去泛素化
去泛素化在NF-κB信号通路调节中的作用

去泛素化在NF-κB信号通路调节中的作用Edward W Harhaj1, Vishva M Dixit21Department of Microbiology and Immunology, Sylvester Comprehensive Cancer Center, The University of Miami, Miller School of Medicine, 1550 NW 10 Avenue, Miami, FL 33136, USA; 2Department of Physiological Chemistry, Genentech Inc., 1 DNA Way,South San Francisco, CA 94080, USA核因子kappa B(NF-κB)是一种非常关键的调控因子,参与多种生物功能,包括先天和适应性的免疫以及细胞的生存等。
NF-κB的激活严格受到一种慢性信号的调节,这一信号会导致持续性的炎症以及癌症。
由关键信号分子E3连接酶介导的泛素化途径对NF-κB信号通路具有重要的调控作用。
去泛素化(DUBs)可抵抗E3连接酶,因此在下调NF-κB信号通路以及维持稳态时,其有重要的作用。
通过特异的去泛素化,如A20和CYLD,可以了解NF-κB信号通路的下调途径,进而为慢性疾病以及癌症提供治疗的可能。
核因子kappa B(NF-κB)/Rel代表了一种真核转录因子家族,这一家族是基因的主要调控因子,可以控制先天和适应性的免疫反应。
NF-κ B 家族的蛋白都含有约300个氨基酸组成的RHD 结构域,这一结构域包括DNA结合区、核定位区以及二聚化区。
NF-κ B 家族成员可以形成同源或异源的二聚体,包括p65/RelA, c-Rel, RelB, p105/p50 以及p100/p52。
在细胞质中,NF-κB被抑制蛋白或IκBs抑制,所以呈现非活性状态,这些抑制蛋白可以阻止NF-κB的核定位以及与DNA的结合。
去泛素化酶
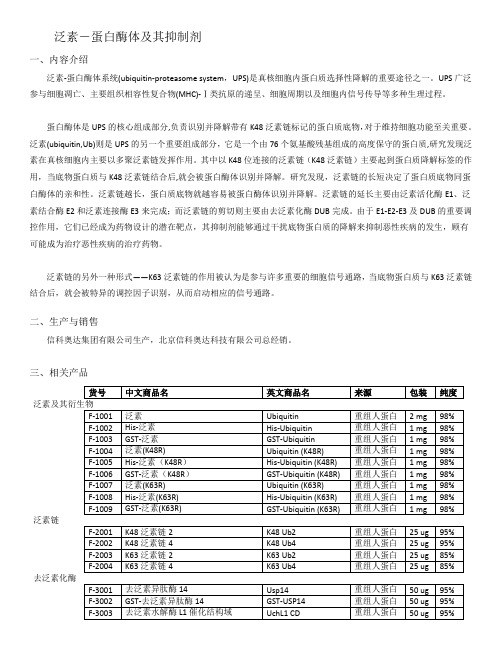
泛素-蛋白酶体及其抑制剂一、内容介绍泛素-蛋白酶体系统(ubiquitin-proteasome system,UPS)是真核细胞内蛋白质选择性降解的重要途径之一。
UPS广泛参与细胞凋亡、主要组织相容性复合物(MHC)-Ⅰ类抗原的递呈、细胞周期以及细胞内信号传导等多种生理过程。
蛋白酶体是UPS的核心组成部分,负责识别并降解带有K48泛素链标记的蛋白质底物,对于维持细胞功能至关重要。
泛素(ubiquitin,Ub)则是UPS的另一个重要组成部分,它是一个由76个氨基酸残基组成的高度保守的蛋白质,研究发现泛素在真核细胞内主要以多聚泛素链发挥作用。
其中以K48位连接的泛素链(K48泛素链)主要起到蛋白质降解标签的作用,当底物蛋白质与K48泛素链结合后,就会被蛋白酶体识别并降解。
研究发现,泛素链的长短决定了蛋白质底物同蛋白酶体的亲和性。
泛素链越长,蛋白质底物就越容易被蛋白酶体识别并降解。
泛素链的延长主要由泛素活化酶E1、泛素结合酶E2和泛素连接酶E3来完成;而泛素链的剪切则主要由去泛素化酶DUB完成。
由于E1-E2-E3及DUB的重要调控作用,它们已经成为药物设计的潜在靶点,其抑制剂能够通过干扰底物蛋白质的降解来抑制恶性疾病的发生,顾有可能成为治疗恶性疾病的治疗药物。
泛素链的另外一种形式——K63泛素链的作用被认为是参与许多重要的细胞信号通路,当底物蛋白质与K63泛素链结合后,就会被特异的调控因子识别,从而启动相应的信号通路。
泛素链去泛素化酶泛素链接酶1、产品质量有保障——产品均由专家研发生产,均可提供试用装2、性价比高——价格比国外同样的产品便宜30%左右,关税、国际运费等均省了3、订货周期短——备有大量现货4、技术健全——泛素相关产品齐全,技术成熟5、相信自己——中国自己的产品跟国外的一样好。
一个新的蛋白质去泛素化酶的鉴定以及功能

05
总结与展望
研究结论
01
鉴定了一个新的蛋白质去泛素 化酶,并对其进行了初步的功 能研究。
02
发现该酶具有去泛素化活性, 能够恢复某些被泛素化的蛋白 质的功能。
03
揭示了该酶在细胞生长、凋亡 及肿瘤发生等生物学过程中的 作用。
研究不足与展望
仍需进一步研究该酶的具体作用机制和 调节方式。
需要明确该酶在人体中的生理和病理作用, 以及其在疾病诊断和治疗中的潜在应用价值 。
03
生物样本收集
收集肿瘤组织及其对应的 正常组织,用于后续实验 。
试剂与仪器
准备相关试剂和仪器,如 抗体、酶标仪等。
实验操作
进行蛋白质提取、免疫印 迹等实验操作。
实验结果与分析
蛋白质表达差异
01
比较肿瘤组织和正常组织中蛋白质的表达情况,发现某些蛋白
质在肿瘤组织中的表达水平有明显变化。
目标蛋白质筛选
研究目的和方法
• 本研究的目的在于鉴定一个新的蛋白质去泛素化酶,并对其 功能进行深入探究。首先,我们将通过生物信息学方法预测 和筛选可能的去泛素化酶;接着,我们将通过实验验证预测 结果,并进一步分析其底物特异性、活性调节机制以及在细 胞中的功能等。
02
蛋白质去泛素化酶的鉴定
实验材料与方法
01
02
03
蛋白质去泛素化酶的功能 研究
实验材料与方法
实验材料
选择本实验室已建立的蛋白质去泛素化酶基因敲除细胞系作 为研究对象,采用已有的蛋白质印迹、免疫共沉淀等方法进 行实验。
实验方法
通过基因敲除技术,构建蛋白质去泛素化酶基因敲除细胞系 ;采用蛋白质印迹法检测敲除前后的蛋白质去泛素化酶表达 情况;采用免疫共沉淀法检测敲除前后的蛋白质去泛素化酶 与目标蛋白的相互作用情况。
去泛素化酶USP19通过对Beclin-1去泛素化调控细胞自噬和抗病毒反应

去泛素化酶USP19通过对Beclin-1去泛素化调控细胞自噬和抗病毒反应细胞自噬是真核细胞内一套高度保守的降解系统,它能够清除毒害性质的蛋白堆积物、衰老的线粒体以及侵染机体的病原微生物。
根据其类型,自噬可以划分为巨自噬、微自噬和分子伴侣介导的自噬三种形式。
巨自噬,也就是常规意义上的自噬,它会形成双层膜结构包裹底物的自噬体,随后自噬体与溶酶体进行融合,在溶酶体中酸性水解酶的作用下将底物降解。
在哺乳动物细胞中,一系列与酵母同源的自噬相关蛋白(ATG)组装成自噬复合体精细操控自噬的过程。
自噬在细胞和生理水平的生物学过程中都发挥至关重要的作用,此外,自噬相关蛋白不但在自噬过程中起作用,而且还参与免疫反应的调节,例如:炎症、抗病毒反应、抗原加工和抗原递呈。
自噬的主要激活剂能够抑制m TOR(mammalian target of rapamycin)信号从而启动自噬。
m TOR信号通路中的关键受体m TORC1复合体抑制ULK1、ATG13、ATG101和FIP200组成的ULK1复合体的活性;而诱导细胞自噬的各种信号能够通过抑制m TORC1的活性,从而激活ULK1复合体,造成自噬的启动,进一步激活下游的Beclin-1复合体。
Beclin-1复合体由Beclin-1、VPS34、VPS15以及ATG14L组成。
Beclin-1复合体的活化导致VPS34催化产生磷脂酰肌醇3磷酸,后者对于自噬体的膜的成核是必需的。
除此之外,在葡萄糖缺失的饥饿条件下,AMPK可以直接磷酸化Beclin-1从而激活VPS34。
Beclin-1是激活VPS34活性进而引发自噬过程的信号中心。
基于Beclin-1在细胞自噬中的核心地位,研究Beclin-1的翻译后水平的调控显得尤为重要。
以往对Beclin-1的研究都集中在磷酸化修饰,对于Beclin-1的泛素化修饰,我们目前还知之甚少。
泛素(ubiquitin)是具有76个氨基酸的高度保守序列的多肽,它们主要在一系列泛素连接酶(E1,E2和E3)的作用下,通过一个或者多个赖氨酸残基连接到蛋白上,从而形成泛素链。
组蛋白的泛素化与去泛素化

a.Histone H2A Monoubiquitination Represses Transcription by Inhibiting PIC formation
•H3K4 methylation is essential for preinitiation complex(PIC) formation. •Ubiquitylation of H2A does inhibit preinitiation complex formation, not in a direct way but indirectly by preventing H3K4 methylation. • Deubiquitylation of histone H2A by USP21 activates transcriptional initiation via trans-histone crosstalk with H3K4 diand tri-methylation.
substrate-specific E3(ligase)
E3主要有两大类: HECT(homologous to the E6-AP carboxyl terminus) 结构域家族,通过与泛素形成催 化作用所必需的硫酯键发挥作用 RING(really interesting new gene)结构域 家族,为E2和底物提供居留位点从而使 E2 催化泛素转移到底物上
X chromosome inactivation, cell cycle progression 2A-DUB :interacts with PCAF USP21: regulator of liver regeneration BAP1: C-terminal hydrolase,repression of HOX genes ✓ H2B deubiquitination Ubp8 :reside within SAGA,Transcription Ubp10 (Dot4):Sir-mediated telomeric and rDNA silencing
泛素化_去泛素化系统的生物化学和生物学功能_李杨

促瘤形成等课题还有待深入研究。
[19] 朱海英 等. 第二军医大学学报, 2005, 26(3): 247—250
[20] Guzman ML et al. Proc Natl Acad Sci USA, 2002, 99: 16220—
参考文献
16225
· 516 ·
《生命的化学》2006 年 26 卷 6 期 CHEMISTRY OF LIFE 2006, 26(6)
治疗的靶点。肿瘤干细胞的基因表达谱也将成为 各种肿瘤的特异性标志。如何能够选择性地杀伤 肿瘤干细胞, 但同时不会抑制正常干细胞而发生
[ 8 ] Bjerkvig R et al. Nat Rev Cancer, 2005, 5(11): 899—904 [ 9 ] Hanahan D et al. Cell, 2000, 100(1): 57—70 [10] Pardal R et al. Nat Rev Cancer, 2003, 3(12): 895—902 [11] van de Wetering M et al. Cell, 2002, 111(2): 241—250
严重的毒副作用已成为学者们研究的热点, 目 前, 已有联合应用去甲氧基柔红霉素和蛋白酶体 抑 制 剂 特 异 性 杀 伤 白 血 病 干 细 胞 的 报 道 [20]。
[12] Lai K et al. Nat Neurosci, 2003, 6(1): 21—27 [13] Dahmane N et al. Development, 2001, 128(24): 5201—5212 [14] Duncan AW et al. Nat Immunol, 2005, 6(3): 314—322 [15] Park IK et al. Nature, 2003, 423(6937): 302—305
去泛素化酶的研究进展

蛋白酶。去泛素化过程是非常精密有序的,它参与 多种重要生命活动,包 括细胞周期调控、基 因 转 录 、 激酶活化、蛋白质降解、D N A 修 复 等 ,去泛素化酶的 异常表达可引发癌症以及神经退行性疾病等多种疾 病 [4]。 1 . 1 泛 素 C 末端水解酶家族
至 今 ,已 有 4 种 U C H 去泛 素 化 酶 ,包 括 。0 1 L 1 、U C H - L 3 、U C H - L 5 和 B A P 1 。泛素 C 末端水解 酶家族(U C H S)属于巯基蛋白酶,其作用是修饰泛素 链 。其催化核心区包含2 3 0 个 氨 基 酸 ,如 U C H - L 1 和 U C H - L 3 [5]。U C H 家族里第一个被发现的酶是 去 泛 素 化 酶 U C H - L 1 ,它可切开泛素链上泛素分子 的 C 端 与 短 肽 构 成 的 化 学 键 ,释 放 泛 素 单 体 。酶 U C H - L 3 的作用原理与酶U C H - L 1 相 似 。U C H 酶 家族中有一个能够协助U C H 酶识别靶蛋白的交叉 环结构[]。 1 . 2 卵巢肿瘤相关蛋白酶家族
卵巢肿瘤相关蛋白酶家族由16个酶蛋白组成, 分 为 4 个 亚 家 族 :O t u b a i n 亚 家 族 、0 T U D 亚 家 族 类 、 A 2 亚 家 族 和 0 T U L I N 亚 家 族 (0 T U L I N ) 。不同 0 T U 家族的中心结构域大小不同,而且这些家族蛋 白酶的氨基酸序列与泛素C 末 端 水 解酶家族、泛素 特 异 性 蛋 白 酶 家 族 不 同 ,对 泛 素 链 的 识 别 具 有 特 异 性 。如 0 T U B 1 和 A 2 0 只 选 择 含 有 K 4 8 位点的泛素 链 ,Cezanne D U B 对 含 K 1 1 位点的泛素链具有特异
去泛素化酶与肿瘤

去泛素化酶与肿瘤唐时珺;陈松;叶茂【摘要】去泛素化是指由去泛素化酶介导的一种蛋白质泛素化的负向调节过程.迄今为止已确定的去泛素化酶有99种,根据其序列和结构的相似性可以分为6大家族,大部分属于半胱氨酸蛋白酶.去泛素化酶的活性直接影响细胞内多种蛋白的周转率、活性、再生以及定位.因此,去泛素化酶对细胞内环境自稳态的维持、蛋白的稳定或降解以及细胞内的信号转导通路起着极其重要的作用.研究表明,去泛素化酶的功能异常与包括肿瘤在内的多种疾病的发生密切相关,以其作为分子靶标,筛选新药物抑制肿瘤的生长已成为抗肿瘤药物开发的热点.【期刊名称】《激光生物学报》【年(卷),期】2014(023)004【总页数】7页(P294-300)【关键词】去泛素化;去泛素化酶;肿瘤【作者】唐时珺;陈松;叶茂【作者单位】湖南大学生物学院分子科学与分子医学实验室,湖南长沙410082;湖南师范大学生命科学学院,湖南长沙410006;湖南大学生物学院分子科学与分子医学实验室,湖南长沙410082【正文语种】中文【中图分类】Q71泛素是由76个氨基酸组成的球形热稳定蛋白。
在真核细胞中,泛素的结构高度保守,蛋白质接受一些信号(如磷酸化、氧化、错误折叠、损伤等)能引发其泛素-蛋白酶体降解途径。
泛素-蛋白酶体降解途径是指靶蛋白通过一系列的酶促反应被泛素链标记进而在蛋白酶体中降解,而泛素分子则可以被释放出来重复利用的过程。
酶促反应包括三步:泛素激活酶E1激活游离的泛素分子并形成E1-泛素复合物;泛素结合酶E2催化E1-泛素复合物形成高能磷酸键相连的E2-泛素复合物;泛素连接酶E3识别靶蛋白并将泛素链连接到靶蛋白上,真核生物中还存在一些E2(如RAD6/UBC2等),不需要E3的参与就能直接识别靶蛋白并标记上泛素链[1]。
近来研究表明,泛素化是一个可逆的过程,去泛素化酶能够水解泛素羧基末端的异肽键,将泛素分子特异性地从底物蛋白或者前体蛋白上水解下来,逆转泛素化过程。
蛋白质泛素化、去泛素化与肿瘤发生的关系

蛋白质泛素化、去泛素化与肿瘤发生的关系【关键词】泛素化泛素-蛋白酶体途径去泛素化去泛素化酶肿瘤泛素蛋白水解酶体途径(Ubiquitin proteasome pathway, UPP)是细胞质和核内蛋白ATP 依托性的非溶酶体降解途径,高效并高度选择性地进行细胞内蛋白转换,还参与某些重要蛋白质的编译后修饰和改造。
因此,对维持细胞正常生理功能具有十分重要的意义。
但随着对UPP研究的深切,发觉蛋白泛素化调剂是一个可逆的进程。
细胞内同时还存在一些特异的去泛素化蛋白酶进行负向调剂[1]。
1 泛素及其功能泛素是由76个氨基酸组成的球形热稳固蛋白,其结构在真核细胞中高度保守,在不同生物体间组成泛素的氨基酸序列不同很小。
泛素是通过一系列泛素启动酶的作用而与靶蛋白连接的。
泛素-蛋白酶体系统的组成(1)泛素:分子量约,普遍存在于真核细胞。
泛素链与蛋白底物的结合形成被蛋白酶体降解的识别信号。
(2)泛素活化酶E1:通过半胱氨酸残基与泛素C端活化的甘氨酸残基形成硫酯键。
(3)泛素转移酶E2:以泛素结合酶方式起作用,活性部位为半胱氨酸。
(4)泛素连接酶E3:识别被降解的蛋白并将泛素连接到底物上。
(5)26S蛋白酶体():蛋白酶体定位于胞核和胞质内。
26S蛋白酶体是由2个环状的19S亚单位和1个20S微粒体状的亚单位组成[2]。
蛋白酶体被以为是细胞内的再生与回收中心,泛素化的靶蛋白在此被分解为短肽和氨基酸,而泛素被回收再利用。
泛素-蛋白酶体降解途径第一时期为泛素与蛋白底物的彼此作用:(1)高能硫酯键E1泛素复合物的形成。
在ATP参与下,游离的泛素被E1激活,即E1的半胱氨酸残基与泛素的C结尾甘氨酸残基形成高能硫酯键。
(2)活化泛素(E1泛素复合物)转移到E2上,释放出E1,形成高能键E2泛素复合物。
(3)底物(被磷酸化、氧化、错误折叠或与辅助蛋白结合的蛋白质)被E3识别并与之结合。
(4)E2泛素复合物上的泛素转移到E3上,形成高能键复合物。
组蛋白的泛素化与去泛素化

12
DNA damage repair
Schematic depiction of histone ubiquitination
Methods. 2011 Jul; 54(3): 315–9325.
Histone deubiquitinating enzyme(DUBs)
✓ H2A deubiquitination USP 16: HOX gene silencing, DNA damage repair
Histone ubquitination&deubquitination
1
The structure of nucleosome
组蛋白是一种11-15KDa 的碱性蛋白,与DNA相 互作用构成核小体。 核心组蛋白的尾巴伸出 来,受到广泛的修饰。
2
Ubquitin
泛素(ubiquitin,Ub)是 高度保守的、含76个氨 基酸的蛋白质,分子量 为8.5 kDa,在真核生物 体内广泛存在。 泛素分子氨基端1-72位 点的氨基酸残基形成一 个紧密球状结构,紧靠 羧基端的4个氨基酸残基 是随机盘绕的(randomcoiled)。
deubquitinase • Histone modifications crosstalk • Involved in multiple cellular events---gene expression
regulation、DNA damage response and repair、 female X chromosome inactive and so on
蛋白质泛素化和去泛素化的机制

蛋白质泛素化和去泛素化的机制蛋白质泛素化和去泛素化是细胞生物学的重要机制,通过这两种机制,细胞可以控制蛋白质的稳定性、代谢、功能和互作等,从而维持生命活动的正常进行。
在本文中,我们将介绍蛋白质泛素化和去泛素化的基本原理、机制和作用,为读者提供一些基础知识和参考。
一、蛋白质泛素化的基本原理和机制蛋白质泛素化指的是一种特殊的蛋白质修饰,它通过将一个小分子泛素(Ub)与目标蛋白结合,从而改变目标蛋白的功能或命运。
泛素本身是由多个氨基酸残基序列(GG)组成的肽链,这些肽链可以与目标蛋白上的特定赖氨酸残基结合,并形成泛素化修饰。
泛素化修饰可以通过三种主要的机制实现,包括单泛素化、多泛素化和链型泛素化。
单泛素化的含义是指某个蛋白质上只有一个泛素分子被修饰。
多泛素化则是指同一蛋白质上有多个泛素分子被修饰。
链型泛素化是指泛素分子之间形成共价二聚体,三聚体或更高级别的链,从而形成链型泛素化修饰。
链型泛素化包括K48-、K63-和K11-链型泛素化等,它们具有不同的结构和作用。
泛素化修饰可以通过一系列酶的协同作用来完成。
首先,泛素激活酶(E1)将泛素与ATP结合,形成一个泛素-E1复合物。
然后,泛素转移酶(E2)将泛素-E1复合物转移到某个特定的E3泛素连接酶上,E3酶与目标蛋白上的特定残基形成复合物,从而实现泛素化修饰。
E3酶还可以选择泛素-E2复合物的结构,以实现单泛素化、多泛素化或链型泛素化等机制。
最后,泛素化修饰的目标蛋白可以通过蛋白酶体或溶酶体等途径被降解或转运。
二、蛋白质去泛素化的机制和作用蛋白质去泛素化是指将已经泛素化的蛋白质上的泛素修饰群体去除的机制。
相比蛋白质泛素化,蛋白质去泛素化的机制更为复杂,因为细胞中存在多种不同类型的去泛素化酶(DUB),它们具有不同的结构、特点和功能。
根据分子机制,DUB可以分为两类,即结构特异性DUB和作用特异性DUB。
结构特异性DUB是指只能识别和切割泛素化群体的特定结构,例如K48-链、K63-链、K11-链等,并能在其中识别和选择特定的链结并将其去除。
Cycloheximide-泛素特异性蛋白酶2a通过去泛素化和稳定RAB1A

【AbMole科研】泛素特异性蛋白酶2a通过去泛素化和稳定RAB1A促进肝癌进展AbMole精研抑制剂十年,最新的科研动态不断与您分享。
本期与您分享的是:泛素特异性蛋白酶2a通过去泛素化和稳定RAB1A促进肝癌进展去泛素化是泛素化的逆过程,它由去泛素酶(DUBs)催化,去泛素酶去除目标蛋白中的泛素,从而阻止其被蛋白酶体降解。
此前,去泛素化被发现参与肝细胞癌(HCC)的发育和进展中。
然而,对于去泛素化在这类癌症的发展和确切作用,人们知之甚少。
研究中利用HCC组织和组织芯片检测DUB泛素特异性蛋白酶2a (USP2a)的表达。
在体外细胞和体内动物模型中评估了USP2a在HCC进展中的关键作用。
通过LCMS/MS分析确定USP2a 在HCC细胞中的潜在靶点,然后研究USP2a对靶点蛋白稳定性和泛素状态的调控。
结果发现,USP2a在HCC组织中显著上调,且高表达与不良预后呈正相关。
随后,进一步研究发现,在体外和体内,沉默USP2a会抑制肝癌细胞的增殖、迁移和侵袭,而外源USP2a 过表达则会产生相反的作用。
LC-MS/MS分析显示,RAB1A是ER和高尔基体囊泡运输系统的关键调控因子,是USP2a在HCC细胞中的潜在靶点。
此外,研究人员发现USP2a可以使RAB1A 去泛素化并稳定,防止其降解,而这一过程是USP2a诱导HCC进展所必需的。
总结来说,研究数据表明,USP2a可以通过去泛素化和稳定RAB1A来促进HCC进展。
这一观察结果表明,DUB靶向治疗可能成为改善肝癌治疗的一种新方法。
Cycloheximide(放线菌酮)(Abmole, M4879,纯度>99%) 是一种蛋白生物合成抑制剂, IC50值为532.5 nM。
作者在研究过程中,采用放线菌酮(CHX) chase法检测RAB1A的半衰期。
简而言之,细胞与蛋白质合成抑制剂CHX (100 μg/ml)处理不同的时间。
然后提取处理后的细胞蛋白,进行Western blotting检测RAB1A的表达。
去泛素化机制

去泛素化机制1. 引言泛素化是一种重要的细胞生物学过程,它在细胞内调控蛋白质的稳定性、活性和功能发挥等方面起着关键作用。
泛素化通过共价地附着小蛋白泛素于目标蛋白上,从而调控目标蛋白的命运。
然而,在某些情况下,细胞需要去除或逆转这种泛素化修饰,以实现特定的调控效果。
因此,去泛素化机制也变得非常重要。
2. 去泛素化酶家族去泛素化是由一类特殊的酶,即去泛素化酶(deubiquitinase, DUBs)来完成的。
DUBs是一类能够切断或去除蛋白质上泛素修饰的酶,它们通过加水解除泛素与目标蛋白之间的共价键,从而去除泛素。
根据其结构和去泛素机制的不同,DUBs可分为结构域特异性DUBs和泛素结构特异性DUBs。
2.1 结构域特异性DUBs结构域特异性DUBs包括UBP/USP家族、UCH家族和MJD家族等。
UBP/USP家族是最大的DUB家族,其成员拥有一个保守的结构域,即UBP结构域。
UBP/USP家族DUBs 通过结构域与泛素结合,并通过水解作用去除目标蛋白上的泛素修饰。
UCH家族DUBs则具有独特的结构域UCH结构域,其机制与UBP/USP家族DUBs有所不同。
MJD家族DUBs则是一类与遗传性神经退行性疾病相关的DUBs,其结构域和去泛素机制也有所特殊。
2.2 泛素结构特异性DUBs泛素结构特异性DUBs是一类通过与泛素结构上特定位点的相互作用来去除泛素修饰的DUBs。
这类DUBs包括OTU家族、JAMM/MPN+家族和MINDY家族等。
OTU家族DUBs通过其OTU结构域与泛素结构上的特定位点相互作用,从而去除泛素修饰。
JAMM/MPN+家族DUBs则通过其JAMM/MPN+结构域与泛素结构上的特定位点相互作用,实现去泛素化。
MINDY家族DUBs则是最新发现的一类泛素结构特异性DUBs,其去泛素机制和特定结构域有所不同。
3. 去泛素化的生理功能去泛素化在细胞生物学中具有重要的生理功能,它参与了多个细胞过程的调控。
最新综述全面总结:去泛素化酶在帕金森病中的作用

最新综述全面总结:去泛素化酶在帕金森病中的作用已有研究表明,线粒体功能障碍(mitochondrial dysfunction)与帕金森病(PD)直接关联,而线粒体自噬(mitophagy)更是与PD密切相关。
线粒体自噬可以选择性地降解线粒体,是线粒体质量控制(mitochondrialquality control)的主要途径之一。
它是由线粒体表面蛋白的泛素修饰触发的,因此线粒体自噬受到去泛素化的抑制。
近年来,许多去泛素化酶(deubiquitinase enzymes,DUBs)被认为是PD的潜在治疗靶点,同时广泛的研究表明,DUBs之间存在相互影响,这可能对PD的进展或阻碍具有协同效应。
基于相关研究结果,在frontiers in Physiology杂志上发表的最新综述'Deubiquitinating Enzymes in Parkinson's Disease'特别介绍了几个在PD领域具有治疗价值的DUBs。
1帕金森病与线粒体质量控制众所周知,PD的主要病理改变是黑质致密部多巴胺能神经元的特异性丢失。
与其他类型细胞不同的是,多巴胺能神经元依赖于特定亚型的钙离子通道来维持其自主起搏活动和多巴胺代谢的高能需求,因此其显得尤为脆弱,而线粒体有氧呼吸产生的副产物ROS就是危害多巴胺能神经元的主要有毒物质。
已有研究证实,PD病患的PINK1和Parkin等线粒体质量控制相关基因发生突变,这一事实进一步支持了PD和线粒体之间的关系。
在PINK1/Parkin通路中,Parkin作为泛素连接酶,当线粒体去极化时,PINK1招募Parkin到线粒体,进而对一些特定的蛋白进行泛素化。
随后,线粒体表面的泛素化蛋白通过募集自噬受体直接与自噬隔离膜相互作用,已经确定的几个线粒体自噬受体包括——p62/SQSTMA、FUNDC1、FKBP8和BNIP3L/Nix等,这些受体的共同特点是包含MAP1LC3/LC3相互作用区域。
去泛素化酶结合蛋白酶体

去泛素化酶结合蛋白酶体去泛素化酶结合蛋白酶体?哎呀,这名字听着就让人头大对不对?你可能觉得这是不是又一个超级复杂的生物学概念,什么都不懂也没关系,我来给你捋捋清楚。
说白了,这个东西就是细胞里的一场大清理行动。
想象一下,就像是你家乱七八糟的房间,墙角堆着不值钱的旧物,床底下有过期的垃圾。
每当有这些“废物”堆积起来,家里就会有些不太舒服,不想让生活质量下降,必须清理。
这时候就需要我们细胞中的“清洁工”来帮忙了,而这帮清洁工就是——去泛素化酶和蛋白酶体。
嗯,说到去泛素化酶和蛋白酶体,这俩名字听起来特别拗口,但其实它们做的事儿可简单了。
去泛素化酶就像是给脏东西做清理的前期准备,它们的工作是把一些垃圾上面的小标签——泛素,给去掉。
你看,泛素就像是一张“淘汰”标签,贴在废旧物品上,告诉蛋白酶体:“嘿,快来处理它!”去泛素化酶就像是个“拆标签工”,把这个泛素标签轻轻松松地拆掉。
然后,这些蛋白就会乖乖地走到蛋白酶体去“见老板”——蛋白酶体会把它们搞定。
怎么搞呢?简单点说,蛋白酶体就像是个大破碎机,把这些不需要的蛋白质给分解掉,分解成小块,然后再做进一步的处理。
嘿,这一系列的动作可不简单,基本上是细胞“自我清理”的高效模式。
有点像我们生活中的垃圾回收站。
你知道,平时家里如果垃圾桶没及时倒,慢慢堆积就会臭气熏天,搞得空气也不清新。
可是如果你定期清理,垃圾就能及时处理,不但环境干净,空气也好闻。
所以这些酶其实是细胞里的一种“环保力量”,它们通过去除不需要的蛋白质,维持细胞的健康,避免病变和其他麻烦。
那种看似不起眼的去泛素化酶,哇,它们可真是功不可没,它们是蛋白质处理的“前台接待”,负责给垃圾贴标签,然后送去处理中心。
而蛋白酶体呢,它就是那个“大清洗”环节的真正大佬。
你可以想象,蛋白酶体就是一个细胞的超级“粉碎机”,把所有不需要的、过期的、损坏的蛋白质都一股脑儿给“吃掉”。
这些蛋白质通过进入蛋白酶体的入口后,被一层层的切割分解,最后变成了简单的小氨基酸,再被细胞用作新的“建筑材料”。
去泛素化的方法

去泛素化的方法
泛素化是一种重要的细胞信号传递过程,它涉及到蛋白质分解、DNA修复、细胞周期调控等多个生物学过程。
然而,当泛素化过程异常时,会引发多种疾病,如肿瘤、自身免疫疾病等。
因此,去泛素化成为了治疗多种疾病的新策略之一。
目前,已经发现了多种去泛素化的方法。
其中,最主要的有以下几种:
1. 酶促去泛素化:利用特定的酶,将泛素从特定的蛋白质上剪
切下来,达到去泛素化的效果。
目前已经发现了多种酶促去泛素化的酶,如USP7、OTUB1等。
2. 蛋白质组学筛选:利用高通量的蛋白质组学技术,筛选出能
够结合到泛素化蛋白质上的小分子化合物,从而达到去泛素化的效果。
目前已经有多个小分子化合物被发现,如PR-619、WP1130等。
3. RNA干扰技术:利用RNA干扰技术,特异性地降低去泛素化
酶的表达,从而达到去泛素化的效果。
这种方法具有较高的特异性和选择性,但也存在许多技术上的挑战。
总的来说,去泛素化是一个新颖的治疗策略,具有广阔的应用前景。
未来,随着对泛素化过程的深入理解和技术的不断更新,相信会有更多的方法被发现和应用。
- 1 -。
otud去泛素化酶的降解

otud去泛素化酶的降解
摘要:
1.OTUD 的作用
2.OTUD 降解的过程
3.OTUD 降解的影响
4.OTUD 降解的研究意义
正文:
OTUD,全称为Ovarian Tumor Domain Containing Ubiquitin Aldehyde Binding Protein 1,是一种泛素羧化酶,主要作用于去泛素化酶,从而参与到蛋白质的降解过程中。
OTUD 去泛素化酶的降解过程主要分为以下几个步骤:首先,OTUD 会与去泛素化酶结合,形成一个稳定的复合物;然后,OTUD 会将复合物上的泛素分子去除,使得去泛素化酶的活性得到恢复;最后,恢复活性的去泛素化酶可以继续参与到其他蛋白质的降解过程中。
OTUD 降解的影响主要体现在对细胞内蛋白质稳态的调控上。
通过调控去泛素化酶的活性,OTUD 可以影响到许多生物学过程,包括细胞生长、细胞凋亡、DNA 损伤修复等。
因此,OTUD 降解对于细胞内环境的稳定至关重要。
OTUD 降解的研究意义主要体现在两个方面:一是对生命过程的理解,通过研究OTUD 降解,我们可以更深入地理解细胞内蛋白质降解的机制,从而对生命过程有更深入的理解;二是对疾病的治疗,由于OTUD 降解对细胞内环境的稳定具有重要作用,因此,研究OTUD 降解可能为疾病的治疗提供新
的思路和方法。
USP14调控RIG-IK63去泛素化抑制RIG-I活化诱导的抗病毒固有免疫应答

USP14调控RIG-IK63去泛素化抑制RIG-I活化诱导的抗病毒固有免疫应答固有免疫系统是人体抵抗外来病原微生物感染的第一道防线。
在病毒感染过程中,模式识别受体通过识别病毒中的病原相关分子模式并活化启动相关信号通路,从而抑制、抵抗病毒的感染。
RLR是机体识别病毒RNA的主要受体,RIG-I能够识别单链RNA病毒,双链RNA 病毒以及具有5’-pp和5’-ppp的平末端RNA病毒,是疱疹性口炎病毒等RNA病毒的主要受体。
泛素化修饰是细胞中蛋白质翻译后修饰的重要方式之一,是高度可逆的。
泛素化修饰发生在细胞活动的各个阶段,在固有免疫应答中具有重要的作用。
蛋白质的泛素化修饰类型包括K6,K11,K27,K29,K33,K48,K63几种,其中K48与K63多聚泛素化修饰的研究相对较为明确。
K63泛素化修饰一般与底物蛋白的活性具有紧密关系,如RIG-I在病毒感染后,需要经过K63泛素化修饰后才能够活化,进而激活下游信号通路。
去泛素化修饰是蛋白质泛素化修饰的逆过程,由去泛素化酶催化。
因此,去泛素化酶在蛋白质翻译后修饰中也具有重要的作用。
在前期实验中,我们用RNA病毒-疱疹性口炎病毒-感染RIG-I稳定过表达的HEK293T细胞,利用免疫共沉淀实验和质谱分析,筛选到包括泛素特异性蛋白酶14在内的能够与RIG-I相互作用的去泛素化酶分子。
该结果提示泛素特异性蛋白酶14可能参与调控RIG-I活化诱导的抗病毒免疫应答。
在本研究中,我们发现泛素特异性蛋白酶14在抗病毒固有免疫应答中发挥负向调控作用。
在小鼠腹腔巨噬细胞及人诱导分化的巨噬细胞系THP1细胞中,干扰泛素特异性蛋白酶14基因可以明显提高RIG-I活化诱导的I型干扰素、IL-6和TNF-α的产生,同时抑制疱疹性口炎病毒的复制。
同样在这两种细胞中,IU1抑制泛素特异性蛋白酶14的活性后也会显著提高RIG-I信号触发的的Ⅰ型干扰素、IL-6和TNF-α的表达,同时抑制疱疹性口炎病毒的复制。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
[分享]讲座教授朱健康《Nature》文章解析表观遗传学[复制链接]
bbioo
Administrator
积分
17006
秀蛋白
3487 个
秀基因
7845 个
帖子
367
楼主
发表于 2009-8-26 08:21 |只看该作者|倒序浏览
|打印
摘要:来自美国加州大学河滨分校植物科学系,植物分子生
物学中心,普渡大学园艺及园林系的研究人员发现了组蛋白
H2B去泛素化(deubiquitination)在表观遗传DNA甲基化和
异染色质沉默方面的调控作用,在DNA甲基化修饰与泛素化之
间建立了新联系,也为DNA甲基化修饰研究提出了新观点,这
一研究成果公布在6月7日《Nature》上。
生物通报道:来自美国加州大学河滨分校植物科学系,植物分
子生物学中心,普渡大学园艺及园林系的研究人员发现了组蛋
白H2B去泛素化(deubiquitination)在表观遗传DNA甲基化
和异染色质沉默方面的调控作用,在DNA甲基化修饰与泛素化
之间建立了新联系,也为DNA甲基化修饰研究提出了新观点,
这一研究成果公布在6月7日《Nature》上。
这一文章的通讯作者是加州大学河滨分校的朱健康教授,其早
年毕业于中国农业大学,现任加州大学河滨分校教授,整合基
因研究所所长,2005年被聘为中国农业大学长江学者讲座教
授。
原文检索:
Nature 447, 735-738 (7 June 2007) |
doi:10.1038/nature05864; Received 23 February 2007;
Accepted 24 April 2007
Control of DNA methylation and heterochromatic silencing
by histone H2B deubiquitination
[Abstract]
在基因组中除了DNA和RNA序列以外,还有许多调控基因的信
息,它们虽然本身不改变基因的序列,但是可以通过基因修饰,
蛋白质与蛋白质、DNA和其它分子的相互作用,而影响和调节
遗传的基因的功能和特性,并且通过细胞分裂和增殖周期影响
遗传,这就是表观遗传学(epigenetics)。
在表观遗传学研究中,小分子RNAs(Short interfering RNAs,
siRNAs)可以引导DNA甲基化,以及异染色质组蛋白修饰,导
致序列特异性转录基因沉默。
目前已知动物和酵母组蛋白H2B
能单泛素化(monoubiquitinate),这能调控H3组蛋白的甲
一领域的一个未解之谜。
在这篇文章中,研究人员发现一种拟南芥去泛素化酶
(deubiquitination enzyme)的突变:SUP32/UBP26能减少
组蛋白H3赖氨酸9的去甲基化,抑制siRNA指导的DNA甲基
化,以及转基因和转座子的异染色质沉默。
同时研究人员也发现拟南芥组蛋白H2B在赖氨酸143位可以发
生单泛素化,并且在sup32突变株中,H2B泛素化及H3赖氨
酸4的三甲基化的水平也增加了。
这些都说明SUP32/UBP26引
起的H2B去泛素化是异染色质组蛋白H3甲基化和DNA甲基化
的一种必要元素。
(生物通:张迪)
附:
朱健康教授
1987年毕业于中国农业大学土化系
1990年取得加州大学河畔分校植物学硕士学位
1993年获得普渡大学植物生理学博士学位
1993--1996朱健康随后任教于纽约市洛克斐勒大学与阿拉
巴马州奥本大学,教授植物生物学
1996年转赴亚利桑那大学吐桑分校任教,2000年成为该校植
物学系教授
现任美国加州大学河滨分校教授,整合基因研究所所长,兼任
国际著名学术刊物《Plant Physiology》、《Plant Molecular
Biology》副主编。
朱健康教授为植物抗逆分子生物学领域世界级领军人物之一,
其及其领导的实验室在植物抗旱、耐盐与耐低温方面的研究硕
果累累,在国内外享有声誉,他的实验室已在《Cell》、
《Science》、《Gene&Development》、《Plant Cell》、PNAS
等国际著名学术刊物发表论文40余篇。