第八章mRNA剪接编辑
mRNA剪切机制简介mRNA专题

mRNA剪切机制简介mRNA专题细胞核内前体mRNA的剪接的执行者是剪接小体,它能够在成熟mRNA出核和翻译之前识别剪接信号,移除不编码内含子,并将能够编码蛋白的外显子拼接在一起。
细胞核内前体mRNA的剪接需要经历2次转酯化反应化步骤去掉内含子才能将相邻的外显子拼接成成熟的mRNA。
在前体mRNA上有三个反应区域分别在5'剪接位点(5'SS),3'剪接位点(3'SS)以及分枝位点(图1)。
除了这三个反应区域外,在多细胞生物体的内含子上还拥有保守性的多聚嘧啶束,它位于3'剪接位点以及分枝位点之间。
图1选择性剪接发生过程示意图选择性剪接由内含子5'剪接位G和U2个核苷酸点以及3'剪接位点A和G 2个核苷酸介导。
分支点A核苷酸非常保守的,一般位于3'剪接位点上游20-50个核苷酸。
剪接反应的过程发生2次转酶化反应,这个过程中需要5个snRNPs复合物(Ul,U2,U4,U5,andU6)。
这些复合物能够聚集在前体mRNA上形成大分子的聚合物。
剪接小体,核内最大的RNP复合物,能够识别这些反应位点并催化前体mRNA发生剪接。
剪接小体中主要的模块是snRNPs复合物。
剪接小体一般包含5种snRNPs:U1、U2、U4,U5和U6 snRNP。
每一个snRNP包含了单个snRNA和至少7种蛋白亚基。
这些snRNP和另外的非snRNP相关蛋白(例如SF1、U2AF和Prp19复合物)一步步有序的聚集在前体mRNA上依次形成前剪接小体E,A,B以及C复合物(图1)。
在这有序的过程中,这些snRNP以及非snRNP相关蛋白和反应位点之间发生复杂的结合与去结合过程,这些复杂的过程为核小体提供了多次检查的机会以保证它们结合的准确性从而提高位点选择的精确性。
在核小体组装之前,U1 snRNP占据5'剪接位点,而SF1结合在分枝位点,这2个过程是ATP依赖的并最终形成前剪接小体E复合物(图1)。
现代分子生物学第八章

以后,照样可以产生有活性的mRN A; 另一些基因, 如SV40 T抗原基因,一旦除去内含子,成熟mRNA运 人细胞质基质的过程就完全被阻断。
10
8.1.3基因家族 • 真核细胞的DNA是单顺反子结构,很 • 少出现置于一个启动子控制之下的操纵子。 真核细胞中许多相关的基因常按功能成套 组合,被称为基因家族。 • 同一家族的成员有时紧密地排列在一起, 成为一个基因簇;但更多的时候,它们分散 在同一染色体的不同部位.甚至位于不同的 染色体上。具有各自不同的表达调控模式。
8
• 选择性剪接:同一基因的转录产物由于不同的剪接方式形成不同 mRNA的过程。 • 选择性剪接的典刑例子:小鼠淀粉酶基因表达的组织特异性变化
9
• 在小鼠肝和唾液腺这两种组织中,淀粉酶mRNA的编码 序列完全相同,但5‘端起始部分长度不同。 • 事实上,L外显子只是唾液腺淀粉酶基因中内含子序列的 一部分,将在mRNA成熟过程中被切除。 • 有实验证明,由S外显子起始的转录产物是由L外显子起 始转录产物的100倍以上。 • 由于一个基因的内含子成为另一个基因的外显子,形成 基因的差异表达,这是真核基因断裂结构的一个重要特 点。
现代分子生物学
指导老师:杨林松 演绎人:徐楸能
1
8.1真核基因表达调控相关概念和一 般规律
•
• 8.1.1基因表达的基本概念 基因组: 一个细胞或病毒所携带的全部遗传 信息或整套基因。 基因:指能产生一条肽链或功能RNA所必需的 DNA片段。它包括编码区和其上下游区域。以 及在编码片段间(外显子)的间断切割序列(内 含子) 基因表达:基因经过转录、翻译,产生具有特 异生物学功能的蛋自质分子或RNA分子的过程。 基因表达调控:受内源及外源信号调控的这个 调控的过程。
第8章 RNA转录后的加工
4-硫尿苷
次黄嘌呤核苷(肌苷)
1-甲基鸟苷
N6 -异戊烯基腺苷
假尿嘧啶核苷
二氢尿苷
真核tRNA内含子的特点:
• 位置相同,都在反密码 子环的下游,内含子和 反密码子配对形成茎环 • 外显子和内含子交界处 无保守序列 • 不同tRNA的内含子长度 和序列各异 • 内含子的剪切是依靠 RNA酶异体催化(自身 不是核酶)
mRNA
蛋白质合成模板
RNA的加工 rRNA和tRNA:不论原核或真核生物的rRNA和tRNA都是以初级 转录本形式被合成的,然后再加工成为成熟的RNA分子。
mRNA: 原核生物的mRNA却不需加工,仍为初级转录本的形式。
真核生物pre-mRNA要经过复杂的加工历程,包括加帽、 加尾和内含子的剪接等。
1、在5’端加帽(cap)) 场所是核内
帽子0:m7 G ppp X 单细胞生物(如酵母) 帽子1:m7 G ppp Xm 多细胞生物,主要形式 帽子2:m7 G ppp XmpYm 占10-15%
三种帽子的 共同位置 在帽子1中 可被甲基化
帽子1
m7Gppp
鸟甘酸转移酶
帽子2
剪接前加帽,剪接后加帽 剪接前加帽
类似的加工过程也可以在某些噬菌体的多顺反子mRNA中见到。例 如,大肠杆菌噬菌体T7的早期基因转录出一条长的多顺反子mRNA, 经RNaseIII切割成5个单独的mRNA和一段5′端前导序列。mRNA的 切割对其中某些早期蛋白质的合成是必要的。推测可能是由于较 长的 mRNA产生二级结构,会阻止有关编码序列的翻译。这种RNA 二级结构(可能还有三级结构)与其功能的调控关系在多种情况 下均可看到,并不仅限于翻译起始的调控。通过 RNA 链的裂解, 改变了RNA的二级结构,从而影响它的功能。
RNA的生物合成.pptx

谢谢聆听
2目0 录
转录起始过程
1. RNA聚合酶全酶(2)与模板结合
2. DNA双链解开3. 在NA聚合酶作用下发生第一次聚合反应, 形成转录起始复合物
5-pppG -OH + NTP 5-pppGpN - OH 3 + ppi
转录起始复合物:
RNApol (2) - DNA - pppGpN- OH 3
RNA
UUUU...…
UUUU...…
茎环(stem-loop)/发夹(hairpin)结构
2019-11-11
谢谢聆听
2目9 录
RNA-pol
5
3
3
5
5´pppG
茎环结构使转录终止的机理
• 使RNA聚合酶变构,转录停顿;
• 使转录复合物趋于解离,RNA产物释放。
2019-11-11
谢谢聆听
谢谢聆听
3目7 录
(二)转录延长
真核生物转录延长过程与原核生物大致相 似,但因有核膜相隔,没有转录与翻译同步的 现象。
RNA-pol前移处处都遇上核小体。 转录延长过程中可以观察到核小体移位和 解聚现象。
2019-11-11
谢谢聆听
3目8 录
转
核小体
录
延
RNA-Pol
长 转录方向
中
的
核
小
RNA-Pol
2019-11-11
谢谢聆听
目8 录
结构基因
5
编码链
3
模板链
转录方向
转录方向
模板链
3
编码链
5
2019-11-11
谢谢聆听
目9 录
不对称转录(asymmetric transcription)
抗肿瘤mRNA剪接调控技术联合纳米载体递送系统的作用机制及其在临床治疗中的应用

抗肿瘤mRNA剪接调控技术联合纳米载体递送系统的作用机制及其在临床治疗中的应用摘要:本文探讨了抗肿瘤mRNA剪接调控技术与纳米载体递送系统的结合作用机制,并分析了其在临床治疗中的潜在应用。
通过对mRNA剪接机制的深入研究和对纳米载体递送系统的优化设计,本文提出了一种创新的联合策略,旨在提高抗肿瘤药物的靶向性和疗效。
通过实验验证,该策略在体外细胞实验和体内动物模型中均显示出显著的抗肿瘤效果。
本文还讨论了该策略可能面临的挑战和未来的发展方向。
关键词:mRNA剪接;纳米载体;抗肿瘤;联合策略;靶向治疗Abstract: This article explores the combined mechanism of action between antitumor mRNA splicing regulation technology and nanocarrier delivery systems, and analyzes its potential applications in clinical treatment. Through indepth research on mRNA splicing mechanisms and optimized design of nanocarrier delivery systems, this article proposes an innovative joint strategy aimed at improving the targeting and efficacy of antitumor drugs. Through experimental verification, this strategy has shown significant antitumor effects in both in vitro cell experiments and in vivo animal models. In addition, this article also discusses the potential challenges and future development directions of this strategy.Keywords: mRNA splicing; Nanocarriers; Antitumor; Joint strategy; Targeted therapy 第一章、引言1.1 研究背景与意义1.1.1 mRNA剪接调控技术的重要性mRNA剪接是真核生物基因表达过程中的一个关键步骤,它决定了一个基因可以产生多种不同的蛋白质。
前体mRNA的剪接过程
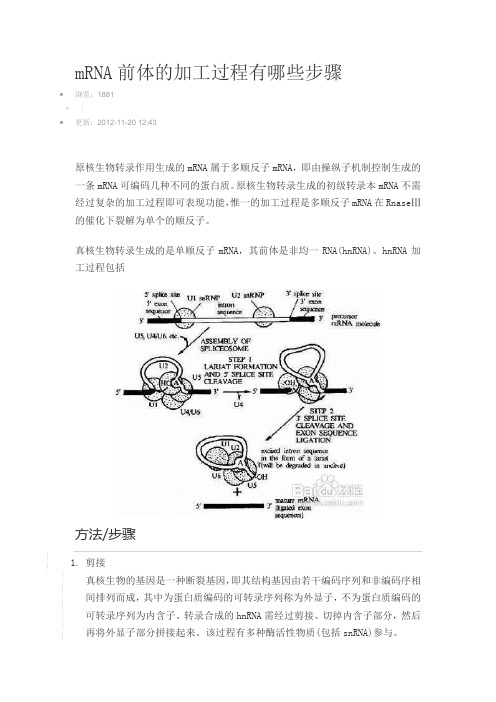
mRNA前体的加工过程有哪些步骤•浏览:1881•|•更新:2012-11-20 12:43原核生物转录作用生成的mRNA属于多顺反子mRNA,即由操纵子机制控制生成的一条mRNA可编码几种不同的蛋白质。
原核生物转录生成的初级转录本mRNA不需经过复杂的加工过程即可表现功能,惟一的加工过程是多顺反子mRNA在RnaseⅢ的催化下裂解为单个的顺反子。
真核生物转录生成的是单顺反子mRNA,其前体是非均一RNA(hnRNA)。
hnRNA加工过程包括方法/步骤1.剪接真核生物的基因是一种断裂基因,即其结构基因由若干编码序列和非编码序相间排列而成,其中为蛋白质编码的可转录序列称为外显子,不为蛋白质编码的可转录序列为内含子。
转录合成的hnRNA需经过剪接、切掉内含子部分,然后再将外显子部分拼接起来。
该过程有多种酶活性物质(包括snRNA)参与。
2.5′末端加“帽”真核细胞成熟mRNA的5′末端均有一个特殊的结构,即m7Gpp-pmnNp,称为“帽”。
帽的生成是在细胞核内进行的,但胞浆中也有酶体系,动物病毒mRNA 加帽过程就是在宿主细胞的胞浆内进行的。
3.3′末端加“尾”mRNA前体分子的3′末端有一段保守序列,由特异的核酸内切酶切去多余的核苷酸,然后在多聚A聚合酶的催化下,由ATP聚合生成多聚A尾。
该反应在核内发生,在胞浆中也可继续进行。
4.碱基修饰mRNA分子中有少量稀有碱基(如甲基化碱基)是在转录后经化学修饰(如甲基化)而形成的。
5.选择性加工某些MRNA前体含有多个3‘剪切位点和多聚腺苷酸化位点,因此利用这些选择性位点可产生具有不同3'端非编码区或者具有不同编码能力的RNA产物。
通过可变剪接途径可以挑先最保留在MRNA中的外显子,结果单个基因可以合成多种不同的蛋白质。
6.RNA编辑在合成并经RNA编辑加工之后,MRNA分子的序列可以发生改变。
个别核苷酸可以被置换,添加或者删除。
编辑过的MRNA翻译产生了较短脱脂基蛋白B48,由于基缺少一个结合受体的蛋白结构域,因此功能受限。
前体mRNA的剪接过程
mRNA前体的加工过程有哪些步骤•浏览:1881•|•更新:2012-11-20 12:43原核生物转录作用生成的mRNA属于多顺反子mRNA,即由操纵子机制控制生成的一条mRNA可编码几种不同的蛋白质。
原核生物转录生成的初级转录本mRNA不需经过复杂的加工过程即可表现功能,惟一的加工过程是多顺反子mRNA在RnaseⅢ的催化下裂解为单个的顺反子。
真核生物转录生成的是单顺反子mRNA,其前体是非均一RNA(hnRNA)。
hnRNA加工过程包括方法/步骤1.剪接真核生物的基因是一种断裂基因,即其结构基因由若干编码序列和非编码序相间排列而成,其中为蛋白质编码的可转录序列称为外显子,不为蛋白质编码的可转录序列为内含子。
转录合成的hnRNA需经过剪接、切掉内含子部分,然后再将外显子部分拼接起来。
该过程有多种酶活性物质(包括snRNA)参与。
2.5′末端加“帽”真核细胞成熟mRNA的5′末端均有一个特殊的结构,即m7Gpp-pmnNp,称为“帽”。
帽的生成是在细胞核内进行的,但胞浆中也有酶体系,动物病毒mRNA 加帽过程就是在宿主细胞的胞浆内进行的。
3.3′末端加“尾”mRNA前体分子的3′末端有一段保守序列,由特异的核酸内切酶切去多余的核苷酸,然后在多聚A聚合酶的催化下,由ATP聚合生成多聚A尾。
该反应在核内发生,在胞浆中也可继续进行。
4.碱基修饰mRNA分子中有少量稀有碱基(如甲基化碱基)是在转录后经化学修饰(如甲基化)而形成的。
5.选择性加工某些MRNA前体含有多个3‘剪切位点和多聚腺苷酸化位点,因此利用这些选择性位点可产生具有不同3'端非编码区或者具有不同编码能力的RNA产物。
通过可变剪接途径可以挑先最保留在MRNA中的外显子,结果单个基因可以合成多种不同的蛋白质。
6.RNA编辑在合成并经RNA编辑加工之后,MRNA分子的序列可以发生改变。
个别核苷酸可以被置换,添加或者删除。
编辑过的MRNA翻译产生了较短脱脂基蛋白B48,由于基缺少一个结合受体的蛋白结构域,因此功能受限。
真核生物mRNA选择性剪接发生的相关因素

真核生物mRNA选择性剪接发生的相关因素摘要:在真核生物细胞中,选择性剪接扮演着非常重要的角色,如蛋白质多样性、细胞代谢、疾病的发生等。
据报道可知,选择性剪接的发生与很多因素相关如剪接因子、剪接相关的保守序列、染色质结构等,其具体机制一直随着科学工作者的不断努力而完善。
由于pre-mRNA选择性剪接在真核生物中具有的独特作用,使得选择性剪接发生的相关因素也成为人们研究的热点课题。
本文就真核生物mRNA选择性剪接发生的相关因素做简要介绍。
关键词:选择性剪接;mRNA;真核生物;剪接相关Motif;染色质结构0前言1978年Gilbert[1]首次提出选择性剪接这一概念后,选择性剪接开始广泛成为人们研究的目标。
人类基因组计划公布的初步研究结果中人类基因数是3.5万个左右,而不是原先预计的8万至10万个。
由此可知一个基因不是只编码一个蛋白质,而是可能编码一个或多个蛋白质。
近期报道显示,在人类含多个外显子的pre-mRNA中,能发生选择性剪接的约占92-94%[2]。
同时选择性剪接与疾病的发生之间有着密切联系,如炎症路径和癌症发生等[25][26]。
这使得选择性剪接成为近年来研究的热点课题。
1选择性剪接的概念与模式pre-mRNA的选择性剪接(Alternative splicing,AS),即一个基因能产生多种mRNA,这极大地丰富了蛋白质组的多样性和基因表达调控的灵活性[7]。
近年来,在不同的真核生物物种中都有选择性剪接事件被报道,如酵母和人类等[3][4]。
选择性剪接发生模式主要包括以下5种[5]:可变的5’剪接位点(alternative 5’splice site):与内含子保守5’剪接位点相竞争,且能与该内含子的3’剪接位点相互作用发生选择性剪接的5’剪接位点;可变的3’剪接位点(alternative 3’splice site):与内含子保守3’剪接位点相竞争,且能与该内含子的5’剪接位点相互作用发生选择性剪接的3’剪接位点;选择性保留某些内含子(intron retention):在剪接过程中选择性保留整个内含子或部分内含子序列,使其成为成熟mRNA的一部分;外显子跳跃(exon skipping)和互斥外显子(mutually exclusive exons):都是表现在剪接过程中外显子被选择性拼接。
第八章 RNA加工(RNA processing)

A snoRNA (小核 仁RNA) base pairs with a region of rRNA that is to be methylated — define methylation sites
Pseudouridine (假尿嘧啶) modification
5
OH
五、RNA的自剪接
Each operon contains one copy of each of the 16S, the 5S and the 23S rRNA sequences. About 1~4 coding sequences for tRNA molecules are also present.
(二)断裂基因的证据
R-looping
The intron regions of the DNA will not find counterparts in the mRNA and so will form unhybridized loops. We call them R loops because hybridization with RNA caused them to form.
3. RNA cleavage
RNase III
Other types of RNase
M5, M16, M23
四、真核生物rRNA的加工
(一)真核生物rRNA基因
The rRNA genes are present in a tandemly repeated cluster (串联重复) containing 100 or more copies of the transcription unit, and are transcribed in nucleolus by RNA Pol I.
第八章mRNA剪接编辑

mRNA的多聚腺苷酸化过程
CPSF因子专一性与加尾信号AAUAAA结合 CstF因子与富GU区结合 CFI、CFII在加尾信号下游的15~25 nt处切割 poly(A)多聚酶催化多聚腺苷酸化,先添加约10
个腺苷酸,速度较慢 poly(A)结合蛋白的结合加快多聚酶延伸速度,
控制多聚腺嘌呤尾巴的长度
CPSF PAP
PAPB
Pol II的CTD促进切割
3 端多聚腺苷酸尾巴的功能
提高mRNA的稳定性 在核-质转运中起作用 提高mRNA的翻译效率 影响最后一个内含子的切除 创造终止密码子UGA 选择性加尾调节基因的表达
3. RNA剪接(Splicing)
真核生物mRNA前体的剪接
mRNA前体的加工内容: 3. 内含子的剪接
1. 5 端添加帽子结构
mRNA的过程.
Spliceosome 剪接体
Self-splicing introns and mechanisms
Alternative splicing (可变剪接): 有些pre-mRNAs 可以通过不同的剪接方式产生不同的成熟的 mRNA. 60% of the human genes are spliced in this manner.
mRNA剪接编辑
ห้องสมุดไป่ตู้
真核生物基因组DNA的结构
真核基因大多是断裂的: 一个基因可由多个内含子和外显子间隔排
列而成。 内含子在真核基因中所占的比例很高,甚
至超过99%。
RNA
部分人类基因中内含子序列所占的比重分析
前体mRNA--hnRNA 5’-Cap 3‘-Poly(A)
mRNA 剪接
hnRNA
11 生物化学习题与解析--RNA的生物合成过程

R N A的生物合成过程一、选择题(一)A型题1.下列关于转录的叙述正确的是A.转录过程需RNA引物B.转录生成的RNA都是翻译模板C.真核生物转录是在胞浆中进行的D.DNA双链一股单链是转录模板E.DNA双链同时作为转录模板2.DNA上某段编码链碱基顺序为5'-ACTAGTCAG-3',转录后mRNA上相应的碱基顺序为A.5'-TGATCAGTC-3'B.5'-UGAUCAGUC-3'C.5'-CUGACUAGU-3'D.5'-CTGACTAGT-3'E.5'-CAGCUGACU-3'3.不对称转录是A.双向复制后的转录B.以DNA为模板双向进行转录C.同一单链DNA,转录时可以交替作为编码链和模板链D.同一单链DNA,转录时只转录外显子部分E.没有规律的转录4.真核生物的转录特点是A.发生在细胞质内,因为转录产物主要供蛋白质合成用B.转录产物有poly(A)尾,DNA模板上有相应的poly(dT)序列C.转录的终止过程需ρ(Rho)因子参与D.转录起始需要形成PIC(转录起始前复合物)E.需要α因子辨认起点5.下列关于转录编码链的叙述正确的是A.能转录生成mRNA的DNA单链B.能转录生成tRNA的DNA单链C.同一DNA单链不同片段可作模板链或编码链D.是基因调节的成份E.是RNA链6.Pribnowbox序列是A.AAUAAAB.TAAGGCC.TTGACAD.TATAATE.AATAAA7.真核生物的TATA盒是A.参与转录起始B.翻译的起始点C.RNA聚合酶核心酶结合位点D.σ因子结合位点E.复制的起始点8.原核生物DNA指导的RNA聚合酶由数个亚基组成,其核心酶的组成是A.α2ββ'(ω)B.α2β(σ)C.α2ββ'σ(ω)D.α2β'(ω)E.αββ' 9.原核生物识别转录起始点的是A.ρ因子B.核心酶C.RNA聚合酶的α亚基D.σ亚基E.RNA聚合酶的β亚基10.ρ因子的功能是A.参与转录的启动过程B.参与转录的全过程C.加速RNA的合成D.参与转录的终止过程E.可改变RNA聚合酶的活性11.在转录延长阶段,RNA聚合酶与DNA模板的结合是A.全酶与模板结合B.核心酶与模板特定位点结合C.结合松弛而有利于RNA聚合酶向前移动D.和转录起始时的结合状态没有区别E.结合状态相对牢固稳定12.下列关于转录因子(TF)的叙述正确的是A.是真核生物RNA聚合酶的组分B.参与真核生物转录的起始、延长和终止阶段C.是转录调控中的反式作用因子D.是真核生物的启动子E.是原核生物RNA聚合酶的组分13.真核生物转录终止A.需要ρ(Rho)因子B.需要释放因子(RF)C.与加尾修饰同步进行D.需要信号肽E.形成茎环形式的二级结构14.外显子是A.DNA的调节序列B.转录模板链C.真核生物的编码序列D.真核生物的非编码序列E.原核生物的编码序列15.DNA复制和转录过程具有许多异同点,下列关于DNA复制和转录的描述中错误的是A.在体内只有一条DNA链转录,而两条DNA链都复制B.在这两个过程中合成方向都为5'→3'C.两个过程均需RNA引物D.两个过程均需聚合酶参与E.通常情况下复制的产物其分子量大于转录的产物16.以下哪些代谢过程需要以RNA为引物A.DNA复制B.转录C.RNA复制D.翻译E.逆转录17.下列有关真核细胞mRNA的叙述,错误的是A.是由hnRNA经加工后生成的B.5'末端有m7GpppN帽子C.3'末端有poly(A)尾D.为多顺反子E.成熟过程中需进行甲基化修饰(二)B型题A.pppGB.PICC.TFD.TATAATE.AATAAA1.顺式作用元件2.反式作用因子3.真核生物的转录起始前复合物A.5'→3'B.3'→5'C.C端→N端D.N端→C端E.一个点向两个方向同时进行4.双向复制5.肽链的生物合成方向6.转录的方向A.DNA指导的RNA聚合酶B.RNA指导的DNA聚合酶C.DNA连接酶D.引物酶E.拓扑酶7.在复制中催化小片段RNA合成的酶8.RNA合成时所需的酶9.DNA合成时所需的酶A.DNA聚合酶B.RNA聚合酶C.逆转录酶D.核酶E.Taq酶10.化学本质为核酸的酶11.遗传信息由RNA→DNA传递的酶12.耐热的DNA聚合酶(三)X型题1.下列关于RNA生物合成的叙述,正确的是A.RNA聚合酶的核心酶能识别转录起始点B.转录复合物是由RNA聚合酶和DNA组成的复合物C.转录在胞质进行从而保证了翻译的进行D.DNA双链中仅一股单链是转录模板E.合成RNA引物2.真核生物mRNA转录后加工方式有A.在3'端加poly(A)尾B.切除内含子,拼接外显子C.合成5'端的帽子结构D.加接CCA的3'末端E.去掉启动子3.下列哪项因素可造成转录终止A.ρ因子参与B.δ因子参与C.在DNA模板终止部位有特殊的碱基序列D.RNA链3'端出现茎环结构E.RNA链3'端出现寡聚U与模板结合能力小4.真核生物的tRNAA.在RNA-polⅢ催化下生成B.转录后5'端加CCA尾C.转录后修饰形成多个稀有碱基(I、DHU、ψ)D.5'端加m7GpppN帽子E.二级结构呈三叶草型5.真核生物rRNAA.单独存在无生理功能,需与蛋白质结合为核蛋白体发挥作用B.在RNA-polⅠ作用下合成rRNA前体C.45S-rRNA剪切为5.8S、18S、28S三种rRNAD.45S-rRNA与蛋白质结合为核蛋白体E.转录后加工在细胞核内进行二、是非题1.复制和转录起始均需RNA引物。
河南农业大学考研专业课《现代分子生物学》考试试卷(913)

河南农业大学考研专业课《现代分子生物学》课程试卷(含答案)__________学年第___学期考试类型:(闭卷)考试考试时间:90 分钟年级专业_____________学号_____________ 姓名_____________1、分析题(5分,每题5分)1. 写出原核表达实验步骤。
答案:原核表达实验步骤如下:(1)按上述步骤提取该组织工作的RNA,反转录为cDNA。
(2)以cDNA为模板,用上述引物扩增出带酶切位点的A基因片段。
(3)用BamHI和EcoRI分别切割目的花絮和载体,连接酶将A 基因和载体连接起来构成重组质粒。
(4)将重组质粒转入感受态的大肠杆菌BL21中,涂布,根据载体所携带的抗性标记筛选出稳定遗传重组质粒媒介的单克隆。
(5)摇菌、扩大培养,待菌液生长到对数生长期,收集菌体,裂解、收集蛋白质。
(6)将总蛋白制备成溶液通过铜蛋白柱,由于A蛋白携带有His标签,可以被吸附在镍柱上才,然后加缓冲液将吸附黏附在镍柱上才的A蛋白洗脱下来,即可得到A蛋白溶液。
解析:2、判断题(55分,每题5分)1. 一染色体DNA经内切酶SalⅠ切割后,产生了若干个具有黏性末端的DNA片段,将这些片段分别在T4 DNA连接酶的作用下自身连接成环,然后导入受体细胞,都可以进行独立地复制。
()答案:错误解析:在T4 DNA连接酶的作用下,若干个具有黏性末端的DNA片段自身连接成弧,导入受体细胞后,并不是都可以进行独立的复制。
2. P53蛋白参与监控细胞核DNA的损伤,因此是一个肿瘤抑制因子。
()答案:正确解析:P53是人体非常重要的抑癌基因,参与监控细胞核DNA的损伤,它可以通过抑制细胞周期的进展,抑制细胞增殖。
但是它的缺失可以导致细胞增殖失控(无限增殖)。
3. 凝胶阻滞实验最初是用来对纯化的原核调控蛋白和DNA的互作进行动力学分析。
()答案:正确解析:凝胶阻滞分析是来研究DNA与特异性蛋白的相互作用,最初是用来对纯化的原核调控蛋白和DNA的互作进行动力学分析,通常是放射性标记的DNA片段与纯化蛋白,或提取物中的蛋白混合物相结合,然后在非变性凝胶中分析该产物。
第8章 RNA转录后加工 作业答案

第8章RNA转录后的剪接与加工一、名词解释:套索结构、顺式/反式剪接、分支点、剪接因子、剪接体、转脂反应、选择性剪接、RNA编辑二、填空题1.真核生物的mRNA 加工过程中,5′ 端加上____,在3′端加上____。
如果被转录基因产不连续的,那么____一定要被切除,并通过____过程将____连接在一起。
这个过程涉及许多RNA 分子,如U1 和U2 等,它们被统称为____。
它们分别与一组蛋白质结合成____。
并进一步地组成40S 或60S 的结构,叫作____。
帽子结构、多聚腺苷尾(poly(A)尾)、内含子、剪接、外显子、snRNA、snRNP、剪接体2.内含子之间以共同序列为界:5′剪接位点的____和3′剪接位点的____。
GU、AG3.真核生物前体t RNA 的加工包括____的切除和____的拼接。
随着5' 端和3' 端的序列切除,3'端加上了序列____,在四膜虫中前tRNA 内含子的切除和外显子的拼接是通过____机制。
内含子、外显子、CCA、自动催化三、单项选择题1、mRNA转录后的加工不包括()A.5’端加帽子结构B.3’端加polyA尾C.切除内含子D.连接外显子E.3’加CCA尾2、关于真核生物mRNA的聚腺苷酸尾巴,错误的说法是()A 是在细胞核内加工接上的B直接在转录初级产物的3’末端加上去的C 维持mRNA作为翻译模板的活性D先切除3’末端的部分核苷酸然后加上去的3、关于外显子和内含子叙述错误的是()A外显子是基因中编码序列,并表达为成熟RNA的核酸序列B 外显子能转录,内含子不能转录C去除内含子,连接外显子的过程叫拼接D 基因中外显子加内含子的长度相当于hnRNA的长度E 基因中外显子和内含子相互间隔排列4、下列关于mRNA的叙述正确的是()A 3’末端含有CCA—OH B二级结构呈三叶草型C 5’末端有“帽子”结构D 含许多稀有碱基5、关于tRNA叙述错误的是()A 在真核细胞核内,由RNA聚合酶Ⅲ催化合成其初级产物B 二级结构呈三叶草型C 3’末端有CCA—OHD 含有许多稀有碱基E 5’末端有多聚A尾巴6、下列哪一种反应不属于转录后修饰()A.腺苷酸聚合B.去除外显子C.端加帽子结构D.去除内含子E.甲基化7、外显子是()A.不能被转录的序列B.被转录但不被翻译的序列C.被转录也被翻译的序列D.以上均不对8、参与RNA剪接的是()A.mRNA B.tRNA C.45SrRNA D.snRNA9、真核生物成熟的mRNA5'端具有()A.m7ApppNp B.m7GpppNp C.m7UpPpNpD.m7CpppNp E.以上都不是10、真核生物结构基因包括()A.外显子和内含子B.内含子C.外显子D.两者都不是11、真核生物转录终止修饰点序列是()A.TATA box B.AATAAA和其下游GT序列C.GC box D.AAUAAA E.Pribnow盒12、在真核细胞中,下列哪种杂交能完全配对()A.DNA-hnRNA B.DNA-mRNAC.DNA-成熟的tRNA D.DNA-18S-rRNA13、关于外显子和内含子的叙述,正确的是()A.外显子在DNA模板上有相应的互补序列,内含予没有B.hnRNA上既有外显子也有内含子序列C.原核生物mRNA有内含子D.除去外显子的过程称为剪接E.成熟的mRNA有内含子14、关于mRNA的描述,哪项是错误的? ()A.原核细胞的mRNA在翻译开始前需加聚A尾巴B.原核细胞的许多mRNA携带着几个多肽链的结构信息C.真核细胞mRNA在端携有特殊的“帽子”结构D.真核细胞转录生成的mRNA经常被“加工”E.真核细胞mRNA是由RNA聚合酶Ⅱ催化合成的15、下列关于RNA分子中“帽子”的叙述,哪一项是正确的? ()A.可使tRNA进行加工过程B.存在于tRNA的-末端C.是由聚A组成D.仅存在于真核细胞的mRNA上E.用于校正原核细胞mRNA翻译中的错误16、转酯反应()。
mrna异常剪接体外验证实验技术的流程

在讨论 mRNA 异常剪接的外部验证实验技术的流程之前,我们首先需要了解 mRNA 异常剪接的含义和意义。
在此之后,我们将介绍这一验证实验技术的流程,以及其在研究中的重要性和应用价值。
我们会共享一些个人观点和理解。
让我们深入探讨。
1. mRNA 异常剪接的含义和意义mRNA 异常剪接指的是在转录后的 mRNA 水平上发生的剪接异常,而不是在 DNA 水平上。
这种异常剪接可能导致编码蛋白质的错误,进而影响基因的功能。
这种异常在许多疾病中都起着关键作用,包括癌症、遗传性疾病和自身免疫性疾病。
研究 mRNA 异常剪接及其验证实验技术对于理解疾病的发生和发展至关重要。
2. mRNA 异常剪接的外部验证实验技术流程外部验证实验技术是通过实验手段对某一假设或猜想进行验证的过程。
在研究 mRNA 异常剪接的外部验证实验技术中,一般包括以下步骤:(1) 样本准备:选择合适的细胞系或组织样本,以及相应的对照样本,进行实验前的样本准备工作。
(2) RNA 提取:从样本中提取RNA,以获得研究所需的mRNA 样本。
(3) RT-PCR:利用逆转录-聚合酶链反应技术,将 mRNA 转录为cDNA,然后进行PCR 扩增,以检测mRNA 异常剪接的存在与比较。
(4) 凝胶电泳:将扩增后的产物进行凝胶电泳分析,以观察不同样本间mRNA 异常剪接产物的差异。
(5) Sanger 测序:对凝胶电泳检测到的异常剪接产物进行 Sanger 测序,以确定具体的剪接位点和剪接类型。
(6) 数据分析:对实验结果进行分析,判断是否存在mRNA 异常剪接,并对其进行定量和定性描述。
3. mRNA 异常剪接的外部验证实验技术在研究中的重要性和应用价值外部验证实验技术是验证和验证科研成果的关键手段之一。
在研究mRNA 异常剪接的外部验证实验技术中,其重要性和应用价值主要体现在以下几个方面:- 确认研究发现:通过外部验证实验技术,可以对初步研究结果进行验证,从而保证研究的可靠性和科学性。
mRNA选择性剪接的分子机制

:+(’ , +(!"#$ %&’&#("$ )(’("$ , !"#$"%& ’((),*+(+)
.//0 (*-1 2 )+-’
! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! !
LM%$@ @N:FOD4 &) *% P : Q:3D5N38; QD5,8(4/" :7 "#$% %39D;(894RD ’23454(6
!+<
!
"#$% 剪接的基本模式
以外显子和内含子为单元的基因结构形式在真
可以被选择保留或切除 (图 !, ; 多个外显子可以 D) 进行不同组合的可变拼接 (图 !, ; 内含子可以被选 7) 择保留在 "#$% 中 (图 !, 等。这些不同的剪接形 6) 式形成了不同的剪接组合, 产生了不同的剪接产物。 有时候这种剪接组合产生的产物数目是极其惊人 的。如果蝇的 ?/58" 基因经选择性剪接产生的产物 达 <H +++ 余种, 超过果蝇整个基因组基因数目的两 倍。
-/01 选择性剪接的分子机制
章国卫,宋怀东,陈 竺
(上海市瑞金医院上海血液学研究所,上海 ’(((’K)
摘
要:真核细胞 5N0E 前体经过剪接成为成熟的 5N0E, 而 5N0E 前体的选择性剪接极大地增加了蛋白质的多
样性和基因表达的复杂程度, 剪接位点的识别可以以跨越内含子的机制 (内含子限定) 或跨越外显子的机制 (外显 子限定) 进行。选择性剪接有多种剪接形式: 选择不同的剪接位点, 选择不同的剪接末端, 外显子的不同组合及内 含子的剪接与否等。选择性剪接过程受到许多顺式元件和反式因子的调控, 并与基本剪接过程紧密联系, 剪接体 中的一些剪接因子也参与了对选择性剪接的调控。选择性剪接也是 + 个伴随转录发生的过程, 不同的启动子可调 控产生不同的剪接产物。5N0E 的选择性剪接机制多种多样, 已发现 N0E 编辑和反式剪接也可参与选择性剪接过 程。 关键词:5N0E;选择性剪接 中图分类号:WK’’ 文献标识码:E 文章编号:(*-14)+-’ (’(()) (+4(+(’4(X
暨南大学《分子生物学》复习题

暨南大学《分子生物学》复习题分子生物学复习题Macromolecules(大分子)Supercoil(超螺旋):DNA双螺旋本身进一步盘绕称超螺旋。
超螺旋有正超螺旋和负超螺旋两种,负超螺旋的存在对于转录和复制都是必要的。
Palindrome(回文序列):DNA序列中一条链从左到右阅读和另一条链从右到左阅读是一样的序列,即两条链由相邻的反向重复序列组成。
melting temperature(解链温度):DNA在溶液中随温度升高逐步变性,当A260达到彻底变性时,A260的一半时的温度就叫解链温度(Tm),Tm与GC比例、序列复杂性等因素有关。
hyperchromic shift(增色效应):当双螺旋DNA融解(解链)时,260nm处紫外吸收增加的现象。
Nucleosome(核小体):真核生物染色体的基本结构单位,由DNA与组蛋白H1、H2A、H2B、H3、H4组成的,DNA以左手螺旋缠绕于组蛋白核心上Chaperones(伴侣蛋白):与新合成的多肽链形成复合物并协助它正确折叠成具有生物功能构向的蛋白质Domain(结构域):多肽链在二级结构或超二级结构的基础上形成三级结构的局部折叠区,它是相对独立的紧密球状实体,称为结构域。
Motif(基序):又称超二级结构,是蛋白质分子中特别是球状蛋白质分子中由若干相邻的二级结构与元件(主要是α螺旋和β折叠片)组合在一起,彼此相互作用,形成种类不多的、有规则的二级结构组合或二级结构串,在多种蛋白质中充当三级结构的构件。
protein family(蛋白质家族):指具有相似结构,有某一功能共性的一组蛋白质。
Proteasome(蛋白酶复合物):蛋白酶体存在于所有真核细胞的胞质及核内,是高度保守具有多种催化功能的蛋白酶复合物。
蛋白酶体具有蛋白水解酶活性,蛋白酶体水解作用需要泛素蛋白参加。
Ubiquitylation(蛋白质泛素化):泛素间隔或连续地附着到被降解的蛋白质赖氨酸残基上,这一过程称为蛋白质泛素化。
第八章RNA的生物合成转录

• 如前所述,某些抗生素如利福霉素和利福平能抑 制细菌RNA聚合酶,因而能抑制细菌RNA的合成。
第18页/共60页
第三节 转录后的加工和修饰
• 真核细胞中转录生成的RNA是初级转录产物,均需经过一定程度的加工才具有活性。原核细胞由于没有细 胞核,其结构基因是连续的核苷酸序列,转录后产生的RNA很少需要加工处理(tRNA例外)就转运到核蛋 白体上参与蛋白质的合成。新生的无活性的RNA转变为有活性的RNA的过程,称为RNA的成熟(转录后的 加工),包括链的断裂、拼接和化学修饰等。
tRNA
氨基酸臂
第30页/共60页
反密码环
• (三)rRNA与蛋白质构成核蛋白体是蛋白质合成的场所 rRNA是一类分子量不等 的非均一性RNA,它们与多种蛋白质互相镶嵌,结合成为显微镜下可见的核蛋白体 颗粒,是氨基酸聚合成肽链的场所。核蛋白体由大、小亚基构成。大亚基有转肽酶 活性和两个tRNA结合部位,一个是结合肽酰-tRNA的部位(P位),另一个是结合 氨基酰-tRNA的部位(A位)。小亚基有结合模板mRNA的功能,在大小亚基之间 有容纳mRNA的部位,核蛋白体能沿着mRNA5′→3′方向阅读遗传密码。
第20页/共60页
• (二).mRNA的剪接 • 真核细胞的mRNA前体称核内不均一RNA(hnRNA),它是由断裂基因转录的,包
含有内含子和外显子的区段,所以其分子量比成熟的mRNA大几倍,甚至数十倍。 剪接就是把hnRNA中的内含子除去,把外显子拚接起来,成为具有翻译功能的模 板—mRNA。
第21页/共60页
第31页/共60页
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
这 三 种 帽 子 都 有 特 殊 面 对 面 核 苷 酸 结 构 ( confronted nucleotide structure)。
HN—CH3
m7G
帽子0
mRNA 5端的帽子位臵 和可被甲基化的位点
三种5’帽结构形式
帽0 普遍存在
m7GpppXpYp------
帽1
m7GpppXmpYp-----
mRNA前体的加工内容: 3. 内含子的剪接
1. mRNA依赖snRNA的拼接
2.选择拼接
3.顺式和反式拼接 4. I型自我拼接 5. II型自我拼接
1. 5 端添加帽子结构 2. 3 端添加多聚腺苷酸
真核生物成熟mRNA形成的过程
断裂基因发现不久,Crick(1978年)提出了一系 列发人深思的问题:
普遍存在
帽2
m7GpppXmpYmp-----
很少发生
帽子结构的功能
1.
2. 3. 4.
5.
有助于mRNA跨越核膜转运; 保护 mRNA 5′不被酶降解; 使mRNA能与核糖体小亚基结合; 被蛋白质合成的起始因子所识别,促进蛋白质 合成; 提高剪接效率
2. Poly A 尾巴
多聚腺苷化的信号及参与的蛋白因子
加帽的顺序
当新生RNA链长度约为25nt 时,其5’端经修饰被加上 一个7-甲基G,以5’-5’三磷酸连接。 RNA三磷酸酶(Triphosphatase) 鸟苷酸转移酶(guanyltransferase) 鸟嘌呤-7-甲基转移酶(guanosine ethyltransferase) 反应步骤如下: 核苷酸水解酶从RNA的5‘-端切掉γ 基团; 鸟苷酸转移酶将来自GTP的GMP连接到RNA 5’-端的β 磷酸上,形成5-5三磷酸,释放出PPI 甲基转移酶催化SAM的甲基转移到Cap-0的N7位。
(1)剪切作用若是通过酶来进行的,那么这种剪接 酶是怎样识别RNA上特定的位点? (2)剪切酶有没有特异性?对不同的RNA是否需要 不同的酶? (3)切除的RNA是以线状还是环状存在的?切下的 RNA的命运将如何?
GU-AG法则
Chambon等分析比较了大量结构基因的内含子切割 位点,发现有2个特点: (1)内含子的两个末端并不存在同源或互补; (2)连接点具有很短的保守序列,亦称边界序列。
拼接体组装 第一次转酯:左外元、内元剪切套索 第二次转酯:exons连接、套索状内元释放 拼接体(spliceosome)解体与套索(lariat)降解 同步
The simplified mechanism of nuclear mRNA precursor
2‘-hydroxyl group of A residue of branchpoint
mRNA的剪接
转录产生的核内RNA前体分子与蛋白质结合,形 成RNA和蛋白质组成的snRNP复合物(ribonucleoprotein particle)。 随着RNA链的延伸,每个内含子5’和3’两端的复 合物成对联结,产生60S的颗粒—剪接体 (spliceosome),进行RNA前体分子的剪接。
Step1
RNA三磷酸酶
鸟苷转移酶
GMP
Step2
SAM
Step3
CAP1:OCH3
CAP2:OCH3
CAP 0
帽子的类型
在末端鸟苷的第7位上存在单个甲基化位点的称O类帽子 (capO);
在次末端核苷酸的核糖上的2′-O位点上还有一个甲基位
点的称1类帽子(cap 1);
此外,在第三个核苷酸的核糖上(2′-O)有甲基化位点 的称2类帽子(cap2)。
100种内含子的5′端都是GU;3′端都是AG,因此称为 GU-AG法则(GU-AG rule),又称为Chambon法则。
5’splice site (5’剪接位点): the exon-intron boundary at the 5’ end of the intron 3’ splice site (3’剪接位点): the exonintron boundary at the 3’ end of the intron Branch point site (分枝位点): an A close to the 3’ end of the intron, which is followed by a polypyrimidine tract (Py tract).
mRNA剪接编辑
真核生物基因组DNA的结构
真核基因大多是断裂的: 一个基因可由多个内含子和外显子间隔排 列而成。 内含子在真核基因中所占的比例很高,甚 至超过99%。
RNA
部分人类基因中内含子序列所占的比重分析
前体mRNA--hnRNA
5’-Cap
3‘-Poly(A)
mRNA 剪接
成熟mRNA的转运
Spliceosome(剪接体)
剪接过程由 U1, U2, U4, U5 , U6 snRNPs催化,也有 其他剪接因子的参与。 这些snRNPs中的RNA成分与 5’-和 3’剪接点及分支 点的多种保守碱基配对。 Splicesome: The large RNA-protein body on which splicing of nuclear mRNA precursors occurs.
hnRNA
hnRNA (heterogeneous nuclear RNA):核内不 均一RNA,是mRNA的原初转录产物在核 内形成的大小不等的中间物,相对分子质 量大,也不均一,成为hnRNA。 hnRNA与mRNA的关系:
两者具有部分相同的序列; 两者都可以做翻译的模板; 两者都有5-端帽子和3-端polyA尾巴; 两者都是由RNA聚合酶转录
hnRNP
hnRNP:核糖核蛋白颗粒
hnRNA的物理结构是核糖核蛋白颗粒 (hnRNP),颗粒中蛋白质包围着hnRNA, 这种hnRNA和蛋白质组成的复合物称为 hnRNP。
1. mRNA 加帽
帽子结构 (m7GpppGp-) 7-甲基鸟核苷三磷酸:它由甲基化 鸟苷酸经焦磷酸化与mRNA的5‘末端核苷酸相连,形成 5’,5‘-三磷酸连接(5’,5‘-triphosphate linkage), mRNA5’端 的这种结构称为帽子(cap)。 反应空间:在mRNA的5‘-端加上m7GTP的结构。此过程 发生在细胞核内,即hnRNA即可进行加帽。 加工过程:首先是在磷酸酶的作用下,将5'-端的磷酸基水 解,然后再加上鸟苷三磷酸,形成GpppN的结构,再对G 进行甲基化。
剪接体中的snRNP 的功能
snRNP snRNA(nt) 在拼接中可能的作用
U1
U2 U5/ U4/ U6
165
185 116/ 145/106
通过序列互补,结合到5拼接位点上
结合到分枝位点(U2AF),催化转酯反 应,与U6配对 三聚体 U5: 结合5与3拼接点外显子 U4:拼接体的组装 U6:结合5拼接点,与U2一起催化转 酯反应
mRNA的多聚腺苷酸化过程
CPSF因子专一性与加尾信号AAUAAA结合 CstF因子与富GU区结合 CFI、CFII在加尾信号下游的15~25 nt处切割 poly(A)多聚酶催化多聚腺苷酸化,先添加约10 个腺苷酸,速度较慢 poly(A)结合蛋白的结合加快多聚酶延伸速度, 控制多聚腺嘌呤尾巴的长度
有效的信号是: AAUAAA及随后的 23-24nt的富GU区, 再后的富U区。 参与的蛋白因子: CPSF,CstF,CFI, CFII
加尾反应需要的顺式元件
3 端添加多聚腺苷酸
长度:20~200 nt 加尾信号: 切割与加尾: a. 剪切/多聚腺苷酸化特异性因子(cleavage and polyadenylation specificity factor, CPSF) b. 剪切刺激因子(cleavage stimulation factor, CstF) c. 剪切因子I,II(CFI, CFII) d. poly(A)多聚酶(polyadenylate polymerase, PAP) e. poly(A)结合蛋白(poly (A) binding protein, PBP)
Spliceosome 剪接体 Self-splicing introns and mechanisms
Alternative splicing (可变剪接): 有些pre-mRNAs 可以通过不同的剪接方式产生不同的成熟的 mRNA. 60% of the human genes are spliced in this manner.
RNA加工过程及其生理功能
断裂基因 hnRNA,hnRNP,sRNA Exons (外显子): the coding sequences Introns (内含子) : the intervening sequences RNA splicing: 去除pre-mRNA的内含子形成成熟 mRNA的过程.
CPSF
PAP
PAPB
Pol II的CTD促进切割
3 端多聚腺苷酸尾巴的功能
提高mRNA的稳定性 在核-质转运中起作用 提高mRNA的翻译效率 影响最后一个内含子的切除
创造终止密码子UGA
选择性加尾调节基因的表达
3. RNA剪接(Splicing)
真核生物mRNA前体的剪接
细胞核中的小分子RNA称为细胞核小RNA(small nuclear RNA,snRNA); 位于细胞质中的称为细胞质小RNA(small cytoplasmicRNA,scRNA)。 在自然状态下,它们以核糖核蛋白颗粒(SnRNP 和scRNP)的形式存在,俗称snurps和scyrps。 在核仁中也存在着一类小的RNA,称为核仁小 RNA(small nucleolarRNA, snoRNA),它们在rRNA 的加工中起作用。 snRNA参与剪接过程,并于其它蛋白一起构成一 个大的颗粒复合体,称为剪接体(splicesome)。