【医学PPT课件大全】 蛋白激酶、蛋白磷酸酶与信号转导
合集下载
蛋白激酶和蛋白磷酸酶蛋白激酶

1. IP3/Ca2+途径 IP3引起细胞膜和内质网 膜上的Ca2+通道开放,胞浆Ca2+显著增加, 导致细胞活化。
2. DAG/PKC途径 DAG特异性激活丝/苏 氨酸激酶-PKC家族。
G蛋白 PI(4,5)P2 PLCb PLCg
其它磷脂酶 离子通道
PI3-K PI3
DAG
腺苷酸环化酶
Ca2+ PKC
氨酸磷酸化后可结合SHP2,进而抑制TCR相
关的NFAT活化。
(二) 蛋白激酶和蛋白磷酸酶
蛋白激酶:将磷酸供体分子如ATP的磷酸基 团转移到底物蛋白氨基酸受体的酶类。
蛋白磷酸酶:催化磷酸化蛋白分子的磷酸 酯键发生去磷酸化反应的酶类。
蛋白酪氨酸激酶
受体型蛋白酪氨酸激酶:PLC、
abcddd PI3K、GAP、STAT等
(五)转录因子
NF-kappaB:发现于B细胞kappa轻链转录调 控,受多种蛋白的精细调控
Rel蛋白家族成员组成的同源或异源二聚体,分 部最广的是p50/RelA。Rel同源结构域(RHD) 存在与其他Rel蛋白、DNA、IkappaB结合区以 及核定位序列。
IkappaB磷酸化、泛素化、降解导致NFkappaB活化、入核
最早发现于T细胞的核转录因子家族,主要 参与钙离子内流有关的膜受体信号传导。
NFAT同源结构域含有核定位序列、钙调磷 酸酶作用位点。
主要的活化方式:钙调磷酸酶催化去磷酸化、 核转位、结合DNA
四、信号传递途径
(一) IP3/Ca2+途径和DAG/PKC途径:
受体相关的磷脂酰肌醇磷脂酶g2(PI-PLCg2) 被PTK磷酸化后激活,水解细胞膜内侧的磷 脂酰4,5-二磷酸肌醇(PIP2)成为1,4,5-三磷酸 肌醇(IP3)和1,2-二酰基甘油(DAG)。
2. DAG/PKC途径 DAG特异性激活丝/苏 氨酸激酶-PKC家族。
G蛋白 PI(4,5)P2 PLCb PLCg
其它磷脂酶 离子通道
PI3-K PI3
DAG
腺苷酸环化酶
Ca2+ PKC
氨酸磷酸化后可结合SHP2,进而抑制TCR相
关的NFAT活化。
(二) 蛋白激酶和蛋白磷酸酶
蛋白激酶:将磷酸供体分子如ATP的磷酸基 团转移到底物蛋白氨基酸受体的酶类。
蛋白磷酸酶:催化磷酸化蛋白分子的磷酸 酯键发生去磷酸化反应的酶类。
蛋白酪氨酸激酶
受体型蛋白酪氨酸激酶:PLC、
abcddd PI3K、GAP、STAT等
(五)转录因子
NF-kappaB:发现于B细胞kappa轻链转录调 控,受多种蛋白的精细调控
Rel蛋白家族成员组成的同源或异源二聚体,分 部最广的是p50/RelA。Rel同源结构域(RHD) 存在与其他Rel蛋白、DNA、IkappaB结合区以 及核定位序列。
IkappaB磷酸化、泛素化、降解导致NFkappaB活化、入核
最早发现于T细胞的核转录因子家族,主要 参与钙离子内流有关的膜受体信号传导。
NFAT同源结构域含有核定位序列、钙调磷 酸酶作用位点。
主要的活化方式:钙调磷酸酶催化去磷酸化、 核转位、结合DNA
四、信号传递途径
(一) IP3/Ca2+途径和DAG/PKC途径:
受体相关的磷脂酰肌醇磷脂酶g2(PI-PLCg2) 被PTK磷酸化后激活,水解细胞膜内侧的磷 脂酰4,5-二磷酸肌醇(PIP2)成为1,4,5-三磷酸 肌醇(IP3)和1,2-二酰基甘油(DAG)。
蛋白磷酸化与蛋白激酶幻灯片

因此可同时与两种以上的其他信号分子结 合。 • 同一类结构域可存在于多种不同的信号分 子中。 • 本身均为非催化结构域。
64
2)非受体酪氨酸蛋白激酶的种类 分为11个家族,至少有30个成员。它
们介导多种生长因子受体、细胞因子受体、 淋巴细胞抗原受体以及黏附分子整合素的 信号转导。
65
a)Src激酶家族 Src是原癌基因c-Src的产物,参与抗
protein,GAP)以及crk、abl和vav原癌基
因产物等。
50
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
51
Cterminal PLC SH2 domain
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HC CH2 OH OC
NH HC CH2 OC
O OP O
O
13
(1)蛋白激酶A(protein kinase A,PKA) 即cAMP依赖性蛋白激酶。 全酶存在胞浆,被cAMP激活后,催化亚
7
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
8
9
(二)蛋白激酶的种类
10
11
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
12
86
2)受体酪氨酸激酶激活信号蛋白的机制 一是信号分子的膜转位; 二是通过结构改变而被激活; 三是通过酪氨酸的磷酸化被激活。
64
2)非受体酪氨酸蛋白激酶的种类 分为11个家族,至少有30个成员。它
们介导多种生长因子受体、细胞因子受体、 淋巴细胞抗原受体以及黏附分子整合素的 信号转导。
65
a)Src激酶家族 Src是原癌基因c-Src的产物,参与抗
protein,GAP)以及crk、abl和vav原癌基
因产物等。
50
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
51
Cterminal PLC SH2 domain
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HC CH2 OH OC
NH HC CH2 OC
O OP O
O
13
(1)蛋白激酶A(protein kinase A,PKA) 即cAMP依赖性蛋白激酶。 全酶存在胞浆,被cAMP激活后,催化亚
7
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
8
9
(二)蛋白激酶的种类
10
11
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
12
86
2)受体酪氨酸激酶激活信号蛋白的机制 一是信号分子的膜转位; 二是通过结构改变而被激活; 三是通过酪氨酸的磷酸化被激活。
第五章蛋白激酶

PLC-1的PH domain与IP3结合
血影蛋白的PH domain与膜磷脂结合
e)PTB结构域
PTB结构也可识别一些含磷酸化酪氨 酸的模体。
Shc PTB Domain
非受体酪氨酸蛋白激酶重要结构域的特点:
• 一个信号分子可含有两种以上的结构域, 因此可同时与两种以上的其他信号分子结 合。
(一)蛋白激酶的结构
共同的结构特征:
• 保守的催化结构域/亚基
• 调节结构域/亚基
• 其他功能结构域
PKC
1. 催化结构域/亚基
催化核心含有12个高度保守的亚区。
功能: • 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
2. 调节结构域/亚基
p p
ANK ANK ANK ANK
Kinase domain
Mig-2
RTK
paxillin
Integrins
ECM
Generously provided to the CMC by Cary Wu (Apr-03
(9)DNA依赖性蛋白激酶 DNA依赖性蛋白激酶(DNA-dependent protein kinaes,DNA-PK)可磷酸 化许多核蛋白,包括核受体、转录因子、 DNA拓扑异构酶和RNA聚合酶Ⅱ等。 DNA-PK可发生自主磷酸化,其催化 亚基和Ku蛋白都依赖于DNA和ATP而磷酸 化。
(2)受体酪氨酸蛋白激酶
PDGF一类的跨膜受体,可磷酸化靶蛋 白的酪氨酸,因此称为受体酪氨酸蛋白激 酶(receptor tyrosine kinase,RTK)。
1)受体酪氨酸激酶的自主磷酸化位点
常位于受体的羧基末端胞质域的非催 化区,或质膜与酪氨酸激酶结构域之间。 主要功能是与靶蛋白的SH2结构域结合, 激活靶蛋白。 靶蛋白与磷酸化位点的结合依赖于 pTyr附近的氨基酸组成及顺序。
蛋白磷酸化与蛋白激酶(医学相关)

优质课件
37
(7)核糖体S6激酶(S6K)
包括S6KⅠ和S6KⅡ,能催化核糖体 S6蛋白磷酸化。
优质课件
38
优质课件
39
(8)整合素连接激酶
整合素连接激酶(intergrin-linked kinase,ILK)可直接磷酸化PKB/Akt, 其活性依赖PI3K。
优质课件
40
PINCH, ILK domain structures & interacting partners
优质课件
12
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HC CH2 OH OC
NH HC CH2 OC
O OP O
O
优质课件
13
(1)蛋白激酶A(protein kinase A,PKA)
• 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
优质课件
7
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
优质课件
8
优质课件
9
(二)蛋白激酶的种类
优质课件
10
优质课件
11
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
• 广泛分布于各组织的胞质,以Ca2+依赖 的形式从胞质中移位到细胞膜上,此过程 称之为转位。PKC转位是其活化的标志。
【医学PPT课件】蛋白激酶A(PKA)介导的信号通路

• 每个调节亚基都含有2个高亲和性的 cAMP结合部位:A和B,位于肽段的羧 基端. 这2个cAMP结合部位都与蛋白激酶 A全酶的解聚以及激活有关,而且cAMP 的结合与全酶的激活呈强的正协同效应. 首先,cAMP与B部位的结合改变了调节 亚基的构象,从而使A部位更易于结合 cAMP. A部位与cAMP的结合则令调节亚
腺苷酸环化酶的激活和cAMP 的产生
• G蛋白和腺苷酸环化酶(ACase)的活化 • 腺苷酸环化酶的磷酸化抑制与脱敏 • cAMP在胞内水平的调节
• 胞外配基(如激素、核苷酸、炎症因子等)
与腺苷酸环化酶偶联的膜受体相结合, 通过激活性GTP-结合蛋白(Gs)的介导, 诱导了腺苷酸环化酶的活化. ACase通过 水解ATP产生环腺苷酸(cAMP)而提高了 胞内的cAMP水平.
• 在哺乳动物中,依据氨基酸顺序的不同, 催化亚基主要分为两类同工酶:Cα和Cβ, 它们催化的底物具有如下共有序列: XArg-Arg-X-Ser-X (X代表任何1个中性氨 基酸,Arg在激酶的底物识别中有着重要 的作用). PKA的催化亚基含有3个结合部 位:Mg-ATP结合部位,催化部位以及底 物肽结合部位.
• 导致了调节亚基的结构变化而与催化亚 基解离产生1个催化亚基二聚体和2个激 活的催化亚基单体[2]. 其解离反应式 可以表示如下:R2C2+4cAMP R2(cAMP)4+2C*(指活化态,对(ⅰ) 蛋白 激酶A的催化亚基. 蛋白激酶A为一丝
氨酸/苏氨酸蛋白激酶,它的催化亚基 可以使特异底物肽段的Ser或Thr残基磷酸 化.
• 随着克隆技术的快速发展,新发现的蛋 白激酶及其同工酶以几何级数的速度增 长着,对它们的研究也日趋深入. 其中, 蛋白激酶A是最小、最简单,也是研究最 为透彻的蛋白激酶之一.
蛋白激酶

protein,GAP)以及crk、abl和vav原癌基
因产物等。
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
Cterminal PLC SH2 domain
c)SH3结构域
可见于多种胞质信号蛋白及肌动蛋白 结合蛋白中。
SH3识别的部位是一些富含脯氨酸的 区域PXXP。
功能:参与PTK介导的蛋白质间的相 互作用,可能在亚细胞定位和细胞骨架蛋 白相互作用中起作用。
SH3 Domain
d)PH结构域
最初于一种血小板内PKC底物 pleckstrin中发现的结构域,称为 pleckstrin 同源 (pleckstrin homology, PH) 结构域。
PINCH, ILK domain structures & interacting partners
Nck-2
SH3 SH3 SH3
SH2
PINCH
LIM5 LIM4 LIM3 LIM2
LIM1
ILK
pp
ANK ANK ANK ANK PH
RTK
CH-ILKBP/actopaxin parvins/affixin
黏附斑激酶(focal adhesion kinase, FAK)在黏附分子整合素介导的细胞与细 胞外基质的黏附和信号转导中具有起始作 用。
整 合 素 结 构 模 型
Linear structure of FAK
(2)受体酪氨酸蛋白激酶
PDGF一类的跨膜受体,可磷酸化靶蛋 白的酪氨酸,因此称为受体酪氨酸蛋白激 酶(receptor tyrosine kinase,RTK)。
因产物等。
SH2能特异地识别磷酸化的酪氨酸残 基以及磷酸化残基的羧基端氨基酸序列并 与其相互结合。
SH2的主要功能是介导胞质内多种信 号蛋白的相互连接,形成蛋白异聚体复合 物,从而调节信号传递。
Cterminal PLC SH2 domain
c)SH3结构域
可见于多种胞质信号蛋白及肌动蛋白 结合蛋白中。
SH3识别的部位是一些富含脯氨酸的 区域PXXP。
功能:参与PTK介导的蛋白质间的相 互作用,可能在亚细胞定位和细胞骨架蛋 白相互作用中起作用。
SH3 Domain
d)PH结构域
最初于一种血小板内PKC底物 pleckstrin中发现的结构域,称为 pleckstrin 同源 (pleckstrin homology, PH) 结构域。
PINCH, ILK domain structures & interacting partners
Nck-2
SH3 SH3 SH3
SH2
PINCH
LIM5 LIM4 LIM3 LIM2
LIM1
ILK
pp
ANK ANK ANK ANK PH
RTK
CH-ILKBP/actopaxin parvins/affixin
黏附斑激酶(focal adhesion kinase, FAK)在黏附分子整合素介导的细胞与细 胞外基质的黏附和信号转导中具有起始作 用。
整 合 素 结 构 模 型
Linear structure of FAK
(2)受体酪氨酸蛋白激酶
PDGF一类的跨膜受体,可磷酸化靶蛋 白的酪氨酸,因此称为受体酪氨酸蛋白激 酶(receptor tyrosine kinase,RTK)。
蛋白激酶与磷酸酶ppt课件

2019/9/4
2
磷酸化修饰的氨基酸
2019/9/4
3
蛋白质的磷酸化修饰
导致受体蛋白的激活或失活; <2% 蛋白质的磷酸化修饰发生在Tyr残基上; 大部分的早期信号转导是由酪氨酸蛋白激酶介导
的(protein tyrosine kinase, PTKs); Ser/Thr磷酸化修饰多发生于信号转导级联反应
18
蛋白质磷酸酶
丝氨酸/苏氨酸蛋白磷酸酶 (PP1, PP2A); 酪氨酸蛋白磷酸酶 (PTP1B); 双特异性蛋白磷酸酶 (DUSP1); 脂质磷酸酶 (PTEN, SHIP)。
2019/9/4
19
Ser/Thr与Tyr蛋白磷酸酶
Ser/Thr磷酸酶 (PP1)
催化机制:
含有双核的金属离子中心; 通过水分子来进行亲核攻击; 磷酸基团不转移到磷酸酶上,
2019/9/4
13
双特异性蛋白激酶
MAPKK可同时催化MAPK激酶的T与Y的磷酸化 修饰:
磷酸化位点的三肽模体 — TXY ERK — TEY p38 — TGY JNK — TPY
MAPK是通过Thr和Tyr的双位点同时磷酸 化而被激活。
2019/9/4
14
磷酸肌醇循环
phosphatidylinositol
Fe
OO
HO
P O-Ser-substrate
2019/9/4
8
蛋白激酶的分类
酪氨酸蛋白激酶
受体激酶 (EGFR,FGFR,PDGFR) 非受体激酶 (JAK,Src,Abl)
丝氨酸/苏氨酸蛋白激酶 (PKC,MAPK,Plk,Rho Kinases)
双特异性蛋白激酶 (MAPKK)
蛋白质的转运与信号传导课件.ppt
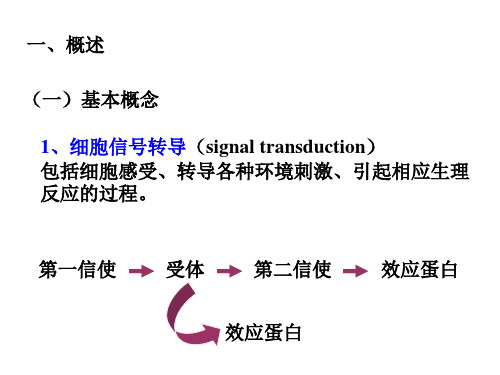
三、反应
同时,植物细胞信号转导系统在某些方面还保留了低等 原核细胞的信号转导机制,例如植物激素乙烯受体ETR1 与细菌双组份信号转导系统之间具有极大的相似性。
二、植物细胞信号转导过程
植物细胞的信号转导过程可以简单概括为: 刺激与感受——信号转导——反应三个重要的环节。
(一)刺激与感受 受体
(二)信号转导
此过程相当复杂,主要包括胞外信号的跨膜转换、细 胞内第二信使系统和信号的级联放大以及蛋白质的可 逆磷酸化。
1、信号跨膜转换
(1)通过离子通道连接受体跨膜转换信号
离子通道(ion channel)是存在于膜上可以跨膜转运 离子的一类蛋白质。
(2)酶促信号直接跨膜转换
该过程的跨膜信号转换主要由酶连受体来完成。
cell
胞外环境信号:指机械刺激、磁场、辐射、温度、风、 光、CO2、O2、土壤性质、重力、病原因子、水分、营养 元素、伤害等影响植物生长发育的重要外界环境因子。
胞间信号:指植物体自身合成的、能从产生之处运到别 处,并对其他细胞作为刺激信号的细胞间通讯分子,通 常包括植物激素、气体信号分子NO以及多肽、糖类、细 胞代谢物、甾体、细胞壁片段等。
3、第二信使(second messenger)
又称次级信使,是指细胞感受胞外环境信号和胞间信 号后产生的胞内信号分子,从而将细胞外信息转换为 细胞内信息。
一般公认的细胞内第二信使有钙离子(Ca2+)、肌 醇三磷酸(inositol 1,4,5-trisphosphate,IP3)、二酰 甘油(1,2-Diacylglycerol,DG)、环腺苷酸 (cAMP)、环鸟苷酸(cGMP)等。
•高特异性 •高的亲合力 •高饱和性 •可逆性 •通过磷酸化(与动物细胞信号传 导的区别)
信号转导教学课件ppt

信号转导与疾病
许多疾病的发生与信号转导异常有关,如肿瘤、自身免疫 性疾病、神经退行性疾病等。
针对信号转导途径的治疗策略已成为许多药物研发的重要 方向,包括抑制剂、激动剂、拮抗剂等。
02
信号转导基本元件
细胞膜受体
定义
01
细胞膜受体是指能够接受外界信号分子,并将其转化为细胞内
信号分子的跨膜蛋白。
种类
肿瘤细胞信号转导与血管生成和侵袭
1 2
肿瘤细胞血管生成与信号转导
详细阐述肿瘤细胞通过哪些信号转导通路促进 血管生成以及血管生成的信号转导机制。
肿瘤细胞侵袭与信号转导
介绍与肿瘤细胞侵袭相关的信号转导通路,如 基质金属蛋白酶(MMPs)及其抑制剂等。
3
肿瘤细胞的转移与信号转导
阐述肿瘤细胞转移过程中涉及的信号转导通路 及机制,包括转移前后的变化及转移过程中的 关键分子作用。
PLC通路的生物学功 能
PLC通路主要参与调节细胞增殖、分 化、凋亡等生物学过程,并在多种疾 病中发挥重要作用。
要点三
PLC通路与其他信号 通路的交叉互动
PLC通路与其他信号通路如MAPK、 PI3K等存在交叉互动,共同调节细胞 生理功能。
04
信号转导与免疫
固有免疫
01
固有免疫应答
是生物体在长期进化过程中形成的、对多种病原体具有防御作用的天
02
细胞膜受体可分为离子通道型受体、G蛋白偶联型受体和酶偶
联型受体等。
功能
03
细胞膜受体在感知外部环境刺激、调节细胞反应和通讯方面起
着重要作用。
G蛋白
定义
G蛋白是指与细胞膜受体结合并被激活的一类蛋 白,其在信号转导中起着关键作用。
第七章 细胞信号转导 PPT课件

细胞内 蛋白质 可逆磷 酸化
PKA
酶蛋白磷酸化修饰
细胞反应
Ca Ca + + CaM + Ca Ca E
+
CaM 对光敏素作用图解:
光
Pr
Pfr <10-6 M[Ca] 10-6 M CaM
CaM
E [CaM
E]
Ca Ca + + CaM + Ca Ca E
+
激活钙ATP 酶、钙通道 、NAD激酶 、多种蛋白 激酶等
孢子发 芽、有 丝分裂 、原生 质流动
植 物 体 内 的 信号传导
(signal transduction)
细胞外
质膜
G蛋白连 接受体 受体 二元组 分系统
细胞内
胞外 信号
Ca/CaM;IP3/DAG 蛋白可逆磷酸化
细胞反应
信号输入
跨膜信号转换
细胞内信号转 导网络
信号输出
第一节 信号与受体结合
• 一、信号
胞外信号
化学信号(chemical signal): 细胞感受刺激后合成并传递到作用 部位引起生理反应的化学物质(激素和 病原因子)。 物理信号(physical signal): 细胞感受到刺激后产生的能够传递 信息作用的光、电和水力等信号。 初级信号(第一信使)
小
G 蛋 白:一个亚基的单体
G蛋白的发现:吉尔曼(Gilman)、罗德贝尔(Rodbell) 获诺贝 尔医学生理奖(1994)
G蛋白有二种类型,一种为异源三聚体G蛋白;另 一种为小G蛋白(与a亚基相似)
• Three subunits
G蛋白 signalling
信号转导通路PPT课件

细胞内信号传递特点
信号的逐级放大
细胞内信号传递过程中,信号分子通过级联反应 逐级放大,使微弱的细胞外信号能够引起强烈的 细胞生理反应。
信号的可调性
细胞内信号传递过程受到多种因素的调节,包括 受体表达水平、信号分子的合成与降解、信号转 导蛋白的活性与定位等,这些调节机制使细胞能 够对外界刺激作出精确而灵活的应答。
免疫细胞信号转导通路的抑制失活
02 如免疫抑制性受体信号转导通路的失活,导致免疫细
胞过度激活和炎症反应。
免疫细胞与靶细胞之间的信号转导异常
03
免疫细胞与靶细胞之间的信号转导异常,导致免疫相
关疾病的发生和发展。
其他常见疾病中信号转导问题
心血管疾病中信号转导异常
如血管内皮细胞信号转导通路的异常,导致动脉粥样硬化和高血 压等疾病的发生。
信号的特异性
细胞内信号传递具有高度的特异性,不同的信号 分子只能激活特定的信号转导途径,引起特定的 细胞生理反应。
信号的整合性
细胞内存在多种信号转导途径,这些途径之间通 过交叉对话和相互调控,实现对细胞生理功能的 整体协调和控制。
02
典型信号转导通路介绍
G蛋白偶联受体介导通路
G蛋白偶联受体(GPCR)是一大类膜蛋白受体的统称 ,介导细胞对多种信号分子的响应。
GPCR与G蛋白结合后,通过激活或抑制下游效应器酶, 将信号传递至细胞内。
常见的GPCR介导的信号转导通路包括cAMP信号通路、 磷脂酰肌醇信号通路等。
酶联受体介导通路
01
酶联受体是一种具有内在酶 活性的受体,其介导的信号 转导通常与受体的酶活性相
关。
02
酶联受体通过催化特定的底 物生成第二信使,从而将信
导通路中的关键基因。
蛋白激酶和蛋白磷酸酶在信号转导中的作用

蛋白激酶:催化蛋白质磷酸化的酶类,反应中需有高能化合物(如ATP)参加。
将A TP的γ磷酸基转移到底物特定的氨基酸残基上,使蛋白质磷酸化的一类磷酸转移酶。
根据其底物蛋白被磷酸化的氨基酸残基种类,可将它们分为5类:蛋白丝氨酸/苏氨酸激酶、蛋白酪氨酸激酶、蛋白组氨酸激酶、蛋白色氨酸激酶和蛋白天冬氨酰基/谷氨酰基激酶。
蛋白磷酸酶:催化磷酸化氨基酸残基脱磷酸的酶。
与蛋白激酶一起配合调节底物蛋白质的磷酸化作用,调控多种细胞生物学过程。
根据底物蛋白质分子上磷酸化的氨基酸残基的种类主要分为蛋白质丝氨酸/苏氨酸磷酸酶、蛋白质酪氨酸磷酸酶和双特异性磷酸酶。
蛋白激酶可使蛋白质磷酸化,蛋白磷酸酶使蛋白去磷酸化。
蛋白磷酸化与去磷酸化是真核细胞信号转导的共同通路,其动态变化几乎涉及从胚胎发育到个体成熟的所有过程,包括细胞的癌变和凋亡。
磷酸化与去磷酸化的平衡主要由蛋白激酶(protein kinases,PK)和磷酸酶(protein phosphatases, PPs)调控。
磷酸化和去磷酸化作为分子开关,是信号转导中最简便而又十分快捷的反应方式,一般是通过磷酸化而激活,去磷酸化而失活。
大量研究结果表明蛋白质的磷酸化与去磷酸化过程在多种信号识别与转导中起重要作用,它是生物体中普遍存在的一种调节过程。
蛋白激酶是一类将ATP γ位的磷酸基团转移到底物的氨基酸残基上引起靶蛋白发生磷酸化的调节酶,它通过促进功能蛋白的磷酸化而使细胞对各种刺激做出相应的反应。
泛素化途径的功能:由于基因突变、自由基破坏、环境胁迫、疾病等导致反常蛋白的产生,需要被及时降解清除,以免干扰正常的生命活动;维持体内的氨基酸代谢库;防御机制的组成部分;蛋白质前体的裂解加工等。
蛋白酪氨酸激酶转导细胞增殖与分化信号ppt课件

激活G蛋白
激活蛋白酪氨酸激酶
去极化与超极化调节蛋白质功 调节蛋白质的功能和
能和表达水平
表达水平,调节细胞
分化和增殖
Biochemistry
一、 细胞内受体多属于转录因子
位于细胞内的受体多为转录因子,与相应配体结 合后,能与DNA的顺式作用元件结合,在转录 程度调理基因表达。
该型受体结合的信息物质有类固醇激素、甲状腺 素、维甲酸、维生素D等,它们进入细胞后,有 些可与其位于细胞核内的受体相结合构成激素受体复合物,有些那么先与其在细胞质内的受体 相结合,然后以激素-受体复合物的方式穿过核 孔进入核内。
Biochemistry
❖ G蛋白经过G蛋白偶联受体〔G proteincoupled receptors,GPCRs〕与各种下游 效应分子,如离子通道、腺苷酸环化酶、 PLC联络,调理各种细胞功能。
Biochemistry
2.低分子质量G蛋白
❖ 低分子量G蛋白〔21kD〕,在多种细胞信号 转导途径中具有开关作用。
Biochemistry
三种膜受体的特点
特性
内源性 配体 结构
跨膜区 段数目 功能 细胞 应答
离子通 道受体 神经递质
寡聚体形 成的孔道
4个
离子通道 去极化与 超极化
G-蛋白偶联受体
单次跨膜受体
神经递质、激素、趋化因子、 外源刺激(味,光)
生长因子 细胞因子
单体
具有或不具有催化活 性的单体
7个
1个
5’ AGGTCATGACCT 3’ 3’ TCCAGTACTGGA 5’
Biochemistry
二、离子通道型膜受体是化学信号与 电信号转换器
❖ 离子通道型受体是一类本身为离子通道的受 体,它们的开放或封锁直接受化学配体的控 制,被称为配体-门控受体通道〔ligandgated receptor channel〕。
《细胞信号转导》PPT课件

molecularbiology生物化学与分子生物学教研室第一节细胞通讯第二节细胞信号转导的分子机制第三节不同受体介导的细胞信号转导通路第四节细胞信号转导与医学细胞外信号细胞内的多种分子的浓度活性位置变化蛋白激酶与蛋白磷酸酶proteinkinaseproteinphosphatasegtp结合蛋白gtpbindingproteinmolecularswitchsgtpgtpgdpgtpgtpgtpg蛋白的主要类型肾上腺素腺苷酸环化酶atpcamp无活性pka活化pka磷酸化酶b激酶糖原合酶糖原分解增加肾上腺素腺苷酸环化酶atpcampg蛋白一类和gtp或gdp结合位于胞膜胞浆面的外周蛋白具有信号转导功能由三个亚基组成非活化形式活化形式proteinactivationpkacampacplcippkacampac11gtp结合蛋白异源三聚体低分子量g蛋白gtp结合形式为活性形式gdp结合形式为非活性形式2130kda称为ras超家族现有50多种具有gtp酶活性13gapgtpaseactivatingproteingtpase激活蛋白sosguanidineexchangefactor鸟苷酸交换因子gefgtpoffgdpgaprasrassosgap第二节细胞信号转导的分子机制15蛋白复合物proteincomplexesclusters是细胞信号转导分子共同构成的基本工作场所是信号转导过程特异性和精确性的保证是网络性调控的基础signalosomestransducisomessignalcomplexsignalcassettessignalingmodules16转录调控复合物17蛋白相互作用是信号转导复合物形成的基础蛋白相互识别的结构基础蛋白复合物的重要结构蛋白衔接蛋白adapterprotein支架蛋白scaffoldprotein1840proteininteractiondomain19sh2domainsrcsh2srchomologydomainpyeei20sh3domainclassrkxxpxxpclasspxxpxrsrchomologydomain蛋白激酶btkphthsh3sh2催化区衔接蛋白grb2sh3sh2sh3转录因子statdna结合区sh2ta细胞骨架蛋白tensinsh2ptb22phosphotyrosine?sh2?ptbapoptosis?dd?ded?car
《蛋白激酶与磷酸酶》课件

要点二
作用机制
这些磷酸酶通过不同的机制将磷酸基团从蛋白质或分子上 转移给水或其他分子,实现对蛋白质功能的调节,参与各 种生物学过程。
01
蛋白激酶与磷酸酶 在生物学中转导中发挥关键作用,通 过调节蛋白质磷酸化状态,影响细胞内信号通路的传递 。
详细描述
半胱氨酸蛋白激酶是一类具有催化半胱 氨酸残基磷酸化的酶,主要参与细胞凋 亡和自噬等过程。
VS
详细描述
半胱氨酸蛋白激酶在细胞凋亡和自噬等过 程中发挥重要作用,通过磷酸化特定的半 胱氨酸残基,调控相关蛋白的活性,触发 细胞死亡或自噬等过程,从而维持细胞内 环境的稳定。
其他蛋白激酶
总结词
除了上述几种常见的蛋白激酶外,还有许多其他类型的蛋白激酶,如钙调蛋白激酶、环氧合酶等,它 们在细胞内不同生物学过程中发挥重要作用。
酪氨酸蛋白激酶
总结词
酪氨酸蛋白激酶主要参与生长因子受体信号转导和免疫信号转导,通过磷酸化酪 氨酸残基来激活或抑制下游信号分子。
详细描述
酪氨酸蛋白激酶在细胞表面受体,如生长因子受体和免疫受体中发挥作用,当这 些受体被配体激活时,酪氨酸蛋白激酶会磷酸化特定的酪氨酸残基,触发一系列 信号转导事件,最终影响细胞生长、增殖和分化等过程。
,通过抑制其活性或调节其表达,可有效抑制肿瘤的生长和扩散。
01
蛋白激酶与磷酸酶 的研究前景与展望
新药研发与治疗策略
针对特定蛋白激酶或磷酸酶的抑制剂或激活剂的研发,用于治疗癌症、神经退行性 疾病、糖尿病等疾病。
利用蛋白激酶或磷酸酶的调节作用,开发新的药物作用靶点,提高药物的疗效和特 异性。
深入研究蛋白激酶或磷酸酶在疾病发生发展中的作用机制,为新药研发提供理论支 持和实践指导。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA依赖性蛋白激酶(DNA-dependent protein kinaes,DNA-PK)可磷酸 化许多核蛋白,包括核受体、转录因子、 DNA拓扑异构酶和RNA聚合酶Ⅱ等。
• 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
(二)蛋白激酶的种类
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
蛋白激酶(protein kinase,PK):
是一类磷酸转移酶,其作用是将 ATP 的 - 磷酸基转移到底物特定的氨基酸残基 上,使蛋白质磷酸化, 发挥其生理生化功 能。
(一)蛋白激酶的结构 共同的结构特征:
• 保守的催化结构域/亚基 • 调节结构域/亚基 • 其他功能结构域
PKC
1. 催化结构域/亚基 催化核心含有12个高度保守的亚区。 功能:
CH1 CH2
K inase dom ain
N -te r LIM S
M igfilin
FERM PH FERM
M ig-2
p a x illin
In te g rin s
ECM
G enerously provided to the C M C by C ary W u (Apr-03
(9)DNA依赖性蛋白激酶
(7)核糖体S6激酶(S6K)
包括S6KⅠ和S6KⅡ,能催化核糖体 S6蛋白磷酸化。
(8)整合素连接激酶
整合素连接激酶(intergrin-linked kinase,ILK)可直接磷酸化PKB/Akt, 其活性依赖PI3K。
PIN C H , ILK dom ain structures & interacting partners
C1是膜结合区,与佛波酯、DAG和PS结合 有关。
C2是Ca2+结合位点。
• 催化区:
C3是ATP结合位点。
C4是底物蛋白结合位点。
PS DAG
Ca2+
ATP
PKC底物:
• 参与信号转导的底物,如表皮生长因子 受体、胰岛素受体、T细胞受体(TCR)、 Ras、GTP酶活化蛋白等;
• 参与代谢调控的底物,如膜上的通道和 泵;
活化需要Ca2+ 、DAG和PS。
B组-新型PKC (new PKC,nPKC):、、 (L) 、、 和 。 活化不需要Ca2+。
C组-非典型PKC (atypical PKC,aPKC): 和 / 。 活化不需要Ca2+和DAG。
各亚型均由一条肽链组成,分为四个保 守区C1~C4和五个可变区V1~V5。 • 调节区:
N c k -2
SH3 SH3 SH3
SH2
PIN C H
LIM 5 LIM 4 LIM 3 LIM 2
LIM 1
IL KΒιβλιοθήκη ppANK ANK ANK ANK PH
RTK
C H -ILK B P/actopaxin p a rv in s /a ffix in
N -te r
Actin C ytoskeleton Filam in
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HCCH2 OH OC
NH
O
HCCH2 OPO
OC
O
(1)蛋白激酶A(protein kinase A,PKA)
即cAMP依赖性蛋白激酶。
• 调节基因表达的底物,如转录因子、翻 译因子、S6K、Raf激酶等。
PKC分布:
• 广泛分布于各组织的胞质,以Ca2+依赖 的形式从胞质中移位到细胞膜上,此过程 称之为转位。PKC转位是其活化的标志。
• 佛波酯(TPA or PMA)是一种促癌剂,其 结构与DAG相似,可持续活化PKC,促进 细胞增殖。
第五章
蛋白激酶、蛋白磷酸酶 与信号转导
【目的要求】
1.掌握蛋白激酶的概念及其催化的蛋白质磷酸 化反应;丝氨酸/苏氨酸蛋白激酶的主要种类及 其激活剂;PKC的分类及其特点;酪氨酸蛋白 激酶的概念、分类及其重要的结构域;受体酪 氨酸蛋白激酶的概念;蛋白磷酸酶的概念及分 类。
【目的要求】
2.熟悉受体酪氨酸蛋白激酶的分类;双重特异 性蛋白激酶;蛋白激酶在信号转导中的作用; 丝氨酸/苏氨酸蛋白磷酸酶的生物学作用;酪氨 酸蛋白磷酸酶的分类及其在信号转导中的作用; 双重特异性蛋白磷酸酶。
乙酸豆蔻佛波
(3)钙/钙调素依赖性蛋白激酶(CaMK)
包括肌球蛋白轻链激酶(myosin light chain kinase,MLCK)、磷酸化酶 激酶、CaMKⅡ等。
(4)CMGC组蛋白激酶
• 脯氨酸依赖性激酶(proline depedent kinase,PDK)
• 酪蛋白激酶Ⅱ(casein kinase Ⅱ ,CK Ⅱ )家族
PDK:
细胞周期素依赖性蛋白激酶(cyclin depedent kinase,CDK)家族
丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)家族
糖原合成酶激酶3(glycogen synthetase kinase 3,GSK3)
CDK样激酶(CDK-like kinase, CLK)家族
全酶存在胞浆,被cAMP激活后,催化亚 基可① 调节代谢;②调节离子通道;③调 节其他信号转导途径的蛋白;④ 进入细胞 核调节基因表达。
(2)蛋白激酶C
即Ca2+和磷脂依赖的蛋白激酶,受 Ca2+ 、DAG和PS激活。
PKC有11种亚型:
A组-典型PKC (classical PKC,cPKC): 、Ⅰ、Ⅱ和
MAPK途径:
EGF途径
(5)蛋白激酶G
即cGMP依赖性蛋白激酶( cGMP – dependent protein kinase,PKG),以 cGMP 为变构剂,在脑和平滑肌中含量较 丰富。
(6)G蛋白偶联受体激酶
有肾上腺素受体蛋白激酶( adrenergic receptor kinase, -ARK)、 -ARK相关激酶和视紫红质激酶等。
• 与蛋白质或多肽底物结合; • 与磷酸供体ATP/GTP结合; • 转移磷酸基到底物相应的氨基酸残基上。
2. 调节结构域/亚基 同源性较低。 作用: 调节酶的活性; 靶向作用,与酶的亚细胞定位有关。
(二)蛋白激酶的种类
真核细胞的蛋白激酶可分为五类: ① 丝氨酸/苏氨酸蛋白激酶 ② 酪氨酸蛋白激酶 ③ 组/赖/精氨酸蛋白激酶 ④ 半胱氨酸蛋白激酶 ⑤ 天冬氨酸/谷氨酸蛋白激酶
蛋白激酶(protein kinase,PK):
是一类磷酸转移酶,其作用是将 ATP 的 - 磷酸基转移到底物特定的氨基酸残基 上,使蛋白质磷酸化, 发挥其生理生化功 能。
(一)蛋白激酶的结构 共同的结构特征:
• 保守的催化结构域/亚基 • 调节结构域/亚基 • 其他功能结构域
PKC
1. 催化结构域/亚基 催化核心含有12个高度保守的亚区。 功能:
CH1 CH2
K inase dom ain
N -te r LIM S
M igfilin
FERM PH FERM
M ig-2
p a x illin
In te g rin s
ECM
G enerously provided to the C M C by C ary W u (Apr-03
(9)DNA依赖性蛋白激酶
(7)核糖体S6激酶(S6K)
包括S6KⅠ和S6KⅡ,能催化核糖体 S6蛋白磷酸化。
(8)整合素连接激酶
整合素连接激酶(intergrin-linked kinase,ILK)可直接磷酸化PKB/Akt, 其活性依赖PI3K。
PIN C H , ILK dom ain structures & interacting partners
C1是膜结合区,与佛波酯、DAG和PS结合 有关。
C2是Ca2+结合位点。
• 催化区:
C3是ATP结合位点。
C4是底物蛋白结合位点。
PS DAG
Ca2+
ATP
PKC底物:
• 参与信号转导的底物,如表皮生长因子 受体、胰岛素受体、T细胞受体(TCR)、 Ras、GTP酶活化蛋白等;
• 参与代谢调控的底物,如膜上的通道和 泵;
活化需要Ca2+ 、DAG和PS。
B组-新型PKC (new PKC,nPKC):、、 (L) 、、 和 。 活化不需要Ca2+。
C组-非典型PKC (atypical PKC,aPKC): 和 / 。 活化不需要Ca2+和DAG。
各亚型均由一条肽链组成,分为四个保 守区C1~C4和五个可变区V1~V5。 • 调节区:
N c k -2
SH3 SH3 SH3
SH2
PIN C H
LIM 5 LIM 4 LIM 3 LIM 2
LIM 1
IL KΒιβλιοθήκη ppANK ANK ANK ANK PH
RTK
C H -ILK B P/actopaxin p a rv in s /a ffix in
N -te r
Actin C ytoskeleton Filam in
1. 丝氨酸/苏氨酸蛋白激酶
丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase,S/T-PK)催化 丝氨酸/苏氨酸的羟基磷酸化。
NH HCCH2 OH OC
NH
O
HCCH2 OPO
OC
O
(1)蛋白激酶A(protein kinase A,PKA)
即cAMP依赖性蛋白激酶。
• 调节基因表达的底物,如转录因子、翻 译因子、S6K、Raf激酶等。
PKC分布:
• 广泛分布于各组织的胞质,以Ca2+依赖 的形式从胞质中移位到细胞膜上,此过程 称之为转位。PKC转位是其活化的标志。
• 佛波酯(TPA or PMA)是一种促癌剂,其 结构与DAG相似,可持续活化PKC,促进 细胞增殖。
第五章
蛋白激酶、蛋白磷酸酶 与信号转导
【目的要求】
1.掌握蛋白激酶的概念及其催化的蛋白质磷酸 化反应;丝氨酸/苏氨酸蛋白激酶的主要种类及 其激活剂;PKC的分类及其特点;酪氨酸蛋白 激酶的概念、分类及其重要的结构域;受体酪 氨酸蛋白激酶的概念;蛋白磷酸酶的概念及分 类。
【目的要求】
2.熟悉受体酪氨酸蛋白激酶的分类;双重特异 性蛋白激酶;蛋白激酶在信号转导中的作用; 丝氨酸/苏氨酸蛋白磷酸酶的生物学作用;酪氨 酸蛋白磷酸酶的分类及其在信号转导中的作用; 双重特异性蛋白磷酸酶。
乙酸豆蔻佛波
(3)钙/钙调素依赖性蛋白激酶(CaMK)
包括肌球蛋白轻链激酶(myosin light chain kinase,MLCK)、磷酸化酶 激酶、CaMKⅡ等。
(4)CMGC组蛋白激酶
• 脯氨酸依赖性激酶(proline depedent kinase,PDK)
• 酪蛋白激酶Ⅱ(casein kinase Ⅱ ,CK Ⅱ )家族
PDK:
细胞周期素依赖性蛋白激酶(cyclin depedent kinase,CDK)家族
丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)家族
糖原合成酶激酶3(glycogen synthetase kinase 3,GSK3)
CDK样激酶(CDK-like kinase, CLK)家族
全酶存在胞浆,被cAMP激活后,催化亚 基可① 调节代谢;②调节离子通道;③调 节其他信号转导途径的蛋白;④ 进入细胞 核调节基因表达。
(2)蛋白激酶C
即Ca2+和磷脂依赖的蛋白激酶,受 Ca2+ 、DAG和PS激活。
PKC有11种亚型:
A组-典型PKC (classical PKC,cPKC): 、Ⅰ、Ⅱ和
MAPK途径:
EGF途径
(5)蛋白激酶G
即cGMP依赖性蛋白激酶( cGMP – dependent protein kinase,PKG),以 cGMP 为变构剂,在脑和平滑肌中含量较 丰富。
(6)G蛋白偶联受体激酶
有肾上腺素受体蛋白激酶( adrenergic receptor kinase, -ARK)、 -ARK相关激酶和视紫红质激酶等。