端粒与端粒酶的研究进展
关于端粒及端粒酶的调查报告

关于端粒及端粒酶的调查报告一:引言2009年10月5日,诺贝尔生理学或医学奖颁发给了美国科学家伊丽莎白·布莱克本、卡罗尔·格雷德和杰克·绍斯塔克,以表彰他们在研究端粒和端粒酶保护染色体的机理方面的贡献。
这篇调查报告将会通过资料查询和逻辑推断等方式论述关于端粒,端粒酶以及它们与肿瘤细胞的相关内容。
二:端粒和端粒酶2.1.1端粒端粒(Telomere)是染色体末端的重复DNA序列,在人细胞中长度约为几千到一两万碱基对,它防止细胞将天然染色体末端识别为染色体断裂,起着保护和稳定染色体的作用。
[1]随着细胞的分裂增殖,端粒会逐渐缩短。
当端粒的长度缩短到一定程度时,细胞的分裂便会停止。
因此,端粒具有调节细胞增殖的作用,是细胞分裂的“时钟”。
端粒的碱基序列具有极高的保守性,但不同物种的端粒仍有差异,例如:四膜虫重复序列为GGGGTT,草履虫为TTGGGG,人类和哺乳动物为TTAGGG.[2]2.1.2端粒的结构端粒通常由富含G的DNA重复序列,以及端粒结合蛋白和端粒相关蛋白组成。
端粒结合蛋白直接保护端粒DNA,端粒相关蛋白通过与端粒结合蛋白的相互作用间接影响端粒的功能。
端粒既可保护染色体不受核酸酶的破坏,又避免了因DNA黏性末端的裸露而发生的染色体融合。
[4][5]2.2.1端粒酶端粒酶(Telomerase),在细胞中负责端粒的延长的一种酶。
在端粒发现之后,人们便开始猜测存在这样一种酶,可以起到延长端粒的作用——因为随着细胞的分裂增殖和染色体的复制,端粒应当越来越短,但是某些细胞(如肿瘤细胞)的端粒长度却能够保持相对不变。
在1997年,Tom Cech实验室的Lingner在Euplotes aediculatus以及酿酒酵母中发现了真正的端粒酶催化亚基。
[3]2.2.2端粒酶的作用机理端粒酶主要依靠两种成分来实现其功能,一种名为端粒酶逆转录酶(TERT)的蛋白酶,另一种是作为模板的一小段RNA序列。
端粒与端粒酶的研究进展

体 外培养的 细胞衰老与体 内衰老 的一个 共 同特征 是染
色体 端 粒 的逐 渐 缩 短 。H r y al 等 率 先证 明 了 正常 细胞 衰 老 e
时的端粒丢失 。 J发现 体外培养的人成纤维细胞端粒平均 他 f ] 长度 以一种复制倚赖性方式缩短 。 这种缩短也与体 内衰老相 关, 老年供体成纤维细胞和外周血 细胞中端粒平均长度 比年 轻 人短 ,lg o m等 研 究 了三个年 龄组发现 , Sa b o 人类有 丝分 裂 细胞 中端粒 长度 与供体年 龄呈高度负相关 , 并报道 外周血 细胞端粒缩短的速率是 3 b / 1p 年 端粒酶是影响端粒长度的 最主要的目索 , 端粒缩短后 可在端牡酶 的作用下得 以合成 而 使端粒延长 , 正常 的体细胞 并无端粒酶活性 为 探讨端 粒 但
至死亡 将这种正确 人体细胞在体外分裂潜能受限制 的现象 称为细胞衰老 , 并认为其是“ 衰老在细胞水平的表现 ” 。现 已 发现 许多其 他娄 型的 人娄 细胞 也存 在着这 种现象 . : 如 表皮 角质细胞 、 平滑肌 细胞 、 晶状 体细胞 、 神经胶质 细胞、 内皮细
胞 、 巴细 胞 以 及 肾上 腺皮 质 细 胞 淋
真 核生物 中都检 测到了端粒酶 的活性 。
端粒酶 属于逆转 录酶 , 是棱 糖核蛋 白复台体 , 主要 成丹 是 RN 和 蛋 白质 . 有 引物 特 异 识别 位 点 , 以 自身 的 A 含 能
端垃和端粒酶是近年来生命科学研 究的热. 端粒随细 氨。 胞的分裂进行性缩 短 , 胞的衰老过 程也就 开始启动了。端 细 粒酶活性和端粒长度 应正相 关 , 间接地 影响了细胞的衰老过 程 。而端粒酶 的活化使一些细胞 获得 无限增殖 的能力 . 在永
端粒和端粒酶的发现历程

端粒和端粒酶的发现历程廖新化引言2009年诺贝尔生理学或医学奖授予了UCSF(加州大学旧金山分校)的Elizabeth Blackburn (简称Liz),Johns Hopkins University(约翰霍普金斯大学)的Carol Greider(简称Carol),以及Howard Medical School(哈佛医学院)的Jack Szostak。
诺贝尔奖主页上介绍她/他们获奖的原因是揭示了“how chromosomes are protected by telomeres and the enzyme telomerase”(染色体是如何被端粒和端粒酶保护的)。
端粒和端粒酶的研究进程中贯穿着“发现现象/问题”-“提出概念/模型”-“实验验证”的思路,整个过程就像相继解开一个个puzzle(智力谜团)一样有趣,充满了思想的光辉。
重现这个思路对科学工作者是有启发意义的。
本文也提供了一个很好的科学问题推演的教学案例。
染色体末端的两个难题以及端粒的概念20世纪70年代初,对DNA聚合酶特性的深入了解引申出了一个染色体的复制问题。
DNA 聚合酶在复制DNA的时候必须要有引物来起始,而且它的酶活性具有方向性,只能沿着DNA5’到3’的方向合成。
染色体复制之初可以由小RNA作为引物起始合成,之后细胞的修复机器启动,DNA聚合酶能够以反链DNA为模板,以之前合成的DNA为引物,合成新的DNA取代染色体中间的RNA引物。
但是线性染色体最末端的RNA引物因为没有另外的引物起始,没有办法被DNA取代。
所以线性染色体DNA每复制一轮,RNA引物降解后末端都将缩短一个RNA引物的长度(图1,简化的示意图,实际上染色体的DNA双链末端不会是平的)。
尽管这个引物不长,但是细胞千千万万代地不断复制,如果不进行补偿,染色体不断缩短,最终就会消失。
James Watson(因为发现DNA双螺旋结构获得诺奖)最早就明确指出了这个“末端隐缩问题”,并猜想染色体也许可以通过在复制前联体(染色体末端跟末端连起来)的方式来解决末端复制的问题[1]。
端粒、 端粒酶的研究进展

况下,端粒作为细胞“分裂时钟”能缩短,最终导 致细胞脱离细胞周期。
端粒酶(telomerase)催化端粒合成,是一种逆转 录酶,能以自身的RNA为模板,反向合成— TTAGGG—,不断地加在DNA未端,肿瘤细胞能够不 断地分裂增生,“端粒酶—RNA”结构的存在是先决条 件。
随着每次细胞分裂,染色体末端逐渐缩短,细
胞进入不可逆的抑制状态,直至细胞衰老,此过程 即称为复制性衰老。人类体细胞遵循这个规则从细 胞出生到衰老,单细胞生物遵循这个规则分裂后定 有其它机制保持单细胞生物传代存活,生殖细胞亦 如此,这些细胞怎样保持细胞具有继续分裂或长期 分裂的能力呢?科学家们发现端粒确实随着每次分 裂而缩短,但也会被新合成的端粒片断再延长。科 学家们怀疑,可能尚有末被发现的酶,该酶具有标 准的DNA多聚酶所不具备的功能,能使已缩短的端 粒延长,使科学家们兴奋的是到1985年首先在四膜 虫中证实了这种能使端粒延长的酶—端粒酶的存在。
体细胞的端粒有限长度(telomere restriction fragments TRFS)大多数明显短于生殖细胞,青年 人的TRFs又显著长于年长者,提示TRFs随着细胞 分裂或衰老,在不断变短,主要是由于DNA聚合酶 不能完成复制成线性DNA末端所致。缺少端粒的染 色体不能稳定存在,这是因为端粒DNA与结构蛋白 形成的复合物如同染色体的一顶“帽子”,它既可
端粒、 端粒酶的研究进展
早在30年代,两名遗传学家Muller和Mcclintock
分别在不同的实验室用不同的生物做实验发现染色 体末端结构对保持染色体的稳定十分重要,Muller 将这一结构命名为端粒(telomere)。直到1985年 Greider等从四膜虫中真正证实了端粒的结构为极 简单的6个核苷酸TTAGGG序列的多次重复后发现 了端粒酶(telomerase TRAP-eze) 。
端粒端粒酶研究进展

端粒端粒酶研究进展端粒是染色体末端的一段DNA序列,它起到保护染色体稳定性和完整性的作用。
然而,由于染色体在每次细胞分裂时会缩短一段,当端粒长度过短时,染色体会发生异常,并最终导致细胞老化和死亡。
端粒酶则是一种重要的酶,它能够补充并保持端粒的长度稳定。
近年来,对于端粒和端粒酶的研究取得了许多重要的进展。
首先,科学家们对于端粒和端粒酶的结构和功能进行了深入的研究。
端粒由重复的TTAGGG序列组成,这些序列会通过端粒酶的作用被补充。
端粒酶主要由两个亚基组成:一个叫做端粒酶反转录酶TERT,另外一个则是端粒酶RNA(TERC)。
TERT具有酶的活性,而TERC则是TERT的模板,用于合成新的端粒DNA。
端粒酶通过不断循环地合成新的端粒DNA来补充端粒的长度,从而延长染色体的寿命。
其次,研究表明端粒和端粒酶在癌症中具有重要的作用。
在正常细胞中,端粒的长度会随着细胞的分裂而缩短,从而限制了细胞的生命周期。
然而,在肿瘤细胞中,端粒酶的活性会显著增加,导致细胞端粒的长度不断维持,并且细胞可以无限制地分裂。
这种增强的端粒酶活性对于肿瘤细胞的免疫逃逸、增殖和转移等方面起着重要的作用。
因此,端粒酶已成为癌症治疗的一个重要靶点,研究人员已经开始开发端粒酶抑制剂,以抑制肿瘤的生长和扩散。
此外,最近的研究发现,端粒和端粒酶在衰老过程中也发挥了重要的作用。
随着年龄的增长,端粒长度会逐渐缩短,从而引发细胞衰老和组织功能下降。
研究人员尝试通过增强端粒酶的活性来抑制细胞衰老,以延长寿命和改善老年病的发生率。
实验证据显示,通过增加端粒酶的表达或给予端粒酶活性的药物可以有效地抑制细胞衰老。
这些发现为老年病的治疗和延长寿命提供了新的研究方向。
总之,端粒和端粒酶在细胞衰老、癌症等疾病方面的研究进展迅速。
研究人员们对于端粒和端粒酶的结构和功能有了更深入的了解,并且逐渐揭示了它们在疾病中的重要作用。
未来的研究将继续深入探究端粒和端粒酶的调控机制,并开发出更具针对性的治疗手段,为人类健康的维护做出更大的贡献。
端粒和端粒酶结构与作用的研究进展

选择已有 的确定点,并且布设一条长度确定的全站仪高程 与管理 ,2018(02):186-187.
闭合 的路线,并且需要联测所有的 GPS网之中的 “点”。
根据国家在测量的精准度方面的四等水准要求 ,必须要利 作者简介 邓勋 (1987-),男,汉族 ,湖南省安化县,助理工程师
用垂直角 中丝法来每部分往返三次 的进 行测 回。经过实际 本科 ,研究方向:测量。
机通常会选用 四五台南方的 9600北极星 GPS。机器 的标准 明相较于其他测量 的方式,在进行矿山的测量工作 中,通
称精度是平面少于或者多于 5mm+lO咱以内。高程是 多于或 过对矿山地面控制的测量的结果表明,更加适合测量 的方
者少于 lOmm+lO 以内,通常采用静态测量 的模式来作业, 法就是 GPS结合全站仪 。GPS技术与全站仪 的有机结合 ,
2
信 息记 录材料 2 01 8年 8月 第 1 9卷第 8期
数 目和长度 因物种而异 ,人类端粒主要 由 5’一(TTAGGG) 方案 。2015年斯坦福大学发表 的一项研究报告给出了新
rr_3’重复 DNA序列构成 。除少数细胞如生殖细胞 、干细胞 、 的启示:可 以借鉴端粒 酶的有力功能,开发一种可控 的功
的要求 。在进行测量 中,观测量采用无约束平差的方式, 更劳 固,前景更加美好 。
一 般进行二维约束平差的时候,选用 1954北京坐标系。
4.3高程测量 GPs网
【参 考文献 】
在进行高程测量中 GPS技术的应用较为广泛。相对来 [1]张高兴 ,李忠金 .GPS结合全站仪在矿 山地面控 制测量 中的
信 息记 录材料 2 01 8年 8月 第 1 9卷 第8期
端粒及端粒酶的研究进展

生物化学与生物物理进展PROGRESS IN BIOCHEMISTRY ANDBIOPHYSICS1999年 第26卷 第5期 Vol.26 No.5 1999端粒及端粒酶的研究进展任建国 周军 戴尧仁摘要 端粒是染色体末端独特的蛋白质-DNA结构,在保护染色体的完整性和维持细胞的复制能力方面起着重要的作用.端粒酶则是由RNA和蛋白质亚基组成的、能够延长端粒的一种特殊反转录酶.端粒长度和端粒酶活性的变化与细胞衰老和癌变密切相关.端粒结合蛋白可能通过调节端粒酶的活性来调节端粒长度,进而控制细胞的衰老、永生化和癌变.研制端粒酶的专一性抑制剂在肿瘤治疗方面有着广阔的前景.关键词 端粒,端粒酶,衰老,永生化,癌变学科分类号 Q50Progress in the Studies of Telomere and Telomerase.REN Jian-Guo, ZHOU Jun, DAI Yao-Ren(Department of Biological Science and Biotechnology, Tsinghua University, Beijing 100084,China).Abstract Telomeres are unique DNA-protein complexes at the terminals of chromosomes that play a critical role in protecting chromosomal integrity and in maintaining cellular replicative potential. Telomerase is a specialized reverse transcriptase, composed of both RNA and protein subunits, that elongates telomeric repeats. The changes in telomere length and telomerase activity are closely linked to cell aging and carcinogenesis. Telomere binding-protein may regulate telomere length by regulating telomerase activity, and then control cell aging, immortalization and carcinogenesis.The development of specific telomerase inhibitors will have broad prospect in the aspect of tumor therapy.Key words telomere, telomerase, aging, immortalization,carcinogenesis 近年来,有关端粒及端粒酶的研究异常活跃,许多新的结构和功能的发现使之成为生物学和医学关注的热点.本文拟对端粒及端粒酶的最新进展予以阐述.1 端粒(telomere) 端粒是真核细胞内染色体末端的蛋白质-DNA结构,其功能是完成染色体末端的复制,防止染色体免遭融合、重组和降解[1~3].从单细胞的有机体到高等的动植物,端粒的结构和功能都很保守.1.1 端粒DNA 大多数有机体的端粒DNA由非常短而且数目精确的串联重复DNA排列而成,富含鸟嘌呤.个别种类的端粒DNA重复单元很长.此外,果蝇的端粒结构非常新颖,重复序列是一个可互换的因子.端粒的DNA序列多种多样,其功能不需要独特的序列来维持.尽管在许多物种中端粒DNA有相当大的变化,但仍可在进化关系非常远的生物中发现相同的端粒序列,如所有的脊椎动物、原生动物锥虫及几种粘菌和真菌都有相同的端粒序列T2AG3.其他情况下,尽管不同的有机体有不同的端粒序列,但彼此总有明显的相关性. 端粒DNA的平均长度因物种而异.在人中大约15 kb,在大鼠中可长达150 kb,在小鼠中一般在5~80 kb之间变化,而在尖毛虫中却只有20 bp.在所有的有机体中,端粒DNA的长度总是波动变化的.酵母的端粒DNA在200到400 bp间随遗传或营养状态的改变而改变.四膜虫和锥虫等有机体的端粒长度在对数期会持续增加.相反,在人体中,随着细胞的持续分裂,端粒会缓慢缩短.1.2 端粒结合蛋白 目前对端粒结合蛋白还了解甚少.在酵母中,端粒的主要结合蛋白是Rap1p,在体外以很高的亲和性与端粒上的许多识别位点相结合.研究表明,Rap1p与端粒长度的调节有关,Marcand等[4]认为Rap1p能够阻止端粒酶接近端粒从而负调节端粒的长度.相反,Ray等[5]的研究结果表明Rap1p可以在端粒周围通过聚集端粒酶或提高端粒酶活性而延长端粒.编码尖毛虫端粒小体蛋白的基因和RAP1的基因没有相似的序列. 该蛋白的结合需要单链的T4G4T4G4尾.同Rap1p不同的是,此蛋白仅限于同染色体的末端相结合,故称之为末端专一性DNA结合蛋白.此外,在爪蟾提取物中也检测到末端专一性结合活性.因此末端限制性结合蛋白可能是端粒染色质的一个普通特性. 在人中已经鉴定出两个端粒重复序列结合蛋白(telomeric repeat-binding factor,TRF).TRF1是一个60 ku的同源二聚体双链TTAGGG重复序列结合蛋白,包含一个Myb型的C端螺旋-转折-螺旋区和一个DNA结合折叠的同源区,N端是酸性疏水区[6].另一个端粒结合蛋白是TRF2,它与TRF1很相似,所不同的是其N端碱性很强[7].两种蛋白在体外都专一性地与双链TTAGGG重复序列结合,在体内则位于端粒.人的hTRF与Rap1p 没有同源性.长期过表达TRF1将导致端粒逐渐地和过程性地变短.该过程可能通过抑制端粒酶活性而实现.TRF2则可以防止染色体末端相互融合[2].最近,Kim等[8]在水稻中也鉴定出三个TTAGGG专一性结合蛋白复合物.这些复合物对双链DNA及富含胞嘧啶的单链序列无亲和性.其功能目前仍不清楚.2 端粒酶(telomerase) 端粒酶是一种自身携带模板的反转录酶,催化端粒DNA的合成,能够在缺少DNA模板的情况下延伸端粒寡核苷酸片段.其活性取决于它的RNA和蛋白质亚基[9].Prescott等发现酵母的端粒酶至少包含两个功能性相互作用的RNA分子,两者都可充当DNA聚合作用的模板,端粒酶至少包含两个活性位点.端粒酶除了具有反转录活性外,还具有核酸内切酶的活性[10].小腔游仆虫中有活性的端粒酶复合物的分子质量大约是230 ku,含有一个约66 ku的RNA亚基及两个123 ku和43 ku的蛋白质亚基,大亚基专一性地和端粒DNA底物结合.端粒酶的主要功能是维持染色体末端的端粒序列,从而抵消因细胞分裂而导致的端粒DNA的消耗.最近发现,端粒酶另外一个重要的功能就是合成串联重复的TTAGGG序列,为TRF2提供结合位点,防止染色体的末端融合[2]. 端粒酶的RNA亚基是合成端粒DNA的模板,对于端粒酶的结构和催化活性都十分重要.四膜虫端粒酶RNA有159个核苷酸,模板区为5′-CAACCCCAA-3′.人端粒酶RNA有455个核苷酸,模板区为5′-CUAACCCUAAC-3′.不同种类的纤毛虫,其端粒酶RNA长度在148~209之间变化,其中9~15个核苷酸具有种的专一性,与特定种类的端粒DNA序列互补.端粒酶RNA重要序列缺乏保守性,但都有保守的二级结构.这对于保持端粒酶的活性极为重要.端粒酶RNA的基因已经在纤毛虫、酵母、小鼠、人等生物中得到了克隆.将突变的RNA基因导入细胞后发现这些改变的序列在端粒DNA中出现,表明端粒酶的RNA决定了端粒DNA的序列.在酵母或乳酸菌中,缺失单拷贝的端粒酶RNA 基因会导致端粒缩短和细胞死亡.这些证据表明模板RNA对端粒酶的活性至关重要. Romero等和McCormick-Graham等推导出一个端粒酶RNA的二级结构模型:从5′到3′方向包含四个保守的双螺旋,双螺旋Ⅰ是最保守的区域,双螺旋Ⅱ、 Ⅲ、Ⅳ是茎环结构,这些保守的茎环通常是蛋白质结合区域.在双螺旋Ⅱ与Ⅲ之间存在模板序列,其上游的保守序列5′-(CU)GUCA-3′限制了模板区的5′边界.在双螺旋Ⅳ中有一个结构上保守的GA结,有助于蛋白质的识别与结合.最近研究表明,模板区的位置因物种而异.Autexier等[11]为了阐明端粒酶中RNA亚基的功能,将一系列缺失或替换一定数量碱基的RNA与野生型端粒酶蛋白质亚基进行酶的重构,研究了RNA特殊二级结构区域对端粒酶活性的影响. 当5′端和茎环Ⅰ、Ⅱ和Ⅳ中的残基缺失或替换时,端粒酶的活性降至野生型的15%~35%.表明这些结构对端粒酶的活性很重要.缺失5′端大于11以上的残基时酶活性完全丧失.说明一些重要的序列或结构上的相互作用都发生在这一区域.有趣的是,影响端粒酶RNA潜在假结的突变、缺失整个茎环Ⅲ和替换茎环Ⅳ中的GA 结,并不明显影响酶的活性. 端粒酶的蛋白质成分不如RNA亚基研究得那样清楚.在过去几年里,端粒酶的催化亚基已经在酵母[12,13]、原生动物[12]和人[14]中分离出来.该蛋白质亚基的功能区与已知的反转录酶(reverse transcriptase, RT)在序列和功能上有明显的相似性,故称为端粒酶反转录酶(telomerase reverse transcriptase, TRT).酵母的Est1p是一个77 ku的多肽,专一性地与RNA亚基结合.缺失该基因,细胞会产生如同缺失端粒酶RNA亚基一样的表型. Weinrich等发现在端粒酶特殊的保守区和RT组分中,单个氨基酸的改变会降低或消除端粒酶的活性,直接证明hTRT是端粒酶的催化蛋白组分.在四膜虫中,发现两个端粒酶相关的蛋白质p80和p95.p80专一性地和端粒酶RNA结合,而p95则可和G链引物交联.在人和啮齿类动物中,已发现p80的同源物[15].从小腔游仆虫中纯化的端粒酶中发现另外两个蛋白质p123和p43,这两个蛋白质似乎与p80和p95没有相关性[12].p123包含有RT组分,是酵母Est2p的同源物[12].Est2p的RT组分对于体内、体外端粒DNA的合成是必需的.Est2p/p123在真核生物中很保守,在反转录酶的进化树上代表一个很早的分支[13].目前,仍然不清楚的是生物界里是否存在两类端粒酶,一类依赖于p80和p95;另一类依赖于p123/Est2p. 端粒酶的特殊性使端粒酶活性的测定在研究中显得尤为重要.早期的测定方法是通过测定细胞提取物将端粒重复片段加到一个合成的寡聚脱氧核苷酸引物3′端的能力进行的.但由于哺乳动物细胞端粒酶含量低,又有干扰现象,故难度较大.Kim等[16]建立了灵敏、快速、高效的端粒重复序列扩增法(TRAP),以后又在引物方面作了改进.此后人们又相继建立了荧光法、原位端粒重复片段扩增法及TRAP与闪烁技术联用的SPA法等敏感的检测手段,在医疗检测中得到了迅速的应用.3 端粒及端粒酶与衰老和癌变的关系3.1 端粒及端粒酶与衰老的关系 越来越多的证据表明端粒长度控制着衰老进程,端粒缩短是触发衰老的分子钟. 在大多数正常的人体细胞中并不能检测到端粒酶的活性,端粒随细胞分裂每次丢失50~200个碱基.Cooke等认为,这是由于正常的人体细胞中端粒酶未被活化,导致了端粒DNA缩短的缘故.保护性端粒酶的减少可能最终制约了细胞的增殖能力.当几千个碱基的端粒DNA丢失后,细胞就停止分裂而衰老.端粒及端粒酶涉及衰老最有力的证据是Bodnar等的工作.Bodnar等[17]将人的端粒酶基因导入正常的细胞中,使得端粒酶异常表达.活化的端粒酶导致端粒序列异常延长,细胞旺盛增殖,细胞寿命大大延长.这一结果首次为端粒钟学说提供了直接的证据.3.2 端粒酶活化与肿瘤 在正常的人体细胞中,端粒程序性地缩短限制了转化细胞的生长能力,这很可能是肿瘤形成的一个抑制机制.端粒酶的重新表达在细胞永生化及癌变过程中起着重要的作用.有人甚至认为表达端粒酶的正常细胞更易癌变.人们在代表不同肿瘤类型的大约1 000多个活检样品中发现大约85%的样品呈端粒酶阳性反应.相反,90%以上的邻近正常组织却是端粒酶阴性.从而将这个酶与永生化和肿瘤的形成密切联系在一起!端粒酶活性与肿瘤的这种特殊关系使之在诊断与治疗方面具有重要的应用价值[18,19].对端粒酶活性的抑制可能对某些类型的肿瘤来说是一个很有意义的治疗方法[20].3.3 衰老和肿瘤发生的分子机制 细胞衰老和癌变与端粒及端粒酶的关系可以表述如下:端粒的数量控制着细胞的增殖能力,是细胞内的分裂钟.端粒酶在生殖细胞系中维持端粒的长度,随着细胞的发育端粒酶活性受到抑制,端粒持续变短.当正常人体细胞的端粒缩短至一定程度时,启动阻止细胞分裂的信号,细胞开始衰老死亡,此即所谓的Hayflick界限(M1期).另外一些细胞由于癌基因、抑癌基因等的突变逃逸M1期,获得一定的额外增殖能力,进入第二死亡期(M2).此时端粒酶仍为阴性,端粒进一步缩短.大部分细胞达到极限而死亡,生存下来的细胞具有无限增殖的能力,端粒酶重新活化,成为永生细胞.在肿瘤形成过程中,端粒的延长是一个重要的甚至是一个必要的步骤! 既然端粒异常缩短后会触发细胞衰老和癌变,那么细胞一定有某种方式监测端粒的长度变化并用这些信息来调节端粒酶的活性,从而将这些重复序列加到染色体的末端.研究酿酒酵母时,发现端粒重复序列结合蛋白Rap1p负调节端粒的延长.最近van Steensel等[21]与Cooper等[22]分别在酵母和人中发现一个新的端粒重复序列结合蛋白,同样阻止端粒的延长.Cooper等[22]在粟酒裂殖酵母中克隆了Taz1p的基因,此蛋白质与端粒DNA的双链结合.值得注意的是,尽管粟酒裂殖酵母,酿酒酵母和人的端粒重复序列不同,Taz1p、Rap1p和TRF1这三个端粒序列结合蛋白却有相似的DNA结合区(类Myb型).在这个结合区以外,Taz1p与TRF1几乎没有同源性,与Rap1p就根本没有同源性.然而,taz1+基因的突变与Rap1p碳末端平头突变却有相似的表型,即端粒片段大大延长.这些新的工作表明端粒长度的调节机制是高度保守的. 细胞究竟是怎样调节端粒的长度的呢?van Steensel等[21]首次报道了人端粒结合蛋白(TRF1)的功能性研究,并提出端粒长度的调节机制.在端粒酶阳性的肿瘤细胞系HI1080中,长期过表达TRF1导致端粒逐渐的和持续性的缩短.相反,当TRF1负显性突变后,失去与端粒DNA结合的功能,最终诱导了端粒的延长.证明TRF1是端粒延长的一个抑制因子,负反馈调节端粒的长度.由于在可检测的水平上并不影响端粒酶的表达,因此,van Steensel等认为TRF1与端粒DNA结合后,顺式抑制端粒酶的活性,从而控制端粒的长度.根据这些结果,他们提出一个简单的端粒长度调节模型:与端粒重复片段结合的TRF1的数量可以调节端粒酶.野生型蛋白的加入,增加了端粒上TRF1的数量,从而为端粒酶提供了一个负信号.然而,通过负显性突变使TRF1功能缺失,却导致端粒酶的活化和端粒的延长.总之,这些研究表明,端粒重复序列的双链结合蛋白负调节端粒的延长.Shore[23]指出:细胞内可能存在一个感受染色体末端重复序列结合蛋白数量的机制,当这个数量超过一定的界限后,就产生一个信号阻止由端粒酶引起的端粒延长,或者,此信号可以活化缩短端粒重复片段的核苷降解或重组的过程.去除重复片段结合位点的不完全复制或降解事件,将消除对端粒酶的抑制.目前,人们还不清楚上述信号是如何产生与传导的.Ku等发现一些细胞周期抑制剂、DNA损伤因子、TopⅡ抑制剂均不能抑制端粒酶的活性,相反,一些蛋白激酶C(protein kinase C, PKC)的抑制剂却能大大地降低端粒酶的活性.究其原因一方面可能因为PKC的失活使得活化端粒酶表达的效应分子不能活化,另一方面PKC可能在体内直接调节端粒酶的活性.c-myc是细胞增殖与凋亡的一个中心调节子,c-myc的表达严格依赖于分裂信号,被生长抑制信号或分化信号所抑制.Fujimoto等发现抑制c-myc的表达能够抑制端粒酶的活性,表明原癌基因c-myc对于端粒酶的调节是必需的.Mandal等发现在HeLa细胞中过表达Bcl-2导致端粒酶的活性增加5~10倍.Maxwell等的结果却表明端粒酶的活性不受P53的过量表达及凋亡的影响.这些证据表明端粒酶活性的调节是一个复杂的过程,它与细胞内一系列信号识别与传导有关系,其详细的调节机制还有待进一步的研究.3.4 端粒假说遇到的挑战 最近的研究表明,端粒酶的活化并非肿瘤细胞中的独特现象,许多正常增殖的细胞中也观察到了端粒酶的活化.Starling等、Kipling等及Broccoli等在小鼠中的研究结果表明,端粒缩短同衰老和肿瘤间并没有密切的联系.在正常人的口腔角化细胞的衰老过程中,也未观察到端粒的缩短.Blasco等[1]通过基因敲除使小鼠中的端粒酶RNA基因缺失,导致端粒酶的活性丧失.发现在快速增殖的器官中,细胞由于缺乏端粒酶而凋亡[3].但丧失端粒酶活性的细胞在培养中能够永生化、被病毒癌基因转化及在裸鼠中形成肿瘤.在某些肿瘤去分化的过程中端粒酶活性也未受到抑制.研究小鼠皮肤乳头状瘤的结果表明,端粒酶的活性与增殖率没有密切联系.总之,澄清这些例外的事实需要更加深入细致的研究,以期找到一个合理的解释. 总之,端粒和端粒酶在衰老和癌变中的作用使得人们对研究前景充满信心.对端粒和端粒酶深入细致的研究将有助于清楚地阐明衰老和肿瘤的机理,为在实践中抗衰老和治疗肿瘤提供新的理论基础.目前关于端粒及端粒酶的研究主要集中在以下几个方面:a.端粒酶的结构和功能.b.端粒酶的纯化和激活机制.c.寻找端粒酶的专一性抑制剂及其在抗癌中的应用.d.端粒的高级结构及结合蛋白的作用机理.这几个方面仍需进一步的探索.衰老和癌变无疑都是多因素作用的结果,但端粒和端粒酶很可能在其中扮演重要的角色.作者单位:清华大学生物科学与技术系,北京 100084参考文献1 Blasco M A, Lee H W, Hande M P, et al. Telomere shortening and tumor-formation by mouse cells lacking telomerase RNA. Cell, 1997, 91(1):25~342 van Steensel B, Smogorzewska A, de Lange T. TRF2 protects human telomeres from end-to-end fusions. Cell, 1998, 92(3):401~4133 Lee H W, Blasco M A, Gottlieb G J, et al. Essential role of mouse telomerase in highly proliferative organs. Nature, 1998, 392(6676):569~5744 Marcand S, Gilson E, Shore D. A protein-counting mechanism for telomere length regulation in yeast. Science, 1997, 275(5302): 986~9905 Ray A, Runge K W. The C-terminus of the major yeast telomere binding-protein Rap1p enhances telomere formation. Mol Cell Biol, 1998, 18(3):1284~12956 Bianchi A, Smith S, Chong L, et al. TRF1 is a dimer and bends telomeric DNA. EMBOJ,1997, 16(7):1785~17947 Bilaud T, Brun C, Ancelin K, et al. Telomeric localization of TRF2, a novel human telobox protein. Nature Genet, 1997, 17(2):236~2398 Kim J H, Kim W T, Chung I K. Rice proteins that bind single-stranded G-rich telomere DNA. Plant Mol Biol, 1998, 36(5):661~6729 Nakamura T M, Cech T R. Reversing time:origin of telomerase. Cell, 1998, 92(5):587~590 10 Greene E C, Bednenko J, Shippen D E. Flexible positioning of the telomerase-associated nuclease leads to preferential elimination of nontelomeric DNA. Mol Cell Biol, 1998, 18(3):1544~155211 Autexier C, Greider C W. Mutational analysis of tetrahymena telomerase RNA: identification of residues affection telomerase activity in vitro. Nucl Acids Res, 1998, 26(3):787~79512 Lingner J, Hughes T R, Shevchenko A, et al. Reverse-transcriptase motifs in the catalytic subunit of telomerase. Science, 1997, 276(5312):561~56713 Nakamura T M, Morin G B, Chapman K B, et al. Telomerase catalytic subunit homologs from fission yeast and human. Science, 1997, 277(5328):955~95914 Meyerson M, Counter C M, Eaton E N, et al. Hest2, the putative human telomerase catalytic subunit gene, Is Up-regulated in tumor-cells and during immortalization. Cell, 1997, 90 (4):785~79515 Harrington L, Mcphail T, Mar V, et al. A Mammalian telomerase-associated protein. Science, 1997,275(5302): 973~97716 Kim N W, Piatyszek M A, Prowse R K, et al. Specific association of human telomerase activity with immortal cells and cancer. Science, 1994, 266(5193):2011~201517 Bodnar A G, Ouellette M, Frolkis M, et al. Extension of life-span by introduction of telomerase into normal human cells. Science, 1998, 279(5349):349~35218 Hoos A, Hepp H H, Kaul S, et al. Telomerase activity correlates with tumor aggressiveness and reflects therapy effect in breast-cancer. Int J Cancer, 1998, 79(1):8~1219 Kyo S, Takaura M, Tanaka M, et al. Telomerase activity in cervical cancer is quantitatively distinct from that in its precursor lesions. Int J Cancer, 1998,79(1):66~7020 Hoos A, Hepp H H, Kaul S, et al. Telomerase activity correlates with tumor aggressiveness and reflects therapy effect in breast cancer. Int J Cancer, 1998, 79(1):8~1221 Vansteensel B, Delange T. Control of telomere length by the human telomeric protein Trf1. Nature, 1997, 385(6618):740~74322 Cooper J P, Nimmo E R, Allshire R C, et al. Regulation of telomere length and function by aMyb-domain protein in fission yeast. Nature, 1997, 385(6618): 744~74723 Shore D. Telomeres-different means to common ends. Nature, 1997, 385(6618): 676~677收稿日期: 1998-07-07, 修回日期: 1998-09-25。
端粒及端粒酶的研究进展

端粒和端粒酶的研究进展

• 端粒和端粒酶简介 • 端粒和端粒酶的研究历史与现状 • 端粒和端粒酶与人类健康 • 端粒和端粒酶的实验研究方法 • 总结与展望
01
端粒和端粒酶简介
端粒的结构和功能
端粒的结构
端粒是由DNA和蛋白质组成的结 构,位于染色体末端,保护染色 体免受损伤和降解。
端粒的功能
端粒的主要功能是维持染色体的 稳定性和完整性,防止染色体融 合和降解,同时参与细胞分裂和 衰老过程。
相关疾病。
端粒和端粒酶的调控机制
03
目前,研究者们正在深入研究端粒和端粒酶的调控机制,以期
更好地理解其在细胞生命活动中的作用。
未来研究方向和展望
01
深入探究端粒和端粒酶的作用机制
未来研究需要进一步深入探究端粒和端粒酶的作用机制,以更好地理解
其在细胞生命活动中的作用。
02
开发基于端粒和端粒酶的治疗方法
未来可以开发基于端粒和端粒酶的治疗方法,用于治疗相关疾病。
03
加强跨学科合作与交流
未来需要加强跨学科合作与交流,促进端粒和端粒酶研究的深入发展。
03
端粒和端粒酶与人类健康
端粒和端粒酶与衰老
端粒与衰老
端粒是染色体末端的保护结构,随着细胞分裂次数的增加,端粒长度逐渐缩短, 导致基因组不稳定和细胞功能异常,最终引发衰老。
端粒酶与衰老
端粒酶是一种维持端粒长度的酶,通过激活端粒酶可以延长端粒长度,从而延缓 衰老过程。
端粒和端粒酶与疾病的关系
端粒与心血管疾病
心血管疾病患者中,端粒长度缩短与动脉粥样硬化、心肌梗死等 疾病的发病风险增加相关。
端粒与癌症
端粒酶的异常激活可以导致细胞无限增殖,形成肿瘤,因此与癌症 的发生和发展密切相关。
端粒及端粒酶的研究进展

Your site here
LOGO
端粒酶应用的展望
肿瘤的诊断 :研究结果表明,85%的
恶性肿瘤组织端粒酶活性均呈阳性,而癌旁 组织的阳性率仅有4%。因此端粒酶活性可 以作为肿瘤诊断的指标之一。可以用许多 材料来检测端粒酶的活性,不必采用手术取 样,因此简单易行,操作方便。且端粒酶活性 水平还可以区分肿瘤的恶性程度,从而选择 恰当的治疗方法。
端粒及端粒酶 研究进展
——王振
宇
LOGO
端粒和端粒酶的概述
2009年,美国的三位科学家Elizabeth H· Blackburn、Carol W· Greider和Jack W· Szostak发表了题为“端粒和端粒酶是 如何保护染色体的”而共同获得诺贝尔生 理学或医学奖。也是从这一重大研究成果 开始,端粒和端粒酶的研究为人类衰老和 肿瘤带来了福音。
Your site here
LOGO
端粒和端粒酶
端粒酶的结构: 一般认为端粒酶由端粒 酶RNA(TR)、端粒酶相关蛋白和端粒酶催 化亚基(TERT)3部分组成。许多物种的TR 已经被成功克隆,如纤毛虫、酵母、人和鼠 等。
Your site here
LOGO
端粒的功能:
1.保护染色体末端 真核生物的端粒DNA-蛋白复合物,如帽 子一般,保护染色体末端免于被化学修饰或 被核酶降解,同时可能还有防止端粒酶对端 粒进行进一步延伸的作用。改变端粒酶的模 板序列将导致端粒的改变,从而诱导细胞衰 老和死亡。
Your site here
LOGO
端粒的功能:
3.决定细胞的寿命
染色体复制的上述特点决定了细胞分裂 的次数是有限的,端粒的长度决定了细胞的 寿命,故而被称为“生命的时钟”。
端粒及端粒酶的研究进展

综述文章编号:1009-0002(2007)04-0667-03端粒及端粒酶的研究进展杨平勋,黄君健军事医学科学院生物工程研究所,北京100850[摘要]端粒是真核细胞染色体末端的特有结构,是由端粒结合蛋白和一段重复序列的端粒DNA组成的一个高度精密的复合体,在维持染色体末端稳定性,避免染色体被核酸酶降解等方面起着重要的作用。
端粒的长度、结构及组织形式受多种端粒结合因子的调控。
由于端粒的重要性,在哺乳动物细胞里,端粒的长度或端粒结构变化与癌症发生及细胞衰老有密切的关系。
由于末端复制问题的存在,随着细胞分裂次数的增加,端粒不断缩短,细胞不可避免的走向衰老或凋亡。
由于在细胞分裂过程中端粒长度的不断缩短与细胞分裂代数增加具有相关性,即端粒长度反应了细胞的分裂次数,因此有人将端粒形象的比喻为生物时钟。
在90%的癌细胞中,端粒酶被重新激活,以此来维持端粒的长度,使细胞走向永生化。
简要综述了端粒、端粒酶及端粒酶结合蛋白的最新研究进展。
[关键词]端粒;端粒酶;端粒结合蛋白[中图分类号]Q26[文献标识码]AAdvancesofTelomereandTelomeraseYANGPing-xun,HUANGJun-jianBeijingInstituteofBiotechnology,Beijing100850,China[Abstract]Telomeresarespecializednucleoproteincomplexesattheendsoflinearchromosomes,consistingofmulti-kilo-baselongarraysofdouble-strandedTTAGGGrepeats,asingle-strandedoverhangofthe3'G-strand,andassociatedpro-teins.Itprotectstheendsoflinearchromosomes.Telomerelength,constructionandorganizationareregulatedbylotsoftelomerebindingproteins.Becauseofthesignificanceoftelomeres,thechangesoftelomeres'lengthorconstructurecancausecellstoentersenescenceorapoptosis.Telomeraseisimportantformaintainingtelomeres,andmostofcancercellshavetelomeraseactivity,so,itcanbeatargetforcurecancer.Thelatestdevelopmentofthetelomere,telomeraseanditsbindingproteinsfunctionwerediscussed.[Keywords]telomereUtelomeraseUtelomere-bindingprotein端粒的发现应归功于20世纪30年代2位遗传学家芭芭拉・麦克林托克和赫尔曼・穆勒的观察结果。
端粒、端粒酶研究及应用进展

疗和延缓衰老。
了 TC除A些B1以蛋外白Y具来有自以于前美未国知斯的坦功福大能学。这医些学新院的的发研现究对人于员以还后确的定
端 粒 酶 成 分 的 确 定 和 端 粒 酶 的 制 造 ,端 粒 酶 活 性 珠 激 活 和 抵 制 药
需要着重研究端粒与肿瘤的关系。了解是什么原因激活了癌细胞
的 端 粒 酶 ,导 致 了 在 正 常 人 体 细 胞 中 产 生 了 端 粒 酶 活 性 的 异 变 。
科技论文与案例交流
161
端 粒 、端粒酶研究及应用进展
韦伟 (西 华 师 范 大 学 西 南 野 生 动 植 物 资 源 保 护 教 育 部 重 点 实 验 室 西 华 师 范 大 学 生 命 科 学 学 院 四 川 南 充 637009)
摘 要 :端 粒 、端 粒 酶 是 一 种 与 人 体 的 肿 瘤 的 预 防 和
究 领 域 研 制 出 延 缓 衰 老 ,抗 癌 的 药 品 。 3.2端粒与肿瘤的关系
端 粒 酶 的 结 构 和 成 份 ,并 了 解 如 何 制 造 和 激 活 它 的 活 性 ,而这将
众 所 周 知 ,恶性肿瘤细胞中具有较高活性的端粒酶,它能维
有助于医学界制造出抑制和激活端粒酶的药物,帮助癌症患者治 持癌细胞端粒的长度,使其无限制扩增。因此下一阶段的研究还
如果能搞清这个问题,将是人为攻克癌症的一大发现。 3.3端粒与心血管疾病
生 态
物的制造都是非常有意义的。
相关研究说明心血管疾病中的端粒缩短可能是冠心病、心血 环
另外还有来自于美国索尔克生物研究所的科研人员发现了 管疾病的致病因素之一。但是至今科学家们仍没有发现两者之间 境
可 以 控 制 端 粒 酶 关 闭 的 开 关 。研 究 者 发 现 的 这 个 “关 闭 ”开关可能 有助于让端粒酶的活跃度低于这一限度。这一开关的发现明显有 助于研究者进一步发现如何激活和抑制端粒酶的活性具有很大
端粒、端粒酶研究及应用进展

身 RN A为模 板 通过 逆转 录 而合成 的 , 端粒 酶通 过 在染色 体末
端添加 端粒序 列 , 从而使 端粒 能够 固定 在某个 特定 长度 _ 7 _ 。人 端粒酶包括 3个重要部分 , 即人端 粒酶 R A( u ntlm rs N h ma o eae e R A, T R A 、 端 粒 酶相关 蛋 白和人 端 粒酶 催 化 亚单 位 。 N h R N )人 端粒 酶 长度 在人 类 各种 细胞 中差别 很大 , a oy等 [发 现端 G vr 8 1
融合及 序列缩短等异 常 , 而这些异常又 可能受端粒酶 的调控 。
1 端 粒 、端粒 酶 的发 现
上 世 纪初 , 名遗 传 学家 Mc l tc 【与 MulrH t发 著 Ci okB】 n J l Jt e 2
端 粒酶属 于逆转 录酶 , 端粒 的核 糖核酸 酶就是端 粒酶 以 自
年代 , afc 究发 现 : 过体外 培养 的正 常人 成纤 细胞 的 Hyi l k研 经
复制过 程并非 细胞 的死 亡过程 , 只是细胞 群 中的大部分 细胞 而
在经历 了数次 分裂增殖 后停滞 在了某 个特定状态 , 仅是基 因 仅
粒酶 3 末端 残基 至少要 求存在 3个核苷 酸片 断 , 才能保 持整 个端 粒酶 的活性 。纯 化端 粒酶 全酶可 知人 类端 粒 酶复合 物 的
逐渐 变短 。基 于 以上末 端复制 特点 , 端粒会 随着细胞 分裂逐 渐 缩短 , 每分 裂 1 次缩短 5 0~ 10h , 0 p 端粒缩 短到其临 界长度时 , 就会 导致细胞 衰老甚 至死亡 。因此 , 正常真 核细胞 的端粒 可被
称 作 “ 命 之 钟 ”I 生 6 1 。
现: 染色体 的稳定性 和完整性是 由染色体 的末端来 维持 的。基 于此 发现 , l rH 将 其命 名 为 “eo ee , 定 义来 源于 希 Mul J e tlm r” 此
端粒及端粒酶活性检测方法研究进展

端粒及端粒酶活性检测方法研究进展
张晓娜;杨镒峰;许保增
【期刊名称】《特产研究》
【年(卷),期】2017(039)004
【摘要】缩短的端粒是细胞老化的信号,检测端粒酶活性可间接测定细胞状态.端粒酶通过p66Shc信号通路对胚胎发育永久性停滞产生影响.本文对哺乳动物卵母细胞或胚胎永久性停育与端粒酶活性水平变化的关系、端粒酶活性检测方法研究进展及各种方法的优缺点进行综述,包括端粒重复扩增法(TRAP)、TRAP-酶联免疫吸附(ELISA)法、TRAP-银染法、杂交链式信号放大反应结合磁分离技术法和亚甲蓝MB作为G-四联体结合探针法、H-GNs比色法、依赖无标记分子信标的级联放大DNA机制法.
【总页数】7页(P40-46)
【作者】张晓娜;杨镒峰;许保增
【作者单位】中国农业科学院特产研究所特种动物分子生物学省部共建国家重点实验室,长春130112;内蒙古大学,呼和浩特01002;中国农业科学院特产研究所特种动物分子生物学省部共建国家重点实验室,长春130112;中国农业科学院特产研究所特种动物分子生物学省部共建国家重点实验室,长春130112
【正文语种】中文
【中图分类】Q753
【相关文献】
1.端粒酶活性检测及检测方法的研究进展 [J], 张艳;刘柏林
2.端粒酶活性检测方法研究进展 [J], 郭林燕;阳明辉
3.端粒酶活性检测方法的研究进展 [J], 姜国彦;李敏;危艳武
4.端粒酶活性检测方法研究进展 [J], 邱广斌;崔娴维;张甦
5.端粒酶活性检测方法的改进及dNTP浓度对端粒酶活性的影响 [J], 吴东林;张玉静;李鹏;陈守义;阮承迈
因版权原因,仅展示原文概要,查看原文内容请购买。
端粒和端粒酶在癌症中的研究进展及意义

端粒和端粒酶在癌症中的研究进展及意义摘要:端粒是位于染色体末端的DNA串联重复序列,对基因组稳定性和完整性起保护作用。
端粒的长度与细胞周期密切相关。
其长度变化机制分为依赖端粒酶活性和端粒重组两类,氧化应激和铅(Pb)与端粒酶的功能蛋白相结合抑制其活性,致使端粒缩短,硒(Se)与二者具有拮抗作用,延缓衰老。
相关数据表明85%肿瘤细胞与端粒酶活性成正相关,以端粒酶活性作为肿瘤治疗靶标称为当代热点之一。
主要对肺癌、乳腺癌等恶性肿瘤与端粒的相关性进行了综述,以期为端粒和端粒酶在癌症治疗研究提供参考依据。
关键词:端粒;端粒酶;肿瘤20世纪30年代,人们开始了解染色体上的一种特殊结构——端粒。
端粒是存在于真核细胞线状染色体末端的一种特殊结构,与端粒结合蛋白一起构成了特殊的“帽状”结构,维持染色体的完整和细胞活性,其实质为一小段DNA-蛋白质复合体。
端粒与有丝分裂有着密切的联系,细胞每分裂一次,端粒就缩短30~200bp,当缩短到2~4kb,会导致细胞复制功能衰退,引起细胞衰老或死亡,被科学家称为“有丝分裂时钟”和“生命时钟”[1,2]。
端粒的延长和重组机制都是通过端粒酶来催化和介导的,端粒酶在保持端粒稳定、基因组完整、细胞活性和潜在的增殖能力等方面发挥重要作用。
鉴于端粒酶在正常组织体细胞中的活性被抑制,而在肿瘤中则被重新激活,可能参与肿瘤恶性转化的机制,成为医学界研究的重点和热点之一。
2009年美国3位科学家因发现端粒和端粒酶结构及其对染色体末端的保护功能,而获诺贝尔生理学或医学奖。
1端粒和端粒酶1.1端粒的结构和功能端粒是位于染色体末端由一个富含G的DNA串联重复序列[3]和端粒结合蛋白组成,每个重复序列一般为5—7bp[4]。
不同物种其重复序列存在l~2个碱基差异,哺乳动物的端粒重复序列为5’-(TTAGGG)n-3’[5],植物的端粒重复序列为5’一(TTTAGGG)n-3’[6]。
端粒长15~20kb,其重复序列成T环结构,像帽子一样能有效防止染色体间末端重组、融合和染色体退化[7]。
端粒与端粒酶
端粒酶结合蛋白(TEP
)
TEP1、生存动力神经细胞基因(SMN) 产物、
、
PinX1、 Est1p 和Est3p
端粒酶是在染色体末端不断合成端粒序 列的酶,它可以维持端粒的长度,维持细胞 增殖潜能。端粒酶以自身RNA为模板合成端 粒酶重复序列,具有逆转录酶活性,它的活 性不依赖于DNA聚合酶,对RNA酶、蛋白酶 和高温均敏感。端粒酶活性表达能稳定端粒 的长度,抑制细胞的衰老,在生殖细胞和干 细胞中可检测到高水平的端粒酶活性。
㈠ 端粒酶的结构 端粒酶在结构上为一核糖核蛋白复合体,由RNA 和 结合的蛋白质组成,是RNA依赖的DNA 聚合酶。它是一种 特殊的能合成端粒DNA的酶,通过明显的模板依赖方式每 次添加一个核苷酸。
端粒酶实质上是一种特殊的逆转录酶
端粒酶RNA(hTR)
端粒酶逆转录酶(TERT)
端粒酶结合蛋白(TEP)
端粒酶延长端粒长度以减慢细胞衰老最早 的 证 据 来 自 Bodnar 等 的 研 究 , 1 9 9 8 年 其 在 Science上刊文报道:将人的端粒酶基因导入 端粒酶阴性的正常人体细胞中激活其表达并培 养细胞,然后与未导入该基因的细胞比较,发 现前者端粒明显增长,细胞分裂旺盛,细胞寿 命比后者大大延长,更令人关注的是细胞并无 肿瘤样改变。
端粒
染色体
细胞分裂
端粒
细胞分裂
细胞将停止分裂而趋于老化
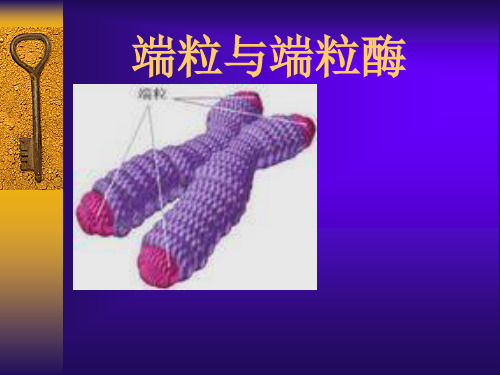
端粒:是真核细胞线性染色体末端特殊结构。
由端粒DNA和端粒相关蛋白组成。 端粒DNA:为不含功能基因的简单、高度重复序列, 在生物进化过程中具有高度保守性。
不同物种的端粒DNA 序列存在差异。
人类及其它脊椎动物染色体端粒的结构是 5′TTAGGG3′的重复序列, 长约15kb。体细胞的端粒有 限长度(telomere restriction fragments TRFS)大多数 明显短于生殖细胞,青年人的TRFs又显著长于年长 者,提示TRFs随着细胞分裂或衰老,在不断变短, 主要是由于DNA聚合酶不能完成复制成线性DNA末 端所致。
端粒和端粒酶的研究进展

3
端粒长度和端粒酶活性与多种疾病(如癌症、衰 老相关疾病等)的发生和发展密切相关。
03 端粒酶的类型、分布及调 控机制
端粒酶的类型与分布
端粒酶的类型
根据结构和功能不同,端粒酶主要分为两种类型,即端粒酶逆 转录酶(TERT)和端粒酶RNA(TERC)。其中,TERT具有催 化活性,而TERC则作为模板参与端粒DNA的合成。
基因组学和转录组学分析
通过基因组学和转录组学技术,全面分析端粒和端粒酶相关基因在 细胞中的表达谱和调控网络。
06 展望与未来研究方向
端粒和端粒酶研究的挑战与机遇
挑战
端粒和端粒酶的研究仍面临许多技术 上的挑战,如难以在体内直接观测端 粒长度和端粒酶活性,以及缺乏特异 性高的端粒酶抑制剂等。
机遇
随着基因编辑、高通量测序等新技术的 发展,端粒和端粒酶的研究将迎来新的 机遇,有望更深入地揭示其在细胞衰老 、肿瘤发生等领域的作用机制。
端粒和端粒酶在肿瘤发生和发展中扮演重要 角色,因此针对端粒和端粒酶的靶向药物研 发有望为肿瘤治疗提供新的策略和方法。
THANKS FOR WATCHING
感谢您的观看
的风险。
端粒酶在肿瘤细胞中的活性
02 端粒酶在多数肿瘤细胞中被重新激活,以维持端粒长
度,促进肿瘤细胞的无限增殖。
端粒酶抑制剂与肿瘤治疗
03
针对端粒酶的抑制剂已成为潜在的肿瘤治疗策略,通
过抑制端粒酶活性来诱导肿瘤细胞衰老或凋亡。
端粒和端粒酶在衰老相关疾病中的作用
01
端粒缩短与衰老
端粒缩短被认为是衰老的生物标 志物之一,与多种衰老相关疾病 的发生发展密切相关。
03
疗也具有重要意义。
国内外研究现状及发展趋势
端粒与端粒酶研究进展

端粒与端粒酶研究进展
王宏斌
【期刊名称】《中山大学研究生学刊:自然科学与医学版》
【年(卷),期】2001(022)001
【摘要】本文介绍了端粒,端粒酶的结构,功能,并较为详细地阐述了端粒-端粒酶假说及其实验证据以及端粒研究的应用前景。
【总页数】3页(P41-43)
【作者】王宏斌
【作者单位】中山大学生命科学学院遗传学99硕,广州510275
【正文语种】中文
【中图分类】Q7
【相关文献】
1.端粒、端粒酶、端粒酶抑制剂与消化系统肿瘤的研究进展 [J], 许兰涛;孙延庆;张玉萍
2.端粒酶活性调节机制与端粒酶抑制剂研究进展 [J], 何国平;税青林
3.人子宫颈组织中端粒酶人端粒酶RNA组分基因表达和人端粒酶蛋白基因编码催化蛋白亚基mRNA的相关性分析 [J], 宋建明;周平;吴俊辉;昝沁;许美权
4.不同食管病变中端粒酶活性与端粒酶RNA组分、端粒酶逆转录催化亚单位的相关性及其意义 [J], 张蕾;温洪涛;张云汉;李靖若
5.端粒酶和端粒酶抑制剂研究进展 [J], 葛晶
因版权原因,仅展示原文概要,查看原文内容请购买。
端粒-端粒酶在神经退行性疾病中的研究进展

端粒-端粒酶在神经退行性疾病中的研究进展
吴志悦;薛晓帆;曾景蓉;黄安琪(综述);许二赫(审校)
【期刊名称】《中风与神经疾病杂志》
【年(卷),期】2024(41)2
【摘要】神经退行性疾病多以β-淀粉样蛋白(β-amyloid,Aβ)、α-突触核蛋白(α-synuclein,α-syn)作为生物标记物来辅助临床进行诊断。
近年来,人们发现染色体末端的端粒能够作为衡量生物衰老程度的指标,并发现端粒长度、端粒酶活性可能作为评估老年神经退行性疾病发病风险、疾病进展、不良预后的血液标记物,但目前相关国内外研究结果并不具有一致性。
了解端粒相关生物标记物在年龄相关疾病中的作用,是可以帮助临床医生更好地了解疾病的发生发展机制。
现就端粒-端粒酶系统在神经退行性疾病中的最新研究进展综述如下,以介绍端粒长度、端粒酶活性对神经退行性疾病的影响以及潜在作用机制。
【总页数】6页(P169-174)
【作者】吴志悦;薛晓帆;曾景蓉;黄安琪(综述);许二赫(审校)
【作者单位】首都医科大学宣武医院神经内科;首都医科大学附属北京朝阳医院神经内科
【正文语种】中文
【中图分类】R742;R749.16
【相关文献】
1.端粒-端粒酶与衰老退行性变的关系
2.端粒、端粒酶、端粒酶抑制剂与消化系统肿瘤的研究进展
3.端粒及端粒酶在恶性血液病中的研究进展
4.端粒/端粒酶在恶性血液病中的研究进展
5.端粒、端粒酶、端粒酶抑制剂与消化系统肿瘤的研究进展
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
端粒与端粒酶的研究进展
【摘要】研究显示,端粒酶活性被激活,可维护端粒的长度,细胞将会延缓衰老,避免癌变。
此外,端粒酶的发现还在理论上丰富和发展了分子肿瘤学,据研究显示90%的人体肿瘤与端粒酶相关,若我们通过端粒酶活性的检测,提前预知肿瘤的发生,从而提前预防和治疗,或者若我们能使癌细胞中的端粒酶再度“休眠”,恶性肿瘤就会停止生长,以此来治疗癌症。
【关键字】端粒端粒酶肿瘤癌症衰老染色体
1.端粒和端粒酶的概述
2009年,美国的三位科学家Elizabeth H·Blackburn、Carol W·Greider和Jack W·Szostak发表了题为“端粒和端粒酶是如何保护染色体的”而共同获得诺贝尔生理学或医学奖。
也是从这一重大研究成果开始,端粒和端粒酶的研究为人类衰老和肿瘤带来了福音。
端粒是真核细胞染色体末端的帽子样的结构,它具有稳定染色体末端结构,防止染色体DNA降解和末端融合,保护染色体结构基因,调节正常细胞生长等作用。
同种生物不同组织的细胞,甚至相同组织的不同细胞由于处于不同的生命时相,端粒的长度也不一样。
由此可发现端粒的长度跟细胞的寿命、衰老与死亡有密切关系,所以端粒的长度被称为“生命时钟”【1】。
端粒酶(telomerase)是一种以自身RNA为模板,将端粒DNA合成至染色体的核糖核蛋白复合物(ribonucleoprotein,RNP)。
端粒长度的维持需要端粒酶的激活。
所以端粒酶在保持端粒稳定、基因组完整、细胞长期的活性和潜在的继续增殖能力等方面有重要作用。
端粒酶的活性存在于人的生殖细胞、肿瘤细胞、永生化细胞系和再生性组织中,一般情况下酶的活性处于抑制状态,只有当端粒体受到损
伤的时候,端粒酶才被激活。
由于端粒和端粒酶对肿瘤和癌症的发生有很大关系,所以近年来,端粒和端粒酶的研究也比较多,且主要是在妇产科学、基础医学、心血管疾病、泌尿科学、外科学等方面,其中端粒酶与肿瘤形成关系的研究占总文献比例最大【2】。
2.端粒和端粒酶的结构
端粒是存在于染色体3'末端的特殊部位,通常由一些简单重复的序列组成。
不同种类的细胞端粒重复序列不同,大多长约5-8bp。
人类的端粒序列由5 '
-TTTAGGG-3 '组成,长约5—15bp。
端粒和结合蛋白组成核蛋白复合物,进化上有高度的保守性【3】。
端粒酶的核心成分是端粒酶RNA(TER)和端粒酶反转录酶(TERT),其中TER 亚基是合成端粒DNA的模板,对于端粒酶的结构和催化活性都十分重要,而TERT仅在端粒酶阳性细胞中正常表达,其它组织细胞不表达或活性较低。
TERT 基因由三个结构域组成:N端、中心结构域和C端。
N端与端粒酶活性、RNA结合、蛋白的聚集和端粒延伸有关,中心结构域含有端粒酶特有的一些基序,保守性较高,相反C端保守性较低【4】。
3.端粒的功能
3.1端粒与衰老:
端粒有长短,一般来说,端粒的长度是随年龄增加而越来越短的。
端粒消失,会使染色体发生畸变,从而使人类细胞丧失复制能力,最终导致细胞衰老。
端粒酶,则具有调控端粒长短的能力,其活性也随年龄大小而不同,年轻时,活性大,较容易延长端粒,这是年轻人不易显老的原因。
此外,男性端粒长度缩短略快于女性,这也是男性平均寿命低于女性的原因。
研究认为,延缓衰老的方法主要是降低身体新陈代谢速率,如少吃少饮。
新陈代谢率越高,细胞分裂次数就越多,端粒缩短越快,寿命也就越短。
另外用药物刺激体内的干细胞,弥补衰老损耗的细胞,也能延长端粒长度,以至延缓老化,其中生长激素的抗衰老效果比较肯定,如用生长激素注射法可以调动干细胞f类似物来延长端粒【5】。
3.2端粒与肿瘤
据实验研究证明,恶性肿瘤细胞中具有较高活性的端粒酶,它能维持癌细胞端粒的长度,使其无限制扩增。
在众多肿瘤疾病中,端粒对妇科肿瘤的发生发展关系最为密切,其中包括宫颈癌、子宫内膜癌、卵巢癌、妊娠滋养细胞肿瘤等。
Sakamoto等用PCR及TRAP法检测了妇科肿瘤组织中端粒酶的表达,发现肿瘤组织中端粒酶的表达显著高于正常组织。
现已知在95%左右的妇科恶性肿瘤中,有60%左右的癌前病变或交界性肿瘤中存在端粒酶活性,而仅3.75 %的良性病变表达出了端粒酶活性。
这些结果说明,端粒酶的活化普遍存在于妇科恶性肿瘤中,并且参与了肿瘤的早期发生,可以认为是肿瘤恶变或恶性肿瘤发生的征兆【6】。
对于端粒酶活性引起的肿瘤发生,近年来也研究了一些抑制剂。
以反义核苷酸抑制剂为代表的端粒酶抑制剂,能通过各自不同的作用机制抑制端粒酶的活性,从而最终致使肿瘤细胞的凋亡,以达到肿瘤治疗的作用【7】。
3.3端粒与心血管疾病:
目前有多项研究表明,端粒与一些年龄相关的疾病(如动脉粥样硬化、冠心病)的形成有关。
Huzen等的研究指出,在活体中,淋巴细胞端粒长度在动脉粥样硬化患者较无动脉粥样硬化对照组短,动脉粥样硬化斑块端粒的长度同斑块的特点及发展与心血管事件相关。
Scott等的研究也提示,心血管疾病中的端粒缩短可
能是冠心病的新的、独立的危险因素。
在近期的研究中也提出,发展成动脉粥样硬化血管组织中的端粒长度,较没有发展成动脉粥样硬化的血管组织中的端粒长度短。
以上多项研究均表明,动脉粥样硬化的发生、发展与端粒长度的变化有明显关系。
目前国内尚无外周血白细胞端粒长度及酶活性与老年痴呆或AD关系的报道,诸多与端粒和衰老有关的基础性问题还有待探讨【8】。
4.端粒及端粒酶的检测方法:
端粒及端粒酶与正常细胞生长调控及恶性肿瘤形成机制密切相关。
于是端粒酶活性的检测对于肿瘤形成机制、诊断及预后评价就具有了重要意义。
目前,端粒酶活性的检测方法主要有以下几种:
端粒重复序列延伸法(Telomere extension assay):它与DNA聚合酶分析方法相似,如:把核酸提取物、代表脊椎动物端粒重复序列的单链DNA前体(TYAGGG)和放射标记的磷酸脱氧核糖一起孵育,然后通过放射自显影检测凝胶上新添加的DNA重复序列。
端粒重复序列扩增法(TRAP)及其改良方法:在TRAP法中,端粒酶反转录出一段DNA片段,经过PCR扩增出大量的含不同重复数目的TFAGGG片段,进行放射自显影或不加同位素,直接电泳。
在电泳图谱中显示出等距离的相差6bp的梯形图谱。
每104个细胞中只有1个永生化细胞,TRAP就能检测到端粒酶活性。
对该法进行改进,在端粒延伸与PCR扩增时,把CX引物作为生物素标记引物(Bio—CX),扩增后加亲合素标记的荧光微球,最后直接用液闪仪计数【9】。
5.展望
端粒和端粒酶的发现和研究,对人类疾病的研究作出了重要贡献。
研究显示,端粒酶活性被激活,可维护端粒的长度,细胞将会延缓衰老,避免癌变。
此外,。