甘油三酯的代谢
脂代谢思维导图

脂代谢思维导图甘油三酯代谢过程合成代谢1、合成部位及原料肝、脂肪组织、小肠是合成的重要场所,以肝的合成能力最强,注意:肝细胞能合成脂肪,但不能储存脂肪。
合成后要与载脂蛋白、胆固醇等结合成极低密度脂蛋白,入血运到肝外组织储存或加以利用。
若肝合成的甘油三酯不能及时转运,会形成脂肪肝。
脂肪细胞是机体合成及储存脂肪的仓库。
合成甘油三酯所需的甘油及脂肪酸主要由葡萄糖代谢提供。
其中甘油由糖酵解生成的磷酸二羟丙酮转化而成,脂肪酸由糖氧化分解生成的乙酰CoA合成。
2、合成基本过程①甘油一酯途径:这是小肠粘膜细胞合成脂肪的途径,由甘油一酯和脂肪酸合成甘油三酯。
②甘油二酯途径:肝细胞和脂肪细胞的合成途径。
脂肪细胞缺乏甘油激酶因而不能利用游离甘油,只能利用葡萄糖代谢提供的3-磷酸甘油。
分解代谢即为脂肪动员,在脂肪细胞内激素敏感性甘油三酯脂的酶作用下,将脂肪分解为脂肪酸及甘油并释放入血供其他组织氧化。
甘油甘油激酶-->3-磷酸甘油-->磷酸二羟丙酮-->糖酵解或有氧氧化供能,也可转变成糖脂肪酸与清蛋白结合转运入各组织经β-氧化供能。
脂肪酸的分解代谢-β-氧化在氧供充足条件下,脂肪酸可分解为乙酰CoA,彻底氧化成CO2和H2O并释放出大量能量,大多数组织均能氧化脂肪酸,但脑组织例外,因为脂肪酸不能通过血脑屏障。
其氧化具体步骤如下:1.脂肪酸活化,生成脂酰CoA。
2.脂酰CoA进入线粒体,因为脂肪酸的β-氧化在线粒体中进行。
这一步需要肉碱的转运。
肉碱脂酰转移酶是脂酸β氧化的限速酶,脂酰CoA进入线粒体是脂酸β-氧化的主要限速步骤,如饥饿时,糖供不足,此酶活性增强,脂肪酸氧化增强,机体靠脂肪酸来供能。
3.脂肪酸的β-氧化。
丁酰CoA经最后一次β氧化:生成2分子乙酰CoA,故每次β氧化1分子脂酰CoA生成1分子FADH2,1分子NADH+H+,1分子乙酰CoA,通过呼吸链氧化前者生成2分子ATP,后者生成3分子ATP。
甘油三酯的代谢

⽢油三酯的代谢20 ~ 20 学年度第学期教师课时授课教案学科系:医学院授课教师:专业:临床科⽬:⽣物化学年⽉⽇年⽉⽇第七章脂类代谢第⼆节⽢油三酯的代谢⽢油三酯是机体主要的脂类,其代谢主要包括分解代谢与合成代谢。
各组织中的⽢油三酯不断地进⾏⾃我更新,其中脂肪组织和肝有较⾼的更新率,其次为肠黏膜和肌⾁组织,⽽⽪肤和神经组织中⽢油三酯更新率最低。
⼀、⽢油三酯的分解代谢(⼀)脂肪动员贮存在脂肪组织中的⽢油三酯,在脂肪酶催化下,逐步⽔解为⽢油和游离脂肪酸(FFA)并释放⼊⾎,经⾎液运输⾄全⾝各组织⽽被氧化利⽤的过程称为脂肪动员。
脂肪动员的过程如下:脂肪⽔解是在⽢油三酯(TG)脂肪酶、⽢油⼆酯(DC)脂肪酶、⽢油⼀酯(MG)脂肪酶的作⽤下逐步完成,上述酶中,以⽢油三酯脂肪酶的活性最低,故⽢油三酯脂肪酶是脂肪动员的限速酶,⽽其活性受多种激素的调控,因此⼜称激素敏感性脂肪酶。
胰岛素、前列腺素E2可降低这种酶的活性,抑制脂肪动员,故称为抗脂解激素;胰⾼⾎糖素、肾上腺素、去甲肾上腺素等可提⾼该酶的活性,促进脂肪动员,称为脂解激素。
机体对脂肪动员的调控就是通过激素对这⼀限速酶的作⽤实现的。
进⾷后胰岛素分泌增加,脂肪动员减弱;当禁⾷、饥饿或处于兴奋时,肾上腺素、胰⾼⾎糖素等分泌增加,脂肪动员加强。
脂肪动员⽣成的脂肪酸和⽢油释放⼊⾎,游离脂肪酸与⾎浆⽩蛋⽩结合成复合物,运输到全⾝组织⽽被利⽤。
(⼆)⽢油的代谢脂肪动员产⽣的⽢油,可在肝、肾等组织氧化供能,也可进⾏糖异⽣。
在⽢油激酶催化下,⽢油磷酸化⽣成-磷酸⽢油,再脱氢⽣成磷酸⼆羟丙酮,后者可循糖代谢途径氧化供能或异⽣成糖,反应如下:(三)脂眆酸的氧化分解除脑组织和成熟红细胞外,⼤部分组织均能氧化脂肪酸,以肝和肌⾁最为活跃。
在氧供应充⾜的情况下,脂肪酸氧化分解为CO2和H20并释放⼤量的能量。
1.脂肪酸的活化脂肪酸在细胞质中进⾏活化。
在脂酰CoA合成酶的催化下,由ATP供能,辅酶A参与,活化⽣成脂酰CoA。
甘油三酯的合成代谢

甘油三酯的合成代谢?甘油三酯(Triglyceride),是长链脂肪酸和甘油形成的脂肪分子,是人体内含量最多的脂类,大部分组织均可以利用甘油三酯分解产物供给能量,同时肝脏、脂肪等组织还可以进行甘油三酯的合成,在脂肪组织中贮存。
人体可利用甘油、糖、脂肪酸和甘油一酯为原料,经过磷脂酸途径和甘油一酯途径合成甘油三酯。
1. 甘油一酯途径:以甘油一酯为起始物,与脂酰CoA共同在脂酰转移酶作用下酯化生成甘油三酯。
2. 磷脂酸途径:磷脂酸,即3-磷酸-1,2-甘油二酯,是合成含甘油脂类的共同前体。
糖酵解的中间产物—类磷酸二羟丙酮在甘油磷酸脱氢酶作用下,还原生成3-磷酸甘油;游离的甘油也可经甘油激酶催化,生成3-磷酸甘油(因脂肪及肌肉组织缺乏甘油激酶,故不能利用激离的甘油)。
3-磷酸甘油在脂酰转移酶作用下,与两分子脂酰CoA反应生成3-磷酸-1,2甘油二酯,即磷脂酸。
此外,磷酸二羟丙酮也可不转为3-磷酸甘油,而是先酯化,后还原生成溶血磷脂酸,然后再经酯化合成磷脂酸。
磷脂酸在磷脂酸磷酸酶作用下,水解释放出无机磷酸,而转变为甘油二酯,它是甘油三酯的前身物,只需酯化即可生成甘油三酯。
甘油三酯所含的三个脂肪酸可以是相同的或不同的,可为饱和脂肪酸或不饱和脂肪酸。
甘油三酯的合成速度可以受激素的影响而改变,如胰岛素可促进糖转变为甘油三酯。
由于胰岛素分泌不足或作用失效所致的糖尿病患者,不仅不能很好利用葡萄糖,而且葡萄糖或某些氨基酸也不能用于合成脂肪酸,而表现为脂肪的氧化速度增加,酮体生成过多,其结果是患者体重下降。
此外,胰高血糖素、肾上腺皮质激素等也影响甘油三酯的合成。
TCA循环等等重要代谢途径哪些步骤有维生素或其辅酶参与反应?1、乙酰CoA与草酰乙酸的羧基进行醛醇型缩合,柠檬酸转变成异柠檬酸:前者由柠檬酸合成酶催化,后者由顺乌头酸酶催化,均为变构酶,需要维生素B12作为变构酶的辅酶,参与一些异构化作用。
2、第一次氧化脱酸:在异柠檬酸脱氢酶作用下,异柠檬酸生成α-酮戊二酸、NADH和CO2 而第二次氧化脱羧:在α-酮戊二酸脱氢酶系作用下,α-酮戊二酸氧化脱羧生成琥珀酰-CoA、。
脂代谢—甘油三酯的代谢(生物化学课件)

=
O
肉 RCH2CH2C~SCoA
AMP 碱
脂酰CoA
PPi
转
合成酶
ATP
运
CoASH
O
载
=
RCH2CH2C-OH
体
脂肪酸
线 粒 体 膜
O
=
RCH2CH2C~SCoA
脂酰CoA
FAD
脱氢酶
FADH2
β αO
RCH=CHC~SCoA
2ATP 呼吸链 H2O
=
=
⊿--2烯酰CoA
H2O
水化β 酶 α O
脂酰CoA
FAD
脱氢酶
β
αO
FADH2
=
RCH=CHC~SCoA
⊿2--烯脂酰CoA 水化酶
H2O
β
αO
=
RCHOHCH2C~SCoA
L(+)-β羟脂酰
NAD+
CoA脱氢酶
NADH+H+
βα O
=
RCOCH2C~SCoA
β酮脂酰CoA O硫解酶
CoA-SH
=
RC~SCoA + CH3CO~SCoA
脂酰CoA 反⊿2-烯脂酰CoA L(+)-β羟脂酰CoA β酮脂酰CoA 脂酰CoA+乙酰CoA
RCHOHCH2C~SCoA
L(+)-β羟脂酰
NAD+
CoA脱氢酶
NADH+H+
βα O
=
RCOCH2C~SCoA
β酮脂酰CoA
3ATP 呼吸链 H2O
硫解酶
O
CoA + CH3CO~SCoA
甘油三酯的代谢

甘油三酯的代谢
甘油三酯(Triglycerides)是一种由甘油和三个脂肪酸分子组成的脂质。
以下是甘油三酯的代谢过程:
1. 摄入甘油三酯:甘油三酯主要通过饮食摄入,尤其是高脂肪食物。
2. 胰岛素的作用:血液中的胰岛素可以促进甘油三酯的合成和储存。
它会刺激脂肪组织摄取血液中的脂肪酸,并与甘油结合形成甘油三酯,储存在脂肪细胞中。
3. 分解甘油三酯:当身体需要能量时,脂肪细胞中的甘油三酯会被分解成甘油和脂肪酸。
4. 脂肪酸的运输和氧化:脂肪酸会与白蛋白结合,在血液中被运输到需要能量的组织。
在线粒体中,脂肪酸会经过β-氧化途径被氧化成乙酰辅酶A,从而产生能量。
5. 甘油的代谢:分解的甘油可以被肝脏转化为葡萄糖,供给需要能量的组织。
总之,甘油三酯的代谢主要涉及甘油和脂肪酸的合成、储存、分解和运输等过程。
这个过程受到胰岛素和其他激素的调控,并参与能量供给和脂肪代谢等生理过程。
甘油三酯和磷脂代谢脂代谢

06
甘油三酯和磷脂代谢的调节策略
药物治疗
贝特类药物
通过激活过氧化物酶体增殖物激活受体( PPAR)来降低甘油三酯和升高高密度脂蛋 白胆固醇(HDL-C),如非诺贝特、苯扎贝 特等。
他汀类药物
通过抑制HMG-CoA还原酶来降低胆固醇, 如阿托伐他汀、瑞舒伐他汀等。
营养干预
控制总热量摄入
01
减少高糖、高脂肪和高热量食物的摄入,以控制体重和血脂水
甘油三酯的合成与分解
甘油三酯的合成
甘油三酯的合成需要脂肪酸和甘油的共同参与,脂肪酸在肝脏、脂肪组织和肌肉 中合成,然后与甘油形成甘油三酯。
甘油三酯的分解
甘油三酯在脂肪酶的作用下分解成甘油二酯和脂肪酸,再进一步分解成二氧化碳 和水,释放能量。
甘油三酯的调节机制
激素调节
胰岛素、胰高血糖素、肾上腺素等激素调节甘油三酯的合成 与分解。
04
甘油三酯与磷脂代谢的关系
甘油三酯对磷脂代谢的影响
甘油三酯合成磷脂
甘油三酯是合成磷脂的重要前体,通过磷脂酶的作用,合成磷脂酰甘油和溶 血磷脂酰甘油。
调节脂肪酶活性
甘油三酯通过调节脂肪酶的活性来影响磷脂的合成和代谢。高浓度的甘油三 酯可以抑制脂肪酶的活性,从而降低脂肪酶分解磷脂的能力。
磷脂对甘油三酯代谢的影响
促进脂肪分解
磷脂酰胆碱等磷脂可以促进脂肪酶的作用,加速脂肪分解,从而降低血液中甘油 三酯的水平。
调节脂肪酸摄取
磷脂可以促进细胞对脂肪酸的摄取和吸收,从而影响甘油三酯的合成和代谢。
甘油三酯与磷脂代谢的交互作用
相互影响脂肪合成
甘油三酯和磷脂的代谢途径相互影响,共同调节脂肪的合成和代谢。
调节脂肪酸氧化
甘油三酯和磷脂可以调节脂肪酸的氧化过程,从而影响能量代谢和脂肪酸的利用。
甘油三酯代谢

与内质网紧密结合的脂肪酸ω-氧化酶系由羧化酶、脱氢酶、NADP、 NAD+及细胞色素P-450(cytochrome P450, Cyt P450)等组成。
脂肪酸ω-甲基碳原子在脂肪酸ω-氧化酶系作用下,经ω-羟基脂肪酸、 ω-醛基脂肪酸等中间产物,形成α, ω-二羧酸。这样,脂肪酸就能从任一 端活化并进行β-氧化。
1904年,努珀(F. Knoop)采用不能被机体分解的苯基标记脂肪 酸ω-甲基,喂养犬,检测尿液中的代谢产物。发现不论碳链长短,如果 标记脂肪酸碳原子是偶数,尿中排出苯乙酸;如果标记脂肪酸碳原子是 奇数,尿中排出苯甲酸。据此,努珀提出脂肪酸在体内氧化分解从羧基 端β-碳原子开始,每次断裂2个碳原子,即“β-氧化学说”。
第三节
甘油三酯的代谢
Metabolism of Triglyceride
本 节
甘油三酯的分解代谢
主
• 脂肪动员
要
• 甘油进入糖代谢
内 容
• 脂酸的β氧化
• 脂酸的其他氧化方式
• 酮体的生成和利用
甘油三酯的合成代谢
脂肪酸的合成代谢
一、甘油三酯氧化分解产生大量ATP供机体需要
(一)甘油三酯分解代谢从脂肪动员开始
较短链 脂酸
(线粒体)
β氧化
3. 丙酰CoA转变为琥珀酰CoA进行氧化
Ile Met Thr Val 奇数碳脂酸 胆固醇侧链
L-甲基丙二酰CoA
消旋酶
CH3CH2CO~CoA
CO2
羧化酶 (ATP、生物素)
D-甲基丙二酰CoA
变位酶 5-脱氧腺苷钴胺素 琥珀酰CoA
TAC
4. 脂肪酸氧化还可从远侧甲基端进行
葡萄糖和甘油三酯的转化

葡萄糖和甘油三酯的转化一、引言葡萄糖和甘油三酯是人体内重要的能量来源,而它们之间的转化也是人体内代谢过程中不可或缺的一部分。
本文将从葡萄糖和甘油三酯的结构、代谢途径以及相关激素等方面进行详细介绍。
二、葡萄糖的结构和代谢途径1. 葡萄糖的结构葡萄糖是一种单糖,化学式为C6H12O6。
它的分子结构包括六个碳原子、十二个氢原子和六个氧原子。
2. 葡萄糖在人体内的代谢途径葡萄糖在人体内主要通过以下两种途径进行代谢:(1)醛基途径:葡萄糖经过一系列反应被转化为丙酮酸,最终通过三羧酸循环产生ATP。
(2)磷酸戊糖途径:葡萄糖经过一系列反应被转化为乙醛-3-磷酸和丙酮醛-3-磷酸,最终通过糖原合成或者葡萄糖氧化产生ATP。
三、甘油三酯的结构和代谢途径1. 甘油三酯的结构甘油三酯是一种三酰基甘油,化学式为C3H5(OH)3(COOCnH2n+1)3。
它的分子结构包括一个甘油分子和三个脂肪酸分子。
2. 甘油三酯在人体内的代谢途径甘油三酯在人体内主要通过以下两种途径进行代谢:(1)脂肪酸β-氧化途径:脂肪酸被转化为乙酰辅酶A,经过一系列反应产生ATP。
(2)脂肪合成途径:乙酰辅酶A和丙酮-3-磷酸合成脂肪酸,最终与甘油形成甘油三酯。
四、葡萄糖转化为甘油三酯的途径1. 葡萄糖通过磷酸戊糖途径转化为甘油三酯当血液中的葡萄糖浓度过高时,葡萄糖通过磷酸戊糖途径被转化为乙醛-3-磷酸和丙酮醛-3-磷酸。
这两种物质可以进入脂肪合成途径,与甘油结合形成甘油三酯。
2. 葡萄糖通过脂肪合成途径转化为甘油三酯当血液中的胰岛素水平升高时,葡萄糖被转化为乙酰辅酶A和丙酮-3-磷酸,进入脂肪合成途径。
在此过程中,乙酰辅酶A和丙酮-3-磷酸与甘油结合形成甘油三酯。
五、影响葡萄糖转化为甘油三酯的因素1. 胰岛素胰岛素是调节血糖水平的重要激素,它可以促进葡萄糖转化为甘油三酯。
2. 饮食高碳水化合物、高脂肪的食物会增加血液中的葡萄糖和甘油三酯浓度,从而促进葡萄糖转化为甘油三酯。
甘油三酯和磷脂代谢

摄入碳水化合物的种类和数量会影响甘油三酯和磷脂 的代谢。
蛋白质
摄入蛋白质的种类和数量会影响甘油三酯和磷脂的代 谢。
运动对甘油三酯和磷脂代谢的影响
有氧运动
有氧运动可以促进甘油三酯的分解和糖异生作用 ,以及磷脂的分解。
力量训练
力量训练可以促进甘油三酯的合成和储存,以及 磷脂的合成。
运动对血脂的影响
探索利用调节甘油三酯和磷脂代谢的膳食补充剂
膳食补充剂可以作为一种有效的手段来调节甘油三酯和磷脂代谢,进而预防和治疗相关疾病。
发现针对甘油三酯和磷脂代谢的新靶点
通过深入研究甘油三酯和磷脂代谢的分子机制,有助于发现新的靶点,为开发新的治疗方法提供基础。
发展更加灵敏和准S
生理意义
磷脂的分解代谢为人体提供了能量和代谢 原料,同时也有助于维持人体的正常生理 功能。
磷脂与细胞膜的结构和功能
磷脂与细胞膜的结构
磷脂双分子层构成了细胞膜的基本骨架,对于维持细胞膜的结构和稳定性具有重要作用。
磷脂与细胞膜的功能
磷脂分子中的亲水性磷酸头部朝向细胞外液,而疏水性的脂肪酸尾部则朝向细胞内,这种结构特点使得磷脂分子 能够有效地分隔细胞内外环境,维持细胞内外渗透压平衡。此外,磷脂分子还可以参与细胞信号转导、细胞运动 、细胞分裂和分化等生命活动。
脂肪酸合成
脂肪酸是合成甘油三酯的基本单位之一。脂肪酸主要在肝脏和脂肪组织 中合成,通过乙酰CoA和NADPH作为原料,经过多个步骤合成。
03
甘油合成
甘油是另一个合成甘油三酯的基本单位。甘油主要在肝脏中合成,由糖
酵解途径中的3-磷酸甘油脱氢酶催化的反应生成。
甘油三酯的分解代谢
甘油一酯水解
甘油三酯在甘油一酯酶的作 用下可水解成甘油二酯和脂 肪酸。这个反应是甘油三酯
简述甘油三酯的分解代谢

简述甘油三酯的分解代谢1.引言1.1 概述甘油三酯(triglyceride)是一种重要的脂类化合物,在人体和动物体内广泛存在。
它是由一种甘油分子和三个脂肪酸分子通过酯键结合而成的。
作为我们日常饮食中主要的脂肪来源之一,甘油三酯在体内具有多种生理功能和作用。
正常情况下,通过食物摄入的脂肪会转化为甘油三酯储存在脂肪细胞中,以提供能量的长期储备。
甘油三酯的分解代谢是指将体内储存的甘油三酯分解成甘油和脂肪酸,以供能量消耗和维持生命活动。
这个过程主要发生在脂肪组织中的脂肪细胞内,通过一系列的酶反应逐步进行。
甘油三酯的分解代谢对人体的能量平衡和体脂调节非常重要。
当我们的能量需求增加时,例如进行体力活动或长时间禁食,体内储存的甘油三酯会被分解释放出来,供给能量消耗。
而在摄入过多能量的情况下,多余的甘油三酯会重新合成并储存起来,导致体重增加和肥胖。
甘油三酯的分解代谢涉及多种酶的参与和多个途径的调控。
其中最关键的酶包括甘油三酯脂肪酶(triglyceride lipase)和激活蛋白激酶A (protein kinase A),它们通过磷酸化等机制促进甘油三酯的分解。
而甘油和脂肪酸的进一步利用则需要通过线粒体内的β-氧化和其它代谢途径进行。
甘油三酯分解代谢的研究对于深入理解脂肪代谢、肥胖等相关疾病的发生机制以及预防和治疗具有重要意义。
还有许多未知的问题等待我们进一步探索和研究,例如甘油三酯分解代谢的调控机制、与疾病发生关联的分子机理等。
因此,对甘油三酯的分解代谢进行深入研究,可以为我们提供更多关于脂质代谢的信息,有助于更好地保护我们的健康,并为未来的临床治疗和疾病预防提供新的思路和方法。
1.2 文章结构文章结构的目的是为读者提供一个清晰的逻辑框架,以便理解和阅读文章的内容。
本文的结构主要分为引言、正文和结论三个部分。
引言部分主要概述了甘油三酯的分解代谢,并简要介绍了本文的结构和目的。
正文部分包括了甘油三酯的定义和作用、甘油三酯的来源和合成、甘油三酯的分解代谢以及甘油三酯分解代谢的相关酶和途径。
脂肪酸和甘油三酯的分解代谢

由胆碱开始,胆碱来源于食物或磷酯酰胆碱的降解
3、磷脂酰丝氨酸的合成
(1)、 丝氨酸与磷脂酰乙醇胺的醇基酶促交换 磷酯酰乙醇胺+丝氨酸—磷酯酰丝氨酸+乙醇胺
动物、大肠杆菌中,磷脂酰丝氨酸可脱羧生成磷脂酰乙醇胺
(2)、 磷脂酸—CDP 二脂酰基甘油一磷脂酰丝氨酸(细菌中) P184反应式
4、磷脂酰肌醇的合成
第五节 鞘脂类的代谢
第六节 胆固醇的代谢
胞固醇的合成(自己看一下,不要求) 胞固醇中27个碳原子全部来源于乙酰CoA 。
3、8—二氨基-5-乙基-6-苯基菲啶溴盐。
a 胆碱+ATP 胆碱激酶
b 磷酸胆碱+CTP c.CDP 胆碱+甘油二酯
磷酸胆碱胞嘧啶核苷酸转移酶 磷酸胆碱转移酶
磷酸胆碱+ADP
CDP 胆碱+ppi 磷
脂酰胆碱+CMP
以上三个合成途径的关系:
糖原合成时,Glc 的活性形式是UDP-葡萄糖(尿嘧啶核苷二磷酸Glc ) 磷脂酰胆碱。
6教学设计-甘油三酯的分解代谢

《甘油三酯的分解代谢》教学设计一、教材分析物质代谢是生命活动的基础,《甘油三酯的分解代谢》这一节在整个生物化学三大营养物质代谢中占有非常重要的地位,本节的内容与前面章节中讲述的糖类代谢紧密联系,是继糖类代谢后的又一物质代谢知识的延伸、应用部分,有利于学生更好理解物质代谢的作用,三大营养物质的氧化分解供能又直接关系着后面的细胞呼吸和代谢类型,以及生态系统的物质循环等知识内容。
所以,怎样完成本节的教学十分重要。
本节教学内容包括脂肪动员、甘油的代谢、脂肪酸的氧化分解、酮体的代谢四个部分。
为了更好地完成教学,每讲一种物质的代谢联系该种物质与健康的关系,另外,我将糖类代谢和蛋白质代谢安排在前两个课时完成,为避免一课时内知识点过于集中的问题,我将甘油三酯的合成代谢放入下一小节去讲述,这样既可以处理好知识间的逻辑关系,又能重新复习前面所讲过的糖代谢和蛋白质代谢的有关知识,做到新课内容与已将知识的衔接。
二、学情分析我所面对的是高职临床医学一年级的学生,他们思维活跃,理解能力强,可以在老师的引导下从感性资料中抽象出理性的概念。
他们学习生物化学的兴趣浓厚,愿意参与与生物化学知识相关的社会问题的探讨。
另外一方面,他们天真、敏感,这个年龄段的学生很容易受到外界事物的影响,缺乏对事物的本质分析和识别能力。
他们正是身体发育的关键阶段,社会上、媒体中、娱乐圈中出现很多不科学的减肥方法,学生很容易盲目效仿,误入歧途,影响身心发育。
因此他们需要科学饮食观念和正确的审美观点。
三、教学设计思路脂类代谢与健康的内容涉及的肥胖问题可以说是当今的热门话题,肥胖的人越来越多,减肥方法五花八门,对学生影响很大。
我们的人才培养方案中提出提高学生的生物科学素养、注意理论联系实际的理念。
所以我在教学中提出有关肥胖这个中心问题,从中心问题出发解决甘油三酯的分解代谢,最后以解决肥胖问题来强化知识。
这样既有利于甘油三酯分解代谢途径的掌握,又有利于学生建立起生物是统一整体及辩证唯物主义的基本观点。
甘油三酯代谢及高甘油三酯血症的治疗
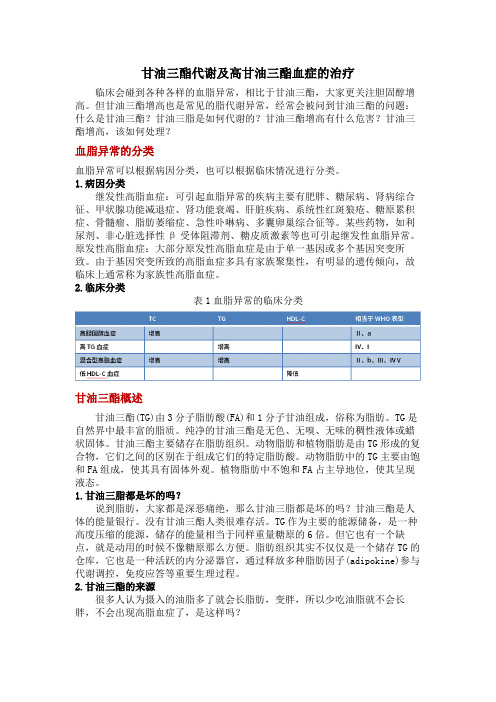
甘油三酯代谢及高甘油三酯血症的治疗临床会碰到各种各样的血脂异常,相比于甘油三酯,大家更关注胆固醇增高。
但甘油三酯增高也是常见的脂代谢异常,经常会被问到甘油三酯的问题:什么是甘油三酯?甘油三脂是如何代谢的?甘油三酯增高有什么危害?甘油三酯增高,该如何处理?血脂异常的分类血脂异常可以根据病因分类,也可以根据临床情况进行分类。
1.病因分类继发性高脂血症:可引起血脂异常的疾病主要有肥胖、糖尿病、肾病综合征、甲状腺功能减退症、肾功能衰竭、肝脏疾病、系统性红斑狼疮、糖原累积症、骨髓瘤、脂肪萎缩症、急性卟啉病、多囊卵巢综合征等。
某些药物,如利尿剂、非心脏选择性β受体阻滞剂、糖皮质激素等也可引起继发性血脂异常。
原发性高脂血症:大部分原发性高脂血症是由于单一基因或多个基因突变所致。
由于基因突变所致的高脂血症多具有家族聚集性,有明显的遗传倾向,故临床上通常称为家族性高脂血症。
2.临床分类表1血脂异常的临床分类甘油三酯概述甘油三酯(TG)由3分子脂肪酸(FA)和1分子甘油组成,俗称为脂肪。
TG是自然界中最丰富的脂质。
纯净的甘油三酯是无色、无嗅、无味的稠性液体或蜡状固体。
甘油三酯主要储存在脂肪组织。
动物脂肪和植物脂肪是由TG形成的复合物,它们之间的区别在于组成它们的特定脂肪酸。
动物脂肪中的TG主要由饱和FA组成,使其具有固体外观。
植物脂肪中不饱和FA占主导地位,使其呈现液态。
1.甘油三脂都是坏的吗?说到脂肪,大家都是深恶痛绝,那么甘油三脂都是坏的吗?甘油三酯是人体的能量银行。
没有甘油三酯人类很难存活。
TG作为主要的能源储备,是一种高度压缩的能源,储存的能量相当于同样重量糖原的6倍。
但它也有一个缺点,就是动用的时候不像糖原那么方便。
脂肪组织其实不仅仅是一个储存TG的仓库,它也是一种活跃的内分泌器官,通过释放多种脂肪因子(adipokine)参与代谢调控,免疫应答等重要生理过程。
2.甘油三酯的来源很多人认为摄入的油脂多了就会长脂肪,变胖,所以少吃油脂就不会长胖,不会出现高脂血症了,是这样吗?甘油三酯来源分为内源性合成及外源性摄取,通过饮食摄入的油脂(植物油和动物油)可以在小肠内转化为甘油三酯储存在体内(外源性摄入)。
甘油三酯的合成代谢

一、甘油三酯的水解(一)组织脂肪酶有三种,脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶,逐步水解R3、R1、R2,生成甘油和游离脂肪酸。
(二)第一步是限速步骤,肾上腺素、肾上腺皮质激素、高血糖素通过cAMP 和蛋白激酶激活,胰岛素和前列腺素E1相反,有抗脂解作用。
二、甘油代谢脂肪细胞没有甘油激酶,所以甘油被运到肝脏,由甘油激酶磷酸化为3-磷酸甘油,再由磷酸甘油脱氢酶催化为磷酸二羟丙酮,进入酵解或异生,并生成NADH。
三、脂肪酸的氧化(一)饱和偶数碳脂肪酸的氧化1. 脂肪酸的活化:脂肪酸先生成脂酰辅酶A才能进行氧化,称为活化。
由脂酰辅酶A合成酶(硫激酶)催化,线粒体中的酶作用于4-10个碳的脂肪酸,内质网中的酶作用于12个碳以上的长链脂肪酸。
生成脂酰AMP中间物。
乙酰acetyl;脂酰acyl2. 转运:短链脂肪酸可直接进入线粒体,长链脂肪酸需先在肉碱脂酰转移酶I催化下与肉碱生成脂酰肉碱,再通过线粒体内膜的移位酶穿过内膜,由肉碱转移酶II催化重新生成脂酰辅酶A。
最后肉碱经移位酶回到细胞质。
3. β-氧化:在线粒体基质进行,每4步一个循环,生成一个乙酰辅酶A。
l脱氢:在脂酰辅酶A脱氢酶作用下,α、β位生成反式双键,即Δ2反式烯脂酰辅酶A。
酶有三种,底物链长不同,都以FAD为辅基。
生成的FADH2上的氢不能直接氧化,需经电子黄素蛋白(ETF)、铁硫蛋白和辅酶Q进入呼吸链。
l水化:由烯脂酰辅酶A水化酶催化,生成L-β-羟脂酰辅酶A。
此酶只催化Δ2双键,顺式双键生成D型产物。
l再脱氢:L-β-羟脂酰辅酶A脱氢酶催化生成β-酮脂酰辅酶A和NADH,只作用于L型底物。
l硫解:由酮脂酰硫解酶催化,放出乙酰辅酶A,产生少2个碳的脂酰辅酶A。
酶有三种,底物链长不同,有反应性强的巯基。
此步放能较多,不易逆转。
4. 要点:活化消耗2个高能键,转移需肉碱,场所是线粒体,共四步。
每个循环生成一个NADH和一个FADH2,放出一个乙酰辅酶A。
生物化学笔记甘油三酯的合成代谢

一、软脂酸的合成(一)乙酰辅酶A的转运合成脂肪酸的碳源来自乙酰辅酶A,乙酰辅酶A是在线粒体形成的,而脂肪酸的合成场所在细胞质中,所以必需将乙酰辅酶A转运出来。
乙酰辅酶A在线粒体中与草酰乙酸合成柠檬酸,通过载体转运出线粒体,在柠檬酸裂解酶催化下裂解为乙酰辅酶A和草酰乙酸,后者被苹果酸脱氢酶还原成苹果酸,再氧化脱羧生成丙酮酸和NADPH,丙酮酸进入线粒体,可脱氢生成乙酰辅酶A,也可羧化生成草酰乙酸。
(二)丙二酸单酰辅酶A的生成乙酰辅酶A以丙二酸单酰辅酶A的形式参加合成。
乙酰辅酶A与碳酸氢根、ATP反应,羧化生成丙二酸单酰辅酶A,由乙酰辅酶A羧化酶催化。
此反应是脂肪酸合成的限速步骤,被柠檬酸别构激活,受软脂酰辅酶A抑制。
此酶有三个亚基:生物素羧化酶(BC)、生物素羧基载体蛋白(BCCP)和羧基转移酶(CT)。
(三)脂肪酸合成酶体系有7种蛋白,以脂酰基载体蛋白为中心,中间产物以共价键与其相连。
载体蛋白含巯基,与辅酶A类似,可由辅酶A合成。
(四)脂肪酸的合成1. 起始:乙酰辅酶A在ACP-酰基转移酶催化下生成乙酰ACP,然后转移到β-酮脂酰-ACP合成酶的巯基上。
2. ACP与丙二酸单酰辅酶A生成丙二酸单酰ACP,由ACP:丙二酸单酰转移酶催化。
3. 缩合:β-酮脂酰ACP合成酶将乙酰基转移到丙二酸单酰基的α-碳上,生成乙酰乙酰ACP,并放出CO2。
所以碳酸氢根只起催化作用,羧化时储存能量,缩合时放出,推动反应进行。
4. 还原:NADPH在β-酮脂酰ACP还原酶催化下将其还原为D-β-羟丁酰ACP。
β-氧化的产物是L-型。
5. 脱水:羟脂酰ACP脱水酶催化生成Δ2反丁烯酰ACP,即巴豆酰ACP。
6. 再还原:烯脂酰ACP还原酶用NADPH还原为丁酰ACP。
β-氧化时生成FADH2,此时是为了加速反应。
7. 第二次循环从丁酰基转移到β-酮脂酰ACP合成酶上开始。
7次循环后生成软脂酰ACP,可被硫酯酶水解,或转移到辅酶A上,或直接形成磷脂酸。
甘油三酯的分解代谢

第二节脂类的代谢一、脂肪的动员储存在脂肪细胞中的脂肪,被脂肪酶逐步水解为游离脂酸(free fatty acid, FFA)及甘油并释放入血以供其他组织氧化利用,该过程称为脂肪的动员。
在脂肪动员中,脂肪细胞内激素敏感性甘油三酯脂肪酶起决定性作用,它是脂肪分解的限速酶。
当禁食、饥饿或交感神经兴奋时,肾上腺素、去甲肾上腺素、胰高血糖素等分泌增加,作用于脂肪细胞膜表面受体,激活腺苷酸环化酶,促进cAMP合成,激活依赖cAMP的蛋白激酶,使胞液内HSL磷酸化而活化。
后者使甘油三酯水解成甘油二酯及脂酸。
这步反应是脂肪分解的限速步骤,HSL是限速酶,它受多种激素的调控,故称为激素敏感性脂肪酶。
能促进脂肪动员的激素称为脂解激素,如肾上腺素、胰高血糖素,ACTH及TSH等。
胰岛素、前列腺素E2及烟酸等抑制脂肪的动员,对抗脂解激素的作用。
脂解作用使储存在脂肪细胞中的脂肪分解成游离脂酸及甘油,然后释放入血。
血浆白蛋白具有结合游离脂酸的能力,每分子白蛋白可结合10分子FFA。
FFA不溶于水,与白蛋白结合后由血液运送至全身各组织,主要由心、肝、骨骼肌等摄取利用。
甘油溶于水,直接由血液运送至肝、肾、肠等组织。
主要是在肝甘油激酶作用下,转变为3-磷酸甘油;然后脱氢生成磷酸二羟丙酮,循糖代谢途径进行分解或转变为糖。
脂肪细胞及骨骼肌等组织因甘油激酶活性很低,故不能很好利用甘油。
二、脂酸的β-氧化脂酸是人及哺乳动物的主要能源物质。
在O2供给充足的条件下,脂酸可在体内分解成CO2及H2O并释出大量能量,以ATP形式供机体利用。
除脑组织外,大多数组织均能氧化脂酸,但以肝及肌肉最活跃。
1.脂肪酸的活化脂酰CoA的生成脂酸进行氧化前必须活化,活化在线粒体外进行。
内质网及线粒体外膜上的脂酰CoA合成酶在ATP、CoASH、Mg2+存在的条件下,催化脂酸活化,生成脂酰CoA。
脂酸活化后不仅含有高能硫酯键,而且增加了水溶性,从而提高了脂酸的代谢活性。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第七章 脂类代谢
第二节 三脂酰甘油的分解代谢
Hale Waihona Puke 第七章 脂类代谢Lipid Metabolism
三脂酰甘油的分解代谢
基本知识点
甘油三酯的水解
甘油的氧化分解
脂肪酸的氧化
一、甘油三酯的水解
由图示可知:
三脂酰甘油水解产物为:一分子甘油及三分子脂肪酸。 催化上述反应所需的酶为:三脂酰甘油(TG)脂肪酶、 二脂酰甘油(DG)脂肪酶、一脂酰甘油(MG)脂肪酶。 其中,三脂酰甘油(TG)脂肪酶是限速酶,且受多种 激素的调节,又称激素敏感性脂肪酶。 脂解激素:肾上腺素、去甲肾上腺素、胰高血糖素、肾 上腺皮质激素等能直接激活脂肪组织中的三脂酰甘油脂 肪酶,促进脂肪动员。 抗脂解激素:胰岛素,可使三脂酰甘油脂肪酶活性降低。
脂肪酸氧化产生的能量计算
以软脂酸(16个C原子的脂肪酸)为例:
⑴活化,消耗2ATP,生成软脂酰CoA(细胞质中) ⑵脂酰CoA脱氢:FADH2 ,产生2分子 ATP。 ⑶β-羟脂酰CoA脱氢:NADH,产生3分子ATP。 ⑷β-酮脂酰CoA硫解:乙酰CoA → TCA,12分子ATP (n-2)脂酰CoA → 第二轮β氧化 共7次β氧化,产生7分子FADH2,7分子NADH+H+及8分子乙酰CoA 。 活化消耗: 2分子ATP β氧化产生: 7×(2+3)ATP = 35 8个乙酰CoA: 8×12ATP = 96 净生成: 131-2=129分子ATP
•脂酰CoA的转运
脂肪酸活化是在胞液中进行的,而催化脂肪酸氧化的 酶系在线粒体基质内,所以,需以脂酰肉碱形式转运至线 粒体内。
• 线粒体内膜外侧(胞质侧):肉碱脂酰转移酶 Ⅰ催化,脂酰CoA将脂酰基转移给肉碱的 β 羟基, 生成脂酰肉碱。 • 线粒体内膜:移位酶将脂酰肉碱移入线粒体并 将肉碱移出线粒体。 • 线粒体内 : 肉碱脂酰转移酶Ⅱ催化,使脂酰基 又转移给CoA,生成脂酰CoA和游离的肉碱。
二、甘油的氧化分解
生成磷酸二羟丙酮后,进入糖酵解途径继续氧化分解 生成CO2和H2O并释放能量,少量也可在肝糖异生为G或Gn。
三、脂肪酸的氧化
脂肪酸氧化分解的部位:原核生物在细胞溶胶 中进行,真核生物在线粒体基质进行。 脂肪酸氧化供能的组织:除脑细胞和成熟红细胞外 的大多数组织都能利用脂肪酸氧化功能,以肝和肌 组织最为活跃。 脂 肪 活化与转运 酸 氧 β-氧化 化 过 乙酰CoA的彻底氧化 程
脂酰CoA的β-氧化
硫 解
再 脱 氢
加 水
脱 氢
脂酰CoA的β-氧化过程
脂 肪 酸 活 化
AMP ATP
脂肪酸
FADH2 H2O
+
脂酰CoA
CoA-SH 硫激酶
PPi
脂酰CoA脱氢酶 А、β烯脂酰CoA 烯脂酰CoA水化酶 β -羟脂酰CoA β-羟脂酰CoA脱氢酶 β-酮脂酰CoA
NADH CoA-SH
脂肪酸的活化与转 运
脂肪酸的活化:
Mg2+
RCOO- + CoA-SH + ATP---→RCO-S-CoA + AMP + PPi
其中,此反应中用到的酶是:脂酰-CoA合成酶。 活化生成的脂酰-CoA为高能化合物,且水溶性强,提高了代谢 活性。 1分子脂肪酸活化成脂酰CoA,实际上等于消耗了2分子ATP。
思考题:
若是硬脂酸(18个C原子组成的脂肪酸)氧化 供能,会产生多少分子的ATP呢?
β-酮脂酰CoA硫解酶
乙酰CoA
少2个C原子的脂酰-CoA
少2个碳原子的脂酰-CoA再经过脱氢、加水、 再脱氢、硫解,进行又一次β-氧化,如此反复 进行,直至脂酰-CoA完全氧化为乙酰CoA。
可见, β-氧化的终产物为乙酰CoA。
乙酰CoA的彻底氧化
生成的乙酰CoA经TCA循环被彻底氧化,生成CO2和 H2O,并释放能量,产生12分子的ATP。 脂肪酸氧化分解可产生多少分子ATP呢?