维管系统介绍
植物体内有机物的合成、代谢、运输与分配

植保051班 3号 叶良妹
一、植物体内有机物的合成
原料:CO2、H2O、光照 产品:直接产物有糖类,包
括蔗糖和淀粉 间接产物有脂肪,蛋白质等。
有机物物的合成来自光合作用
绿色植物吸收光能,同化二氧化碳和水,制造有机物并释放 氧气的过程,称为光合作用
1、光合作用的三大步骤:
糖在无氧状态下分解成丙酮酸的过程,又 称EMP途径。糖酵解过程在细胞原生质内 进行
二、三羧酸循环(TCAC) z 糖酵解的产物丙酮酸在有氧条件下进入
线粒体逐步氧化分解,形成水和二氧化碳 的过程
三、磷酸戊糖途径(PPP) PPP是细胞存在。由于 磷酸戊糖是该途径的中间产物,故该途径 称为磷酸戊糖途径
的O2分子数或固定的 CO2分子数 光系统Ⅰ( PSⅠ)
其反应中心色素分子吸收700 nm的红光并 发生光化学反应。 PSⅠ颗粒较小,存在于 间质片层和基粒的非垛叠区。它与 NADPH 的生成有关
光系统Ⅱ( PSⅡ) 其反应中心色素分子吸收 680nm的红光
并发生光化学反应。PSⅡ颗粒较大,存在 于基粒片层的垛叠区它与 H2O的氧化即氧 气的释放有关
z 呼吸作用的场所:线粒体
主要糖类的代谢
z 淀粉(叶绿体内)、蔗糖(细胞质)经过 水解成葡萄糖,又经过糖酵解生成丙酮酸 若经过无氧呼吸则生成酒精或乳酸和少量 的ATP;若经过有氧呼吸则生成CO2、H2O 和大量的ATP。
三、植物体内有机物的运输和 分配
有机物的运输
1、有机物质运输的途径 z 维管系统是专门执行运输功能的输导组织,由
韧皮部和木质部组成,贯穿植物全身 z 有机物的运输途径是由韧皮部担任,主要运输
组织是韧皮部里的筛管和伴胞。
浅谈植物体形态结构,功能与其环境的适应性

浅谈植物体形态结构,功能与其环境的适应性伊宁市第八中学生物教研组:帕提曼.玉山结构与功能相统一的观点包括两层意思:一是有一定的结构就必然有与之相对应功能的存在;二是任何功能都需要一定的结构来完成。
例如叶的表皮是无色透明的,表皮细胞排列紧密,向外一面的细胞壁上有透明而不易透水的角质层。
表皮的这种结构的存在,即利于阳光透过,又能防止叶内水分过多地散失,还能保护叶内部不受外来的伤害;而阳光透入,防止水分散失,保护叶内组织,又需要一定的结构来完成,这就是表皮。
一、分生组织位于植物的生长部位,具有持续或周期性分裂能力的细胞群,称为分生组织。
分生组织的细胞排列紧密,细胞壁薄,细胞核相对较大,细胞质浓,细胞器丰富。
根据分生组织在植物体内的位置不同,可将分生组织分为顶端分生组织、侧分生组织和居间分生组织三类;此外,也可根据来源将分生组织分为原分生组织、初生分生组织和次生分生组织三类。
原分生组织位于根尖和茎尖的顶端,由一群胚性的原始细胞组成,能长期地保持分裂能力。
初生分生组织由原分生组织的细胞分裂而来,一方面初生分生组织的细胞可继续分裂,另一方面开始初步分化,逐渐向成熟组织过渡。
初生分生组织有原表皮、基本分生组织和原形成层三种。
次生分生组织就是侧分生组织,由已分化成熟的薄壁组织细胞恢复分裂能力转变而来,有维管形成层和木栓形成层两类。
细胞的特点:外形细小,近于等径,细胞排列紧密、无胞间隙、细胞核大、薄壁、液泡细小、多、细胞质浓厚、生活力强。
1.顶端分生组织:顶端分生组织存在于根尖和茎尖的分生区部位,由短轴或近于等径的胚性细胞构成,细胞排列紧密,能较长时期地保持旺盛的分裂能力。
2.侧分生组织:侧分生组织包括维管形成层和木栓形成层,它分布于植物体的周围,平行排列于所在器官的边缘。
侧分生组织细胞的形状为长轴形和等径状,其功能是使植物体变粗。
3.居间分生组织:居间分生组织分布于成熟组织之间,进行一段时间的分裂活动后失去分裂能力,完全分化为成熟组织。
分生组织

㈡成熟组织
10
11
B. 气孔
12
C. 表皮的附属物:表皮毛
13
2) 次生保护组织:周皮
14
15
小 结:
保护组织覆盖在器官表面,有表皮和周皮两种。表皮最 显著的特征是细胞外壁角化并形成角质层,为初生保护组 织。周皮是木栓层、木栓形成层、栓内层的总称,其中的 木栓层细胞壁高度栓化,不透水,不透气,是代替表皮的 次生保护组织。
导组织的概念,应该是担负物质长途运输的组织,而不是“担负物 质运输的组织”。因为薄壁组织也能担负短距离的物质运输。 39
(二)六类组织中应重点掌握输导组织
在六类组织中,分布最广泛、体积最大的是薄壁组织和输导组织
。但薄壁组织 结构简单,容易掌握,而输导组织结构复杂,不易理解,特别是导
管和导管分子、筛管和筛管分子、导管和管胞的区别,不好掌握。
有不同方式的增厚并木化。根据侧壁增厚方式导管可分为环纹导管、
管代替其行使输导功能。
22
管胞
是一种狭长而两端斜尖的细胞,与导管的
主要区别是端壁不形成大穿孔而为具缘纹孔,彼此 不连接成长管。在蕨类植物和裸子植物中,管胞是 唯一的输水结构,在被子植物中,管胞和导管同时 存在。 筛管和伴胞 筛管由长管状的生活薄壁细胞纵向 连接而成。每一个细胞称为筛管分子,分化成熟的 筛管分子没有细胞核,其端壁特化成为筛板,筛板 上有许多筛孔。通过筛孔连系着上下两个筛管分子 的细胞质索称为联络索。较老的筛管由于胼胝体的 形成而暂时或永久地丧失输导能力。紧贴筛管分子 旁边有 1至数个小型的薄壁细胞,与筛管分子是由
维管束

维管束
叶植物中。位于后生木质部与后生韧皮部之间的原形成层,如能继续发育为维管形成层(即束中形成层),并与维管束之间薄壁组织产生的形成层(束间形成层)相接,向外产生次生韧皮部,向内分化次生木质部,这种能继续生长的维管束,称为无限维管束,常见于裸子植物和木本双子叶植物中。
在初生植物体内,维管束相互连接、错综复杂。茎中的维管束进入叶子里,是通过茎皮层到叶柄基部的一段维管束,即叶迹。同样,侧芽发生后,由枝迹将茎的维管束与侧枝维管束相互连接。根中的维管组织的排列与茎不同,它们之间的联系是通过一个过渡区,即由根的木质部束与韧皮部束交替排列的方式,逐渐转变成茎中木质部与韧皮部内外排列成维管束的形式。总之,根通过过渡区与茎维管束相连,茎再通过枝迹与叶迹同侧枝与叶子的维管束连接,这样,在初生植物体中构成一个完整的维管组织系统,主要起输导和支持作用。
维管束
维管束(vascular bundle)是维管植物(蕨类植物、裸子植物和被子植物)的叶和幼茎等器官中,由初生木质部和初生韧皮部共同组成的束状结构。维管束彼此交织连接,构成初生植物体输导水分,无机盐及有机物质的一种输导系统——维管系统,并兼有支持植物体的作用。
分布
组成
பைடு நூலகம்类型
外韧维管束
蕨类植物(精)

蕨类植物的形态特征
• 蕨类植物的叶子中,有仅进行光合作用的 叶,称为营养叶或不育叶;也有些叶子的 主要作用是产生孢子囊和孢子,称为孢子 叶或能育叶。 • 有些蕨类的营养叶和孢子叶是不分的,而 且形状相同,称同型叶;也有孢子叶和营 养叶形状完全不相同的,称为异型叶。 • 在系统演化过程中,同型叶是朝着异型叶 的方向发展的。
蕨类植物的形态特征
• 孢子萌发后,形成为配子体。配子体又称原叶体,小型, 结构简单,生活期较短。 • 极大多数蕨类的配子体为绿色、具有腹背分化的叶状体, 能独立生活,在腹面产生颈卵器和精子器,和苔类植物相 似,但精子多鞭毛。像卷柏和水生蕨类等异孢种类,配子 体是在孢子内部发育的,已趋向于失去独立性的方向发展。 • 配子体产生的精子和卵,在受精时还不能脱离水的环境。 受精卵发育成胚,幼胚暂时寄生在配子体上,长大后配子 体死亡,孢子体即行独立生活。 • 蕨类植物都有世代交替。少数为配子体占优势 ,而大多 数为孢子体世代占优势。
真蕨亚门:薄囊蕨纲:水龙骨目
• 绝大多数为陆生或附生种类。孢子囊聚生成各式 孢子囊群。孢子同型。为蕨类植物门中最大的1目。 • 茎为根状茎,有分枝,横卧地下,在土壤中蔓延 生长,生不定根并被有棕色的茸毛。叶每年从根 状茎上生出并钻出地面,有长而粗壮的叶柄。叶 片大,幼叶拳曲,成熟后平展,呈三角形,2—4 回羽状复叶。 • 孢子囊群生于孢子叶的背面,沿叶缘生长。囊群 有1条纵裂的环带
蕨类植物的形态特征
• 在蕨类植物中,存在着孢子体不经过孢子而产生配子体的 现象,称为无孢子生殖。 • 配子体也可以不经过配子的结合,而直接产生孢子体的, 这种现象称为无配子生殖。 • 无配子生殖在蕨类植物中相当普遍,有时在1种植物中, 同时可具有无配子生殖和无孢子生殖的现象,许多无孢子 生殖产生的配子体,能正常地产生精子器和颈卵器,这种 配子体的染色体数目为2n,由此产生的配子配合后,形成 了4n的孢子体,这种四倍体(tetraploid)的孢子体,也可 诱导形成四倍的无孢子生殖的配子体。真蕨类中这种诱导 成的无孢子生殖的二倍体(diploid)的配子体,也可以与 单倍染色体(hapoid)的配子体相互交配,由此产生了三 倍体(triploid)的孢子体。
普通生物学名词解释

第一章绪论:生物界与非生物界1、生物圈(biosphere)2、熵(en tropy)3、耗散结构(dissipative structure4、应激性(irritability)5、适应:包含有两方面的涵义6、稳态(homeostasis):7、生物多层次结构8、五界系统9、双名法(binomial nomendature)第二章生命的化学基础1、同位素示踪2、必需元素(essential element)3、生物大分子(macromolecule)4、多聚体(polymer)5、糖类&非必需氨基酸7、必需氮基酸8、蜡(wax)9、固醇(steroid)10、氨墓酸(ami no acid)11、肽键(peptide bond)12、肽(peptide)和多肽(polypeptide)13、蛋白质的一级结构14、蛋白质的二级结构15、蛋白质的三级结构16、蛋白质的四级结构17、蛋白质的变构作用⑻losteric effect)18、蛋白质的变性(denaturation)19、核昔酸20、ATP。
第三章:细胞结构与细胞通讯1、细胞学说2、细胞质(cytplasm )3、生物膜(biomembrane )4、核膜(nuclear envelope )5、核纤层(nuclear lamina)&染色质(chromatin )7、常染色质(euchromatin )8、异染色质(heterochromatin )9、染色体(chromosome )10、组蛋白(histone)12、高尔基复合体(Golgi complex )13、质体(plastid )14、液泡15、细胞连接(cell junctions)16、细胞通讯第四章:细胞代谢1、代谢(metabolism)2、势能(potential energy)3、热力学(thermogynamics)4、自由能(free energy)5、活化能(activation energy)。
植物学整理

植物学总结第一章绪论植物细胞1,植物细胞的结构(纹孔是细胞间物质交流的区域)2,具体的起源,内共生学说3,叶绿体基因组的特性4,植物细胞的分化与死亡(物理死亡和凋亡)第二章植物组织1,植物组织的概念胞间连丝是组织形式的物质基础2,植物组织的产生和演化是长期适应自然环境和自然选择的结果3,植物组织的类型分生组织的概念类型发育时期和活动结果成熟组织的类型,薄壁组织、机械组织、保护组织(表皮和周皮的区别)、疏导组织(包括导管管胞筛管筛胞)、维管束、分泌组织重点1,以苔藓和蕨类对比简述孢子体配子体的生活史2,定距式检索表(以根茎叶花果实种子为顺序挑选性状对比明显的,例如草本植物和木本植物,裸子植物和被子植物,有地上茎和无地上茎,常绿和落叶,叶对生和叶互生,叶披针形和叶心形,头状花序和总状花序,唇形花和舌状花)罂粟(有乳汁侧膜胎座蒴果),垂柳,红松,一串红,向日葵,蒲公英,杏3,以松鼠植物为例,简述松柏类(裸子植物)的生活史4,简述云冷山的区别和重要特征5,【重】花的演化意义(例如两性花配子的形成,以及具有雌蕊子房发育成果实的意义)6,【重点】双受精现象的意义7,APG系统的概念和优势,以及它的全称被子植物中发生以及英文名Angiosperm Phylogeby Group8,协同进化和趋同进化9,以无油樟为例简述现生最古老的被子,植物类群的特征(现身被子植物最基部类群)10,从花序和果实类型出发的名词解释(例如聚花果和聚合果的差异)11,请问如何分别小枝和复叶12,以木兰科为例简述其原石特征的花托伸长(壳斗科总苞)13,毛莨科性状特征14,简述一个类群的初生次生结构……第一章绪论植物细胞1,植物细胞的结构(纹孔是细胞间物质交流的区域)木质部或韧皮部的结构及其功能。
木质部:由导管、管胞、木纤维和木薄壁细胞构成。
功能是输送水和无机盐。
韧皮部:是输送有机营养物质的通道,由筛管或筛胞、伴胞、韧皮薄壁细胞和韧皮纤维构成。
第三章 第四节 营养器官间的相互关系及营养器官的变态

叶迹和叶隙、枝迹和枝隙的图解
• 3.营养器官在植物生长中的相互影响
• (1)地下部分与地上部分的相互关系 • ①种子萌发时是先长根后长苗。 • ②根系从地上部分,特别是叶获取有机养料; 根系向地上部分提供水、无机盐和根系制造的某 些氨基酸、维生素及其它生理活性物质; • ③地上部分从地下部分获取水和无机盐,还有 一些生理活性物质,如氨基酸、维生素、一些激 素等;地上部分向地下部分提供有机养料,如葡 萄糖、氨基酸、维生素、生长素等。 • “根深叶茂,本固枝荣”,说明根与茎叶之间 是相互依存、相互制约的辩证关系。
葡萄的茎卷须
黄瓜的茎卷须
• (3) 叶状茎(叶状枝)
• • • • • • • • • 茎变态为叶状,绿色扁平,能进行光合作用。 假叶树 侧枝变为叶状枝,叶退化为鳞片状,叶腋内可生小花。 因鳞片极小,不易辩认,常被人们误认为叶上开花。 天门冬 叶腋内也产生叶状枝,而叶极小。 竹节蓼 叶状枝极显著,叶小或全缺。 再“奇怪”的形态,即使是从未见过,也应利用植物 形态的基本知识进行分析,无论变态为什么样子,茎总是 具有茎的形态特征。 譬如,叶上长花的问题,可以从植物学角度去分析, 叶上会开花吗?什么地方会开花?只有茎、枝上会开花, 那么,由此,我们可以肯定,这个着生花的结构长得再像 “叶”,它也是茎。
• (一)变态根
• 变态根主要有贮藏根、气生根、寄生根三种类型
•
• • • • •
1.贮藏根
存贮养料,肥厚多汁,形状多样,常见于二年生 或多年生的草本双子叶植物。 贮藏根是越冬植物的一种适应,所贮藏的养料可 供来年生长时的需要,使根上能抽出枝来,并开花 结果。 根据来源可分为肉质直根和块根两大类。 肉质直根:主要由主根发育成。一株上仅有一个 肉质直根。 块根:主要由不定根或侧根发育而成。一株上可 形成多个块根。
(完整版)植物生活史小结
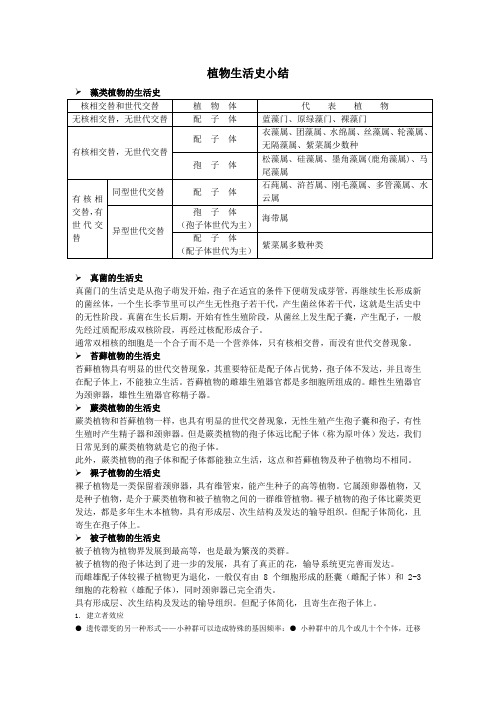
植物生活史小结➢真菌的生活史真菌门的生活史是从孢子萌发开始,孢子在适宜的条件下便萌发成芽管,再继续生长形成新的菌丝体,一个生长季节里可以产生无性孢子若干代,产生菌丝体若干代,这就是生活史中的无性阶段。
真菌在生长后期,开始有性生殖阶段,从菌丝上发生配子囊,产生配子,一般先经过质配形成双核阶段,再经过核配形成合子。
通常双相核的细胞是一个合子而不是一个营养体,只有核相交替,而没有世代交替现象。
➢苔藓植物的生活史苔藓植物具有明显的世代交替现象,其重要特征是配子体占优势,孢子体不发达,并且寄生在配子体上,不能独立生活。
苔藓植物的雌雄生殖器官都是多细胞所组成的。
雌性生殖器官为颈卵器,雄性生殖器官称精子器。
➢蕨类植物的生活史蕨类植物和苔藓植物一样,也具有明显的世代交替现象,无性生殖产生孢子囊和孢子,有性生殖时产生精子器和颈卵器。
但是蕨类植物的孢子体远比配子体(称为原叶体)发达,我们日常见到的蕨类植物就是它的孢子体。
此外,蕨类植物的孢子体和配子体都能独立生活,这点和苔藓植物及种子植物均不相同。
➢裸子植物的生活史裸子植物是一类保留着颈卵器,具有维管束,能产生种子的高等植物。
它属颈卵器植物,又是种子植物,是介于蕨类植物和被子植物之间的一群维管植物。
裸子植物的孢子体比蕨类更发达,都是多年生木本植物,具有形成层、次生结构及发达的输导组织。
但配子体简化,且寄生在孢子体上。
➢被子植物的生活史被子植物为植物界发展到最高等,也是最为繁茂的类群。
被子植物的孢子体达到了进一步的发展,具有了真正的花,输导系统更完善而发达。
而雌雄配子体较裸子植物更为退化,一般仅有由8个细胞形成的胚囊(雌配子体)和2-3细胞的花粉粒(雄配子体),同时颈卵器已完全消失。
具有形成层、次生结构及发达的输导组织。
但配子体简化,且寄生在孢子体上。
1. 建立者效应●遗传漂变的另一种形式——小种群可以造成特殊的基因频率:●小种群中的几个或几十个个体,迁移到它处定居下来,与原种群隔离开来,自行繁殖形成新的种群;有些等位基因没有带出来,导致新种群与原种群的基因频率的差异;●新种群的基因频率取决于建立者(定殖者)——分离出来的几个或几十个个体●意义:通过自然选择,有可能形成新物种。
园林技术班《植物学》A卷和B卷试题及答案

园林技术班期末考试《植物学》试题(A卷)90分钟(一共6页)(答案写在答卷内容后)学号姓名班级分数一、名词解释(20分)1 根的次生生长,2 心皮,3 花序,4 异形叶性,5世代交替,6 维管系统,7 原生质体,8 种(物种),9 叶迹,10 树皮二、填空题(25分,共50空,每空0.5分)1 种子留土萌发和出土萌发在萌发过程中的主要区别,在于。
2 常见的地下茎有可分为、、和四类。
3 生物的六界系统是指、、、、和界。
4 植物细胞的后含物类型有、、及其有关的物质,还有成结晶的无机盐和其他有机物。
5 成熟组织可按功能分为、、、和。
6 维管植物的主要组织系统可分为、、三种。
7 种子植物营养器官是、、、繁殖器官是、和。
8 侧根起源于。
9 种子植物中和真菌共生的根称为。
10 就叶片的形状来讲,一般指整个单叶叶片的形状,但有时可指、、的形状,可作为识别植物和分类的依据之一。
11 叶序主要有三种基本类型、和。
12 同类器官,长期进行不同的生理功能,以适应不同的环境,就导致功能不同,形态各异,成为器官;反之,相异的器官,长期进行相似的生理功能,以适应某一外界环境,就导致功能相同,形态相似,成为器官。
13 植物的繁殖方式可以区分为三种类型:、和。
14 胚珠着生的心皮壁上,往往形成肉质突起,称为。
15 被子植物的受精作用完成以后,便发育为种子,发育为果实。
16 果实的类型中,称为真果,称为假果。
17 植物的学名是由、和组成。
18 花是,并且进一步发展成为果实和种子。
三、单项选择题(12分)1在绿藻门的下述特征,与高等植物特征相同。
A、叶绿素a和bB、尾鞭型鞭毛C、接合生殖D、光合产物为真淀粉2被子植物生活史中,两个世代交替的转折点是。
A、减数分裂B、受精作用C、花粉和胚囊发育成熟D、胚形成E、种子萌发F、A和BG、B和E3马铃薯(土豆)的食用部分是。
A、块茎B、块根C、球茎D、根状茎4在进行,为了促进生根,常使用一些生长素。
微管微缆系统技术简介

微管微缆系统技术简介概述微缆气吹(JETnet)技术是荷兰NKF光缆公司全球首创的光缆气吹布放技术,采用管道吹缆专用微型管道光缆气吹敷设工艺(即母管+子管+微缆敷设技术)。
该技术利用气吹敷缆的方法,先将微管吹进已敷设的母管中,然后根据客户需求,可分批次将微缆吹进微管中。
该技术可广泛应用于骨干网、城域网、接入网中。
一、JETnet 系统结构分析:图1:JETnet系统结构示意图JETnet 的基本组件就是“母管—子管—光缆”。
母管一般采用HDPE/硅芯管,也可采用PVC管,子管用HDPE材料挤制而成,光缆为微型中心管式光缆。
(一)母管:母管直径HDPE 母管有φ25mm、φ32mm、φ40mm、φ50mm、φ63mm 几个规格。
母管的功能是引导和保护含有微缆的微管不被损坏。
为了确保微管顺利吹入母管,母管必须能够承受必要的压力。
同时母管内壁必须光滑、干燥和清洁,可以减少微管敷设时和母管之间的摩擦系数。
(二)微管:子管直径有φ7mm和φ10mm两种规格。
1.微管的类型:微管是气吹系统的一个重要的组成部分。
在微缆敷设前微管必须先敷设到目的地,并且要求一次性敷设到位。
因为纽绞的问题,在一根母管内的微管是不能分批敷设的。
微管在制作工艺等方面具有较高的技术含量,其内壁采用具有同步挤压的永久性固体润滑硅芯层,选择纵向导气槽(凹槽)结构,进一步降低了摩擦系数。
目前国外的微管按结构区分大约有三种:(1)集束管:有点象我们市话管道上常用的蜂窝管,是由微管束和外护套两部分在厂家组合而成,但是微管和微管之间是不连接的,可以有相对的位移。
集束管的优点在于管道的密度高,可以在有效的空间内容纳最多的微管。
缺点是灵活性较差,管道连接不方便。
(2)微管束:它是利用微管超级气吹机 (SUPERJET MD)根据母管的内径尺寸将一定数量的微管束吹进已经敷设好的母管内,根据气吹原理,微管束不能填充整个管道,因此微管和母管之间需要有一定的空间,这种空间同时增强了微管的机械强度,保证了母管在发生轻度变形时,不会直接影响到微管。
蕨类植物

鉴于上述情况,本教材仅对其中种类较多,分布较广,具有代表 性的石松亚门、楔叶亚门和真蕨亚门的代表种类进行重点介 绍。
一、石松亚门的代表种类
石松亚门的主要识别特征:
1、孢子体小型,直立或匍匍; 2、小型叶,鳞片状,螺旋状排列,或呈4行排列; 3、孢子叶密集枝端形成孢子叶球; 4、孢子同型或异形; 5、配子体微小,生地下与真菌共生,或部分绿色; 6、精子长卵形,具2条鞭毛。
地下生); 4)配子体生活期很长,部
分或全部靠菌根营养; 5)精子器和颈卵器同生于
配子体的上面,并埋在 组织中。
3、配子体及有性生殖器官的特征:
4、胚的形成特征:
受精 合子 胚
第一次分裂
第二次分裂
第四次分裂
幼胚
特点:合子的第一次分裂为横裂,胚胎分化较迟缓 。
(二)卷柏目的特征:
代表植物卷柏属(Selaginella)
10、孢子的产量及其形 态
11、原叶体(配子)的 形态结构
12、有性生殖器官的位 置和构造
16、植物地理学 17、古植物学
第三节 蕨类植物的代表种类
最早的蕨类植物化石发现于距今约4亿年前的志留纪晚期。现 存蕨类植物约12000种。中国有2600种,其中云南即有1000余 种,享有“蕨类植物王国”之称。
厚孢子囊5-10个 生于孢囊柄六 角形盘状体下 面
极少为厚孢子囊, 绝大多数为薄孢 子囊。孢子囊集 聚成囊群,生于
孢子叶背面或背
缘,多具囊群盖
孢 子
孢子同型
有的孢子同 型有(的石 孢松子)异,孢子异型 型(卷柏)
孢子同型,具 弹丝
竹原纤维-维管束的组成及其类型

维管束的组成及其类型维管束(vascular bundle) 是植物体内的一种束状结构,组成植物的输导系统,同时对植物器官有支持作用。
是由原形成层分化产生的几种组织共同构成的复合组织。
植物根﹑茎﹑叶內运输水分的细胞上下排列成管状,运输养分的细胞也上下排列成管状,两者聚集成束状,称为维管束,叶部的维管束称为叶脉。
维管束运输水分的构造叫做木质部,运输养分的构造叫做韧皮部。
维管系统:一株植物或一个器官的全部维管组织称为维管系统。
木质部输送水分:土壤中的水分及矿物质由根毛进入植物体內,到达根部的维管束后,由维管束的木质部运送到茎及叶等各部分的细胞。
植物的根部有許多细丝状的根毛,是由根部表皮细胞向外突出所形成的构造,与土壤直接接触;根毛的数目相当多,可以增加根部吸收面积。
韧皮部输送养分:植物叶所产生的养分由叶部维管束內的韧皮部运送到茎及根等各部分的细胞,供植物细胞利用或储藏。
储藏的养分在植物需要時可以再分解,並由韧皮部运输到所需要的地方。
例如甘薯的养分大部分储藏在根內,而使根部膨胀形成块根;以块根繁殖時,储藏的养分分解并运输到芽,供芽生长利用。
维管束有多种类型:⑴无限维管束:韧皮部与木质部之间有束中形成层。
⑵有限维管束(闭锁性维管束):韧皮部与木质部之间无束中形成层。
⑶外韧维管束:韧皮部排列在木质部的外方。
⑷双韧维管束:木质部的内外方均有韧皮部。
⑸周木维管束:木质部围绕着韧皮部呈同心排列。
⑹周韧维管束:韧皮部围绕着木质部呈同心排列。
双子叶植物多具有无限外韧维管束,有些具有双韧维管束。
单子叶植物具有有限外韧维管束。
维管束及其类型2010-8-3 17:55【大中小】【我要纠错】维管植物-维管系统由许多维管束组成。
韧皮部-筛管、伴胞、筛胞、韧皮薄壁细胞、韧皮纤维木质部-导管、管胞、木薄壁细胞与木纤维维管植物由三种系统组成,皮组织系统、维管组织系统和基本组织系统。
1、有限外韧维管束——韧外、木内、中无形成层。
单子叶茎2、无限外韧维管束——韧外、木内、中有形成层。
被子植物的器官系统详解

木栓形成层的分裂产生:向内形成栓内层
向外形成木栓层
细胞分裂方向:
1.切向分裂(平周分裂):横切面上,细胞分裂方向平行于周 径,结果增加细胞层次。
2.径向分裂(垂周分裂):横切面上,细胞分裂方向垂直于周 径,结果扩大器官或组织的周径。
3.横向分裂:纵切面上,细胞分裂方向垂直于纵轴,结果使器 官或组织伸长。
小麦茎
2.禾本科植物茎的结构
表皮:有气孔器(保卫细胞哑铃型)
机械组织:厚壁组织
基本组织:有或无髓腔 维管束:有限维管束,无形成层
维管束
表皮
玉米茎
基本 组织
(四)双子叶植物茎的次生结构
周皮 木栓形成层 皮层
初生韧皮部 次生韧皮部
维管形成层
次生木质部
初生木质部
双子叶植物茎次生结构
木射线
边材 心材
多年生木本植物茎结构
根冠:根尖的顶端,起保护作用 分生区:具分裂能力,使根生长 伸长区:细胞逐渐分化并伸长 成熟区(根毛区):细胞生长停止,
形成各种组织
(三)根的初生结构
由初生分生组织生长所形成的结构称初生结构。
1.双子叶植物根的初生结构
表皮:具根毛 皮层:外皮层
中皮层 内皮层:凯氏带
维管柱(中柱):
[1]中柱鞘:中柱最外层的1-2层薄壁细胞 [2]初生韧皮部:筛管、伴胞
次生韧皮部
维管形成层
次生木质部
初生木质部
(初生结构部分) (次生结构部分)
周皮
二、茎
功能:(1)运输和支持; (2)营养、贮藏; (3)繁殖
(一)茎的形态 ◇有节和节间之分 ◇在节上着生叶和芽 ◇着生叶和芽的茎为枝条
顶芽
植物学形态解剖学名词

植物学(上)名词术语中英文对照植物学(Botany)植物形态学(Plant morphology)植物解剖学(plant anatomy)原生质体(protoplast)细胞壁(cell wall)显微结构(microscopic structure)亚显微结构(submicroscopic structure)超微结构(ultramicroscopic structure)质体(Plastid)叶绿体(chloroplast)类囊体(thylakoid)基粒(granum)基粒间膜(基质片层,fret)基质(stroma或matrix)有色体(或称杂色体,chromoplast)白色体(leucoplast)造油体(elaioplast)前质体(proplastid)液泡(vacuole)液泡膜(tonoplast)细胞液(cell sap)纹孔(pit)胞间连丝(plasmodesmata)后含物(ergastic substance)淀粉粒(starch grain)淀粉体(amylop1ast)脐点(hilum)拟晶体(crystalloid)糊粉粒(aleuronegrain)糊粉层(aleurone layer)胞质分裂(Cytokinesis)成膜体(phragmoplast)细胞板(cellplate)微管周期(microtubule cycle)细胞分化(cell differentiation)反分化(或脱分化dedifferentiation)组织(tissue)分生组织(meristematic tissue或meristem)顶端分生组织(apical meristem)侧生分生组织(lateral meristem)居间分生组织(intercalarymeristem)形成层(cambium)木栓形成层(cork cambium或phellogen)原分生组织(promeri-stem)初生分生组织(primary meristem)次生分生组织(secondary meristem)保护组织(protective tissue)薄壁组织(parenchyma)机械组织(mechanical tissue)输导组织(conducting tissue)分泌结构(secretory structure)表皮(epidermis)周皮(periderm)气孔(stoma)皮孔(lenticel)保卫细胞(guard cell)吸收组织(absorptive tissue)根毛(root hair)木栓(phellem或cork)栓内层(phelloderm)同化组织(assimilating tissue)储藏组织(storage tissue)储水组织(aqueous tissue)通气组织(aerenchyma)传递细胞(transfer cell)厚角组织(collenchyma)厚壁组织(sclerencnyma)石细胞(sclereid或stone cell)纤维(fiber)木质部(xylem)韧皮部(phloem)管胞(tracheid)导管分子(vesselelement或vesselmember)穿孔(perforation)导管(vessel)筛管分子(sieve-tube element或sieve-tube member)筛管(sieve tube)筛孔(sieve pore)筛孔(sieve pore)筛板(sieve plate)原生质联络索(connecting strand)胼胝质(callose)筛域(sive area)伴胞(companioncell)胼胝体(callus)筛胞(sieve cell)腺表皮(glandular epidermis)腺毛(glandular hair)蜜腺(nectary)排水器(hydathode)吐水(guttation)水孔(waterPore)通水组织(epithem)分泌细胞(secretorycell)分泌腔(secretorycavity)分泌道(secretorycanal)乳汁管(laticifer)无节乳汁管(nonar-ticulatelaticifer)有节乳汁管(arti-culatelaticifer)组织系统(tissue system)皮组织系统(dermal tissue system)维管组织系统(vascular tissue system)基本组织系统(fundamental tissue system或ground tissue system)皮系统(dermal system)维管系统(vascular system)基本系统(fundamental system或ground system)种子(seed)胚(embryo)胚乳(endosperm)种皮(seed coat,testa)外胚乳(perisperm)胚根(radicle)胚芽(plumule)胚轴(hypocotyl)子叶(cotyledon)种脐(hilum)种阜(caruncle)种脊(raphe)有胚乳种子(albuminousseed)无胚乳种子(exalbuminous seed)胚芽鞘(coleoptile)胚根鞘(coleorhi- za)盾片(scutellum)外胚叶(epiblast)种子萌发(seed germination)子叶出土的幼苗(epigaeous seedling)子叶留土的幼苗(hypogaeous seedling)器官(organ)营养器官(vegetative organ)根(root)根系(root system)主根(main root)直根(tap root)初生根(primaryroot)侧根(lateral root)次生根(secondaryroot)不定根(adventitiousroot)定根(normal root)种子根(seminal root)直根系(taprootsystem)须根系(fibrousrootsystem)原始细胞(initialcell)不活动中心(或称静止中心,quiescentcentre)根尖(roottip)根冠(root cap)分生区(meristematiczone)伸长区(elongationzone)成熟区(maturationzone)维管柱(vascular cylinder)皮层(cortex)切向分裂(弦向分裂,tangentialdivision)平周分裂(periclinalkivision)径向分裂(radialdivision)横向分裂(transversedivision)垂周分裂(anticlinaldivision)根毛区(roothairzone)初生生长(primary growth)初生组织(primary tissue)初生结构(primary structure)根被(velamen)外皮层(exodermis)内皮层(endoder-mis)凯氏带(Casparian strip)通道细胞(passage cell)中柱鞘(pericycle)髓(pith)初生木质部(primary xylem)初生韧皮部(primary phloem)外始式(exarch)原生木质部(protoxylem)后生木质部(metaxylem)木质部脊(xylem ridge)二原型(diarch)三原型(triarch)四原型(tetrarch)五原型(pentarch)六原型(hexarch)多原型(polyarch)原生韧皮部(protophloem)后生韧皮部(meta-phloem)根原基(root primordium)内起源(endogenousorigin)形成层环(cambium ring)木射线(xylemray)韧皮射线(phloemray)维管射线(vascularray)木栓形成层(phellogen或cork cambium)栓内层(phelloderm)木栓(phellem或cork)周皮(periderm)共生(symbiosis)根瘤(root nodule)菌根(mycorrhiza)外生菌根(ectotrophic mycorrhiza)内生菌根(endotrophic mycorrhiza)内外生菌根(ectendotrophicmycorrhiza)茎(stem)节(node)节间(internode)枝或枝条(shoot)叶痕(leafscar)维管束痕(bundle scar,简称束痕)芽鳞痕(bud scalescar)芽(bud)枝芽(branch bud)叶芽(leafbud)花芽(floralbud)叶原基(leaf primordium)腋芽原基(axillary bud primordium)侧枝原基(lateral branch primordium)枝原基(branchprimordium)芽轴(bud axis)定芽(normalbud)不定芽(adventitiousbud)顶芽(terminal bud)腋芽(axillary bud)侧芽(lateral bud)副芽(accessory bud)叶柄下芽(subpetiolar bud)裸芽(naked bud)被芽(protected bud)鳞片(scale)芽鳞(bud scale)鳞芽(scaly bud)混合芽(mixed bud)活动芽(active bud)休眠芽(dormant bud)潜伏芽(latent bud)直立茎(erect stem)缠绕茎(twining stem)攀援茎(climbing stem)匍匐茎(creeping stem)纤匍枝(runner)单轴分枝(monopodial branching)合轴分枝(sympodial branching)假二叉分枝(falsedichotomous branching)二叉分枝(dichotomousbranching)分蘖(tiller)原表皮(protoderm)基本分生组织(ground meristem)原形成层(procambium)生长点(growing point)生长锥(growing tip)茎端(stem apex)根端(root apex)枝端或苗端(shoot apex)茎尖(stemtip)根尖(root tip)组织原学说(histogen theory)表皮原(dermatogen)皮层原(periblem)中柱原(plerome)原套-原体学说(tunica-corpus theory)原套(tunica)原体(corpus)细胞学分区概念(concept of cytologicalzonation)叶原座(leaf buttress)初生组织(primary tissue)初生结构(primary structure)通气组织(aerenchyma)淀粉鞘(starch sheath)无限维管束(open bundle)有限维管束(closed bundle)外韧维管束(collateral bundle)双韧维管束(bicollateral bundle)周韧维管束(amphicribral bundle)周木维管束(amphivasal bundle)同心维管束(concentric bundle)中柱(stele)原生中柱(protostele)管状中柱(siphonostele)中央柱(centralcylinder)维管柱(vascularcylinder)内始式(endarch)环髓带(perimedullaryzone)髓腔(pith cavity)髓射线(pith ray)初生射线(primary ray)树脂道(resin canal)维管束鞘(bundle sheath)下皮(hypodermis)初生加厚分生组织(primary thickening meristem)束中形成层(fascicularcambium)。
植物生物学题库+答案

一、名词解释1、植物生物学:是从细胞、组织、器官、个体、类群、生态系统等不同层次,阐述植物的形态、构造、生理、分类、分布、遗传变异和进化及其与环境相互关系的一门课程。
2、纹孔: 初生壁完全不被次生壁覆盖的区域。
3、胞间连丝:是穿过细胞壁的原生质细丝,它连接相邻细胞间的原生质体。
它是细胞原生质体之间物质和信息直接联系的桥梁,是多细胞植物体成为一个构造和功能上统一的有机体的重要保证。
4、完全叶:具有叶片、叶柄和托叶三局部的叶称为完全叶.5、不完全叶:缺少叶片、叶柄和托叶中任一局部或两局部的称为不完全叶。
6、单叶:在一个叶柄上生有一个叶片的叶7、复叶:在一个总叶柄上生有多个小叶片的叶8、叶序:叶在茎上的排列方式,一般有3种根本类型:互生、对生和轮生。
9、等面叶:叶肉中无栅栏组织和海绵组织的分化10、异面叶: 叶肉中有栅栏组织和海绵组织的分化11、细胞型胚乳:从初生胚乳核分裂开场,即伴随细胞壁的形成,以后各次分裂都是如此。
12、核型胚乳:是被子植物普遍的胚乳发育形式。
初生胚乳核在最初一段发育时期进展细胞核的分裂而细胞质不分裂,不形成细胞壁,直至胚乳发育后期才发生细胞质分裂,产生细胞壁,形成胚乳细胞。
13、初生生长:顶端分生组织经过分裂、生长、分化产生各种成熟组织的过程。
14、初生构造:植物茎端或根端的顶端分生组织细胞经过分裂、分化和生长形成的构造。
15、次生构造:由根和茎的维管形成层和木栓形成层产生的构造16、无限花序:其开花的顺序是花轴下部的花先开,渐及上部,或由边缘开向中心,如油菜的总状花序17、有限花序:花序中最顶点或最中心的花先开,渐及下边或周围,如番茄的聚伞花序。
18、传粉: 指花粉粒由花粉囊中散出,经媒介的作用而传送到柱头上的过程。
19、双受精:花粉管到达胚囊后,其末端破裂,释放出的两个精子,一个与卵细胞融合,成为二倍体的受精卵(合子),另一个与两个极核(或次生核)融合,形成三倍体的初生胚乳核。
植物的基础知识--输导系统

植物的基础知识--输导系统编者按:植物的基础知识,属于《药用植物学》的内容,是掌握《中药鉴定学》的基础。
因此,我们为大家总结了一系列的植物学基本概念,以期帮助大家更好地掌握知识。
(一)维管束及其类型:维管束是由几个不同的组织联合在一起组成的束状结构,在植物体中构成复杂的输导系统。
从蕨类植物开始出现维管束。
维管束主要由韧皮部与木质部构成。
韧皮部主要由筛管、伴胞、韧皮薄壁细胞与韧皮纤维组成,这部分质较柔韧,故称韧皮部,木质部主要由导管、管胞、木薄壁细胞与木纤维组成,这部分木质坚硬,故称木质部。
双子叶植物和裸子植物根和茎的维管束,在韧皮部和木质部之间有形成层存在,能继续增生长大,所以称为无限维管束(开放性维管束)。
单子叶植物和蕨类植物根和茎的维管束没有形成层,不能增生长大,所以称为有限维管束。
根椐维管束中韧皮部和木质部相与互间排列方式的不同,以及形成层的有无,维管束可分为下列几种类型。
1.有限外韧维管束:韧皮部位于外侧,木质部位于内侧,两者并行排列,中间无形成层如单子叶植物茎的维管束。
2.无限外韧维管束:与有限外韧维管束的不同点是韧皮部与木质部之间有形成层。
如裸子植物和双子叶植物茎中的维管束。
3.双韧维管束:木质部的内外两侧都有韧皮部。
常见于茄科、葫芦科、桃金娘科等植物的茎中。
如颠茄、南瓜茎的维管束。
4.周韧维管束:木质部在中间,韧皮部围绕在木质部的四周。
常见于蕨类某些植物的茎、叶中。
如贯众。
5.周木维管束:韧皮部在中间,木质部围绕在韧皮部的四周。
常见于百合科(轮叶王孙属)、鸢尾科、天南星科(菖蒲属)、莎草科、仙茅科等某些植物茎中。
6.辐射维管束:韧皮部和木质部交互间隔排列,呈辐射状。
存于某被子植物根的初生构造中。
考试大网站整理(二)中柱及其类型:中柱是维管束植物住状器官(茎、根)皮层以内的部分,包括中柱鞘、维管束和髓(如存在时)。
中柱鞘位于中柱最外层,由一至数层薄壁或厚壁细胞组成,中柱鞘的薄壁细胞在一定时期可恢复分生能力,产生侧根、不定根、不定芽、木栓形成层等。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
? 2 结构
? 3 生长发展
? 4 运输ห้องสมุดไป่ตู้程 [编辑] 组成
由筛分子,薄壁组织和厚壁组织细胞构成.
[编辑] 筛分子
包括筛管和筛胞。
筛管分布于被子植物,成熟后的筛分子会损失掉大部分细胞器,只能由旁边的伴胞提供营养。筛分子和伴胞来源于同一筛母细胞。筛管分子顶端相互连接,胞壁之间穿孔,形成筛板。联络索通过筛板孔上下贯穿,以调节运输。伴胞通过胞间连丝与筛管分子联系,保持筛管分子的形态与渗透压,并为之提供营养和能量。
[编辑] 生长发展
根据出现时间的早晚和来源,木质部可划分为初生木质部(primary xylem)和次生木质部(secondary xylem)。
[编辑] 初生木质部
来自于原形成层(Procambium),由薄壁组织和木纤维组成,但没有木射线。根据分化的早晚,初生木质部又可分为原生木质部(Protoxylem)和后生木质部(Metaxylem)。
目录
[隐藏]
? 1 结构
o 1.1 导管
o 1.2 管胞(假导管)
o 1.3 木射线
o 1.4 薄壁组织
o 1.5 木纤维
? 2 生长发展
o 2.1 初生木质部
? 2.1.1 原生木质部
? 2.1.2 后生木质部
o 2.2 次生木质部
? 3 运输过程
3.真中柱:真中柱是外韧管状中柱由于叶隙和髓射线割裂成束而形成的。横切面为一圈环状排列的无限外韧维管束,有射线间隔;纵面观呈网状。为一般双子叶植物和裸子植物所具有。
4.散生中柱:散生中柱是有限外韧维管束或周木维管束,散生于基本组织中。为一般单子叶植物的茎或根茎所具有。如玉蜀黍茎、射干根茎等。
(二) 中柱及其类型:中柱是维管束植物住状器官(茎、根)皮层以内的部分,包括中柱鞘、维管束和髓(如存在时)。中柱鞘位于中柱最外层,由一至数层薄壁或厚壁细胞组成,中柱鞘的薄壁细胞在一定时期可恢复分生能力,产生侧根、不定根、不定芽、木栓形成层等。由于植物及器官不同,中柱鞘有的明显,有的不明显。中柱中的维管束类型和维管束排列方式的不同,而有各种不同类型的中柱。这些类型在根和茎类中草药鉴定上有一定意义。
? 4 注释
? 5 参考文献 [编辑] 结构
木质部由导管、管胞、木射线、薄壁组织和木纤维构成。
[编辑] 导管
存在于大部分的被子植物、裸子植物的买麻藤目和一部分蕨类植物(如欧洲蕨)之中。上下胞壁穿孔而互相衔接,因此液体在其中的流动速度比在管胞中的快。每个导管细胞被称做导管分子。生长初期的导管细胞是活细胞,但后来成熟时细胞质解离,细胞死亡,胞壁木质化。在木质化的过程中,胞壁会出现纹孔。通过纹孔,水分可以横向流动。从演化的角度看,导管是由原始的管胞发展出来的。但已不像管胞那样具有支持功能了。而支持功能则有另一种从原始管胞发展而来的结构即木纤维承担。
1.原生中柱:原生中柱是中柱内的维管系统形成一个实心柱,中心无髓部,中央是木质部,周边是韧皮部。
(1)典型原生中柱:木质部呈圆柱形实心柱,韧皮部围绕其周围。在蕨类植物如海金沙的根茎中可以见到。
(2)星状中柱:木质部呈星状(辐射状)的实心柱,韧皮部位于木质部的星角之间。如星木属(Asteroxylon)植物的中柱属这种类型。
双子叶植物和裸子植物根和茎的维管束,在韧皮部和木质部之间有形成层存在,能继续增生长大,所以称为无限维管束(开放性维管束)。单子叶植物和蕨类植物根和茎的维管束没有形成层,不能增生长大,所以称为有限维管束。根椐维管束中韧皮部和木质部相与互间排列方式的不同,以及形成层的有无,维管束可分为下列几种类型。
[编辑] 管胞(假导管)
少数被子植物、裸子植物和蕨类植物只有管胞而无导管,而大部分被子植物则两者兼有。管胞细胞呈纺锤状,即两端尖细,中间膨大。其生活史与导管相似,最初为活细胞,后来胞质解离细胞死亡。胞壁也会木质增厚,出现纹孔,纹孔膜具有高度渗透性。裸子植物具有典型的具缘纹孔。水分的流动可通过纹孔塞的活动被调节。在较原始的植物中,管胞具有运输和支持功能。
筛胞则存在于蕨类和裸子植物的韧皮部。筛胞和筛管分子结构相似,但并无筛板。筛胞由蛋白质细胞(protein cell)(在松树的针叶横切面可见),又名斯特拉斯堡细胞,提供营养。但蛋白质细胞和筛胞并不来自同一母细胞。
[编辑] 厚壁细胞
可分成纤维和石细胞两种。
[编辑] 薄壁组织
与木质部相似,分为轴向薄壁组织细胞和射线薄壁组织细胞。其中有一部分细胞薄壁内突,这些细胞特化为传递细胞,起着调节运输的作用。
4.周韧维管束:木质部在中间,韧皮部围绕在木质部的四周。常见于蕨类某些植物的茎、叶中。如贯众。
5.周木维管束:韧皮部在中间,木质部围绕在韧皮部的四周。常见于百合科(轮叶王孙属)、鸢尾科、天南星科(菖蒲属)、莎草科、仙茅科等某些植物茎中。
6.辐射维管束:韧皮部和木质部交互间隔排列,呈辐射状。存于某被子植物根的初生构造中。
[编辑] 运输过程
主条目:植物体无机盐运送途径
木质部负责将从根部吸收的水分及溶解其中的离子往上运输。动力则来自植物的蒸散作用和根压
韧皮部是维管植物的运输组织,负责将光合作用的产物——蔗糖,由进行光合作用的器官运输到植物的其他部位。
目录
[隐藏]
? 1 组成
o 1.1 筛分子
o 1.2 厚壁细胞
维管植物有两个重要的辨别方法:
1. 维管植物具有维管组织,可以让植物生长到一个较大的体积。非维管植物则一直保持着较小的体积。
2. 在维管植物,其主要生成阶段是孢子体。
在木质部和韧皮部,水分皆会被不停运送:木质部将水和无机溶质从根部运送往叶片,而韧皮部则会把植物中的有机溶质运送往全株植物。 维管植物都含有木质化的组织(即木质导管或管胞)。
网状中柱:是由双韧管状中柱演变而来,在横切面上见有环状徘列的周韧维管束,一般在各维管束外都有维管束鞘和内皮层,而纵面观则呈网状。多数真蕨类植物具有这种网状中柱,如贯众。
(2)外韧管状中柱:是只在管状木质部的外方有一圈韧皮部,如阴地蕨。有的外韧管状中柱的韧皮部仍连成环,但木质部开始分散成束(在系统发育上可看成是典型管状中柱和真中柱的过渡类型),如紫箕。
维管束是指维管植物(包括蕨类植物、裸子植物和被子植物)的维管组织,由木质部和韧皮部成束状排列形成的结构。维管束多存在于茎(草本植物和木本植物幼体)、叶(叶中的维管束又称为叶脉)等器官中。维管束相互连接构成维管系统,主要作用是为植物体输导水分、无机盐和有机养料等,也有支持植物体的作用。
类型
维管束根据木质部和韧皮部的排列方式可以被分为三大类:[1][2]
? 外韧型:韧皮部位于木质部的外侧。
? 双韧型:维管束内外周均为韧皮部(内生韧皮部和外生韧皮部)。如南瓜的茎。
? 同心型:由一种维管组织包着另一种维管组织。又可细分为两类:
o 周木型:木质部在外周,成圈状包围里面的韧皮部。
o 周韧型:韧质部在外周,成圈状包围里面的木皮部。
此外,根据能否继续发育,还可以将维管束分为有限维管束和无限维管束两类:
[编辑] 后生木质部
分化于原生木质部之后,次生木质部之前,若植物具有次级生长的话。后生木质部发生于初生植物仍然生长的时候,成熟于植物的伸长活动之后,所以避免了被拉长破坏。后生木质部除具有薄壁组织,管状分子外,还有木纤维。
[编辑] 次生木质部
具有次级生长的植物,可以通过维管形成层的活动产生次生木质部。根据其排列方向,又可分为轴向系统和径向系统。这些植物正是通过次生木质部的不断往外生长,中心不断外移木质化,而实现径向加粗的。
1.有限外韧维管束:韧皮部位于外侧,木质部位于内侧,两者并行排列,中间无形成层如单子叶植物茎的维管束。
2.无限外韧维管束:与有限外韧维管束的不同点是韧皮部与木质部之间有形成层。如裸子植物和双子叶植物茎中的维管束。
3.双韧维管束:木质部的内外两侧都有韧皮部。常见于茄科、葫芦科、桃金娘科等植物的茎中。如颠茄、南瓜茎的维管束。
形成层是植物中纵向贯穿根和茎的一层组织,位于木质部和韧皮部之间,形成层一般由多层细胞所组成,严格说来,其中只有一层原始细胞,可以不断地向内外分裂而增生细胞,不断增生的子细胞分化为木质部和韧皮部以及射线的细胞,使得植物的根和茎能不断地生长加粗。属于侧生分生组织。
形成层一般存在于裸子植物和被子植物的双子叶植物中,但在单子叶植物中基本没有形成层,所以单子叶植物生长不能不断地加粗,绝大部分为草本植物,只有棕榈科具有乔木状,也是依靠维管束发展,不能有充分的次生生长能力
[编辑] 木射线
起横向(即与根茎纵轴垂直)运输和贮存作用的薄壁组织,在维管形成层内次生木质部中成辐射发散状排列。
[编辑] 薄壁组织
可以说,这个概念与木射线有重叠。木射线其实就是横向排列的薄壁组织。除木射线外,木质部还有与根茎主轴平行的轴向薄壁细胞,起贮存作用。
[编辑] 木纤维
如上所述,木纤维是原始管胞的一个发展方向,是两端尖细的厚壁组织细胞,具有木质化的次生壁,因此具有支持植物体的作用。
[编辑] 叶脉
在叶中,维管束又被称为叶脉。叶脉通常位于叶肉的海绵组织中,其木质部朝向叶的正面,而韧皮部则朝向叶的背面。由于韧皮部在植物中是用于传输糖分等营养物质的,所以经常可以发现蚜虫多聚集于叶子的背面而非正面。
维管植物是指具有维管组织的植物,维管植物拥有专门的组织来运输水分和养分。它包括蕨类和种子植物,种子植物又分为裸子植物和被子植物。
? 有限维管束:不具有形成层,不能发育出新的木质部和韧皮部。蕨类植物和单子叶植物的维管束多属于此类。
? 无限维管束:在木质部和韧皮部之间有形成层,可以产生新的木质部和韧皮部。这类维管束可以使植物的枝干不断加粗。一般存在于裸子植物和双子叶植物中。