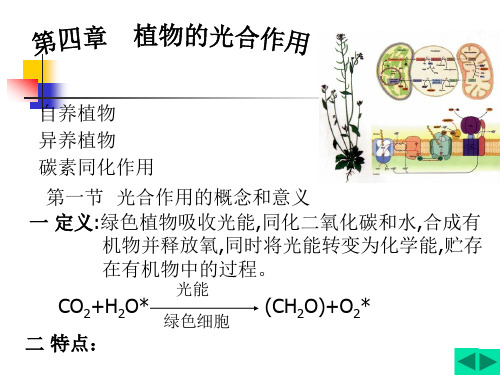
4 第四章 植物的光合作用
植物生理学光合作用课件

类型
非环 式电 子传 递
环式 电子 传递
原初电子供体P 原初电子受体A 次级电子供体D 蛋白质
维持微环境
直接供给电子的物质
光化学反应
D·P ·A
h→ v
* D· P ·A
++
-
-
→ D·P ·A →D ·P ·A
由光引起的反应中心色素分子与原初 电子受体、供体间的氧化还原反应
二、电子传递与光合磷酸化+来自ee-e eD ·P ·A
场所:光合膜 特点:受光促进,不受温度影响
光化学反应
激发态
第二单线态
放热
第一单线态
放荧 热光
放热
磷 光
Chl
三线态
基态
吸收光能
基态
激发态
第一单线态 第二单线态 第一三线态
第一单线态Chl分子的去向:
放热
发射荧光(溶液) 进入第一三线态
浪费!
光化学反应 (活体)
第二单线态Chl分子不能直接用于光合作用
吸收光谱---
PSI和PSII串联
二处逆电势梯度 PQ穿梭(ΔμH+ )
(三)水的光解和放氧 P156 光
2H2O* + 2A 叶绿体 2AH2 + O2*
氧化剂
----希尔反应
(四)光合磷酸化
1.概念 P158
条件--光下 部位--叶绿体 原料--ADP+Pi 产物--ATP
2.类型
驱动力---ΔμH+
一、原初反应
场所:光合膜
特点:速度快,需光
概念:P152
光能吸收
浙教版初中科学教案《植物的光合作用》

浙教版初中科学教案《植物的光合作用》一、教学内容本节课选自浙教版初中科学教材第四章第一节《植物的光合作用》。
教学内容详细阐述了植物光合作用的基本概念、原理、过程以及在实际生活中的应用。
二、教学目标1. 让学生理解光合作用的概念、原理及过程,掌握光合作用公式的书写和含义。
2. 培养学生运用所学知识解释自然现象和实际问题的能力。
3. 增强学生的环保意识,培养学生关爱植物、保护生态环境的观念。
三、教学难点与重点教学难点:光合作用的过程及其公式的理解。
教学重点:光合作用的概念、原理及在实际生活中的应用。
四、教具与学具准备教具:多媒体课件、实物模型、实验器材。
学具:笔记本、笔、实验报告单。
五、教学过程1. 导入:通过展示植物生长的图片,引导学生思考:为什么植物能够生长?生长过程中需要什么?2. 新课导入:介绍光合作用的概念,引导学生学习光合作用的原理。
3. 实践情景引入:进行光合作用实验,让学生观察实验现象,引导学生探讨光合作用的过程。
4. 例题讲解:讲解光合作用的相关例题,帮助学生巩固所学知识。
5. 随堂练习:布置随堂练习,检验学生对光合作用知识的掌握。
6. 知识拓展:介绍光合作用在实际生活中的应用,如绿色能源、环保等。
六、板书设计1. 板书植物的光合作用2. 板书内容:光合作用概念光合作用原理光合作用过程光合作用应用七、作业设计1. 作业题目:解释光合作用的概念及公式。
描述光合作用的过程。
论述光合作用在实际生活中的应用。
2. 答案:光合作用是指植物利用光能将二氧化碳和水合成有机物和氧气的过程。
光合作用过程包括光反应和暗反应两个阶段。
光合作用在生活中有广泛应用,如绿色能源、环保等。
八、课后反思及拓展延伸本节课通过实践情景引入、例题讲解、随堂练习等方式,帮助学生掌握了光合作用的相关知识。
课后,教师应关注学生对光合作用的理解程度,及时解答学生的疑问。
同时,可引导学生进行课后拓展学习,如研究光合作用与生态环境的关系,培养学生的环保意识。
第四章-光合作用

原初反应 (primary reaction) 指光合作用中最 初旳反应,从光合色素分子受光激发起到引起 第一种光化学反应为止旳过程,它涉及光能旳 吸收、传递与光化学反应。原初反应旳成果使 反应中心发生电荷分离。
光合单位 (photosynthetic unit) =聚光色素+反应中心
反应中心色素分子 (reaction center pigment) 是处于反应中心中旳 一种特殊性质旳叶绿素 a 分子,它不但能捕获光能,还具有光化学活 性,能将光能转换成电能。光系统Ⅰ和光系统Ⅱ旳反应中心色素分子 分别是 P700 和 P680 ,这里 P 代表色素 (pigment) , P 后旳数值代 表色素分子在受光激发被氧化时,该色素分子吸收光谱中变化最大旳 波长位置,也即用氧化态吸收光谱与还原态吸收光谱间旳差值最大处 旳波长来作为色素分子旳标志。 P700 和 P680 表达它们受光激发被 氧化时,吸收光谱中变化最大旳波长位置分别是近 700nm 和 680nm 处。是光能旳“捕获器”、“转换器”。
➢ Chl(基态)+hυ 10-15s Chl*(激发态)
激发态旳叶绿素分子 回至基态时,能够光 子形式释放能量。
处于第一单线态旳叶 绿素分子回至基态时 所发出旳光称为荧光。
而处于三线态旳叶绿 素分子回至基态时所 发出旳光称为磷光。
➢因为叶绿素分子吸收旳光能有一 部分消耗在分子内部旳振动上,且 荧光又总是从第一单线态旳最低振 动能级辐射旳,辐射出旳光能肯定 低于吸收旳光能,所以叶绿素旳荧 光旳波长总要比被吸收旳波长长些。
温保鲜旳原因之一
(3) 营养元素
➢ 叶绿素旳形成必须有一定旳营养元素。
➢ 氮和镁是叶绿素旳构成成份,铁、锰、铜、锌等则在叶绿素旳生物合 成过程中有催化功能或其他间接作用。
第四章第一节绿色植物通过光合作用制造有机物

点
2、确保实验顺利进行,对实验结果进行深入的分析。
课前准备
总体要求:1.“统一”设计“分段”教学;2.围绕“三维”落实“三问”;3.充实“心案”活化“形案”。
教学流程
分课时
环节
与时间
教师活动
学生活动
△设计意图
◇资源准备
□评价○反思
新授
一、实验
绿叶在光下制造淀粉
指导学生实验
并提示注意事项
出示指导学生实验的分组表格。
教学设计
题目
第一节绿色植物通过光合作用制造有机物
总课时
1
学校
星火一中
教者
孙亚东
年级
七
学科
生物
设计来源
自我设计
教学时间
教材分ຫໍສະໝຸດ 析这是一节以实验为主的课。在这节课的教学中,不能简单地让学生按照传统的教学方式,照猫画虎地完成实验,而应利用这个实验训练学生设计实验的技能。可以在实验开始前,让学生先尝试设计检验绿叶在光下制造有机物的实验方案,然后对照教科书上的实验指导,找出自己设计的实验的不足。然后引导学生讨论。
学情分析
注意引导学生思考每个实验步骤中包含的科学原理和方法,让学生带着问题做实验,使学生逐渐认识到在科学探索的过程中,要遵循一定的科学原理、科学方法,还需要掌握一定的操作技能。
教
学
目
标
1.阐明绿色植物通过光合作用制造有机物。
2.运用实验的方法检验绿叶在光下制造淀粉。
重
点
1、探究绿叶能否在光下制造淀粉。
巡视指导学生实验。
二、分析实验结果----
光合作用需要光,能产生淀粉
三、光合作用的意义
指导学生看书,引导学生作出概括。
4植物的光合作用和呼吸作用(原卷版)

一、光合作用1、绿色植物的光合作用步骤:暗处理→叶片遮光→光照→酒精脱色→漂洗→滴碘液检验。
a.暗处理的目的:让叶片中原有的淀粉运走耗尽。
b.叶片遮光的目的:进行对照实验。
c.酒精脱色:溶解叶绿素;酒精要水浴加热,目的是防止酒精燃烧发生火灾。
且受热均匀。
①结果分析:叶片遮光部分遇碘不变蓝;光照部分遇碘变蓝。
①结论:光是光合作用的必要条件,淀粉是光合作用的产物之一。
步骤:植株暗处理→光照几小时→叶片酒精脱色→漂洗→滴碘液检验。
a.暗处理的目的:让叶片中原有的淀粉运走耗尽。
b.密封花盆的目的:防止外界空气的干扰。
c.氢氧化钠溶液:吸收瓶内二氧化碳。
d.清水:起对照作用。
①结果分析:放置氢氧化钠的一组遇碘液不变蓝;放置清水的一组遇碘液变蓝。
①结论:二氧化碳是光合作用的原料。
(1)提高光能利用效率的方法:合理密植等。
(2)提高作物产量的方法:增加二氧化碳的浓度;增加光照强度和延长光照时间;适当调节昼夜温差等。
二、绿色植物的呼吸作用1、影响光合作用、呼吸作用外界因素①光照强度。
光照增强,光合作用随之加强。
但光照增强到一定程度后,光合作用不在加强。
夏季中午,由于气孔关闭,影响二氧化碳的进入,光合作用反而下降。
因而中午光照最强的时候,并不是光合作用活动最强的时候。
①二氧化碳浓度。
二氧化碳是光合作用的原料,其浓度影响光合作用的强度。
温室种植蔬菜可适当提高大棚内二氧化碳的浓度以提高产量。
①温度。
植物在10~35①正常进行光合作用,其中25~30①最适宜,35①以上开始下降,甚至停止。
温度、水分、氧气和二氧化碳浓度是影响呼吸作用的主要因素。
①温度。
温度对呼吸作用强弱影响最大。
温度升高,呼吸作用加强;温度过高,呼吸作用强度减弱。
①水分。
植物含水量增加,呼吸作用加强。
①氧气。
一定范围内随氧气浓度的增加,呼吸作用显著加强。
①二氧化碳。
二氧化碳浓度大,抑制呼吸作用。
在贮藏蔬菜、水果、粮食时用低温、干燥、充加二氧化碳等措施可以延长贮藏时间。
植物生理学 4.光合作用
组成:由核心复合体、 PS ΙΙ捕光复合体和放氧复合体 (OEC)组成。
核心复合体:由6种多肽组成。 其反应中心=Tyr+P680+pheo
捕光复合体:LHCΙΙ
放氧复合体:OEC,位于PS ΙΙ的类囊体膜腔表面,
由多肽和与放氧有关的锰复合体、氯和钙离子组
成。水在光照下经过PS ΙΙ的作用,发生水裂解,
(二)光系统
1 红降现象:
2 双光增益效应(爱默生效应): 3 光系统:光系统Ι (PS Ι )、光系统ΙΙ (PS ΙΙ ) PS I 为小颗粒,存在于基质片层和基粒片层的非垛叠区。 组成:反应中心P700、电子受体和PS Ι 捕光复合体三
部分组成。 光反应:适合长光波反应。
PS ΙΙ
其颗粒较大,受敌草隆抑制。存在于基粒片层的垛叠区。
(二)叶绿体的结构
叶绿体膜 外膜:透性大 内膜:透性小,主要控制物质进出的屏障。
组成:主要为可溶性蛋白质(酶)和其它代谢活跃的
基质
物质,呈高度流动性状态,具有固定二氧化碳
(间质)
的能力。(光合作用的暗反应即淀粉的形成与
贮存是在此进行的 。)
嗜饿颗粒(滴)(脂滴):是一类易与饿酸结合的颗
粒,其主要成分是亲脂性的醌类物质。功能是:
叶绿素a/叶绿素b=3/1 叶黄素/胡萝素=2/1
2 红色: 气温、可溶性糖、花色素(红色)
3 黄色:
叶绿素受破坏
光反应:在光下, 1 原初反应(指对光能的吸收、传递和转
在叶绿体的类囊
换的过程。)
体膜上进行的, 由光所引起的光
光 化学反应。实质
光能 原初反应
电能(电子)
(光量子)
2 电子传递和光合磷酸化(指把原初反应
植物生理学之 第四章 植物的光合作用

第四章植物的光合作用一、名词解释1.光合作用2.光合午休现象3.希尔反应4.荧光现象与磷光现象5.天线色素6.光合色素7.光合作用中心8.光合作用单位9.红降现象10.双光增益现象11.C3途径12.C4途径13.光合磷酸化14.非环式光合磷酸化l5. 量子效率16.暗反应17.同化力18.光反应19.CAM途径20.光呼吸21.表观光合速率22.光饱和点23.光补偿点24.CO2饱合点25.CO2补偿点26.光能利用率27.瓦布格效应28.原初反应29.碳素同化作用30.叶面积指数二、将下列缩写翻译成中文1.CAM 2.Pn 3.P700 4.P680 5.LHC 6.PSl 7.PSⅡ8.PQ 9.PC 10.Fd 11.Cytf12 12.RuBP 13.3-PGA 14.PEP l5.GAP 16.DHAP 17.OAA 18.TP 19.Mal 20.ASP 21.SBP 22.G6P 23.F6P 24.FDP 25.LAI 26.X5P 27. Fe-S 28. Rubisco 29.P* 30.DPGA三、填空题1.叶绿体的结构包括______、______、______和片层结构,片层结构又分为_____和______。
2.光合色素可分为______、______、______三类。
3.叶绿素可分为______ 和______两种。
类胡萝卜素可分为______和______。
4.叶绿素吸收光谱的最强吸收带在______ 和______。
5. 光合作用原初反应包括光能的______过程。
6. 叶绿体色素中______称作用中心色素,其他属于______。
7. 缺水使光合速率下降的原因是______、______、______。
8. 卡尔文循环中,同化1分子CO2需消耗______分子ATP和______ 分子NADPH+H+。
9. 高等植物CO2同化的途径有______、______、______三条,其中最基本的是______。
八年级科学上册第4章植物的物质和能量的转化4《植物的光合作用和呼吸作用》教案华东师大版(最新整理)

《植物的光合作用和呼吸作用》教案教学目标1。
知识与技能概述绿色植物光合作用与呼吸作用的关系、测量光合作用的指标以及它们之间的关系.2.过程与方法(1)通过解析有关绿色植物光合作用与呼吸作用的练习题,一方面学会解析题目的方法,另一方面理解绿色植物光合作用与呼吸作用的关系,并能总结出测量光合作用的指标以及它们之间的关系。
(2)运用所学绿色植物光合作用与呼吸作用的知识解析实际生活中的自然现象。
3.情感态度与价值观(1)通过小组比赛的形式激发学习研究的热情,逐步形成锲而不舍、探索创新的学习态度,促进自身的可持续发展。
(2)增强关注自然、关注生活、热爱生活的积极的生活态度。
(3)采用小组合作的学习方式,在合作交流中获得知识、促进理解、提高能力,潜移默化地培养学生正确的价值观和人生观。
教学重点测量光合作用的指标以及它们之间的关系。
教学难点运用测量光合作用的指标以及它们之间的关系解析练习题。
教学过程课前准备比赛每个学习小组解析两道有关绿色植物光合作用与呼吸作用的练习题。
导入多媒体演示新疆素有“瓜果之乡"的美称。
在民间流传着一首优美的“顺口溜”,对新疆不同地区的不同瓜果加以概括。
【学生活动】讨论为什么新疆的水果比其他内地地区的水果含糖等有机物多?【教师】①吐鲁番地区属于高纬度地区,日照时间比内地地区长,光合作用时间长;②吐鲁番地区属于沙漠性气候,日夜温差大,有利有机物质的积累。
通过这节课的学习我们会有更全面的认识。
【教师】关键一绿色植物光合作用与呼吸作用的关系。
【学生活动】讨论总结.【教师】绿色植物光合作用与呼吸作用的关系绿色植物既进行光合作用,也进行呼吸作用,光合作用进行的必要条件是有光,呼吸作用则时刻进行与光无关。
【学生活动】分析下图,植物体中光合作用与呼吸作用的关系。
【教师】光合作用等于呼吸作用,植物不从大气中吸收CO2也不释放O2到大气中。
光合作用小于呼吸作用,植物从大气中吸收O2且释放CO2到大气中。
植物生理学第四章光合作用

光合作用的全过程分为三大步骤:
①原初反应 ②电子传递和光合磷酸化
(光反应)
类囊体膜上进行
③碳素同化 (暗反应)基质中进行
光反应
光能的吸 收、
传递和转 换
电子传递和 光合磷酸化
ATP 形成同化力
NADPH
一、原初反应 原初反应指从光合色素分子被光激发开始到引
起第一个光化学反应为止的过程。
一、原初反应(primary reaction)
四、叶绿素的形成
1. 叶绿素的生物合成(图4-8) ⑴起始物质:谷氨酸或α-酮戊二酸; ⑵重要中间产物:δ-氨基酮戊酸(5-氨基酮戊
酸,原卟啉Ⅸ (protoporphyrin Ⅸ)等;
2. 影响叶绿素形成的条件 ① 光:原叶绿酸酯转变为叶绿酸酯需要光照;但强光下
叶绿素会被氧化. ② 温:最低温2℃、最适温30℃、最高温40℃,高温下
光合链始端是H2O光解产生电子,终端是还原NADP+ 产生NADPH+H+ 。
过程:H2O→PSⅡ复合体→PQ→Cytb6f复合体→ PC →PSⅠ复合体→Fd→NADP+(产生NADPH+H+)
⑶非循环式电子传递 (noncyclic electron transport)
定义:高能电子从H2O到NADP+的跨类囊体膜传 递途径是非闭合的,称为非循环式电子传递。
H2O
Cytb6f
环式光合电子传递
⑸假环式光合电子传递 (pseudocyclic electron transport)
H2O光解所产生的电子不是被NADP+接受,而 是传递给分子态氧(O2),形成超氧阴离子自由基 (O-·2)。
PSⅡ
O-·2 O2
八年级生物上册第四章知识点

八年级生物上册第四章知识点第四章绿色植物的呼吸作用1、光合作用的概念:绿色植物通过叶绿体,利用光能,把二氧化碳和水转化成储存着能量的有机物,并且释放出氧气的过程。
2、光合作用的意义:绿色植物通过光合作用制造的有机物,不仅满足了自身生长、发育、繁殖等生命活动的需要,而且为生物圈的其他生物提供了基本的食物来源。
3、光合作用的主要器官:叶。
4、光合作用的场所:叶绿体。
5、光合作用的物质变化:无机物→有机物。
6、光合作用的能量变化:光能→生物体有机物中的化学能。
7、光反应阶段和暗反应阶段的关系:光反应阶段和暗反应阶段属于光合作用的两个阶段,它们既相互独立,又相互,没有光便没光合作用,有光不一定有光合作用。
8、光合作用的应用:可提高农作物的产量,改善环境等。
9、光合作用和呼吸作用的关系:植物的绿色部分含有叶绿体,叶绿体的内膜相当于半透膜,而细胞内的线粒体和核糖体等细胞器产生的二氧化碳和水蒸气可以通过细胞膜和叶绿体膜散失到细胞外。
当光照强度比较弱时,光合作用产生的氧气量小于呼吸作用产生的二氧化碳量,而当光照强度比较强时,光合作用产生的氧气量大于呼吸作用产生的二氧化碳量。
当光合作用强度大于呼吸作用强度时,植物才真正制造出氧气。
10、绿色植物维持大气中二氧化碳和氧的平衡的意义:绿色植物通过光合作用维持大气中二氧化碳和氧的平衡,不仅为生物圈中的其他生物提供了基本的食物来源,而且对维持整个生物圈的稳态具有重要意义。
生物圈为生物的生存提供了基本条件:营养物质、阳光、空气和水,还有适宜的温度和一定的生存空间。
装片的制作方法(洋葱鳞片叶内表皮细胞装片、人体口腔上皮细胞装片)。
系统及组成(消化系统、呼吸系统、循环系统、神经系统、运动系统)。
花、果实、种子是生殖器官,是植物体的第二器官。
生殖过程:产生生殖细胞、受精、胚胎发育、分娩、婴幼儿的产生和发展(包括青春期)。
植物的生殖(有性生殖和无性生殖)及特点。
生物是一种生命现象,一切有生命的物质都具备这样的特征。
植物生理学-第四章ppt课件

第二节 叶绿体与光合色素
一、叶 绿 体
二、光合色素
1 分类
叶绿素类 (chlorophyll)
类胡萝卜素类 (carotenoid)
叶绿素类a
(蓝绿色)
叶绿素类b
(黄绿色)
磷 光
~ 31千卡
叶绿素分子受光激发时电子能量水平图解
叶绿素的生物合成
合成前体: ð- 氨基酮戊酸
合成途径:
合成条件:
光照 温度 矿质元素
光合作用的机理
原初反应
光
反 应 电子传递和
光合磷酸化
光能的吸收、传递与转换
(光能转换成电能)
基粒片层上
(电能 活跃的化学能)
暗 反 碳素同化 应
(活跃的化学能
H2O的光解和O2的释放,但不能形 成NADPH。(NADP+不足)
光合磷酸化机理
化学渗透学说(P. Mitchell 1961)
第四节 二氧化碳的固定与还原
• C3 途径(还原的戊糖途径、卡尔文循环
The Calvin cycle):C3植物
• C4 途径(C4 pathway)(四碳双羧酸途径):
电子传递和光合磷酸化(photophosphorylation) (电能转换成活跃的化学能)
两个光系统
光合链(“Z”链)
光系统 I : 光系统 II :
证明:“红降”现象 双光增益效应(爱默生效应Emerson effect)
光合电子传递链(“Z”链)
光合磷酸化
在光下叶绿体把光合电子传递与磷
photophosphorylation 酸化作用相偶联,使ADP与Pi形
植物生理学单元自测题

植物生理学单元自测题第四章植物的光合作用一、填空题1.光合作用本质上是一个氧化还原过程。
其中是氧化剂,是还原剂,作为还原的氢的供体。
2.类囊体膜上主要含有四类蛋白复合体,即、、、和。
由于光合作用的光反应是在类囊体膜上进行的,所以称类囊体膜为膜。
3.反应中心色素分子是一种特殊性质的分子,其余的叶绿素分子和辅助色素分子一起称为色素。
4.叶绿体是由被膜、、和三部分组成。
叶绿体中起吸收并转变光能的部位是膜,而固定和同化CO2的部位是。
5.叶绿素对光最强的吸收区有两处:波长640~660nm的光部分和430~450nm的光部分;类胡萝卜素的吸收带则在400~500nm的光区。
6.光合作用可分为:反应、传递和磷酸化、以及等三个阶段。
7.原初反应包括光能的、和反应。
8.光合电子传递链呈侧写的形,电子的最终供体是,电子最终受体是。
9.根据电子传递到Fd后去向,将光合电子传递分为式电子传递、式电子传递和式电子传递三种类型。
10.C3途径是在叶绿体的中进行的。
全过程分为、和三个阶段。
11.核酮糖-1,5-二磷酸羧化酶/加氧酶简称,它既能使RuBP与CO2起,推动C3碳循环,又能使RuBP进行反应而引起光呼吸。
12.C3途径形成的磷酸丙糖可运出叶绿体,在中合成蔗糖或参与其它反应;形成的磷酸己糖则可在中转化成淀粉而被临时贮藏。
13.光呼吸生化途径要经过体、体和体三种细胞器。
光呼吸的底物是。
14.C4植物的光合细胞有细胞和细胞两类。
15.C4植物是在同一和不同的完成CO2固定和还原两个过程;而CAM植物则是在不同和同一完成上述两个过程的。
二、选择题1.光合作用的原初反应是指光能转变成的过程。
A.电能 B.化学能 C.同化力 D.碳水化合物2.夜间,CAM植物细胞的液泡内积累大量的。
A.氨基酸 B.糖类 C.有机酸 D.CO23.与能量转换密切有关的细胞器是。
A.高尔基体与中心体 B.中心体与叶绿体 C.内质网和线粒体 D.线粒体和叶绿体4.光合碳同化的途径有。
北京农学院,植物生理生化,期末题,考研题 光合作用习题

《植物生理学习题集》第四章植物的光合作用Ⅰ 教学大纲基本要求和知识要点一、教学大纲基本要求了解光合作用的概念、意义、研究历史、光合作用总反应式;了解叶绿体的结构、光合色素的种类;了解光合作用过程以及能量吸收转变的情况;了解光合碳同化的基本生化途径以及不同碳同化类型植物的特性;理解光呼吸的含义、基本生化途径和可能的生理意义;了解光合作用的测定方法;了解影响光合作用的内部和外部因素;理解光合作用与作物产量的关系;掌握提高光能利用率的途径与措施。
二、知识要点光合作用是光合生物利用光能同化 CO 2 生成有机物的过程。
CO 2 + 2 H 2 18 O → (CH 2 O) + 18 O 2 + H 2 O植物的光合作用能氧化水而释放氧气,它在光能转化、有机物制造和环境保护等方面都有巨大的作用。
叶绿体是进行光合作用的细胞器。
叶绿体中的类囊体是光反应的场所,在类囊体膜上存在 PS Ⅰ、PS Ⅱ、 Cytb 6 /f 、 ATP 酶四类蛋白复合体 ( 图 5.1) 。
叶绿体中的基质是暗反应的场所,内含同化 CO 2 的全部酶类。
高等植物中的光合色素有两类: (1) 叶绿素,主要是叶绿素 a 和叶绿素 b ; (2) 类胡萝卜素,其中有胡萝卜素和叶黄素。
叶绿素生物合成的起始物是δ- 氨基酮戊酸,该合成过程要有光照,并受温度和矿质元素等的影响。
根据能量转变的性质可将光合作用分为三个过程: (1) 光能的吸收、传递和转换,由原初反应完成; (2) 电能转变为活跃的化学能,由电子传递和光合磷酸化完成; (3) 活跃化学能转变为稳定的化学能,由碳同化完成 ( 表 5.1) 。
表 5.1 光合作用能量转变的过程和反应部位图 5.1 光合膜上的电子与质子传递LHC Ⅱ等受光激发后将接受的光能传到PS Ⅱ反应中心,并在那里发生光化学反应,同时将激发出的 e - 传给质体醌 (PQ) ,从而推动了PS Ⅱ的最初电子传递。
PS Ⅱ反应中心失去 e - 后,变成一个强的氧化剂,它向位于膜内侧的放氧复合体争夺电子而引起水的分解,并将产生的氧气和 H + 释放在内腔。
第四章植物的物质和能量代谢104

structure:
吡咯环
The functions of chlorophyll?
卟啉环
叶绿素醇
甲烯基 副环
叶绿素a C55H72O5N4Mg
叶绿素b C55H70O6N4Mg
1.2.1.2 类胡萝卜素carotenoid
types:叶黄素:黄色 胡萝卜素:橙黄色
两者比例 2:1
6CO2+18ATP+12NADPH F-6- P+18ADP+12NADP++17pi
1.3.3.2 C4途径pathway 60年代中期,发现一些原产热带的植物
如最甘早蔗所、测玉到米14C和不高是梁在等三,碳它酸们中在,饲而喂是14在CO四2后碳, 酸,如OAA、Mal和Asp中,进一步研究说明 这些植物除有和一般植物相同的C3循环途径外, 还 有 一 个 和 光 合 循 环 相 联 系 的 CO2 固 定 途 径 C4——二羧酸途径,该途径是Hatch和Stack 发现的,故也称Hatch-Stack途径,有这种途 径的植物就称为C4植物;而仅有C3途径的植 物称为C3植物
1.3.2.1 光系统 light system
当用长于685nm(远红光)的单色光照射植物 时虽然仍被叶绿素大量吸收,但光合效率明显下 降,这种现象称为“红降”(red drop)现象,后 来又发现用远红光照射植物的同时,如补充以红 光(650nm),则其光合效率比用两种波长的光分 别照射时的总和要大,这种现象称为双光增益效 应或爱默生效应(Emerson effect)
光 叶绿体
2AH2+O2
反离应体—叶—绿希体尔在反光应下(H所ill进re行ac水tio分n)解,并放出O2的
植物生理学--光合作用 ppt课件

淀粉
6CO2+11H2O+18ATP+12NADPH+12H+
PP磷T课酸件 己糖+18ADP+17Pi+115 2NADP+
卡尔文循环的调节
C3途径中的酶 RuBP羧化酶 NADP-GAP脱氢酶
FBP酯酶
SBP酯酶
Ru5P激酶
都属于光调节酶
这些酶在光下活化,以满足光合;而在暗中钝 化,减少底物消耗,使C3循环得以自动调节。
RuBP羧化酶的活性与叶绿体间质中的pH值和 Mg2+含量有密切关系。
PPT课件
16
(二)C4 途径
M.D.Hatch和C.R.Slack(1966)研究证实,在一
光合效率高的植物中,其光合固定CO2 后的第一 个稳定性产物是C4 -二羧酸,由此发现了另一条 CO2 的同化途径——C4 途径,也称C4 -二羧酸途 径或Hatch-Slack循环。
第四章 光合作用
第3节 光合作用机理
二、电子传递与光合磷酸化
(一)光合电子传递 Z 形光合链
①两个光系统串联,最终电子供体是H2O,最终 电子受体是NADP+。
②两个光系统间有一系列的电子载体。 ③传递过程偶联着磷酸化作用,形成ATP。
④各电子载体是以氧化还原电位高低成Z形串联
排列,两处是“上坡”其余“下坡”。
• “不怕太阳晒,也不怕那风雨狂,只怕先生骂我 笨,没有学问无颜见爹娘 ……”
• “太阳当空照,花儿对我笑,小鸟说早早早……”
(二)光合磷酸化
( 1 )非环式光合 磷酸化
( 2 )环式光合磷 酸化
PPT课件
机理
P.Mitchell (1961)提出了 化学渗透学说
《植物生理学》第四章植物的光合作用复习题及答案

《植物生理学》第四章植物的光合作用复习题及答案一、名词解释1.光合作用(photosynthesis):通常是指绿色植物吸收光能,把二氧化碳和水合成有机物,同时释放氧气的过程。
从广义上讲,光合作用是光养生物利用光能把二氧化碳合成有机物的过程。
2. 光合午休现象:指植物的光合速率在中午前后下降的现象。
引起光合"午休"的主要因素是大气干旱和土壤干旱。
另外,中午及午后的强光、高温、低CO2浓度等条件也会使光合速率在中午或午后降低。
3.希尔反应(Hill reaction):希尔(Robert.Hill)发现在分离的叶绿体(实际是被膜破裂的叶绿体)悬浮液中加入适当的电子受体(如草酸铁),照光时可使水分解而释放氧气,这个反应称为希尔反应(Hill reaction) 。
其中的电子受体被称为希尔氧化剂(Hill oxidant)。
4. 荧光(fluorescence)和磷光(phosphorescence)现象:激发态的叶绿素分子回到基态时,可以光子形式释放能量。
处在第一单线态的叶绿素分子回至基态时所发出的光称为荧光,而处在三线态的叶绿素分子回至基态时所发出的光称为磷光。
5. 天线色素(antenna pigment):又称聚(集)光色素(light harvesting pigment),指在光合作用中起吸收和传递光能作用的色素分子,它们本身没有光化学活性。
6. 光合色素:指参与光合作用中光能的吸收、传递和原初反应的各种色素。
包括叶绿素、类胡萝卜素、藻胆素。
可分为聚光色素与作用中心色素两类。
7. 光合作用中心:指在叶绿素或载色体中,进行光合作用原初反应的最基本色素蛋白结构,至少包括一个光能转换色素分子、一个原初电子受体和一个原初电子供体。
8. 光合单位(photosynthetic unit):最初是指释放1个O2分子所需要的叶绿素数目,测定值为2500chl/O 2。
若以吸收1个光量子计算,光合单位为300个叶绿素分子;若以传递1个电子计算,光合单位为600个叶绿素分子。
第四章植物的光合作用知识要点

第四章植物的光合作用一、教学大纲基本要求了解光合作用的概念、意义、研究历史、光合作用总反应式;了解叶绿体的结构、光合色素的种类;了解光合作用过程以及能量吸收转变的情况;了解光合碳同化的基本生化途径以及不同碳同化类型植物的特性;理解光呼吸的含义、基本生化途径和可能的生理意义;了解光合作用的测定方法;了解影响光合作用的内部和外部因素;理解光合作用与作物产量的关系;掌握提高光能利用率的途径与措施。
二、本章知识要点(一)名词解释1.光合作用(photosynthesis) 常指绿色植物吸收光能,把二氧化碳和水合成有机物,同时释放氧气的过程。
从广义上讲,光合作用是光养生物利用光能把二氧化碳合成有机物的过程。
2.碳素同化作用(carbon assimilation) 自养植物吸收二氧化碳,将其转变成有机物质的过程。
植物的碳素同化作用包括细菌光合作用、绿色植物的光合作用和化能合成作用三种类型。
3.光合细菌(photobacteria) :能进行光合作用的一类原核生物。
可分为两类:一些仅有光系统Ⅰ,是不释放分子氧的种类,属于无氧光细菌亚纲,包括红螺菌目和绿菌目;另一些有光系统Ⅰ和光系统Ⅱ,是释放分子氧的种类,属于生氧光细菌亚纲,包括蓝细菌目(又称蓝绿藻)和原绿菌目。
4.希尔反应(Hill reaction) 离体叶绿体在有适当的电子受体存在时,光下分解水并放出氧气的反应。
希尔(Robert.Hill,1939)发现在分离的叶绿体(实际是被膜破裂的叶绿体)悬浮液中加入适当的电子受体(如草酸铁),照光时可使水分解而释放氧气,(同时高铁盐被还原成低铁盐),这个反应被称为希尔反应。
其中的电子受体被称为希尔氧化剂。
5.光反应(light reaction) 光合作用中需要光的反应。
为发生在类囊体上的光的吸收、传递与转换、电子传递和光合磷酸化等反应的总称。
6.暗反应(dark reaction) 光合作用中的酶促反应,即发生在叶绿体间质中的同化二氧化碳生成碳水化合物等有机物的反应。
植物生理学-光合作用完整版本

一个羧基在副环(E)上以酯键与甲醇结合—甲基酯化; 另一个羧基(丙酸)在D环上与植醇(叶绿醇)结合— 植醇基酯化; 非极性,亲脂,插入类囊体膜的疏水区,起定位作用。
叶绿素提取:
纯的有机溶剂不能打破叶绿体色素与蛋白质的联系,所以 必须用能与水混溶的有机溶剂并有少量水存在时,才能将 叶绿体色素提取出来。
人类面临 五大问题
人口 粮食 能源 资源 环境
依赖 光合生产
因此,深入探讨光合作用的规律,揭示光合作用的机理, 使之更好地为人类服务,愈加显得重要和迫切。
第2节 能量转换细胞器 —— 叶绿体
叶片是光合作用的主要器官, 叶绿体(chloroplast)是光合作用最重要的细胞器。
叶绿体的基本结构:
绿 490~ 550 230
黄橙红 550~ 585~ 640~ 585 640 700 212 196 181
远红 700~ 740 166
红外 >740
85 低
光合色素分子对光能的吸收及能量的转变示意图
基态:能量的最低状态 激发态:高能、不稳定状态
物质吸收光子→原子中的e重新排列→分子从基态跃迁到激发态 对于Chl分子: Chl + hγ= Chl* Chl*处于不同激发态:吸收红光→第一单线态;吸收蓝光→第二 单线态。第二单线态的能量>第一单线态。
荧光(fluorescence): 第一单线态的叶绿素分子回至基态时所发出的光。
荧光现象: 叶绿素溶液在透射光下呈绿色,而在反射光下呈红色 的现象。
叶绿素的荧光 (反射光下)叶绿素是叶绿酸的酯(叶绿酸是双羧酸,其中一个羧基被甲醇酯化, 另一个被叶绿醇酯化)。 叶绿素可以与碱起皂化反应而生成醇(甲醇和叶绿醇)和叶绿酸的盐, 产生的盐能溶于水中,用此法可将叶绿素与类胡萝卜素分开。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第四章植物的光合作用光合作用(photosynthesis)通常是指绿色植物吸收光能,把二氧化碳和水合成有机物,同时释放氧气的过程。
地球上一年中通过光合作用约吸收2.0×1011t碳素(6400t/s),合成5×1011t有机物,同时将3.2×1021J的日光能转化为化学能,并释放出5.35×1011t氧气。
光合作用是地球上规模最巨大的把太阳能转变为可贮存的化学能的过程,也是规模最巨大的将无机物合成有机物和从水中释放氧气的过程。
自从有了光合作用,需氧生物才得以进化和发展。
由于光合作用中氧的释放和积累而逐渐形成了大气表面的臭氧(O3)层,O3能吸收阳光中对生物有害的紫外辐射,使生物可从水中到陆地上生活和繁衍。
光合作用是生物界获得能量、食物以及氧气的根本途径,所以光合作用被称为“地球上最重要的化学反应”。
没有光合作用也就没有繁荣的生物世界。
当今人类社会面临着日趋严峻的食物不足、能源危机、资源匮乏和环境恶化等问题,这些问题的解决无一不与植物的光合作用有着密切的关系。
因此深入探讨光合作用的规律,揭示光合作用的机理,使之更好地为人类服务,愈加显得重要和迫切。
第一节光合作用研究的历史一、光合作用总反应式的确定18世纪以前,人们都认为植物是从土壤中获得生长所需的全部元素的。
1771年英国化学家普利斯特利(J.Priestley)发现将薄荷枝条和燃烧的蜡烛放在一个密闭的钟罩里,蜡烛不易熄灭;将小鼠与植物放在同一钟罩里,小鼠也不易窒息死亡。
因此,他提出植物可以“净化”空气,现在就把1771年定为发现光合作用的年代。
以后又经许多人的研究(见绪论),到了19世纪末,人们写出了如下的光合作用的总反应式:6CO2+6H2O→ C6H12O6+6O2 (4-1)从(4-1)式中可以看出:光合作用本质上是一个氧化还原过程。
其中CO2是氧化剂,CO2中的碳是氧化态的,而C6H12O6中的碳是相对还原态的,CO2被还原到糖的水平。
H2O 是还原剂,作为CO2还原的氢的供体。
(4-1)式用了几十年,后来又把它简化成下式:CO2+H2O→(CH2O)+O2(△G°†=4.78³105J) (4-2)(4-2)式用(CH2O)表示一个糖类分子的基本单位,比较简洁。
用叶绿体代替绿色植物,说明叶绿体是进行光合作用的场所。
由于葡萄糖燃烧时释放2870 kJ²mol-1的能量,因而每固定1mol CO2(即12g碳)就意味着转化和贮存了约478kJ的能量。
应该注意到光合作用反应式中所有的反应物和产物都含有氧,而上面两式并没有指出释放的O2是来自CO2还是H2O。
很多年来,人们一直以为光能将CO2分解成O2和C,C与H2O 结合成(C H2O ),然而以下三方面研究证实了光合作用释放的O2来自于H2O 。
1.细菌光合作用能进行光合作用的细菌称之为光合细菌(photosynthetic bacteria)。
光合细菌包括蓝细菌、紫细菌和绿细菌等。
其中蓝细菌的光合过程与真核生物相似,紫细菌和绿细菌则不能分解水而需利用有机物或还原的硫化物等作为还原剂。
例如:紫色硫细菌(purple-sulfur bacteria)和绿色硫细菌(green-sulfur bacteria)利用H2S为氢供体,在光下同化CO2:CO2+2H2S→(CH2O)+2S+H2O (4-3)光合细菌在光下同化CO2而没有O2的释放。
因此,细菌光合作用是指光合细菌利用光能,以某些无机物或有机物作供氢体,将CO2还原成有机物的过程。
1931年微生物学家尼尔(C.B.Van Niel)将细菌光合作用与绿色植物的光合作用加以比较,提出了以下光合作用的通式:CO2+2H2A→(CH2O)+2A+H2O (4-5)这里的H2A代表一种还原剂,可以是H2S、有机酸等,对绿色植物而言,H2A就是H2O,2A就是O2。
绿色植物光合作用中的最初光化学反应是把水分解成氧化剂(OH)与还原剂(H)。
还原剂(H)可以把CO2还原成有机物质;氧化剂(OH)则会通过放出O2而重新形成H2O。
绿色植物和光合细菌都能利用光能将CO2合成有机物,它们是光养生物。
从广义上讲,所谓光合作用,是指光养生物利用光能把CO2合成有机物的过程。
2.希尔反应 1939年英国剑桥大学的希尔(Robert.Hill)发现在分离的叶绿体(实际是被膜破裂的叶绿体)悬浮液中加入适当的电子受体(如草酸铁),照光时可使水分解而释放氧气:4Fe3++2H2O→4Fe2++4H++O2 (4-6)这个反应称为希尔反应(Hill reaction)。
其中的电子受体被称为希尔氧化剂(Hill oxidant),铁氰化钾、草酸铁、多种醌、醛及有机染料都可作为希尔氧化剂。
希尔不但证明了给叶绿体照光可使水分解放氧,氧的释放与CO2还原是两个不同的过程,而且也是第一个用离体的叶绿体做试验,把对光合作用的研究深入到细胞器水平,为光合作用研究开创了新的途径。
以后发现生物中重要的氢载体NADP+也可以作为生理性的希尔氧化剂,从而使得希尔反应的生理意义得到了进一步肯定。
在完整的叶绿体中NADP+作为从H2O到CO2的中间电子载体,其反应式可写为:2NADP++2H2O→2NADPH+2H++O2 (4-7 )CO2也可看作为一种生理性的希尔氧化剂,因为向完整的叶绿体悬浮液中充入CO2或加入能产生CO2的试剂如NaHCO3,照光时叶绿体能发生放氧反应。
3.18O的研究更为直接的证据是标记同位素的实验。
1940年美国科学家鲁宾(S.Ruben)和卡门(M.D.Kamen)等用氧的稳定同位素18O标记H2O或CO2进行光合作用的实验,发现当标记物为H218O时,释放的是18O2,而标记物为C18O2时,在短期内释放的则是O2。
这清楚地指出光合作用中释放的O2来自于 H2O。
CO2+2 H218O→(CH2O)+ 18O2+H2O(4-8)为了把CO2中的氧和H2O中的氧在形式上加以区别,表明光合作用中释放的O2全来自于H2O,而CO2中的一个O又被还原成H2O,因此,可用下式作为光合作用的总反应式。
CO2+2H2O →(C H2O )+ O2+2 H2O (4-9)二、光反应和暗反应光合作用需要光,然而是否其中每一步反应过程都需要有光呢?20世纪初英国的布莱克曼(Blackman)、德国的瓦伯格(O.Warburg)等人在研究光强、温度和CO2浓度对光合作用影响时发现,在弱光下增加光强能提高光合速率,但当光强增加到一定值时,再增加光强则不再提高光合速率。
这时要提高温度或CO2浓度才能提高光合速率。
据测定,在10~30℃的范围内,如果光强和CO2浓度都适宜的话,光合作用的Q10=2~2.5(Q10为温度系数,即温度每增加10℃,反应速度增加的倍数)。
按照光化学原理,光化学反应是不受温度影响的,或者说它的Q10接近1;而一般的化学反应则和温度有密切关系,Q10为2~3,这说明光合过程中有化学反应的存在。
用藻类进行闪光试验,在光能量相同的前提下,一种用连续照光,另一种用闪光照射,中间隔一定暗期,发现后者光合效率是连续光下的200%~400%。
这些实验表明了光合作用可以分为需光的光反应(light reaction)和不需光的暗反应(dark reaction)两个阶段。
1954年美国科学家阿农(D.I.Arnon)等在给叶绿体照光时发现,当向体系中供给无机磷、ADP和NADP时,体系中就会有ATP和NADPH 产生。
同时发现,只要供给了ATP和NADPH+,即使在黑暗中,叶绿体也可将CO2转变为糖。
由于ATP和NADPH是光能转化的产物,具有在黑暗中同化CO2为有机物的能力,所以被称为“同化力”(assimilatory power)。
可见,光反应的实质在于产生“同化力”去推动暗反应的进行,而暗反应的实质在于利用“同化力”将无机碳(CO2)转化为有机碳(CH2O)。
图 4-1 光合作用中“光”反应与“暗”反应的主要产物光合作用中光反应和碳同化(暗反应)分别在叶绿体的不同区域内。
光反应所需要的ATP和NADPH底物合成的一系列反应发生在叶绿体类囊体膜上。
光反应产物在碳同化反应中一系列的基质酶的作用下固定CO2转化为碳水化合物。
当然,进一步研究发现光、暗反应对光的需求不是绝对的。
即在光反应中有不需光的过程(如电子传递与光合磷酸化),在暗反应中也有需要光调节的酶促反应。
现在认为,“光”反应不仅产生“同化力”,而且产生调节“暗”反应中酶活性的调节剂(图4-1),如还原性的铁氧还蛋白。
三、光合单位释放一个氧分子需要吸收几个光量子?需要多少个叶绿素分子参与?在研究这些问题的过程中,提出了“光合单位”的概念。
在研究光能转化效率时,需要知道光合作用中吸收一个光量子所能引起的光合产物量的变化(如放出的氧分子数或固定CO2的分子数),即量子产额(quantum yield)或叫量子效率(quantum efficiency)。
量子产额的倒数称为量子需要量(quantum requirement)即释放1分子氧和还原1分子二氧化碳所需吸收的光亮子数。
1922年,瓦伯格等计算出最低量子需要量为4,而他的学生爱默生(R.Emersen)等则测定出最低量子需要量为8。
后来的实验证据都支持了爱默生的观点,于是8的最低量子需要量得到了普遍的承认,这个数值相当于0.125的量子效率。
根据光化学定律(一个分子吸收一个量子,发生一次光化学变化),如果植物的每个叶绿素分子都能进行光化学反应,按还原1个CO2和释放1个O2需吸收8个光量子算,则每当有8个叶绿素分子在一起时,一次足够强的闪光就会造成1个O2的释放。
但在1932年,爱默生及阿诺德(W.Arnold)对小球藻(chlorella)悬浮液做闪光试验,计算每次闪光的最高产量是约2 500个叶绿素分子产生1个O2分子,似乎在光合组织中是以2 500个叶绿素分子组成1个集合体进行放氧的,于是当时就把释放1分子氧或同化1分子CO2所需的2 500个叶绿素的分子数目称作1个“光合单位”(photosynthetic unit)。
以后又认为,光合是以吸收光量子开始的,应以量子基础计算“光合单位”,1个光合单位应是300(2 500÷8≈300)个叶绿素分子。
为什么要300个叶绿素分子吸收1个光子?其解释是:闪光可能被几百个叶绿素分子吸收,可是激发能需传递到1个能够产生光化学反应的“反应中心”(reaction center)区域才能有效。
这个反应中心的反应中心色素分子(reaction center pigment)是一种特殊性质的叶绿素a分子,它不仅能捕获光能,还具有光化学活性,能将光能转换成电能。