生物化学(第三版)第十章 酶的作用机制和酶的调节课后习题详细解答_ 复习重点
生物化学第三版课后章习题详细解答

生物化学(第三版)课后--章习题详细解答————————————————————————————————作者:————————————————————————————————日期:生物化学(第三版)课后3 -13章习题详细解答第三章氨基酸习题1.写出下列氨基酸的单字母和三字母的缩写符号:精氨酸、天冬氨酸、谷氨酰氨、谷氨酸、苯丙氨酸、色氨酸和酪氨酸。
[见表3-1]表3-1 氨基酸的简写符号名称三字母符号单字母符号名称三字母符号单字母符号丙氨酸(alanine) Ala A 亮氨酸(leucine) Leu L 精氨酸(arginine) ArgR 赖氨酸(lysine)LysK天冬酰氨(asparagines) Asn N 甲硫氨酸(蛋氨酸)(methionine)Met M天冬氨酸(aspartic acid) Asp D苯丙氨酸(phenylalanine)Phe F Asn和/或Asp Asx B半胱氨酸(cysteine) CysC脯氨酸(praline)ProP 谷氨酰氨(glutamine) Gln Q 丝氨酸(serine) Ser S 谷氨酸(glutamic acid)Glu E苏氨酸(threonine) Thr TGln和/或Glu Gls Z甘氨酸(glycine) Gly G 色氨酸(tryptophan) TrpW组氨酸(histidine) HisH 酪氨酸(tyrosine) Tyr Y异亮氨酸(isoleucine)Ile I 缬氨酸(valine) Val V2、计算赖氨酸的εα-NH3+20%被解离时的溶液PH。
[9.9]解:pH=pKa +lg20% pKa= 10.53(见表3-3,P133)pH=10.53+lg20%=9.833、计算谷氨酸的γ-COOH三分之二被解离时的溶液pH。
[4.6]解:pH =pKa+ lg2/3% pKa =4.25pH = 4.25 + 0.176 = 4.4264、计算下列物质0.3mol/L溶液的pH:(a)亮氨酸盐酸盐;(b)亮氨酸钠盐;(c)等电亮氨酸。
大学生物化学(第三版)课后习题解答

第一章糖类1.环状己醛糖有多少个可能的旋光异构体,为什么?[25=32]解:考虑到C1、C2、C3、C4、C5各有两种构象,故总的旋光异构体为25=32个。
2.含D-吡喃半乳糖和D-吡喃葡萄糖的双糖可能有多少个异构体(不包括异头物)?含同样残基的糖蛋白上的二糖链将有多少个异构体?[20;32]解:一个单糖的C1可以与另一单糖的C1、C2、C3、C4、C6形成糖苷键,于是α-D-吡喃半乳基-D-吡喃葡萄糖苷、β-D-吡喃半乳基-D-吡喃葡萄糖苷、α-D-吡喃葡萄糖基-D-吡喃半乳糖苷、β-D-吡喃葡萄糖基-D-吡喃半乳糖苷各有5种,共5×4=20个异构体。
糖蛋白上的二糖链其中一个单糖的C1用于连接多肽,C2、C3、C4、C6用于和另一单糖的C1形成糖苷键,算法同上,共有4×4=16个,考虑到二糖与多肽相连时的异头构象,异构体数目为16×2=32个。
3.写出β-D-脱氧核糖、α-D-半乳糖、β- L-山梨糖和β-D-N-乙酰神经氨酸(唾液酸)的Fischer投影式,Haworth式和构象式。
4.写出下面所示的(A).(B)两个单糖的正规名称(D/L,α/β,f/p),指出(C).(D)两个结构用RS系统表示的构型(R/S)[A、β- D-f-Fru;B、α-L- p-Glc; C、R; D、S]5. L7-葡萄糖的α和β异头物的比旋[αD20]分别为+112.2°和+18.70°。
当α-D-吡喃葡糖晶体样品溶于水时,比旋将由+112.2°降至平衡值+52.70°。
计算平衡混合液中α和β异头物的比率。
假设开链形式和呋喃形式可忽略。
[α异头物的比率为36.5%,β异头物为63.5%]解:设α异头物的比率为x,则有112.2x+18.7(1-x)=52.7,解得x=36.5%,于是(1-x)= 63.5%。
6.将500 mg糖原样品用放射性氰化钾(K14CN)处理,被结合的14CN—正好是0.193μmol,另一500 mg同一糖原样品,用含3% HCl的无水甲醇处理,使之形成还原末端的甲基葡糖苷。
生物化学(第三版)课后习题详细解答

生物化学(第三版)课后习题详细解答第三章氨基酸提要α-氨基酸是蛋白质的构件分子,当用酸、碱或蛋白酶水解蛋白质时可获得它们。
蛋白质中的氨基酸都是L型的。
但碱水解得到的氨基酸是D型和L型的消旋混合物。
参与蛋白质组成的基本氨基酸只有20种。
此外还有若干种氨基酸在某些蛋白质中存在,但它们都是在蛋白质生物合成后由相应是基本氨基酸(残基)经化学修饰而成。
除参与蛋白质组成的氨基酸外,还有很多种其他氨基酸存在与各种组织和细胞中,有的是β-、γ-或δ-氨基酸,有些是D型氨基酸。
氨基酸是两性电解质。
当pH接近1时,氨基酸的可解离基团全部质子化,当pH在13左右时,则全部去质子化。
在这中间的某一pH(因不同氨基酸而异),氨基酸以等电的兼性离子(H3N+CHRCOO-)状态存在。
某一氨基酸处于净电荷为零的兼性离子状态时的介质pH称为该氨基酸的等电点,用pI表示。
所有的α-氨基酸都能与茚三酮发生颜色反应。
α-NH2与2,4-二硝基氟苯(DNFB)作用产生相应的DNP-氨基酸(Sanger反应);α-NH2与苯乙硫氰酸酯(PITC)作用形成相应氨基酸的苯胺基硫甲酰衍生物(Edman反应)。
胱氨酸中的二硫键可用氧化剂(如过甲酸)或还原剂(如巯基乙醇)断裂。
半胱氨酸的SH基在空气中氧化则成二硫键。
这几个反应在氨基酸荷蛋白质化学中占有重要地位。
除甘氨酸外α-氨基酸的α-碳是一个手性碳原子,因此α-氨基酸具有光学活性。
比旋是α-氨基酸的物理常数之一,它是鉴别各种氨基酸的一种根据。
参与蛋白质组成的氨基酸中色氨酸、酪氨酸和苯丙氨酸在紫外区有光吸收,这是紫外吸收法定量蛋白质的依据。
核磁共振(NMR)波谱技术在氨基酸和蛋白质的化学表征方面起重要作用。
氨基酸分析分离方法主要是基于氨基酸的酸碱性质和极性大小。
常用方法有离子交换柱层析、高效液相层析(HPLC)等。
习题1.写出下列氨基酸的单字母和三字母的缩写符号:精氨酸、天冬氨酸、谷氨酰氨、谷氨酸、苯丙氨酸、色氨酸和酪氨酸。
生物化学第三版习题答案

生物化学第三版习题答案生物化学是生命科学中的一个重要分支,它研究生物体内化学过程和物质的变化。
第三版的生物化学教材通常会包含许多习题,帮助学生巩固和深化对知识点的理解。
以下是一些习题及其答案的示例,供参考:习题1:酶的催化作用问题:简述酶的催化作用原理。
答案:酶是生物体内催化化学反应的蛋白质,它们能够显著降低反应的活化能,从而加速反应速率。
酶的催化作用原理主要基于其活性部位与底物的特异性结合,形成酶-底物复合物。
这种结合使得底物分子在酶的活性部位发生结构变形,更易于反应。
反应完成后,产物从酶上分离,酶恢复其原始状态,可以继续催化下一个底物分子。
习题2:DNA的复制问题:描述DNA复制的基本过程。
答案:DNA复制是一个半保留的过程,包括以下步骤:首先,DNA双链被解旋酶解旋成两条单链;接着,DNA聚合酶识别复制起点,并在引物RNA的帮助下开始合成新的互补链;随后,新的链沿着模板链延伸,形成新的DNA双螺旋。
最终,每个原始链都与新合成的链配对,形成两个完整的DNA分子。
习题3:氨基酸的结构和分类问题:列举氨基酸的几种基本结构特征,并简述其分类。
答案:氨基酸是蛋白质的基本组成单元,具有以下基本结构特征:一个α-羧基、一个α-氨基、一个氢原子和一个侧链(R基)。
根据侧链的化学性质,氨基酸可以分为20种标准氨基酸,主要分为四类:非极性疏水氨基酸、极性疏水氨基酸、酸性氨基酸和碱性氨基酸。
习题4:细胞呼吸问题:简述有氧呼吸和无氧呼吸的区别。
答案:有氧呼吸和无氧呼吸是细胞产生能量的两种方式。
有氧呼吸需要氧气作为最终电子受体,通过糖酵解、三羧酸循环(TCA循环)和电子传递链产生大量的ATP。
无氧呼吸,又称为发酵,是在没有氧气的情况下进行的,通过糖酵解产生少量的ATP,同时产生酒精或乳酸作为代谢终产物。
习题5:基因表达调控问题:解释基因表达调控的基本机制。
答案:基因表达调控是指细胞内控制基因转录为mRNA的过程,进而影响蛋白质的合成。
生物化学学习题酶的催化作用和调控机制

生物化学学习题酶的催化作用和调控机制酶是生物体内的一类特殊蛋白质,它在生物化学过程中起着催化和调控作用。
酶的催化作用和调控机制是生物化学学习中的重要内容。
本文将通过解答一些生物化学学习题,来探讨酶的催化作用和调控机制的原理和应用。
1. 什么是酶的催化作用?酶的催化作用是指酶作为催化剂,在生物化学反应中加速反应速率而本身不参与反应的过程。
酶能够降低活化能,使反应更容易发生。
酶与底物结合形成酶-底物复合物,通过调整底物分子的构象,提供合适的反应环境或为反应过程提供必要的功能基团,从而促进和加速生物化学反应。
2. 酶的催化过程中发生了哪些重要事件?酶的催化过程中,发生了以下几个重要事件:(1)底物与酶结合:酶通过底物识别位点与底物结合,形成酶-底物复合物。
(2)底物结构改变:酶可以通过改变底物分子的构象,使之更有利于反应发生。
(3)催化反应:酶通过提供功能基团、调节反应环境等方式催化底物的转化,包括底物的分解、合成、转移等。
(4)生成产物:反应发生后,产生新的物质,酶释放产物,恢复到催化循环中。
3. 酶的活性如何被调控?酶的活性可以通过多种方式被调控,包括:(1)温度:酶的活性随着温度的变化而变化。
适宜的温度范围内,酶的活性增加,但过高或过低的温度会使酶的活性降低甚至失活。
(2)pH值:酶对pH值也非常敏感,适宜的pH值范围内酶的活性最高,但过高或过低的pH值会影响酶的构象和功能基团的离子化状态,从而影响酶的活性。
(3)底物浓度:酶的活性受底物浓度调控。
在底物浓度适宜时,酶的催化作用展现最佳效果,但当底物浓度过高时,酶的活性可能受到抑制。
(4)辅因子:某些酶活性依赖于辅助因子的存在,例如金属离子、辅酶等。
这些辅因子能够与酶结合,形成活性辅因子-酶复合物,从而激活酶的催化作用。
4. 酶在生物体内的调控机制有哪些?酶在生物体内的调控机制有多种,包括:(1)底物浓度反馈抑制:当底物浓度过高时,产物可以通过反馈抑制的方式抑制酶的活性,从而保持底物的代谢平衡。
生化简答题(附答案)

生化简答题(附答案)一、题目本题要求考生对酶的作用机制进行简要阐述,并举例说明酶在生物技术中的应用。
本题考查学生对生物化学知识的掌握,以及将理论知识应用于实际问题的能力。
二、简答题内容1. 酶的作用机制酶是一种生物大分子,主要由蛋白质组成,具有催化化学反应的作用。
酶的作用机制主要包括以下几个方面:(1)降低活化能:酶通过降低化学反应的活化能,使反应速率加快。
活化能是指反应物转变为产物所需的最小能量。
酶与反应物结合形成酶-底物复合物,从而降低反应所需的活化能,加速反应进行。
(2)特异性:酶具有高度的特异性,即一种酶只能催化一种或一类化学反应。
这种特异性是由酶的活性中心与底物的结构互补所决定的。
(3)可逆性:酶催化反应是可逆的,即酶既能催化正反应,也能催化逆反应。
(4)调节性:酶的活性受到多种因素的调控,如酶的浓度、底物浓度、pH、温度等。
这种调节性使酶在生物体内能够适应不同的生理需求。
2. 酶在生物技术中的应用(1)酶在食品工业中的应用:酶在食品工业中具有广泛的应用,如淀粉酶用于淀粉糖浆的生产,蛋白酶用于肉类嫩化,脂肪酶用于油脂改性等。
例题:简述酶在面包制作中的应用。
答案:在面包制作过程中,酶制剂如淀粉酶、蛋白酶和脂肪酶等起到了重要作用。
淀粉酶能分解面粉中的淀粉,产生可发酵的糖类,为酵母提供能量。
蛋白酶能分解面粉中的蛋白质,使面团具有良好的延展性。
脂肪酶能改善面包的口感和风味。
(2)酶在医药领域的应用:酶在医药领域具有广泛的应用,如溶菌酶用于治疗感染性疾病,尿激酶用于治疗血栓性疾病等。
例题:简述酶在药物研发中的应用。
答案:酶在药物研发中具有重要作用。
首先,酶可作为药物靶点,通过抑制或激活酶的活性,达到治疗疾病的目的。
其次,酶可作为生物催化剂,用于合成药物。
此外,酶还可用于药物分析,如通过检测酶活性来判断药物的含量和纯度。
(3)酶在环境保护中的应用:酶在环境保护领域具有重要作用,如降解有机污染物、处理工业废水等。
王镜岩高级生物化学 酶的作用机制和酶的调节
H2N
补:酶的调节部位 Regulatory site
某些酶分子中存在着一些可以与其他分子发生 某种程度的结合的部位,从而引起酶分子空间 构象的变化,对酶起激活或抑制作用。
酶催化作用的本质
酶催化作用的本质是酶的活性中心与底 物分子通过短程非共价力(如氢键,离子 键和疏水键等)的作用,形成E-S反应中 间物。 其结果使底物的价键状态发生形变或极 化,起到激活底物分子和降低过渡态活 化能作用。
肠激酶
-
-胰凝乳蛋白酶活性 最高,但不稳定 -胰凝乳蛋白酶稳定, 活性只有的40%
胃蛋白酶的激活
思考
机体为会么先产生酶原后再经过剪切加 工形成活性酶,为会么不直接产生活性 酶,其生理意义何在?
(三)可逆的共价修饰
1、共价调节酶:可被共价修饰而改变酶活 性。 2、可逆的共价修饰:通过其它酶对其多肽 链上某些基团进行可逆的共价修饰,使酶 处于活性与非活性的互变状态,从而调节 酶活性。 3、例如:蛋白激酶通过蛋白磷酸化和去磷 酸化来失现酶活性与非活性的互变。
非常 保守
胰凝乳蛋白酶上的催化基团
(胰凝乳蛋白酶)
胰凝 乳蛋 白酶 催化 反应 的详 细机 制
三、酶活性的调节控制
(一)别构调控 1、别构调节:酶分子的非催化部位与某些 化合物可逆地非共价结合后发生构象的改 变,进而改变酶活性状态的作用。 2、别构酶:具有上述这种调节作用的酶。 3、效应物:能使酶分子发生别构作用的物 质。可分为正效应物(别构激活剂)和负 效应物(别构抑制剂)。 *ATCase, 3-磷酸甘油醛脱氢酶
习题
* 关于别构酶,正确的表达是 A、它们一般是寡聚酶 B、它们一般是单体酶 C、当效应剂与别构酶非共价结合后,引起别构效应。 D、当效应剂与别构酶共价结合后,引起别构效应。 E、别构酶的动力学性质符合米氏方程式。 F、别构酶的动力学性质不符合米氏方程式。 下列哪一个酶的催化活性需要金属离子? A、溶菌酶 B、羧肽酶 C、胰凝乳蛋白酶 D、胰蛋白酶 胰蛋白酶属单体酶。 别构酶动力学曲线的特点都是呈S形曲线。 酶活性中心是亲水的介电区域。 溶菌酶的实现催化功能需金属离子Mg2+参与。 4、 简述酶活性调控的几种机制。 同工酶有何生理意义?
第10章酶的作用机制和酶的调节

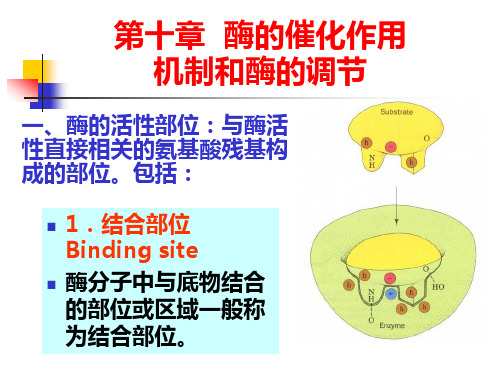
第10章酶的作用机制和酶的调节第10章酶的作用机制和酶的调节教学目的:掌握酶的活性部位结构与功能、酶活性的别构调节、酶原激活,了解酶高效性原因教学重点:酶活性部位的结构与功能及酶的活性的别构调节教学难点:酶活性的别构调节教学方法:多媒体教学内容:一、酶的活性部位及确定方法(一)酶活性部位概念及特点1、酶的活性中心(活性部位):指酶分子中的表面有一个必需基团比较集中、并构成一定空间结构的微小区域。
酶活性中心的基团,按其功能可分为结合基团和催化基团。
活性中心的基团都是维持酶活性的必需基团,2、酶活性部位的共同点:(1)酶活性部位仅占酶体积的很小一部分,通常只占整个酶分子体积的1~2%,酶分子是大分子物质,由很多氨基酸构成,而活性部位仅由几个氨基酸残基组成催化部位一般由2~3个氨基酸残基组成。
结合部位氨基酸残基数目,不同的酶有所不同。
可能是一个,也可能是多个。
(2)酶的活性部位具有三维结构,构成酶活性中心的基团,可位于同一条肽链上,也可位于不同的肽链上,在一级结构上可能相距甚远,但在空间结构上位置必须相互靠近;酶的空间结构受物理或化学因素影响时,酶的活性部位可能会遭破坏,酶会失活。
(3)活性中心的结合基团与底物专一性结合,这需要活性部位的基团精确排列。
活性部位具有一定的柔韧性,活性部位的结构并不是与底物的结构正好互补。
在酶与底物结合过程中,酶活性中心的构象在底物的诱导下可发生形变,然后嵌合互补形成中间产物,而底物在酶活性中心的诱导下也可发生形变,变的易与酶结合,有时是两者的构象同时发生变化后才互补契合(诱导契合学说)。
(4)酶活性部位位于酶分子表面的一个裂缝内,底物分子或底物分子的一部分结合到裂缝中,裂缝内的非极性基团较多,形成一个疏水环境,提高与底物的结合能力,也有极性的氨基酸残基,以便与底物结合并催化底物发生反应。
(5)底物通过较弱的次级键与酶结合。
组成酶活性中心的氨基酸残基,常见的有:组氨酸、赖氨酸、天冬氨酸、谷氨酸、丝氨酸、半胱氨酸和酪氨酸3、研究酶活性部位的方法(1)共价修饰(2)亲和标记法(3)切除法(4)X射线晶体结构分析法二、酶促反应机制(一)基元催化的分子机制:酶的催化作用包括若干基元催化。
生物化学__酶的作用机制和酶的调节

2006-1-7
羧 肽 酶 结 合 底 物 前 后 构 象 变 化
2006-1-7
Glu270
Zn
Tyr248 Arg145
Zn
底物
(三)酸碱催化
酶分子的一些功能基团起瞬时质子供体或质子受体的作用。
氨甲酰磷酸
ATCase
天冬氨酸
+-
ATP
氨甲酰天冬氨酸 UMP UTP
CTP
2006-1-7
E.coli的ATCase的亚基排列
c:催化亚基(catalytic subunit) r:调节亚基( regulative subunit)
ATCase
c3r3)
半 分 子 ( 的 结 构
2006-1-7
2006-1-7
邻近效应与定向效应对反应速度的影响: ①使底物浓度在活性中心附近很高 ②酶对底物分子的电子轨道具有导向作用 ③酶使分子间反应转变成分子内反应 ④邻近效应和定向效应对底物起固定作用
2006-1-7
(二)底物的形变和诱导契合
酶中某些基团可使底物分子的敏感键中某些基团的电子云密度变化,产生电 子张力,降低了底物的活化能。
Glu
级联系统调控糖原 分解示意图
肾上腺素或 胰高血糖素
肾上腺素或 胰高血糖素
(极微量)
(5)酶具有复杂的折叠结构。
1、腺苷酸环化酶
2006-1-7
2006-1-7
接触残基:R1、R2、R6、R8、R9、 R163
辅助残基:R3、R4、R5、R164、R165
结构残基:R10、R162、R169
酶的作用机制和酶的调节

天冬氨酸转氨甲酰酶(ATCase)
• 动力学特征:双底物反应,固定氨甲酰磷酸, 变化[Asp],其s-v图为S形,是别构酶。 效应物:S(同促)、ATP(正协同,异促)、 CTP(负协同,异促)。
(2)3-磷酸甘油醛脱氢酶
• 共四个亚基,Km1和Km2都较小,易与 NAD+结合,即在低底物浓度时反应较 快;而Km3则增大了100倍,很难与NAD +反应。这是由构象变化引起的。在生物 体内,当NAD+不足时可以保证酵解的 进行,而当NAD+过多时则供给其它反 应,避免造成酸中毒。
(1)临近定向效应
在酶促反应中,底物分子结合到酶的活性中心,一方 面底物在酶活性中心的有效浓度大大增加,另一方面, 由于活性中心的立体结构和相关基团的诱导和定向作 用,使底物分子中参与反应的基团相互接近,并被严 格定向定位,使酶促反应具有高效率和专一性特点。
邻近效应 (proximity effect)与定向排列 (orientation
+
OH HN NH
-COO -,
..
-NH 2,
-S ,-
O- :N NH
广义酸基团 (质子供体)
广义碱基团 (质子受体)
(4)共价催化
催化剂通过与底物形成反应活性很高的共价过 渡产物,使反应活化能降低,从而提高反应速 度的过程,称为共价催化。
酶中参与共价催化的基团主要包括 His 的咪唑 基,Cys 的巯基,Asp 的羧基,Ser 的羟基等。
一.酶的活性部位(active site)
• 存在于酶分子表面的具有结合和催化底物形成产物的 空间区域
• 活性中心=结合基团+催化基团 • 必须基团=结合基团+催化基团+活性中心外必须基团
生物化学 第10章 酶作用机制和酶调节-精品文档

3、X射线晶体结构分析法
二、影响酶催化效率的有关因素
ห้องสมุดไป่ตู้
底物和酶的邻近效应与定向效应 底物的形变和诱导契合 酸碱催化 共价催化 金属离子催化 多元催化和协同效应 活性部位微环境的影响
(一)底物和酶的邻近效应与定向效应
邻近效应:在酶促反应中,由于酶和底物分子之间的 亲和性,底物分子向酶的活性中心靠近,最终结合到 酶的活性中心,使底物在酶活性中心的有效浓度大大 增加的效应。 定向效应:当专一性底物向酶活性中心靠近时,会诱 导酶分子构象发生改变,使酶活性中心的相关基团和 底物的反应基团正确定向排列,同时使反应基团之间 的分子轨道以正确方向严格定位,使酶促反应易于进 行。
Chapter10 酶的作用机制和酶的调节
一、酶的活性部位
(一)酶活性部位的特点 在酶蛋白中只有少数特异的氨基酸残基参与底物结合及催化作用, 这些特异的氨基酸残基比较集中的区域,即与酶活力直接相关的区域 称为酶的活性部位或活性中心(通常将酶的结合部位和催化部位总称 为酶的活性部位或活性中心)。
(二)研究酶活性部位(活性中心)的方法
2. 掌握别构酶、同工酶的概念 3. 了解酶工程的含义及酶的应用
ATP
ADP
Thr Ser -OH
蛋白激酶
Thr Ser -O-PO32-
Tyr
蛋白磷酸酶 酶蛋白
Pi H2O
Tyr
酶蛋白
酶的磷酸化与脱磷酸化
四、同工酶
•
同工酶(isoenzyme)是指催化相同的化学反应,
而酶蛋白的分子结构、理化性质乃至免疫学
性质不同的一组酶。
同工酶主要由于基因倍增(duplication)和 趋异(divergence)所致。
第10章_酶的作用机制和酶的调节.

活性部位微环境的影响
酶分子表面有一个裂缝,活性部位就 位于疏水环境的裂缝中。 在非极性环境中的介电常数较在水介 质的介电常数低。 在非极性环境中两个带电基团之间的 静电作用比在极性环境中显著增高
胰凝乳蛋白酶
四、酶催化反应机制的实例
溶菌酶(2)
溶菌酶(3)
溶菌酶催化作用
溶菌酶(4)
丝氨酸蛋白酶(7)
催化三联体 底物(一种多肽)
活性部位 疏水口袋
胰凝乳蛋白酶
酶-底物复合物
胰凝乳蛋白酶反应的详细机制
丝氨酸蛋白酶(8)
②
Ser195-OH的O与肽键的羰基C共价结合,H结合于His57的N上
胰凝乳蛋白酶反应的详细机制
丝氨酸蛋白酶(9)
产物1
③
His提供H给肽键的N形成-NH2,肽键断裂,释放第一个产物
研究酶活性部位的方法(1)
1 酶分子侧链基团的化学修饰法 (1)非特异性共价修饰 (2)特异性共价修饰 (3)亲和标记法 2 动力学参数测定法 3 X射线晶体结构分析法
4 定点诱变法
研究酶活性部位的方法(2)
酶分子侧链基团的化学修饰法
用带标记的化合物与酶的活性部位氨基酸残基侧链基团共价结合,然后 水解该酶,分离出带有标签的肽段,即可分析出活性部位的氨基酸残基。
二、酶催化反应的独特性质
(1)酶反应可以分为两类,一类反应仅仅涉及到电子的转移, 另一类反应涉及到电子和质子两者或其他基团的转移 (2)酶的催化作用是由氨基酸侧链上的功能基团和辅酶为媒介的
(3)酶催化反应的最适pH范围通常狭小 (4)与底物相比较,酶分子很大,而活性部位通常只比底物稍大一些 (5)酶除了具有进行催化反应所必需的活性基团外,还有别的特性
生物化学(第三版)第十章酶的作用机制和酶的调节课后习题详细解答_复习重点

生物化学(第三版)第十章酶的作用机制和酶的调节课后习题详细解答_复习重点第十章酶的作用机制和酶的调节提要酶的活性部位对于不需要辅酶的酶来说,就是指酶分子中在三维结构上比较靠近的几个氨基酸残基负责与底物的结合与催化作用的部位,对于需要辅酶的酶来说,辅酶分子或辅酶分子上的某一部分结构,往往也是酶活性部位的组成部分。
酶活性部位有6个共同特点。
研究酶活性部位的方法有:酶分子侧链基团的化学修饰法,动力学参数测定法,X射线晶体结构分析法和定点诱变法,这些方法可互相配合以判断某个酶的活性部位。
酶是催化效率很高的生物催化剂,这是由酶分子的特殊结构所决定的。
经研究与酶催化效率的有关因素有7个,即底物和酶的邻近效应与定向效应,底物的形变与诱导契合,酸碱催化,共价催化,金属离子催化,多元催化和协同效应,活性部位微环境的影响。
但这些因素不是同时在一个酶中其作用,也不是一种因素在所有的酶中起作用,对于某一种酶来说,可能分别主要受一种或几种因素的影响。
研究酶催化的反应机制,始终是酶学研究的一个重点,通过大量的研究工作,已经对一些酶的作用机制有深入了解,该章对溶解酶、胰核糖核酸酶A、羧肽酶A、丝氨酸蛋白酶、天冬氨酸蛋白酶等的催化作用机制进行了详尽的讨论。
酶活性是受各种因素调节控制的,除了在第8章中已介绍的几种因素外,主要还有①别构调节,例如ATCase。
②酶原的激活,如消化系统蛋白酶原的激活及凝血系统酶原的激活。
③可逆共价修饰调控,如蛋白质的磷酸化,一系列蛋白激酶的作用。
通过以上作用,使酶能在准确的时间和正确的地点表现出它们的活性。
别构酶一般都是寡聚酶,有催化部位和调节部位,别构酶往往催化多酶体系的第一步反应,受反应序列的终产物抑制,终产物与别构酶的调节部位相结合,由此调节多酶体系的反应速率。
别构酶有协同效应,[S]对υ的动力学曲线呈S形曲线(正协同)或表现双曲线(负协同),两者均不符合米氏方程。
ATCase作为别构酶的典型代表,已经测定了其三维结构,详细研究了别构机制和催化作用机制。
生物化学第三版课后习题答案

第一章1. 举例说明化学与生物化学之间的关系。
提示:生物化学是应用化学的理论和方法来研究生命现象,在分子水平上解释和阐明生命现象化学本质的一门学科.化学和生物化学关系密切,相互渗透、相互促进和相互融合。
一方面,生物化学的发展依赖于化学理论和技术的进步,另一方面,生物化学的发展又推动着化学学科的不断进步和创新。
举例:略。
2.试解释生物大分子和小分子化合物之间的相同和不同之处。
提示:生物大分子一般由结构比较简单的小分子,即结构单元分子组合而成,通常具有特定的空间结构。
常见的生物大分子包括蛋白质、核酸、脂类和糖类。
生物大分子与小分子化合物相同之处在丁: 1) 共价键是维系它们结构的最主要的键;2)有一定的立休形象和空间大小; 3)化学和|物理性质主要决定于分子中存在的官能团。
生物大分子与小分子化合物不同之处在于: (1) 生物大分子的分子量要比小分子化合物大得多,分子的粒径大小差异很大; (2) 生物大分子的空间结构婴复杂得多,维系空间结构的力主要是各种非共价作用力; (3) 生物大分子特征的空间结构使其具有小分子化合物所不具有的专性识别和结合位点,这些位点通过与相应的配体特异性结合,能形成超分子,这种特性是许多重要生理现象的分子基础。
3. 生物大分子的手性特征有何意义?提示:生物大分子都是手性分子,这种结构特点在生物大分子的分子识别及其特殊的生理功能方面意义重大。
主要表现在: (1) 分子识别是产生生理现象的重要基础,特异性识别对于产生特定生物效应出关重要; (2) 生物大分了通过特征的三维手性空间环境能特异性识别前手性的小分子配体,产生专一性的相互作用。
4.指出取代物的构型:6.举例说明分子识别的概念及其意义。
提示: :分子识别是指分子间发生特异性结合的相互作用,如tRNA分子与氨酰tRNA合成醉的相互作用,抗体与抗原之间的相互作用等。
分子识别是生命体产生各种生理现象的化学本质,是保证生命活动有序地进行的分子基础。
生物化学(第三版)课后习题解答

第一章糖类提要糖类是四大类生物分子之一,广泛存在于生物界,特别是植物界。
糖类在生物体内不仅作为结构成分和主要能源,复合糖中的糖链作为细胞识别的信息分子参与许多生命过程,并因此出现一门新的学科,糖生物学。
O)n的实验式,其化学本质是多羟醛、多羟酮及其衍生物。
糖类按其聚合度分为单多数糖类具有(CH2糖,1个单体;寡糖,含2-20个单体;多糖,含20个以上单体。
同多糖是指仅含一种单糖或单糖衍生物的多糖,杂多糖指含一种以上单糖或加单糖衍生物的多糖。
糖类与蛋白质或脂质共价结合形成的结合物称复合糖或糖复合物。
单糖,除二羟丙酮外,都含有不对称碳原子(C*)或称手性碳原子,含C*的单糖都是不对称分子,当然也是手性分子,因而都具有旋光性,一个C*有两种构型D-和L-型或R-和S-型。
因此含n个C*的单糖有2n个旋光异构体,组成2n-1对不同的对映体。
任一旋光异构体只有一个对映体,其他旋光异构体是它的非对映体,仅有一个C*的构型不同的两个旋光异构体称为差向异构体。
单糖的构型是指离羧基碳最远的那个C*的构型,如果与D-甘油醛构型相同,则属D系糖,反之属L 系糖,大多数天然糖是D系糖Fischer E论证了己醛糖旋光异构体的立体化学,并提出了在纸面上表示单糖链状立体结构的Fischer投影式。
许多单糖在水溶液中有变旋现象,这是因为开涟的单糖分子内醇基与醛基或酮基发生可逆亲核加成形成环状半缩醛或半缩酮的缘故。
这种反应经常发生在C5羟基和C1醛基之间,而形成六元环砒喃糖(如砒喃葡糖)或C5经基和C2酮基之间形成五元环呋喃糖(如呋喃果糖)。
成环时由于羰基碳成为新的不对称中心,出现两个异头差向异构体,称α和β异头物,它们通过开链形式发生互变并处于平衡中。
在标准定位的Hsworth式中D-单糖异头碳的羟基在氧环面下方的为α异头物,上方的为β异头物,实际上不像Haworth式所示的那样氧环面上的所有原子都处在同一个平面,吡喃糖环一般采取椅式构象,呋喃糖环采取信封式构象。
生物化学第10章 酶的作用机理和酶的调节

别够调节可发生在底物-底物、调节物-底物、调节物-调节 物之间,可以是正协同也可以是负协同。
2.别构酶的动力学
别构酶的[S]对V0的动力学曲线不是双曲线,而是S形曲线(正协 同)或表观双曲线(负协同),二者均不符合米氏方程。
定向效应: 底物会诱导酶分子构象改变,使酶活性中心的相 关基团和底物的反应基团正确定向排列,使反应基团之间 的分子轨道以正确方向严格定位,使酶促反应易于进行。
2. 底物的形变(distortion)与诱导契合
当酶遇到其底物时,酶中某些基团或离子可以使底物分子 内敏感键中的某些基团的电子云密度增高或降低,产生“电子 张力”,使敏感键的一端更加敏感,底物分子发生形变,底物 比较接近它的过渡态,降低了反应活化能,使反应易于发生。
[S] (10-4molL-1)
(NAG)2 (NAG)3 (NAG)4 (NAG)5 (NAG)6 (NAG)8
相对水解率
0 1 8 4000 30000 30000
××
ABCDEF
NAG-NAM-NAG-NAM-NAG-NAM
××
NAG-NAG-NAG
NAG-NAG-NAG-NAG NAG-NAG-NAG-NAG-NAG NAG-NAM-NAG-NAM-NAG-NAM
酶与底物给合时构象变化的示意图
3.多元催化和协同效应
在酶催化反应中,几个基元催化反应配合在一起起作用, 如:胰凝乳蛋白酶是通过Asp102, His57,Ser195组成电荷中继网 催化肽键水解,包括亲核和酸碱共同催化共同作用。
4. 活性部位微环境的影响
生物化学第10章酶的作用机制和酶的调节

胰凝乳蛋白酶的三维结构
胰凝乳蛋白酶分子是一个紧密的椭球体,大 小为5.1×4.0×4.0nm。多数芳香和疏水残基包埋 在蛋白质内部,而多数带电的或亲水的残基分布 在 分 子 表 面 。 3 个 极 性 残 基 His57 、 Asp102 和 Ser195 在 活 性 部 位 形 成 催 化 三 联 体 ( catalytic triad)。这三个残基在胰蛋白酶和弹性蛋白酶中 也存在,但这三种蛋白酶的底物结合部位形状不 同,导致它们裂解不同氨基酸羧基侧的肽键。
特异性共价修饰
此类修饰剂特异性地修饰酶活性部位的残基, 同时使酶失活。此类修饰剂与酶的活性部位特异 性结合,同时修饰酶活性部位的基团。例如二异 丙基氟磷酸(DFP)能特异性地与许多酶的活性 部位的Ser残基共价结合,使酶活力丧失,而不修 饰非活性部位的Ser残基。当用DFP修饰酶后,部 分水解酶蛋白,得到含有二异丙基磷酰基的肽段, 分析此肽段的氨基酸序列,可得到活性部位Ser附 近的肽链序列。
胰蛋白酶、胰凝乳蛋白酶、 弹性蛋白酶的底物结合口袋
Trypsin
Chymotrypsin
Elastase
蓝色 His57
红色 Asp102
黄色 Ser195
eglin C
胰 凝 乳 的结合 蛋白酶与
蓝色带状物 为 eglin C (水蛭蛋白 酶抑制剂)
人工底物
对一些底物是大分子聚合物的酶,在研究 其动力学时,一般使用小分子的人工底物。乙 酸-p-硝基苯酯是一种特别有用的模式底物,因 为其水解产物对硝基苯酚在400nm有强的光吸 收,便于监测其变化。
非特异性共价ห้องสมุดไป่ตู้饰
此类修饰剂可以修饰活性部位内或外的残基 侧链基团。若修饰程度与酶活力丧失成正比,则 说明被修饰的基团位于活性部位。当发现修饰程 度与酶活力丧失成正比时,可在加入底物或竞争 性抑制剂的条件下进行修饰,若加入底物或竞争 性抑制剂可阻遏修饰剂对酶活力的破坏作用,则 进一步证明被修饰的基团位于活性部位。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第十章酶的作用机制和酶的调节提要酶的活性部位对于不需要辅酶的酶来说,就是指酶分子中在三维结构上比较靠近的几个氨基酸残基负责与底物的结合与催化作用的部位,对于需要辅酶的酶来说,辅酶分子或辅酶分子上的某一部分结构,往往也是酶活性部位的组成部分。
酶活性部位有6个共同特点。
研究酶活性部位的方法有:酶分子侧链基团的化学修饰法,动力学参数测定法,X射线晶体结构分析法和定点诱变法,这些方法可互相配合以判断某个酶的活性部位。
酶是催化效率很高的生物催化剂,这是由酶分子的特殊结构所决定的。
经研究与酶催化效率的有关因素有7个,即底物和酶的邻近效应与定向效应,底物的形变与诱导契合,酸碱催化,共价催化,金属离子催化,多元催化和协同效应,活性部位微环境的影响。
但这些因素不是同时在一个酶中其作用,也不是一种因素在所有的酶中起作用,对于某一种酶来说,可能分别主要受一种或几种因素的影响。
研究酶催化的反应机制,始终是酶学研究的一个重点,通过大量的研究工作,已经对一些酶的作用机制有深入了解,该章对溶解酶、胰核糖核酸酶A、羧肽酶A、丝氨酸蛋白酶、天冬氨酸蛋白酶等的催化作用机制进行了详尽的讨论。
酶活性是受各种因素调节控制的,除了在第8章中已介绍的几种因素外,主要还有①别构调节,例如ATCase。
②酶原的激活,如消化系统蛋白酶原的激活及凝血系统酶原的激活。
③可逆共价修饰调控,如蛋白质的磷酸化,一系列蛋白激酶的作用。
通过以上作用,使酶能在准确的时间和正确的地点表现出它们的活性。
别构酶一般都是寡聚酶,有催化部位和调节部位,别构酶往往催化多酶体系的第一步反应,受反应序列的终产物抑制,终产物与别构酶的调节部位相结合,由此调节多酶体系的反应速率。
别构酶有协同效应,[S]对υ的动力学曲线呈S形曲线(正协同)或表现双曲线(负协同),两者均不符合米氏方程。
ATCase作为别构酶的典型代表,已经测定了其三维结构,详细研究了别构机制和催化作用机制。
为了解释别构酶协同效应的机制,有两种分子模型受到人们重视,即协同模型和序变模型。
酶原经过蛋白水解酶专一作用释放出肽段,构象发生变化,形成酶的活性部位,变成有活性的酶,这个活化过程,是生物体的一种调控机制。
可逆地共价修饰调控作用是通过共价调节酶进行的,通过其他酶对其多肽链某些基团进行可逆地共价修饰,使处于活性与非活性的互变状态,从而调节酶活性。
共价修饰的基团主要是磷酸化、腺苷酰化、尿苷酰化及ADP-核糖基化等。
同工酶是指催化相同的化学反应,但其蛋白质分子结构、理化性质和免疫性能等方面不同的一组酶。
同工酶是研究代谢调节、分子遗传、生物进化、个体发育、细胞分化和癌变的有力工具,在酶学、生物学及医学研究中占有重要地位。
LDH同工酶研究的比较清楚,是由良种不同亚基组成的四聚体,有5种同工酶,在不同组织中含量不同,反映了同工酶的组织特异性。
习题1.阐明酶活性部位的概念。
可使用那些主要方法研究酶的活性部位?答:酶的活性部位对于不需要辅酶的酶来说,就是指酶分子在三维结构上比较靠近的几个氨基酸残基负责与底物的结合与催化作用的部分;对于需要辅酶的灭来说,辅酶分子或辅酶分子上的某一部分结构,往往也是酶活性部位的组成部分。
研究酶活性部位的方法有:酶分子侧链基团的化学修饰法、动力学参数测定法、X射线晶体结构分析法和定点诱变法。
2.简要阐明胰Rnase A的活性部位如何确定?答:用化学修饰法研究Rnase A活性的必须氨基酸残基。
在pH5.5下,用等摩尔碘乙酸处理Rnase A,羧甲基化的His119是主要产物,而羧甲基化的His12产物较少。
Rnase A中其他组氨酸对这个试剂的反应弱得多。
所得Rnase A的两个羧甲基化的衍生物均无活性,因此可推测His119和His12为酶活性的必需集团。
2,4二硝基氨苯可选择地同酶Lysε-NH2反应,酶引起失活。
该结果表明Lys41也是酶活性部位的必需氨基酸。
以上研究结果可认为His119、His12、Lys41构成了Rnase A的活性部位。
3.与酶催化效应有关的因素有哪些?它们是怎样提高反应速率的?答:与酶催化速率有关的因素有7个:1.底物和酶的邻近效应与定向效应:酶和底物复合物的形成过程既是专一性的识别过程,更重要的是分子间反应变成分子内反应的过程。
在这一过程中包括两种效应、邻近效应和定向效应。
邻近效应是指酶与第五结合形成中间复合物以后,使底物和底物(如双分子反应)之间,酶的催化基团与底物之间结合于同一分子而使有效浓度得以极大的升高,从而使反应速率大大增加的一种效应。
定向效应是指反应物的反应基团之间和酶的催化基团与底物的反应基团之间的正确取位产生的效应。
2.底物的形变和诱导契合:当酶遇到其专一性底物时,酶中某些基团或离子可以使底物分子内敏感键中的某些基团的电子云密度增高或降低,产生“电子张力”,使敏感键的一端更加敏感,底物分子发生形变,底物比较接近它的过渡态,降低了反应活化能,使反应易于发生。
3.酸碱催化:通过瞬时的向反应物提供质子或从反应物接受质子以稳定过渡态,加速反应的一类催化机制。
4.共价催化(亲核催化或亲电子催化):在催化时,亲核催化剂或亲电子催化剂能分别放出电子或吸取电子并作用于底物的缺电子中心或负电中心,迅速形成不稳定的共价中间复合物,降低反应活化能,使反应加速。
5.金属离子催化:金属离子以3种主要途径参加催化过程:(1)通过结合底物位反应定向;(2)通过可逆地改变金属离子的氧化态调节氧化还原反应;(3)通过静电稳定或屏蔽负电荷,作用比质子强,不少金属离子有络合作用,并且在中性pH溶液中,H+浓度很低,但金属离子却容易维持一定浓度。
金属离子通过电荷的屏蔽促进反应。
金属离子通过水的离子化促进亲核催化。
6.多元催化和协同效应:酶催化反应中常常几个基元催化反应配合在一起共同起作用,加速酶反应。
7.活性部位微环境的影响:在酶分子的表面有一个裂缝,而活性部位就位于疏水环境的裂缝中,大大有利于酶的催化作用。
4.推测下列寡聚糖被溶菌酶水解的相对速率:(G:N-乙酰氨基葡萄;M:N-乙酰氨基葡萄乳酸)(1)M-M-M-M-M-M;(2)G-M-G-M-G-M;(3)M-G-M-G-M-G5.假设在合成(NAG)时D和E糖残基之间的糖苷氧已为18O所标记,当溶菌酶水解时,18O将出现在哪个产物中?答:18O将出现在由A、B、C、D残基组成的四聚体中。
6.请比较溶菌酶、羧肽酶A、胃蛋白酶和胰凝乳蛋白酶:(1)哪一种酶的催化活性需要金属离子?(2)哪种酶只含一条多肽链?(3)哪种酶被DFP迅速地失活?(4)哪种酶是由酶原激活成的?答:(1)羧肽酶A;(2)溶菌酶;(3)胰凝乳蛋白酶;(4)羧肽酶A、胃蛋白酶、胰凝乳蛋白酶。
7.上题4种酶的催化机制中是否有从酶到底物的质子转移过程?若有请指出它们的质子供体是什么?答:溶菌酶中的Glu35的-COOH提供一个H+;羧肽酶A中的Glu270的-COOH提供一个H+;胃蛋白酶中的Asp32的-COOH提供一个H+;胰凝乳蛋白酶中Ser195提供一个H+。
8.TPCK是胰凝乳蛋白酶的亲和标记试剂,它对His57烷基化后使胰凝乳蛋白酶失活。
(1)为胰蛋白酶设计一个像TPCK那样的亲和标记试剂。
(2)你认为怎样可以检验它的专一性?(3)能被胰蛋白酶的这个亲和标记试剂失去活性的还有哪个丝氨酸蛋白酶?9.胰凝乳蛋白酶、胰蛋白酶和弹性蛋白酶作为催化剂有哪些相似之处?有哪些不同之处?在酶的分子结构上是哪些因素引起这些差异?答:相似之处:①执行相同的反应——裂解肽键;②其结构和作用机制很相似;③相对分子质量范围在2.5×103,并且具有相似的顺序和三级结构;④3个极性残基——His57、Asp102和Ser195在活性部位形成催化三联体。
不同之处:①专一性不同:胰蛋白酶裂解碱性氨基酸Arg和Lys羧基侧链肽;胰凝乳蛋白酶选择裂解芳香氨基酸如Phe和Tyr羰基侧链;弹性蛋白主要裂解小的中性氨基酸残基羰基侧链肽;②酶活性部位不同:胰蛋白酶口袋中,有一个负电荷Asp189在底部,有利于结合正电荷Arg和Lys残基;胰凝乳蛋白酶口袋被疏水氨基酸环绕,大的足以容纳一个芳香残基;弹性蛋白口袋浅,开口处具有大的Thr和Val残基,仅小的,部大的残基能够容纳在它的口袋中。
10.ATCase是一种别构酶,其活性部位与别构效应物结合部位分别处于不同亚基之上,有可能设想别构酶上这两种部位存在于同一亚基上吗?为什么?11.对于ATCase来说,琥珀酸起着Asp(两个底物中的一个)的竞争抑制作用。
υ对[Asp]的依赖关系见图10-71A(假设这些实验中第二种底物是过量的并可忽略)。
在图10-71B种[Asp]维持在低水平(图10-71A种箭头所指处)不变,并加入一系列含量递增的琥珀酸。
琥珀酸不能作为底物参与反应。
请解释这些结果。
答:(图略)琥珀酸的结合导致协同由T型向R型转变。
12.试解释为什么胰凝乳蛋白不能像胰蛋白酶那样自我激活?13.羧肽酶A促使底物的水解,下面哪个是其重要的机制:(1)结构重排将必需氨基酸残基靠近敏感键。
(2)形成一个C端环肽的共价中间物。
(3)活性部位Try残基脱质子形成亲核作用。
(4)通过结合Zn2+的活化水。
14.左边裂处的每一种酶,按照提出的催化机制,从右边选择出它们适当的过渡态或化学本质。
(1)溶菌酶——(4)(1)Zn2+的活化水(2)RNA酶——(3)(3)羧肽酶A——(1)(4)胰凝乳蛋白酶(2)(5)(2)氧阴离子(3)五价磷(4)碳正离子(5)胃蛋白酶——(6)(5)四面体肽键(6)天冬氨酸——活化水15.对蛋白酶A的叙述中哪一个是正确的?(1)通过ATP活化。
——(×)(2)在没有激活剂时有2个催化亚基(C)和两个调节亚基(R)组成。
——(×)(3)激活剂结合后解离成一个C2和2个R亚基。
——(×)(4)在C亚基中含有一个假底物顺序。
——(√)16.苯甲脒(Ki=1.8×10-5mol/L)和亮抑蛋白酶肽(Leupeptin Ki=3.8×10-7mol/L)是胰蛋白酶的两种特异竞争性抑制剂,试解释它们的抑制机制。
设计亮抑蛋白酶肽的类似物抑制胰凝乳蛋白酶和弹性蛋白酶。