.45b1395.79ʃ23.57b1190.58ʃ78.16c黄豆苷元含量/(μg g-1)Daidzeincontent40.80ʃ0.13c159.67ʃ3.62a55.01ʃ2.91b36.14ʃ1.35cd32.02ʃ0.02d总异黄酮含量/(μg g-1)Totalisoflavonescontent5794.92ʃ97.36b6475.92ʃ90.63a6480.76ʃ87.45a5150.74ʃ52.32c5155.45ʃ112.20c2.4㊀不同乙烯利浓度下大豆芽菜酚酸组成成分的变化由表2可见:大豆芽菜中总酚酸含量在乙烯利浓度为0.025mmol L-1时达到最大值ꎬ比对照增加34.79%ꎮ当用0.050mmol L-1乙烯利处理后ꎬ大豆芽菜中结合酚酸和总酚酸比对照分别增加47.43%和16.48%ꎮ乙烯利浓度为0.500mmol L-1时ꎬ大豆芽菜中酚酸总量显著低于对照ꎬ说明适宜乙烯利浓度可有效促进大豆芽菜中酚酸的富集ꎬ过高的乙烯利浓度对酚酸的合成会产生抑制作用ꎮ乙烯利处理后大豆芽菜酚酸各组分含量变化趋势不同ꎮ大豆芽菜中没食子酸含量在乙烯利浓度为0.050mmol L-1时达到最大值ꎻ原儿茶酸在乙烯利浓度为0.025mmol L-1时显著增加并达到最大值ꎬ比对照增加5.86%ꎮ当乙烯利浓度大于0.025mmol L-1时ꎬ原儿茶酸的合成受到抑制ꎬ含量显著下降ꎮ在乙烯利浓度为0.050mmol L-1时ꎬ大豆芽菜中香草酸㊁对香豆酸㊁阿魏酸㊁芥子酸含量均显著增加ꎬ分别比对照增加1.17㊁0.49㊁0.68㊁2.88倍ꎮ大豆芽菜中对羟基苯甲酸含量极低ꎬ且丁香酸主要以游离态存在ꎮ表2㊀不同乙烯利浓度下大豆芽菜中酚酸含量的变化Table2㊀Theindividualphenolicacidofsoybeansproutswithdifferentethephonconcentrations组分Component乙烯利浓度/(mmol L-1)Ethephonconcentration酚酸含量/(μg g-1)Phenolicacidcontent游离态Free结合态Bound总计Total没食子酸Gallicacid原儿茶酸Protocatechuicacid对羟基苯甲酸P ̄hydroxybenzoicacid香草酸Vanillicacid丁香酸Syringicacid0 0.25ʃ0.00c0.25ʃ0.00d0.02527.57ʃ0.15b0.47ʃ0.02b28.04ʃ0.16b0.05032.55ʃ0.00a0.68ʃ0.09a33.23ʃ0.09a0.10014.32ʃ0.40c0.34ʃ0.02c14.66ʃ0.38c0.5000.34ʃ0.00c0.34ʃ0.00d0128.78ʃ21.07ab0.85ʃ0.05d129.63ʃ21.13ab0.025134.71ʃ5.29a2.52ʃ0.03a137.23ʃ5.26a0.05080.62ʃ2.79cd1.94ʃ0.00b82.56ʃ2.79c0.10075.86ʃ9.73d1.00ʃ0.01c76.87ʃ9.75c0.500104.61ʃ2.86bc0.44ʃ0.08e105.05ʃ2.94bc0 2.34ʃ0.14b2.34ʃ0.14b0.025 3.70ʃ0.40a3.70ʃ0.40a0.050 4.05ʃ0.10a4.05ʃ0.10a0.100 1.77ʃ0.14c1.77ʃ0.14c0.5002.41ʃ0.02b2.41ʃ0.02b0 23.96ʃ0.17e23.96ʃ0.17d0.0258.03ʃ0.08a42.17ʃ0.20b50.20ʃ0.12a0.0505.72ʃ1.45b46.20ʃ0.93a51.91ʃ2.38a0.100 26.61ʃ1.91d26.61ʃ1.91c0.5004.54ʃ0.15b33.58ʃ0.11c38.13ʃ0.04b011.90ʃ0.80a 11.90ʃ0.80a0.02512.13ʃ0.70a 12.13ʃ0.70a0.05010.17ʃ0.01b 10.17ʃ0.01b0.1007.95ʃ0.00c 7.95ʃ0.00c0.500 263㊀第2期王冕ꎬ等:乙烯利对大豆芽菜生长代谢和酚类物质合成的影响续表2㊀Table2continued组分Component乙烯利浓度/(mmol L-1)Ethephonconcentration酚酸含量/(μg g-1)Phenolicacidcontent游离态Free结合态Bound总计Total对香豆酸P ̄coumaricacid阿魏酸Ferulicacid芥子酸Erucicacid总酚酸Totalphenolicacids0 45.63ʃ0.59c45.63ʃ0.59c0.025 54.33ʃ0.17b54.33ʃ0.17b0.0505.41ʃ1.64b62.41ʃ2.22a67.82ʃ3.86a0.1007.30ʃ1.31b33.91ʃ0.48d41.21ʃ0.82c0.50011.16ʃ0.06a31.53ʃ0.07d42.70ʃ0.13c0 8.58ʃ0.00c8.58ʃ0.00d0.0250.16ʃ0.00c13.56ʃ0.00a13.73ʃ0.01b0.0500.75ʃ0.00a13.68ʃ0.19a14.44ʃ0.19a0.1000.28ʃ0.00b8.16ʃ0.48c8.44ʃ0.48d0.50011.19ʃ0.04b11.19ʃ0.04c0 4.74ʃ0.16c4.74ʃ0.16d0.0257.38ʃ0.08d9.14ʃ0.01a16.52ʃ0.09b0.0509.23ʃ0.00c9.18ʃ0.00a18.41ʃ0.00a0.10010.45ʃ0.76b4.48ʃ0.20c14.92ʃ0.96c0.50012.59ʃ0.00a5.81ʃ0.15b18.40ʃ0.15a0140.68ʃ21.87b62.37ʃ0.94c203.05ʃ22.81c0.025189.97ʃ6.31a83.73ʃ0.58b273.70ʃ5.73a0.050144.56ʃ6.02b91.95ʃ2.41a236.51ʃ8.43b0.100114.61ʃ12.58b49.66ʃ0.93d164.28ʃ13.51d0.500132.90ʃ2.77b51.73ʃ0.32d184.63ʃ3.09cd2.5㊀不同乙烯利浓度下大豆芽菜抗氧化能力的变化由图3可知:当乙烯利浓度为0.025和0.050mmol L-1时ꎬ大豆芽菜中ABTS和DPPH自由基清除能力均比对照显著提高ꎻ而乙烯利浓度为0.500mmol L-1时ꎬ大豆芽菜的ABTS自由基清除能力受到抑制ꎬ说明一定浓度乙烯利可以有效提升大豆芽菜的抗氧化能力ꎬ但浓度过高时会影响大豆芽菜的生长发育ꎬ酚类物质积累水平受到影响ꎬ从而降低自身抗氧化能力ꎮ图3㊀乙烯利对大豆芽菜抗氧化能力的影响Fig 3㊀Effectofethephonontheantioxidantabilityofsoybeansprouts㊀㊀ABTS:2ꎬ2ᶄ-联氮-双-3-乙基苯并噻唑啉-6-磺酸2ꎬ2ᶄ ̄azino ̄bis(3 ̄ethylbenzothiazoline ̄6 ̄sulfonicacidꎻDPPH:1ꎬ1-二苯基-2-三硝基苯肼1ꎬ1 ̄diphenyl ̄2 ̄picrylhydrazyl.经0.050mmol L-1乙烯利处理的大豆芽菜ꎬ其游离酚㊁结合酚的ABTS㊁DPPH自由基清除能力均达到最大值ꎬ其中大豆芽菜中游离酚和结合酚的ABTS自由基清除能力分别是对照的1.1和1.3倍ꎬDPPH自由基清除能力分别是对照的1.04和1.3倍ꎮ大豆芽菜中总酚主要以游离酚形式存在ꎬ经0.05mmol L-1乙烯利处理后ꎬ其游离酚的ABTS㊁DPPH自由基清除能力显著增加ꎬ是影响总酚抗氧化能力的重要因素ꎮ363463南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第44卷3 讨论研究表明ꎬ大豆芽菜在发芽形成芽苗过程中ꎬ不仅保持部分原有营养成分ꎬ而且膳食纤维㊁γ-氨基丁酸(GABA)和酚类等功能成分大量累积ꎬ其营养和功能品质均得到提高[19]ꎮ本研究发现ꎬ不同浓度乙烯利处理对大豆芽菜的芽长㊁鲜/干重比㊁呼吸速率等生理指标均有显著影响ꎬ且适宜浓度的乙烯利可增强大豆芽菜中细胞的生长代谢水平ꎬ改变外观及营养品质ꎮ刘红开[8]研究表明乙烯利处理绿豆芽可促进绿豆芽下胚轴的横向生长以及细胞壁的合成代谢ꎬ从而增加其体积和鲜重ꎬ改善外观品质及营养价值ꎮ此外ꎬ施加外源乙烯利可促进植物内源乙烯的产生ꎬ增加植物的呼吸强度[20]ꎮJiao等[16]研究发现ꎬ一定浓度的乙烯利会引起蓝莓内源乙烯的积累ꎬ提高蓝莓呼吸代谢速率ꎬ本研究结果与之相一致ꎮ本研究表明ꎬ乙烯利不仅改善大豆芽菜的外观品质ꎬ对其内部酚类物质的合成也有显著影响ꎬ其中在乙烯利浓度为0.050mmol L-1时ꎬ大豆芽菜的总酚含量比对照增加7.26%ꎬ尤其结合酚比对照增加40%ꎻ总黄酮㊁异黄酮比对照分别增加30.43%㊁11.84%ꎻ酚酸中没食子酸含量变化较大ꎬ比对照增加131.92倍ꎮ乙烯利(0.05mmol L-1)可调控大豆芽菜中酚类物质合成关键酶的活性ꎬ如PAL㊁POD㊁肉桂醇脱氢酶(CAD)等ꎬ促进酚类物质的富集ꎬ显著提高ABTS㊁DPPH自由基清除能力ꎬ增强植物自身抗氧化能力[21]ꎮ刘红开[8]研究发现ꎬPAL活性与总酚含量成正相关ꎬ绿豆芽经乙烯利处理后PAL活性显著提高ꎬ总酚含量显著增加ꎮGuo等[22]研究表明ꎬ大豆发芽5d后ꎬPAL基因表达水平显著提高ꎬ内源总酚和总黄酮大量富集ꎮTian等[23]发现ꎬ发芽能够提高燕麦酚类物质的含量ꎬ归因于谷物发芽时酶活性的提高ꎮ荞麦萌发过程中可生成酚酸㊁黄酮等重要的植物次生代谢产物ꎬ主要通过苯丙烷代谢途径合成ꎬ其中PAL是重要限速酶[24]ꎮ另有研究认为ꎬPAL酶异构体亚细胞定位的特异性可能和该酶的代谢活性调节密切相关[25]ꎮ因此ꎬPAL作为苯丙烷代谢过程的关键限速酶ꎬ在酚类物质合成途径中具有重要作用ꎮ乙烯利可通过提高大豆芽菜中苯丙烷代谢途径的关键酶活性ꎬ促进酚类物质的富集以及自身抗氧化能力的提高ꎮ本研究表明ꎬ乙烯利可促进大豆芽菜中总酚㊁总黄酮及异黄酮的合成ꎬ其中黄豆苷㊁染料木苷㊁丙二酰基黄豆苷㊁黄豆苷元等异黄酮单一组分均比对照显著增加ꎬ但丙二酰基黄豆黄苷和丙二酰基染料木苷的含量有所降低ꎬ说明乙烯利对大豆芽菜中异黄酮单一组分的富集作用存在差异ꎮ综上ꎬ浓度为0.050mmol L-1的乙烯利可促进大豆芽菜下胚轴的横向生长ꎬ改善大豆芽菜的外观品质ꎬ同时促进大豆芽菜中酚酸㊁黄酮等功能成分的累积ꎬ有效提高大豆芽菜的营养品质ꎮ乙烯利浓度过高则会对大豆芽菜的生理代谢和酚类物质合成产生抑制ꎮ乙烯利是否通过影响苯丙烷代谢途径中关键酶的表达调控酚类物质的合成有待研究ꎮ参考文献References:[1]㊀TianSꎬNakamuraKꎬKayaharaH.Analysisofphenoliccompoundsinwhitericeꎬbrownriceꎬandgerminatedbrownrice[J].JournalofAgriculturalandFoodChemistryꎬ2004ꎬ52(15):4808-4813.[2]XuJGꎬTianCRꎬHuQPꎬetal.Dynamicchangesinphenoliccompoundsandantioxidantactivityinoats(AvenanudaL.)duringsteepingandgermination[J].JournalofAgriculturalandFoodChemistryꎬ2009ꎬ57(21):10392-10398.[3]MahmoudiHꎬHuangJꎬGruberMYꎬetal.Theimpactofgenotypeandsalinityonphysiologicalfunctionꎬsecondarymetaboliteaccumulationꎬandantioxidativeresponsesinlettuce[J].JournalofAgriculturalandFoodChemistryꎬ2010ꎬ58(8):5122-5130.[4]胡庆辉.盐与干旱胁迫诱导烤烟叶片细胞程序性死亡及多酚含量变化的研究[D].北京:中国农业科学院ꎬ2012.HuQH.Thestudyofprogrammedcelldeathandpolyphenolscontentschangesofflue ̄curedtobaccoleavesinducedbysaltanddroughtstress[D].Beijing:ChineseAcademyofAgriculturalSciencesꎬ2012(inChinesewithEnglishabstract).[5]WangHꎬQiuCSꎬAbbasiAMꎬetal.EffectofgerminationonvitaminCꎬphenoliccompoundsandantioxidantactivityinflaxseed(LinumusitatissimumL.)[J].InternationalJournalofFoodScience&Technologyꎬ2015ꎬ50(12):2545-2553.[6]HerediaJBꎬCisneros ̄ZevallosL.Theeffectsofexogenousethyleneandmethyljasmonateontheaccumulationofphenolicantioxidantsinselectedwholeandwoundedfreshproduce[J].FoodChemistryꎬ2009ꎬ115(4):1500-1508.[7]曲文颖ꎬ刘真真ꎬ谢琳淼ꎬ等.外源脱落酸和乙烯利对蓝莓重要品质的调控[J].江苏农业科学ꎬ2017ꎬ45(17):126-129.QuWYꎬLiuZZꎬXieLMꎬetal.Regulationofexogenousabscisicacidandethephononimportantqualityofblueberries[J].JiangsuAgriculturalSciencesꎬ2017ꎬ45(17):126-129(inChinese).[8]刘红开.乙烯对绿豆芽下胚轴生长代谢的调控效应研究[D].北京:中国农业大学ꎬ2014.LiuHK.Reasearchofethyleneinregulatinggrowthandmetabolismofmungbean[Vignaradiata(L.)Wilczek]sproutshypocotyl[D].Beijing:ChinaAgriculturalUniversityꎬ2014(inChinesewithEnglishabstract).[9]VannKꎬTechaparinAꎬApiraksakornJ.BeansgerminationasapotentialtoolforGABA ̄enrichedtofuproduction[J].JournalofFoodScience563㊀第2期王冕ꎬ等:乙烯利对大豆芽菜生长代谢和酚类物质合成的影响andTechnologyꎬ2020ꎬ57(11):3947-3954.[10]KimMAꎬKimMJ.Isoflavoneprofilesandantioxidantpropertiesindifferentpartsofsoybeansprout[J].JournalofFoodScienceꎬ2020ꎬ85(3):689-695.[11]EumHLꎬParkYꎬYiTGꎬetal.Effectofgerminationenvironmentonthebiochemicalcompoundsandanti ̄inflammatorypropertiesofsoybeancultivars[J].PLoSOneꎬ2020ꎬ15(4):e0232159.[12]王学奎.植物生理生化实验原理和技术[M].2版.北京:高等教育出版社ꎬ2006.WangXK.PrinciplesandTechniquesofPlantPhysiologicalBiochemicalExperiment[M].2nded.Beijing:HigherEducationPressꎬ2006(inChinese).[13]MaYꎬWangPꎬZhouTꎬetal.RoleofCa2+inphenoliccompoundmetabolismofbarley(HordeumvulgareL.)sproutsunderNaClstress[J].JournaloftheScienceofFoodandAgricultureꎬ2019ꎬ99(11):5176-5186.[14]ChenZJꎬMaYꎬWengYꎬetal.EffectsofUV ̄Bradiationonphenolicaccumulationꎬantioxidantactivityandphysiologicalchangesinwheat(TriticumaestivumL.)seedlings[J].FoodBioscienceꎬ2019ꎬ30:100409.[15]IslamMZꎬParkBJꎬLeeYT.Effectofsalinitystressonbioactivecompoundsandantioxidantactivityofwheatmicrogreenextractunderorganiccultivationconditions[J].InternationalJournalofBiologicalMacromoleculesꎬ2019ꎬ140:631-636.[16]JiaoCFꎬYangRQꎬZhouYLꎬetal.Nitricoxidemediatesisoflavoneaccumulationandtheantioxidantsystemenhancementinsoybeansprouts[J].FoodChemistryꎬ2016ꎬ204:373-380.[17]IslamMZꎬParkBJꎬKangHMꎬetal.Influenceofseleniumbiofortificationonthebioactivecompoundsandantioxidantactivityofwheatmicrogreenextract[J].FoodChemistryꎬ2020ꎬ309:125763.[18]HeWWꎬWangYXꎬLuoHꎬetal.EffectofNaClstressandsupplementalCaCl2oncarotenoidaccumulationingerminatedyellowmaizekernels[J].FoodChemistryꎬ2020ꎬ309:125779.[19]BuenoDBꎬdaSilvaSIꎬJrꎬChiarottoSABꎬetal.Thegerminationofsoybeansincreasesthewater ̄solublecomponentsandcouldgenerateinno ̄vationsinsoy ̄basedfoods[J].LWT:FoodScienceandTechnologyꎬ2020ꎬ117:108599.[20]ShaoYFꎬXuFFꎬSunXꎬetal.Phenolicacidsꎬanthocyaninsꎬandantioxidantcapacityinrice(OryzasativaL.)grainsatfourstagesofdevelopmentafterflowering[J].FoodChemistryꎬ2014ꎬ143:90-96.[21]XieGFꎬFengYCꎬChenYꎬetal.Effectsof1 ̄methylcyclopropene(1 ̄MCP)andethyleneonpostharvestlignificationofcommonbeans(PhaseolusvulgarisL.)[J].ACSOmegaꎬ2020ꎬ5(15):8659-8666.[22]GuoXBꎬWangLZꎬChangXXꎬetal.Influenceofplantgrowthregulatorsonkey ̄codinggenesexpressionassociatedwithphytochemicalsbiosynthesisandantioxidantactivityinsoybean[Glycinemax(L.)Merr]sprouts[J].InternationalJournalofFoodScience&Technologyꎬ2019ꎬ54(3):771-779.[23]TianBQꎬXieBJꎬShiJꎬetal.Physicochemicalchangesofoatseedsduringgermination[J].FoodChemistryꎬ2010ꎬ119(3):1195-1200. [24]ZhangGꎬXuZCꎬGaoYYꎬetal.Effectsofgerminationonthenutritionalpropertiesꎬphenolicprofilesꎬandantioxidantactivitiesofbuckwheat[J].JournalofFoodScienceꎬ2015ꎬ80(5):H1111-H1119.[25]BassardJEꎬMuttererJꎬDuvalFꎬetal.AnovelmethodformonitoringthelocalizationofcytochromesP450andotherendoplasmicReticulummembraneassociatedproteins:atoolforinvestigatingtheformationofmetabolons[J].TheFEBSJournalꎬ2012ꎬ279(9):1576-1583.责任编辑:范雪梅。
江苏省普通高校研究生科技创新计划-南京农业大学研究生院
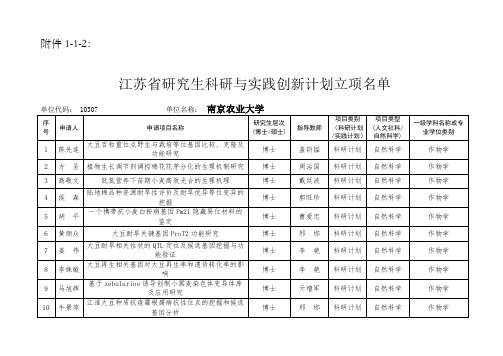
附件1-1-2:江苏省研究生科研与实践创新计划立项名单单位代码: 10307 单位名称:南京农业大学序号申请人申请项目名称研究生层次(博士/硕士)指导教师项目类别(科研计划/实践计划)项目类型(人文社科/自然科学)一级学科名称或专业学位类别1 陈先连大豆百粒重位点野生与栽培等位基因比较、克隆及功能研究博士盖钧镒科研计划自然科学作物学2 方圣植物生长调节剂调控棉花花芽分化的生理机制研究博士周治国科研计划自然科学作物学3 高敬文低氮营养下苗期小麦高效光合的生理机理博士戴廷波科研计划自然科学作物学4 侯森陆地棉品种资源耐旱性评价及耐旱优异等位变异的挖掘博士郭旺珍科研计划自然科学作物学5 胡平一个携带抗小麦白粉病基因Pm21隐藏易位材料的鉴定博士曹爱忠科研计划自然科学作物学6 黄颜众大豆耐旱关键基因ProT2功能研究博士邢邯科研计划自然科学作物学7 姜伟大豆耐旱相关性状的QTL定位及候选基因挖掘与功能验证博士李艳科研计划自然科学作物学8 李姝璇大豆再生相关基因对大豆再生率和遗传转化率的影响博士李艳科研计划自然科学作物学9 马旭辉基于zebularine诱导创制小黑麦染色体变异体库及应用研究博士亓增军科研计划自然科学作物学10 牛景萍江淮大豆种质抗疫霉根腐病抗性位点的挖掘和候选基因分析博士邢邯科研计划自然科学作物学11 潘秀才水稻全基因组DNA双链断裂位点的定位及表观特征研究博士张文利科研计划自然科学作物学12 邵宇航镁与干旱锻炼缓解小麦灌浆期高温胁迫生理机制博士王绍华科研计划自然科学作物学13 汪翔全基因组关联分析水稻低温发芽力博士华健科研计划自然科学作物学14 王佩斯野生二粒小麦抗白粉病基因HSM1的精细定位及克隆博士马正强科研计划自然科学作物学15 王青百萨偃麦草4JL染色体变异体精确鉴定与物理作图博士亓增军科研计划自然科学作物学16 王维领水杨酸在低温锻炼诱导小麦抗寒性中的作用机制研究博士姜东科研计划自然科学作物学17 张贺滨海盐碱地水盐及磷素影响棉花产量的生理生态机制博士孟亚利科研计划自然科学作物学18 张静棉花内生菌中抗菌蛋白抗棉花黄萎病的功能研究博士唐灿明科研计划自然科学作物学19 周雪松与棉花耐逆相关的八氢番茄红素合酶基因的功能分析博士郭旺珍科研计划自然科学作物学20 杨彬水稻种子休眠基因定位与克隆研究博士张红生科研计划自然科学作物学21 胡波甜菜夜蛾谷胱甘肽S—转移酶的代谢功能及调控机制研究博士苏建亚科研计划自然科学植物保护22 丁园氯噻啉纳米多肽竞争物的研制与应用基础研究博士王鸣华科研计划自然科学植物保护23 高原西来稗对二氯喹啉酸抗药性受体机理的研究博士董立尧科研计划自然科学植物保护24 王龙气候变化下转Bt玉米氮素利用与内(外)源防卫物质协调抗虫机理研究博士陈法军科研计划自然科学植物保护25 田盼盼原生共生菌对寄主专化型棉蚜氨基酸需求的调控作用博士刘向东科研计划自然科学植物保护26 盛成旺杀虫剂fluralaner对二化螟的毒力及其作用机制研究博士韩召军科研计划自然科学植物保护27 郝佳Bt Cry 1F毒素抗独特型抗体制备及活性机理研究博士刘贤金科研计划自然科学植物保护28 任维超禾谷镰孢菌细胞自噬参与三唑类杀菌剂的药理学研究博士陈长军科研计划自然科学植物保护29 王帅帅疫霉菌效应蛋白SCR2调节植物免疫反应的机制研究博士郑小波科研计划自然科学植物保护30 张丽娜有益微生物群对辣椒疫病生态防治机理研究博士郭坚华科研计划自然科学植物保护31 孙画婳褐飞虱对醚菊酯抗性的生化与靶标机制博士刘泽文科研计划自然科学植物保护32 赵小珍菠萝泛菌引致玉米褐腐病的致病机制博士高学文科研计划自然科学植物保护33 乔策策生物有机肥提升作物产量的微生物生态机理研究博士李荣科研计划自然科学农业资源与环境34 熊丽气候变化下,根系分泌物对土壤有机质化学结构的影响博士李恋卿科研计划自然科学农业资源与环境35 刘俊丽菌根特异诱导番茄质膜SlHA8的功能和调控机制研究博士陈爱群科研计划自然科学农业资源与环境36 陈旭铜绿假单胞菌(Pseudomonas aeruginosa)持留态形成的分子机制研究博士李辉信科研计划自然科学生态学37 郭瑞抗生素抗性细菌对C.elegans繁殖影响机制研究博士李辉信科研计划自然科学生态学38 王建生物炭中多环芳烃污染特征及还田作物风险研究博士高彦征科研计划自然科学农业工程39 秦超小分子有机污染物对DNA降解的影响及作用机理研究博士高彦征科研计划自然科学农业工程40 王婷婷水稻糖分调控因子OsSPR在氮磷养分平衡机制中的研究博士徐国华科研计划自然科学农业资源与环境41 丁元君分子标志物表征水稻土有机质分子的周转和来源的研究博士潘根兴科研计划自然科学农业资源与环境42 艾昊水稻OsLPR3基因在缺磷胁迫响应中的功能研究博士孙淑斌科研计划自然科学农业资源与环境43 郑海平土壤微生物降解农业秸秆的特征研究博士徐阳春科研计划自然科学农业资源与环境44 刘秀丽水稻磷转运蛋白基因OsPHT2;1的功能研究博士孙淑斌科研计划自然科学农业资源与环境45 吴鹏白菜类作物糖代谢相关基因的系统进化及与耐寒的关系博士侯喜林科研计划自然科学蔬菜学46 高立伟结球白菜叶球形态类型相关基因ARF3.1的克隆和功能验证博士侯喜林科研计划自然科学蔬菜学47 王文丽NFL介导不结球白菜短日照下抽薹开花的分子机理博士侯喜林科研计划自然科学蔬菜学48 袁敬平白菜IQD35对干旱的响应及其调控机制的研究博士李英科研计划自然科学蔬菜学49 张皖皖菊花乙烯信号路径基因CmHRE2响应蚜虫分子机制研究博士陈素梅科研计划自然科学观赏园艺学50 范莲雪萝卜Vc 和SF含量性状形成关键基因鉴定及功能分析博士柳李旺科研计划自然科学蔬菜学51 却枫胡萝卜水培技术体系建立及应用的研究博士熊爱生科研计划自然科学蔬菜学52 刘志薇茶树中氮素对茶氨酸代谢调控的分子机制博士庄静科研计划自然科学茶学53 吕善武不结球白菜中与TuMV-NIb 蛋白相关的寄主因子的筛选及功能验证博士侯喜林科研计划自然科学蔬菜学54 纠松涛葡萄果皮着色相关myb类调节基因的基因型及其调控着色的机理博士房经贵科研计划自然科学果树学55 陆壮高温环境对肉鸡糖脂代谢的影响及其机理研究博士高峰科研计划自然科学畜牧学56 李伯江利用RNA-seq鉴定影响猪肌内脂肪的候选基因博士刘红林科研计划自然科学畜牧学57 薛艳锋妊娠毒血症绵羊肝脏糖脂代谢改变及其调控机制探究博士毛胜勇科研计划自然科学畜牧学58 姚晓磊维生素D参与山羊卵泡发育过程中分子机制研究博士王锋科研计划自然科学畜牧学59 李新宇SIVA1与TGF-β信号通路相互作用调控猪卵巢颗粒细胞凋亡机制博士李齐发科研计划自然科学畜牧学60 夏斯蕾不同淀粉类饲料原料对团头鲂营养代谢、健康和品质的影响博士刘文斌科研计划自然科学畜牧学61 刘余我国粮食经营规模是否适度?——基于江苏省稻农效率的测算博士周应恒科研计划人文社科应用经济学62 赵丹丹生产集聚、农业机械化水平与粮食生产率博士周宏科研计划人文社科农林经济管理63 张骏逸农户秸秆直接还田技术采用行为与政策激励研究博士徐志刚科研计划人文社科农林经济管理64 刘家成中国粮食生产环节外包边界及外包模式研究博士钟甫宁科研计划人文社科农林经济管理65 宁可互联网、生活方式变化与中国农村青少年健康博士钟甫宁、徐志刚科研计划人文社科农林经济管理66 刘畅子女外出务工、老龄化与留守老人健康博士易福金科研计划人文社科农林经济管理67 王莹生产服务可获得性、外包行为与农户生产效率博士陈超科研计划人文社科农林经济管理68 王越人力资本投资与代际收入流动——基于公共投资的视角博士何军科研计划人文社科农林经济管理69 高修歌马度米星铵引起鸡心肌细胞巨泡式死亡的毒性作用及机制研究博士江善祥科研计划自然科学兽医学70 董雨豪应激激素对嗜水气单胞菌生长及致病性调控的研究博士刘永杰科研计划自然科学兽医学71 马可CRISPR-Cas系统对无乳链球菌毒力的影响及调控机制博士刘永杰科研计划自然科学兽医学72 彭婕马链球菌兽疫亚种PurC蛋白影响中性粒细胞跨血脑屏障迁移的机制博士范红结科研计划自然科学兽医学73 袁律峰表面活性素对猪肠道囊膜病毒的抗病毒机理博士杨倩科研计划自然科学兽医学74 张瑜娟鸡BCRP过表达细胞系的构建及其在鸡常用药物渗透性检测中的应用博士王丽平科研计划自然科学兽医学75 王洪OTA对肠上皮细胞屏障功能的影响及其氧化应激机制研究博士黄克和科研计划自然科学兽医学76 谢青云DEXH/D-box RNA解螺旋酶RHA参与调控宿主细胞对PEDV感染的免疫应答博士刘斐科研计划自然科学兽医学77 李林泌乳生物学及乳品质调控博士张源淑科研计划自然科学兽医学78 孟凡强植物乳杆菌素163合成代谢调控机制博士陆兆新科研计划自然科学食品科学与工程79 周婷多胺调控绿豆芽菜植酸降解机理研究博士顾振新科研计划自然科学食品科学与工程80 邱佳容基于Ⅰ型PKS 基因挖掘新型微生物次级代谢产物的研究博士辛志宏科研计划自然科学食品科学与工程81 刘强基于电子鼻检测桃果实病害及病害对果实气味的影响机制博士屠康科研计划自然科学食品科学与工程82 陈贵杰基于肠道菌群调控能量代谢研究茶多糖调节脂肪代谢机理博士曾晓雄科研计划自然科学食品科学与工程83 杜雅珉新型N-糖酰胺酶的重组表达、酶学特性及应用研究博士刘丽科研计划自然科学食品科学与工程84 焦琳舒L-天冬酰胺酶分子设计及改良的研究博士吕凤霞科研计划自然科学食品科学与工程85 邢路娟火腿成熟过程中抗氧化钛的活性变化研究博士张万刚科研计划自然科学食品科学与工程86 李静农户测土配方施肥技术采纳的影响评价及其激励政策选择博士冯淑怡科研计划人文社科公共管理87 张国磊资源禀赋、联镇包村与基层社会治理差异化研究博士张新文科研计划人文社科公共管理88 王健“营改增”对地方政府土地财政及其行为影响研究博士吴群科研计划人文社科公共管理89 许明军“土地规模经营”和“服务规模经营”是两难选择吗?博士冯淑怡科研计划人文社科公共管理90 张海滨动态能力视角下大学学科成长力模型构建及实证研究——以涉农学科为例博士董维春科研计划人文社科公共管理91 李发志快速城镇化地区土地利用变化与生态效应及优化布局研究博士孙华科研计划人文社科公共管理92 张绍阳中国式分权、府际博弈与土地约束性指标管控政策执行偏差博士欧名豪科研计划人文社科公共管理93 高富岗地方政府工业用地供应:制度环境、合约结构与引资效应博士石晓平科研计划人文社科公共管理94 于帅辣椒在中国的传播与本土化研究博士王思明科研计划人文社科科学技术史95 陈加晋民国时期饲料科技近代化研究博士李群科研计划人文社科科学技术史96 李恒征电喷镀Ni-P/BN复合镀层的应用研究博士康敏科研计划自然科学农业工程97 程准大型拖拉机液压机械无极变速箱最优传动比控制规律研究博士鲁植雄科研计划自然科学农业工程98 杨振杰土壤蒸汽消毒机的设计与研究博士汪小旵科研计划自然科学农业工程99 宋长友miR-144对团头鲂氧化应激调控的研究博士徐跑科研计划自然科学水产100 张武肖饲料中亚麻酸与VE对团头鲂幼鱼氨氮胁迫的营养调节研究博士戈贤平科研计划自然科学水产101 宋飞彪Igf3基因在福瑞鲤性腺发育中的作用机制研究博士董在杰科研计划自然科学水产102 刘勇男磷脂酰肌醇在热诱导灵芝次级代谢中的调控机理研究博士赵明文科研计划自然科学生物学103 陈乐麦草畏脱甲基酶Dmt66 的优化改良与转基因应用的研究博士何健科研计划自然科学生物学104 王远丽根瘤菌S41及其突变菌风化黑云母的转录组学研究博士盛下放科研计划自然科学生物学105 刘磊ZmCCaMK自磷酸化位点鉴定及其在BR诱导抗氧化防护中的功能博士张阿英科研计划自然科学生物学106 刘雪松DNA甲基化调节OsZIP1转录表达及镉转运机理的研究博士杨志敏科研计划自然科学生物学107 宋萍拟南芥磷脂酶D调控微管形态和应答高温胁迫的分子机理博士章文华科研计划自然科学生物学108 黄俊伟转pdmAB拟南芥与sphingobiumsp.1017-1联合修复环境中取代脲类除草剂的研究博士蒋建东科研计划自然科学生物学109 吴雪小白菜重金属镉阻隔剂及阻隔调控机理研究博士崔瑾科研计划自然科学生物学110 张毅华一氧化氮介导甲烷缓解渗透胁迫对绿豆萌发的抑制博士沈文飚科研计划自然科学生物学111 侯志U1 RNA参与表观遗传调控的mRNA 3’polyA位点选择的分子机理博士腊红桂科研计划自然科学生物学112 刘宇海滨雀稗水通道蛋白PvPIP1的磷酸化位点和耐盐功能鉴定博士黄炳茹科研计划自然科学草学113 石晓磊金融信息化背景下中国家庭消费行为研究博士董晓林科研计划人文社科应用经济学114 江峥嵘糖启动水稻弱势粒灌浆的生理生化机制硕士丁艳锋实践计划自然科学农业推广115 陈学君抗纹枯病小黑麦种质创新与评价硕士亓增军实践计划自然科学农业推广116 张璐手性噁唑啉类扁桃酰胺的设计、合成及抑菌构效关系研究硕士李圣坤实践计划自然科学农业117 韦珊银杏内生真菌Chaetomium globosumYSC5次生代谢产物研究硕士叶永浩实践计划自然科学农业118 桑程巍灰霉病菌对咯菌腈抗药性性状在后代中遗传分化研究硕士陈长军实践计划自然科学农业119 沈阳UASB-倒置A2/O联合处理高氨氮养猪场废水工艺优化研究硕士王电站实践计划自然科学环境工程120 汪迎高附加值茯砖茶的研制与开发硕士庄静实践计划自然科学农业推广硕士121 梁超凡不结球白菜小孢子培养技术研究与创新硕士李英实践计划自然科学农业推广硕士122 夏美玲薄皮甜瓜新品种Gsb6-4DUS检测及其富硒功能瓜的开发硕士钱春桃实践计划自然科学农业推广硕士123 王华微型耐弱光盆栽荷花产业化生产技术体系的优化硕士徐迎春实践计划自然科学农业推广硕士124 吴胜规模化猪场舍内颗粒物和有害气体分布规律及颗粒硕士李春梅实践计划自然科学畜牧学物化学成分分析125 戴婕徐州市种植业与养殖业合作社运行机制的差异性分析硕士何军实践计划人文社科农业推广硕士126 陈子豪农户对生防技术采纳行为研究——以桔小食蝇饵剂为例硕士胡浩实践计划人文社科农业推广硕士127 刘虹富含GABA发芽青稞营养粉的研制硕士韩永斌实践计划自然科学食品工程128 曹念念基于近红外和计算机视觉的黄桃脆片品质分级研究硕士潘磊庆实践计划自然科学食品工程129 王丹五种豆类水溶性多糖的提取、活性研究及功能性饮品研发硕士李伟实践计划自然科学食品工程130 张仁康鳄鱼肉脯产品开发硕士肖红梅实践计划自然科学食品加工与安全131 赵空暖水稻生长模型参数自动校正与栽培方案设计平台开发硕士姜海燕实践计划自然科学农业硕士132 杨乐基于多元时间序列数据的水稻全生育期气候预测数据集重建研究硕士姜海燕实践计划自然科学农业硕士133 徐晓辉优势视角下提升社区单亲家庭儿童抗逆力的实务研究硕士姚兆余实践计划人文社科农业硕士134 尹圣珍江苏休闲茶业创意发展的路径研究硕士朱世桂实践计划人文社科农业硕士135 高建波重金属污染土壤温和淋洗剂研究与应用硕士董长勋实践计划自然科学化学工程136 文进康旋耕刀轴自动焊接设备研发硕士康敏实践计划自然科学机械工程137 高空基于农业装备驾驶室人机工程试验台的自动调节机构开发硕士杨飞实践计划自然科学机械工程138 雷波物联网技术在装配式建筑供应链管理中的应用研究硕士金德智实践计划自然科学物流工程139 魏昌曼日本中餐馆菜名日译研究——以大阪市为中心硕士李红实践计划人文社科翻译硕士140 甄亚乐中国文学“走出去”之中外译者合作模式研究硕士曹新宇实践计划人文社科翻译硕士141 王赫仲戊基TeA生产工艺及大田试验硕士陈世国实践计划自然科学生物工程142 王婧杰长三角地区重金属Cd低积累叶菜品种的筛选硕士郑录庆实践计划自然科学生物工程143 张晔农村小微企业融资需求自我抑制:类型、程度与缓解硕士董晓林实践计划人文社科金融144 范文静“政银担”农业信贷模式创新研究——基于安徽、山东两省实践硕士黄惠春实践计划人文社科金融145 艾歆基于“沙集模式”的农村网商融资方式选择研究硕士周月书实践计划人文社科会计146 王翠玲金融知识对金融资产持有行为及投资获利的影响硕士王翌秋实践计划人文社科金融147 张钰基于价值链的成本效益研究-以邳州宿羊山为例硕士陈东平实践计划人文社科会计148 许志鹏多年生黑麦草种质资源的耐热性筛选与评价硕士徐彬实践计划自然科学草业联系人:联系电话:传真号码:电子信箱:研究生教育管理部门(公章)年月日。
菊花新品种DUS_测试指南测试性状的选择

DOI:10.19462/ki.zgzy.20231216001菊花新品种DUS测试指南测试性状的选择徐 旭 苏江硕 邓 波 张 飞 陈素梅 房伟民 陈发棣 管志勇(南京农业大学园艺学院/南京农业大学作物遗传与种质创新国家重点实验室/农业农村部景观设计重点实验室,江苏南京210095)摘要:随着我国菊花育种活跃度的不断提升,菊花品种的形态多样性不断丰富,迫切需要提高菊花DUS测试指南适用性。
本研究连续2年对210份形态多样的菊花品种进行观测统计,以NY/T 2228—2012《植物新品种特异性、一致性和稳定性测试指南菊花》涉及的68个性状及本研究群体中发现的9个多样性性状或表达状态为观测依据,对现有菊花品种DUS测试指南做出优化,建议在2012年版测试指南中增加9个测试性状,剔除1个选测性状,并调整2个性状的名称表述,旨在提高菊花新品种鉴定的效率。
关键词:菊花;DUS指南;形态多样性;测试性状Selection of Test Characters in DUS Test Guideline for NewChrysanthemum VarietiesXU Xu,SU Jiangshuo,DENG Bo,ZHANG Fei,CHEN Sumei,FANG Weimin,CHEN Fadi,GUAN Zhiyong(College of Horticulture,Nanjing Agricultural University/State Key Laboratory of Crop Genetics and Germplasm Innovation,Nanjing Agricultural University/Key Laboratory of Landscape Design,Ministry of Agriculture and Rural Affairs,Nanjing 210095)植物新品种特异性(Distinctness)、一致性(Uniformity)和稳定性(Stability)测试(以下简称DUS测试)是新品种保护的技术基础和授权的科学依据[1]。
南农硕士毕业要求

南农硕士毕业要求南京农业大学是一所以农业、生物科学技术为主体的综合性大学。
南农拥有丰富的科研资源和广阔的发展空间,培养出了大量的优秀人才。
作为南农的研究生,在毕业要求方面需要达到以下要求:一、学术要求1.毕业时至少发表或录用一篇CSSCI或SCI来源论文。
作为研究生,需要具备一定的科研能力,要求学生在研究生学习期间至少发表或录用一篇CSSCI或SCI来源论文,提高自己的学术影响力。
2.在导师的指导下完成独立的研究课题并作出成果。
毕业要求学生能够在导师的指导下,独立进行一项课题研究,并取得相应的成果。
这能够锻炼学生的科研能力,提高其对科研问题的解决能力。
3.掌握一门外语,具备一定的阅读和撰写科技论文的能力。
要求研究生掌握一门外语,以便进行国际间的学术交流以及阅读和撰写外文论文等。
1.完成独立的教学任务并取得较好的教学效果。
毕业要求学生完成独立的教学任务,具备很强的教学能力,能够独立设计课程和实验,并有较好的教学效果。
2.参与教学和科研管理的相关工作,积极参加各类学术交流活动。
毕业要求学生积极参与教学和科研管理的相关工作,例如参与科研项目申报和管理,积极参加学术会议、学术讲座等各类学术交流活动。
这对学生的职业发展有着非常重要的作用。
三、素质要求学生的素质是评价学生的综合素养的标准之一,毕业要求研究生具备以下素质:1.具备很强的创新精神和创新能力。
研究生需要具备很强的创新精神和创新能力,这有助于学生更好地解决科研问题,掌握先进的科技知识,迎接未来的挑战。
2.注重团队合作精神。
研究生需要注重团队合作精神,要具备团队合作的能力,与导师和同学之间建立起良好的合作关系,团结协作完成各项科研任务。
3.具备扎实的基础理论知识和专业知识。
研究生需要具备扎实的基础理论知识和专业知识,深入掌握专业技能,为今后的科研发展打下坚实的基础。
总之,南京农业大学要求研究生在学术、教学、素质等多方面都达到一定的水平,掌握先进的科技知识和科研方法,培养优秀的研究生人才,为国家和社会做出贡献。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
一、成果简介
1.主要解决的研究生教育实践问题
研究生教育是我国高等教育的最高层次,研究生教育国际化承担着培养具有国际竞争力高水平人才的重要任务,是建设世界一流大学和一流学科的重要驱动力。
但是,目前我国高校尤其是农林院校研究生教育国际化程度很低,主要表现在:
1)具有海外交流经历的研究生规模很小。
由于海外交流成本很高,所以受众面很小,出国交流人数占研究生总人数比例很低。
2)研究生校园国际化交流氛围不浓厚。
研究生绝大部分为中国本土学生,难以形成多元文化共存的校园氛围。
3)研究生课程国际化程度不高。
研究生课程设置及实施缺乏国际化理念,全英文课程建设不成体系。
4)研究生国际化教育制度机制不完善。
学校研究生国际化教育尚处于发展阶段,国际化运作机制不成熟。
2. 解决实践问题的方法
在学校国际化发展战略的推动下,我校研究生教育以培养具有国际视野及国际竞争力的高层次人才理念为引领,突破传统的“小规模、贵族式”国际化教育方式,积极探索新路径,在理念、形式和内容上做出转变,探索“大规模、大众化”的研究生国际化教育。
主要经验有:
1)首创研究生选派机制,提高选派学生质量,多形式多渠道不断扩大研究生出国学术交流规模。
2)开创以研究生为主导的研究生国际学术会议,提升留学生招生规模和培养水平,营造校园国际交流氛围,搭建研究生国际化交流平台。
3)建设研究生全英文课程体系,为国际化教育提供支撑。
3. 创新点
1)创新研究生选派机制,研究生出国质量和规模逐年扩大,全面拓宽了研究生国际视野;首创直博生全英文论坛、国外访学新模式。
2)开创以研究生为主导的国际学术会议,扩大了农林学科海外影响力,为农林高校研究生打开了国际交流窗口。
3)建立与世界接轨、体现农林院校特色的全英文课程体系,在兄弟院校中示范推广。
构建了科学、规范、长效的研究生国际化培养体系,研究生国际化教育理念深入人心。
4.推广应用成果及贡献
通过全英文答辩遴选,研究生出国规模及质量逐年提升,全面拓宽了研究生国际视野。
大部分研究生从国外归来均发表了高水平论文,成为了我国农业科技发展的优秀青年领军人才。
研究生自己组织筹划的国际学术会议,为研究生搭建了一个高水平的学术交流平台,促进了我校及国内外涉农领域研究生的深度交流,产生了深远的影响力。
会议受到中国社会科学报等多家社会媒体关注报道,《学位与研究生教育》杂志与我校约稿,交流研究生国际会议举办的实践经验。
建立与世界接轨、体现农林院校特色的全英文课程体系,深受研究生们欢迎,63%的学术型研究生选修了全英文课程。
“学术学位研究生全英文教学改革实践与研究”被江苏省教育厅设为重点建设项目,并建议在省内高校甚至全国农林高校中示范推广。
《高级植物营养学》课程已成为教育部“2013年度来华留学英语授课品牌课程”。
《有机肥与土壤微生物》等5门课程入选江苏省高校省级英文授课精品课程。
研究生教育成果突出,培养质量不断提高。
人均SCI收录论文篇数由2005年0.4上升到2015年1.5;SCI单篇累计平均影响因子从2012年的3.0上升到2015年的4.4;博士学位论文已连续三年在国家抽检中全部合格。
研究生国际化培养成效受到各界认可和媒体报道。
西北农林科技大学等众多高校来校交流经验;江苏省教育频道《教改再扬帆》、中国社会科学报、江苏省教育厅有关领导等高度评价了我校研究生国际化培养成效。
研究生国际化教育体系日趋完善,研究生、导师及教育管理人员的国际化培养理念全面提升,形成研究生国际化培养体系创新与实践长效发展的合力。
2。