草本植物抗旱性研究
藜麦种质资源及抗旱和耐盐的研究进展

藜麦种质资源及抗旱和耐盐的研究进展藜麦是一种野生草本植物,主要分布在南美洲、西亚和欧洲。
近年来,由于其高蛋白、高营养价值以及良好的环境适应性等特点,藜麦在全球范围内受到越来越多的关注。
目前,全球各地对藜麦种质资源的研究已经展开,涉及到藜麦的产量、品质、适应性以及抗逆性等不同方面。
藜麦种质资源目前藜麦种质资源的收集和保存工作还比较落后,尤其是大部分藜麦种质资源都集中在南美洲,其他地区种质资源数量比较有限。
但是随着人们对藜麦种质资源的重视,国内外一些研究机构开始积极开展藜麦种质资源的收集保存工作。
已经发现了大量藜麦的荒漠型、山地型、杂交型等不同类型藜麦资源,这些资源可以为藜麦的种质改良提供丰富的遗传基础。
抗旱性研究藜麦对干旱的适应性非常强,其生长期间仅需少量水分就可以生长良好。
但是藜麦在生长初期对干旱的适应性比较差,要想让藜麦具备更强的抗旱能力,需要通过遗传改良等手段进行研究和改进。
目前,国内外一些科学家通过对藜麦抗旱相关基因的研究,发现了一些重要的基因,这些基因可以为藜麦的抗旱育种提供重要的参考。
耐盐性研究藜麦对盐碱地的适应性非常强,其可以在盐碱地上正常生长和发育。
这主要得益于藜麦对盐分的耐受能力很强,其种子和幼苗可以承受高浓度的盐分。
目前,国内外一些研究机构都在积极探索藜麦的耐盐基因,并通过基因编辑等技术手段实现对耐盐基因的精准修饰,从而进一步提高藜麦在盐碱地上的适应性和产量。
总体来说,藜麦的种质资源丰富,其抗旱和耐盐性能力突出,为其在不同地理环境中的应用提供了很好的基础。
但是目前藜麦相关研究还比较缺乏,未来需要加强对藜麦的研究和开发。
香根草、菖蒲、香蒲的耐淹抗旱性研究

香根草、菖蒲、香蒲的耐淹抗旱性研究消落带是湖库河道滨岸下方、水体上方干湿交替呈现的地段。
因此,消落带种植的植物必须兼具耐水淹性和抗旱性。
为筛选丹江口水库消落带适生植物,本试验选择了三种已经在南阳地区种植多年的草本植物——香根草、菖蒲和香蒲,对其进行不同程度水淹和干旱试验,分析了三种植物水淹和干旱下的生长形态、生理变化,比较了耐淹抗旱性。
主要研究结果如下:1.刈去三种植物地上部分后,对其地下部分进行6个月水淹,结果表明:1)水淹结束后,统计三种植物的新芽萌发率由高到低的顺序为:菖蒲(80%)>香根草(49%)>香蒲(0%);2)水淹6个月后,菖蒲株高、根长、生物量较对照组均显著增加,而香根草株高虽有增加但不明显;3)水淹6个月后,与对照相比,香根草、菖蒲的最大光合效率、净光合速率、蒸腾速率、气孔导度、叶绿素含量、根活力、根际土壤微生物量均有不同程度的增长,但综合来看菖蒲增量最大;4)水淹6个月后,香根草SOD活性相比对照显著提高,菖蒲虽提高但无显著差异;香根草、菖蒲MDA量较对照相比均减少但差异不显著,但菖蒲的减量最小;香根草CAT活性较对照相比显著提高,菖蒲有所增高但差异不显著;以上结果表明,6个月根淹下,菖蒲的新芽萌发率最高,恢复生长能力最强,对根淹环境的适应力最佳,其次是香根草,最不耐水淹的是香蒲。
2.以正常浇水为对照,对三种植物幼苗设置了不同深度(浅淹5 cm、半淹10 cm、深淹15 cm)的水淹培养试验,试验进行6个月。
结果表明:1)水淹结束后,统计三种植物不同水深下的存活率由高到低的顺序为:香根草浅淹(83.1%)>半淹(72.9%)>深淹(60.4%);菖蒲浅淹(91.8%)>半淹(82.9%)>深淹(69.4%);香蒲各水淹组均为0;2)水淹6个月后,与对照比,浅淹下香根草的株高、根长、生物量均显著增长,而半、深淹下均有不同程度的减少;菖蒲在各水深下上述指标相比对照均有不同程度增加,总体看菖蒲的增量要大于香根草;3)水淹6个月后,香根草、菖蒲各水深下的最大光合效率、净光合速率、蒸腾速率、气孔导度大多高于对照;香根草叶绿素含量与对照相近,而菖蒲与对照相比显著增加;浅淹下香根草的根活力与对照相比显著增加,其余水淹下与对照接近,半、深淹下菖蒲较对照也显著增加,但浅淹下香根草的增量最大;在三种水淹深度下香根草、菖蒲的根际微生物量相比对照均显著增长,其中菖蒲深淹组的增量最大;4)水淹6个月后,香根草、菖蒲各处理的SOD活性相比对照均无显著差异;香根草各组CAT 活性相比对照虽有增加但差异不显著,菖蒲各组CAT活性相比对照均显著增加;浅淹深度的香根草MDA量相比对照显著降低,其他处理虽有降低但较对照无差异,菖蒲各处理的MDA量与对照相近;以上结果表明,三种植物幼苗水淹下,菖蒲耐淹性最强,其次香根草,最后为香蒲。
植物抗旱途径

植物抗旱途径随着全球气候变化的加剧,干旱问题越来越严重。
而植物作为生物体,在干旱环境下生存,需要具备一些特殊的适应性。
本文将探讨植物在抗旱途径方面的表现及其机制。
一、植物的抗旱途径植物在遭遇干旱环境时,为了保持生命活动,会采取一系列的途径来适应环境。
这些途径包括:调节气孔开合、调节根系生长、积累水分、调节植物体内的代谢物质等。
1. 调节气孔开合气孔是植物体内水分的主要通道,调节气孔开合是植物在干旱环境下的一种适应性。
在干旱环境下,植物会通过调节气孔的开合来减少水分的散失。
植物通过调节气孔的大小和数量,来控制水分的流动和蒸发,从而达到保持水分的目的。
2. 调节根系生长植物的根系是吸收水分的主要器官,根系的生长和分布对于植物的抗旱能力有着重要的影响。
在干旱环境下,植物会通过调节根系的生长和分布来适应环境。
一些植物会增加根系的生长深度,以便吸收更深层的水分;另一些植物则会增加根毛的生长,以增加吸收水分的面积。
3. 积累水分植物在干旱环境下,为了保持水分,会积累水分。
一些植物会通过在植物体内积累水分来适应干旱环境。
例如,一些多年生草本植物可以在雨季时积累水分,并在干旱季节中利用这些水分来维持生命活动。
4. 调节植物体内的代谢物质植物在干旱环境下,为了维持生命活动,会调节植物体内的代谢物质。
例如,植物会调节蛋白质的合成和降解,以适应干旱环境。
植物还会调节酶的活性,以增加植物体内的代谢反应速率。
二、植物的抗旱机制植物在抗旱途径方面的表现,是由植物的抗旱机制所支撑的。
植物的抗旱机制包括:调节水分的平衡、调节渗透压、调节脱水蛋白、调节生长素等。
1. 调节水分平衡植物在干旱环境下,为了保持水分平衡,会通过调节水分的吸收和释放来适应环境。
植物会减少水分的散失,同时增加水分的吸收。
植物还会利用水分的蒸发来降低植物体内的温度,从而减少水分的蒸发。
2. 调节渗透压植物在干旱环境下,为了保持水分平衡,会通过调节渗透压来适应环境。
广东绿化用佛甲草

广东绿化用佛甲草佛甲草(学名为禾本科佛甲草属,Heteropogon contortus)来历可以追溯到古印度,在公元前二世纪形成的《美颜经》提到:佛甲草的发芽对人体有很好的益处,但更重要的是,它是一种非常有效的抗旱草本植物,因此在农业改革中占据着重要的地位。
相传,佛甲草是在南印度布哈拉邦(今泰米尔纳德邦)的香蕉园里发现的,在全国范围内,佛甲草在广东的发挥了重要的作用,在广东的土壤条件下,这种草本植物发展良好,有效地解决了广东地区由于贫瘠土壤导致的水源缺乏和土壤流失的问题。
佛甲草的有效性源于其独特的地质结构,这种复杂的结构可以有效地利用微小空间,捕捉水分,可以节约灌水时间,佛甲草在上面可以把水分长期储存在关联结构中,以便后期再利用,可以把大量的水分储存起来,以减轻广东地区的水短缺问题。
另外,佛甲草具有极强的抗旱能力,它可以在低温、旱涝耐受度范围内长期生存,而且抗旱能力强,低温抗性强,抗病虫草能力高,抗逆能力极强,可以把土壤的水分贮存起来,确保广东植物的长期生长。
此外,佛甲草对生态系统也有重要的作用,它可以防止沙尘风暴,改善环境污染,增加氧气,改善气候,保护生物多样性,降低土壤污染。
另外,这种植物也可以提供良好的子财物,可以丰富植物群落,从而改善环境质量。
因此,佛甲草在广东的绿化工作中发挥了重要作用,在广东的贫瘠土壤、水资源紧缺等情况下,佛甲草的抗旱、节水、抗逆、抗病虫草能力被普遍采用,成为广东省绿化工作的重要组成部分,为改善广东省环境质量和植被结构作出了重要贡献。
佛甲草作为一种抗旱草本植物,一直受到各界的高度重视,在全国各地以及世界各地都有大量的研究论文、案例研究和调研报告为佛甲草的研究和应用提供了有价值的参考。
在未来,佛甲草在广东省绿化工作中的作用将得到进一步发挥,为改善广东省的水资源、天气和环境质量等作出更大的贡献。
总的来说,佛甲草在广东的绿化工作中发挥了重要作用,它的特殊的地质结构可以有效地利用微小空间,捕捉水分,从而节约灌水时间,可以把水分长期储存起来;此外,它还具有极强的抗旱能力,抗病虫草能力强,抗逆能力极强,有助于改善广东省环境质量和植被结构,为改善广东绿化作出了重要贡献。
三种三叶草抗旱性分析
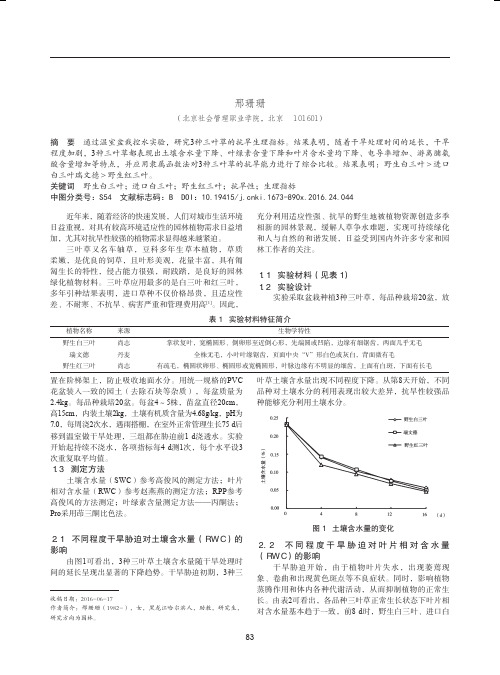
图1 土壤含水量的变化
2.2 不同程度干旱胁迫对叶片相对含水量(RWC)的影响
干旱胁迫开始,由于植物叶片失水,出现萎蔫现象、卷曲和出现黄色斑点等不良症状。
同时,影响植物蒸腾作用和体内各种代谢活动,从而抑制植物的正常生长。
由表2可看出,各品种三叶草正常生长状态下叶片相对含水量基本趋于一致,前8 d时,野生白三叶、进口白
图2 水分胁迫下叶片叶绿素总量的变化2.4 不同程度干旱胁迫对游离脯氨酸(Pro)含量的影响
从表3可看出,各品种正常生长状况下,游离脯氨酸的含量次为:野生红三叶>进口白三叶瑞文德>野生红三叶。
随着干旱胁迫的延长,各品种游离脯氨酸开始增加,各品种三叶草间积累游离脯氨酸的能力差别很大,尤其在第12天时,游离脯氨酸的积累达到高峰,品种间差异达到显著水平,三叶草品种间积累游离脯氨酸
的能力依次为:野生白三叶>进口白三叶瑞文德>野生。
高粱抗旱性研究进展

㊀山东农业科学㊀2024ꎬ56(1):164~173ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2024.01.022收稿日期:2023-03-27基金项目:国家重点研发计划项目(2022YFD1500602-1)ꎻ国家现代农业产业技术体系项目(CARS-06-14.5-B16)作者简介:王晓东(1996 )ꎬ男ꎬ研究实习员ꎬ主要从事高粱栽培与育种工作ꎮE-mail:1009124737@qq.com通信作者:肖继兵(1976 )ꎬ男ꎬ研究员ꎬ主要从事旱作农业研究ꎮE-mail:xiaojb2004@126.com高粱抗旱性研究进展王晓东ꎬ李俊志ꎬ窦爽ꎬ肖继兵ꎬ辛宗绪ꎬ吴宏生ꎬ朱晓东(辽宁省旱地农林研究所ꎬ辽宁朝阳㊀122000)㊀㊀摘要:干旱是限制植物生产力和威胁粮食安全的重要因素之一ꎮ高粱(SorghumbicolorL.Moench)是全球主粮和饲料作物ꎬ因其具有较强的抗旱性和能够在恶劣的环境条件下生存而广泛种植于干旱半干旱地区ꎬ在作物抗旱领域中具有重要的研究价值ꎮ深入解析干旱胁迫下高粱的形态和生理特性㊁鉴定和筛选抗旱品种㊁挖掘相关抗旱基因ꎬ对推动高粱抗旱育种进程㊁提高品种抗旱性㊁提高产量具有重要意义ꎮ本文从干旱胁迫对高粱生长的影响㊁高粱对干旱胁迫的生理响应㊁高粱耐旱性鉴定方法和鉴定指标㊁高粱抗旱性分子生物学和提高高粱抗旱性方法5个方面对高粱抗旱性研究进展进行综述ꎬ并对高粱抗旱性研究方向进行展望ꎬ以期为进一步研究高粱抗旱的形态㊁生理特性及分子机制奠定基础ꎮ关键词:高粱ꎻ干旱胁迫ꎻ生理响应ꎻ分子生物学ꎻ鉴定ꎻ抗旱性中图分类号:S514㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2024)01-0164-10ResearchProgressonDroughtResistanceofSorghumWangXiaodongꎬLiJunzhiꎬDouShuangꎬXiaoJibingꎬXinZongxuꎬWuHongshengꎬZhuXiaodong(LiaoningInstituteofAgriculture&ForestryinAridAreasꎬChaoyang122000ꎬChina)Abstract㊀Droughtisoneoftheimportantfactorsthatlimitplantproductivityandthreatenfoodsecurity.Asaglobalstaplefoodandforagecropꎬsorghum(SorghumbicolorL.Moench)hasgoodcharacteristicsindroughtresistancealongwithabilitiestosurviveinharshenvironmentsꎬandiswidelyplantedinaridandsemi ̄aridareasꎬwhichgiveitimportantresearchvaluesinthefieldofcropdroughtresistance.Itisofgreatsignifi ̄canceinacceleratingbreedingprocessofdrought ̄resistantvarietiesandincreasingdroughtresistanceandyieldofsorghumtofurtheranalyzethemorphologicalandphysiologicalcharacteristicsunderdroughtstressꎬidentifyandscreentheexcellentdrought ̄resistantvarietiesꎬanddigoutdrought ̄resistantgenes.Inthispaperꎬthere ̄searchprogressindroughtresistanceofsorghumwasreviewedfrominfluencesofdroughtstressonsorghumgrowthꎬphysiologicalresponsesofsorghumtodroughtstressꎬidentificationmethodsandindexesꎬmolecularbiologyꎬandimprovementmethodsꎬandtheprospectofresearchdirectionofdroughtresistanceinsorghumwasproposedꎬinordertolayafoundationforfurtherstudyofthemorphologicalandphysiologicalcharacteris ̄ticsandmolecularmechanismsofdroughtresistanceinsorghum.Keywords㊀SorghumꎻDroughtstressꎻPhysiologicalresponseꎻMolecularbiologyꎻIdentificationꎻDroughtresistance㊀㊀干旱是限制作物生产发展的最重要因素之一ꎬ有发生范围广㊁频次高㊁持续时间长等特点[1-2]ꎮ目前ꎬ世界上有三分之一以上总陆地面积的干旱和半干旱地区ꎬ我国现有干旱㊁半干旱和亚湿润干旱区近300万km2ꎬ占国土总面积近四成[3]ꎮ其中ꎬ绝大部分是因为缺乏灌溉条件而以雨养农业为主ꎬ其作物产量占全国总产量的比重较小ꎮ选育耐旱性强的作物品种是保证干旱地区高产稳产的重要举措ꎮ干旱可能会发生在作物生长发育的各个阶段ꎮ然而ꎬ在干旱和半干旱地区ꎬ作物生长季开始和结束时发生干旱的可能性较高ꎮ生长季节开始时的干旱胁迫严重影响植物的生长发育ꎮ如果干旱发生在作物开花期或灌浆期ꎬ可能会导致产量严重下降或歉收[4]ꎮ高粱(SorghumbicolorL.Moench)是禾本科一年生草本植物ꎬ主要种植于热带㊁亚热带和温带的干旱半干旱区ꎬ也是我国主要的杂粮作物之一ꎬ是重要的酿用㊁食用㊁饲用㊁帚用作物ꎬ同时也是全球仅次于水稻㊁玉米㊁小麦㊁大豆种植面积的第五大粮食作物ꎮ高粱具有很强的抗旱㊁耐涝㊁耐盐碱㊁耐瘠薄㊁耐高温等抗逆特性[5]ꎮ高粱不同品种间抗旱能力存在较大差异ꎮ近些年从多个方面开展了高粱抗旱性遗传和抗旱品种选育相关研究[6-7]ꎮ本文综述干旱胁迫对高粱生长的影响㊁高粱耐旱性鉴定方法和鉴定指标ꎬ以及高粱对干旱的生理响应ꎬ并从转录组分析㊁抗旱QTL定位和全基因组关联分析方面进行梳理和整合ꎬ并对高粱抗旱性的分子调控机制㊁鉴定体系及抗旱性品种选育进行展望ꎬ以期为后人开展相关研究提供理论参考ꎮ1㊀干旱胁迫对高粱生长的影响1.1㊀干旱胁迫对高粱种子萌发和幼苗生长的影响水分缺乏使植物发育迟缓ꎬ干旱胁迫达到一定阈值时ꎬ会显著抑制种子萌发和幼苗生长[8]ꎮ王志恒等[9]研究了高粱萌发阶段受干旱胁迫的响应特性ꎬ发现随着干旱胁迫程度的增加ꎬ高粱种子的发芽率㊁发芽势等显著降低ꎬ种子残留干重逐渐增加ꎬ干物质转移㊁转化效率逐渐下降ꎬ根冠比逐渐增大ꎬ比根重逐渐减小ꎮ长期干旱胁迫降低幼苗的苗高㊁叶长ꎬ幼苗地上部和根的鲜重不同程度的下降[10]ꎮ1.2㊀高粱萌发期及苗期的抗旱性研究大多数农作物在种子萌发㊁幼苗形成和开花阶段对干旱胁迫较为敏感ꎬ干旱胁迫下萌发期和苗期表现出耐旱性是作物生长发育的前提ꎮ对高粱萌发期和苗期耐旱性的研究发现ꎬ高粱萌发期和苗期的耐旱性是不一致的ꎮ张笑笑[11]对73份高粱品种进行萌发期和苗期耐旱性鉴定ꎬ初筛结果发现萌发期和苗期都抗旱的品种5份ꎬ苗期抗旱品种10份ꎬ田间和室内采用多重表型分析最终得到苗期抗旱品种1份ꎮ郝培彤等[12]在20%PEG干旱胁迫下评价21份饲草高粱材料的耐旱性ꎬ筛选出萌发期耐旱和苗期耐旱材料各3份ꎬ萌发期和苗期共同耐旱材料1份ꎮ由此可见ꎬ高粱品种萌发期和苗期耐旱性是不同的ꎬ萌发期耐旱品种苗期不一定耐旱ꎬ苗期耐旱品种萌发期也可能不耐旱ꎮ针对高粱萌发期和苗期耐旱性ꎬ许多学者是分开进行研究的ꎬ而在大田干旱生产条件下ꎬ种子从萌发阶段就已经受到干旱胁迫的影响ꎮ因此研究植物的耐旱性ꎬ应该从种子萌发到苗期进行不间断的干旱胁迫处理ꎬ这样可以更加全面地反映出植物在萌发期和苗期对干旱胁迫的各种反应ꎮ1.3㊀干旱胁迫对高粱光合作用的影响Zhang等[13]研究发现ꎬ在干旱胁迫处理后ꎬ高粱叶片叶绿素总含量及叶绿素a㊁叶绿素b含量降低ꎬ且叶绿素a的降低幅度显著大于叶绿素bꎮ干旱胁迫下叶绿素含量降低主要是由于叶绿素生物合成下降ꎬ从而导致叶绿素加速分解ꎮ植物进行光合作用时ꎬ要保证充足的光照ꎬ然而光照过强ꎬ会造成叶片吸收的光能超出同化所需ꎬ进而造成光抑制或者光破坏[14]ꎮ因此植物会通过植物激素㊁外源物质等来减缓由于光能过多引起的光抑制ꎬ促进光合活性ꎬ避免PSⅡ系统受到破坏[15]ꎮ张姣等[16]的研究表明ꎬ干旱胁迫下ꎬ高粱叶片的净光合速率(Pn)㊁气孔导度(Gs)㊁最大光化学效率(Fv/Fm)㊁光化学淬灭系数(qP)㊁电子传递速率(ETR)出现不同程度的下降ꎬ初始荧光(Fo)与对照组相比有所升高ꎬZhang等[13]也得出相同的结论ꎮ说明干旱胁迫会使光合相关酶活性丧失ꎬ导致光能过剩而产生积累ꎬ通过热耗散等途径消耗多余的光能ꎬ可以让作物适应干旱胁迫环境ꎮ干旱胁迫导致光合作用能力下降主要原因是非气孔限制[17]ꎮ王祁等[18]的研究还发现ꎬ在轻度干旱胁迫下ꎬ高粱叶片PSⅡ系统结构和功能损伤较小ꎬ然而在重度胁迫下ꎬ叶片PSⅡ系统遭到破坏ꎬ进而发生光抑制现象ꎮ光合作用的强弱可以直接反映出植物抵御干旱胁迫的能力ꎬ保证叶绿素含量的稳定㊁保护光合相关酶活性ꎬ可抵561㊀第1期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀王晓东ꎬ等:高粱抗旱性研究进展御干旱胁迫对其光合作用的影响ꎮ2㊀高粱对干旱胁迫的生理响应2.1㊀有机渗透调节干旱胁迫下ꎬ植物细胞通过调节可溶性物质的浓度来维持细胞内外渗透压平衡ꎬ进而应对干旱胁迫带来的影响ꎮ参与渗透调节的物质可分为两类ꎬ第一类是外界环境提供的无机离子ꎬ第二类是胞内合成的有机溶质ꎮ第二类渗透调节物质主要包括甜菜碱㊁脯氨酸㊁糖和糖醇等有机化合物[19]ꎮ近些年来ꎬ可溶性蛋白㊁可溶性糖㊁脯氨酸等渗透调节物质被广泛研究ꎮ脯氨酸的积累可以使许多植物应对渗透胁迫反应ꎬ作为一种相容的渗透剂ꎬ其可以提高细胞或组织的保水能力ꎬ同时可以作为碳水化合物的来源ꎻ作为一种酶的保护剂ꎬ也可以减轻蛋白质变性ꎬ具有很强的抗氧化能力ꎮ张玉霞等[20]用聚乙二醇溶液模拟干旱胁迫ꎬ结果表明饲用高粱品种脯氨酸含量与对照组相比显著升高ꎮ王艳秋等[21]研究发现ꎬ干旱胁迫下高粱叶片的脯氨酸含量显著增加ꎬ且其显著性较大ꎬ是高粱调节适应干旱胁迫的重要指标ꎮ可见ꎬ脯氨酸在干旱胁迫下至关重要ꎬ但是也有不同观点:董喜存等[22]的研究发现ꎬ在不同程度干旱胁迫下ꎬ甜高粱品种叶片脯氨酸含量变化趋势并不一致ꎮ因此认为ꎬ单纯测定脯氨酸含量不能准确反映抗旱性ꎬ可以将其作为一种抗旱胁迫下的保护性反应ꎮ可溶性糖主要包括葡萄糖㊁蔗糖㊁果糖和半乳糖ꎮ可溶性糖既可以为植物生长发育提供能量ꎬ并且具有信号功能ꎬ又是植物生长发育的重要调节因子[23]ꎮ何玮等[24]研究不同干旱胁迫下甜高粱叶片可溶性糖含量的变化时发现ꎬ轻度干旱胁迫下可溶性糖含量先下降然后突然升高ꎬ再下降之后突然升到最高ꎻ在重度干旱胁迫下ꎬ可溶性糖含量先下降ꎬ然后升到最高ꎬ再下降ꎮ总体表明ꎬ在受到干旱胁迫时ꎬ高粱叶片可溶性糖含量整体呈升高趋势ꎮGill等[25]研究不同非生物胁迫下高粱可溶性糖含量变化的结果表明ꎬ干旱胁迫下总的可溶性糖含量呈升高趋势且高于对照ꎬ其中果糖含量始终高于葡萄糖和蔗糖ꎮ此外ꎬ在干旱胁迫下ꎬ可溶性糖还可以作为蛋白质渗透保护剂而发挥作用ꎮ可溶性蛋白含量的变化可以直接反映植物渗透调节能力的大小ꎬ它不仅可以提高细胞的保水能力ꎬ而且可以有效地保护生物膜以及细胞的生命物质ꎮ荣少英等[26]研究不同高粱品种在不同干旱条件下可溶性蛋白的变化时发现ꎬ甜高粱㊁普通高粱和对照相比可溶性蛋白含量随着干旱胁迫的加剧呈上升趋势ꎮ有研究[27-28]表明ꎬ在逆境胁迫下ꎬ膜质过氧化产物丙二醛抑制蛋白质的生物合成ꎻ长时间重度干旱使植物体内分解代谢加剧ꎬ导致大量可溶性蛋白分解ꎮ2.2㊀抗氧化防御系统活性氧具有很强的氧化能力ꎮ植物在进行有氧代谢的过程中会产生活性氧ꎬ低浓度的活性氧可以作为信号分子参与调控植物非生物胁迫反应[29-30]ꎮ抗氧化防御系统具有维持植物体内活性氧平衡的功能[31]ꎮ该系统包括两大类ꎬ一类是非酶促抗氧化物质ꎬ其中最为重要的是水溶性抗坏血酸(Asc)ꎬ其次是谷胱甘肽(GSH)ꎬ还有脂溶性生育酚㊁类胡萝卜素等ꎻ另一类是酶促抗氧化剂ꎬ包括超氧化物歧化酶(SOD)㊁过氧化物酶(POD)和过氧化氢酶(CAT)[32]ꎮ植物抗氧化调控系统中ꎬ提高酶活性和抗氧化物的表达量是作物抵御逆境胁迫的关键因素ꎮ陈敏菊等[33]研究发现ꎬ高粱幼苗叶片SOD和CAT活性因干旱胁迫的强度不同而存在差异ꎬ土壤含水量在55%~60%时SOD活性逐渐升高ꎬ随着干旱程度加剧ꎬSOD活性逐渐下降ꎻCAT活性在土壤含水量为40%~60%时高于对照ꎬ随着干旱加剧活性逐渐降低ꎻPOD活性的变化规律和SOD一致ꎮ这表明轻度干旱胁迫可以提高高粱幼苗叶片抗氧化酶活性ꎮ卢峰等[34]研究高粱幼苗不同生长阶段受到干旱胁迫时酶活性的变化情况表明ꎬ胁迫6㊁8㊁12㊁24d时SOD活性显著高于对照ꎬ干旱胁迫12d时酶活性达到峰值ꎬPOD活性的变化和SOD基本一致ꎮ说明高粱幼苗在受到干旱胁迫时ꎬ通过提高叶片保护酶活性来抵御其危害ꎮ2.3㊀激素调节植物激素参与干旱胁迫调节ꎮ通过外源激素来提高作物的抗旱性是现阶段重要的科学途径之一[35-36]ꎮ细胞分裂素通过促进细胞分裂ꎬ延缓植物叶片中叶绿素的降解来提高植物的抗旱性[37]ꎮ661山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀生长素可以正向调控四种抗氧化酶(SOD㊁CAT㊁POD㊁GR)活性来降低干旱胁迫对植株的抑制作用ꎻ同时生长素可以通过调节根生物量㊁增加根的分支来提高水分吸收效率进而提高抗旱性[38]ꎮ脱落酸(ABA)对植物在逆境胁迫下的应答起着关键作用ꎬ其参与气孔的开关ꎬ保卫细胞的通道活动ꎬ调节转录钙调蛋白的表达ꎬ诱导相关基因的表达[39]ꎮ植物在受到干旱胁迫时ꎬ也可以通过减少赤霉素的方式来适应胁迫环境[40]ꎮ关于干旱胁迫下植物激素调节机制ꎬ国内外对水稻㊁小麦等作物的报道较多ꎬ逆境胁迫下高粱中植物激素的作用机制还需进一步探讨和研究ꎮ高粱的抗旱性在生理上涉及到三个方面:第一是干旱胁迫下需要维持高的含水量ꎬ维持高粱水分平衡ꎬ通过增加脯氨酸㊁可溶性糖㊁可溶性蛋白质等物质含量提高渗透调节能力ꎬ维持细胞或者组织持水ꎬ进而维持膨压ꎻ第二是干旱胁迫下保证其基本的生理功能ꎬ通过激素调节㊁酶活性提高等来维持高粱正常的生理功能ꎻ第三是干旱胁迫解除时高粱含水量和生理功能的恢复能力ꎮ做好以上三点ꎬ可以有效地抵御干旱胁迫带来的负面影响ꎮ3㊀高粱耐旱性鉴定方法及鉴定指标因为各个时期的耐旱机制不同ꎬ一般将高粱耐旱性鉴定分为萌发期㊁苗期和全生育期鉴定ꎮ萌发期是作物在干旱胁迫条件下能否完成生长周期的关键时期[41]ꎬ对高粱群体结构和数量起着决定性作用ꎮ高粱萌发期抗旱性鉴定多采用聚乙二醇(PEG)㊁葡萄糖溶液等模拟干旱胁迫环境进行ꎬ通过种子发芽率㊁萌发抗旱指数等反映高粱的抗旱性ꎮ其中PEG-6000是目前被广泛应用的鉴定萌发期抗旱性较为理想的溶液ꎮ陈冰嬬等[41]使用15份保持系㊁18份恢复系和8份杂交种ꎬ通过PEG-6000水溶液模拟干旱胁迫环境ꎬ筛选出1份恢复系和1份保持系萌发期抗旱性亲本材料ꎻ通过抗旱性因子分析ꎬ认为萌发抗旱指数㊁根长和剩余干物质量可以作为高粱萌发期抗旱性筛选的鉴定指标ꎮ候文慧等[42]利用15%的聚乙二醇溶液进行干旱胁迫处理ꎬ采用隶属函数分析方法对8个饲用高粱萌发期抗旱性进行排序ꎬ得出SU9002为抗旱性最强的材料ꎬBJ0602为抗旱性最为敏感的材料ꎻ并利用主成分和聚类分析方法ꎬ对萌发期5个抗旱指标进行分析ꎬ结果表明ꎬ发芽指数和发芽率可以作为饲用高粱萌发期抗旱性评价的指标ꎮ采用聚乙二醇等高渗溶液不仅方法简单ꎬ而且排除了外界环境的干扰ꎬ可以获得更加准确的数据ꎬ有效地缩短了鉴定周期ꎬ提高鉴定效率ꎮ苗期是高粱整个生长发育阶段的关键时期之一ꎬ其生长好与坏直接影响着最终的产量和品质ꎬ因此ꎬ苗期抗旱性鉴定尤为重要ꎮ高粱苗期抗旱性鉴定方法可以分为三种ꎮ第一种较为常见的是使用PEG-6000溶液模拟干旱胁迫环境ꎮ赵晓倩[43]采用25%PEG-6000对259份高粱品种进行干旱胁迫处理ꎬ筛选出极抗旱品种14份㊁极敏感品种33份ꎬ并通过主成分分析方法对9个指标进行分析ꎬ结果表明ꎬ苗高㊁成活率㊁根冠比㊁根长和根鲜重可以作为评价高粱苗期抗旱性的指标ꎮ第二种是干旱复水法ꎬ是指在干旱胁迫后进行复水处理ꎬ用复水后的恢复能力指标评价高粱抗旱性ꎮ刘婷婷等[44]利用盆栽控水法对8个高粱品种幼苗进行干旱复水处理ꎬ通过研究生物量㊁水势㊁渗透式㊁光合参数等生理指标的变化情况来分析不同高粱品种的抗旱能力以及干旱适应能力和旱后复水恢复能力的关系ꎬ分析鉴定出了一份抗旱性强的品种辽杂21和旱后复水能力强的品种Moench.cv.Gadambaliaꎮ干旱胁迫时维持较高的叶片净光合速率和相对含水量有助于其提高干旱复水能力ꎬ因此ꎬ叶片净光合速率和相对含水量可以作为筛选高粱苗期抗旱性的生理指标ꎮ第三种是反复干旱法ꎬ是指通过高粱苗期连续两次干旱胁迫控水ꎬ以材料存活率为评价指标的一种鉴定方法ꎬ适用于大批量的品种鉴定ꎮ李舒凡[45]通过反复干旱法ꎬ对200份高粱品种进行苗期耐旱性鉴定ꎬ将叶片与根系的长势作为抗旱性的评价指标ꎬ能够从存活的质量上区别品种的抗旱性差异ꎬ进一步提高了筛选抗旱性品种的准确性ꎮ对高粱苗期抗旱性的鉴定只能反映出营养生长阶段的情况ꎬ需要结合生殖生长阶段的抗旱性ꎬ对不同品种抗旱能力进行综合评价ꎮ作物全生育期抗旱性鉴定对于抗旱新品种的选育㊁抗旱机制的研究以及抗旱基因的挖掘有着重要意义ꎬ共有两种鉴定方式ꎮ一种是通过人工761㊀第1期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀王晓东ꎬ等:高粱抗旱性研究进展控制水分和环境ꎬ通过干旱棚㊁人工气候箱等模拟干旱环境ꎬ研究各个生育期干旱胁迫对籽粒产量和品质的影响ꎮ汪灿等[46]通过在干旱棚内设置两个处理对50份酒用高粱材料进行成株期抗旱性鉴定ꎬ测定了成株期8个农艺性状ꎬ筛选出两个成株期酒用高粱抗旱性强的品种粱丰141-3和粱丰247-3ꎻ通过主成分㊁灰色关联度分析ꎬ认为分蘖数㊁穗粒数和单株粒重可作为酒用糯高粱资源成株期抗旱性评价指标ꎮ另一种是自然环境法ꎬ设置干旱和水地两个处理ꎬ操作简便ꎬ没有设备的要求ꎬ测定的结果更具说服力ꎬ但受环境因素影响较大ꎬ需要多年的试验数据进行支撑ꎮ袁闯等[47]采用自然环境法ꎬ设置灌水和干旱两个处理ꎬ通过测定高粱成熟期株高㊁穗重㊁千粒重㊁产量等10个性状ꎬ对22份不同品系的甜高粱进行成熟期耐旱性鉴定ꎬ筛选出3份抗旱品种和2份抗旱敏感性品种ꎻ通过主成分分析和逐步线性回归分析ꎬ认为千粒重㊁单株粒重㊁穗粒数和穗茎粗可以作为甜高粱成熟期抗旱性的评价指标ꎮ现阶段ꎬ高粱各抗旱指标评价鉴定基本都是局限于某一个时期ꎬ因此ꎬ需要综合高粱生长发育每个时期的指标来进行综合分析ꎬ建立综合指标评价体系ꎬ以提高高粱品种抗旱性鉴定的可靠性和真实性ꎮ4㊀高粱抗旱性分子生物学研究现阶段ꎬ国内外对于高粱抗旱性鉴定㊁抗旱生理生化以及干旱对农艺性状影响的研究已趋于完善ꎬ并且对于以基因为基础的转基因和分子标记技术也广泛应用到抗旱性分子遗传研究领域ꎬ通过转录组分析㊁QTL定位和全基因组关联分析(GWAS)构建分子遗传图谱ꎬ挖掘抗旱相关基因是高粱抗旱性分子遗传研究的发展方向ꎮ4.1㊀转录组分析转录组分析对于研究未知基因功能和特定调节基因的作用机制起着关键作用[48]ꎮ近年来新一代的转录组测序技术(RNA-seq)应运而生ꎬ它可以研究作物在干旱胁迫下的基因表达模式㊁分析抗旱分子机制㊁确定候选基因并进行功能注释[49]ꎮDugas等[50]通过渗透胁迫和脱落酸对高粱植物的转录组进行了分析ꎬ利用转录组测序技术揭示高粱的抗旱机制和基因筛选ꎮZhang等[51]使用转录组测序方法对干旱胁迫下高粱的叶和根进行转录组分析ꎬ鉴定出了差异表达基因ꎬ通过富集(GO)分析出耐旱性相关转录因子ꎮ王志恒等[52]用PEG-6000对甜高粱进行干旱胁迫ꎬ对高粱幼苗进行转录组测序分析并建立包含cDNA的文库ꎬ对差异表达基因进行GO富集分析和KEGG分析ꎬ发现有两个代谢通路与干旱胁迫响应相关ꎬ这两个通路都属于遗传信息代谢通路ꎮ表明甜高粱通过激活与干旱胁迫相关的蛋白表达和与碳水化合物相关的基因表达而增强渗透调节能力来响应干旱胁迫ꎮXu等[53]对两个抗旱性不同的高粱品种进行转录组分析ꎬ运用转录组测序技术确定了候选基因并进行了基因功能注释ꎬ分析了代谢通路ꎮ转录组测序技术促进了基因功能和表达水平的研究ꎬ通过分析干旱胁迫下基因的表达网络和富集通路ꎬ挖掘相关的新基因ꎬ可为今后进一步揭示干旱胁迫调节机制提供理论支撑ꎮ4.2㊀抗旱基因QTL定位植物的抗旱性是受多基因控制的数量性状ꎬ遗传复杂ꎮ干旱对作物的影响程度变化较大ꎬ常规育种方法费时㊁费力ꎬ难以选育优质的抗旱品种ꎮ随着分子生物学的发展ꎬQTL分析被广泛应用到分子遗传领域ꎮ赵辉[54]利用籽粒高粱654和甜高粱LTR108组成244个RIL群体ꎬ并构建了分子遗传连锁图谱ꎬ利用QTL定位分析耐旱性相关性状ꎬ分别在1㊁4㊁6㊁7染色体上检测出3㊁1㊁1㊁3个与抗旱系数相关的QTLs位点ꎬ并且在LG-1㊁LG-6㊁LG-7上定位到5个影响株高的QTLsꎮHaussmann等[55]用IS9830和N13与E36-1分别构建226个RIL群体ꎬ通过构建遗传图谱ꎬ发现标记分别位于10个连锁群和12连锁群中ꎬ利用复合区间作图检测到的3个性状的QTL数量在5个到8个之间ꎬ解释了31%和42%的遗传变异ꎮSakhi等[56]对107份孕穗期的高粱材料进行干旱胁迫处理ꎬ使用10条染色体上98个SSR标记位点的基因型数据对23对性状进行关联分析ꎬ鉴定出9个QTL与8个抗旱性状相关ꎮ持绿性是高粱干旱胁迫耐受性的一个组成部分ꎮSukumaran等[57]对Tx436(非持绿性)和00MN7645(持绿性)构建重组自交系进行遗传定位ꎬ利用全基因组单标记扫描和复合区间影射互补方法ꎬ检测到了15个与抗旱性状相关的QTLꎻ在1号染色体上发现了籽粒产量QTLꎬ解释了8%~16%的表型变异ꎬ861山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第56卷㊀在第2㊁6㊁9号染色体上发现了开花时间QTLꎬ解释了6%~11%的表型变异ꎬ在3㊁4号染色体上发现了持绿性QTLꎬ解释了8%~24%的表型变异ꎮ有关高粱QTL定位的研究可为后续高粱抗旱基因的精细定位㊁挖掘抗旱性相关基因和分析抗旱性机理奠定基础ꎮ4.3㊀全基因组关联分析(GWAS)GWAS是对多个个体在全基因组范围内进行遗传标记多态性检测ꎬ将基因型和表型进行关联并应用到寻找遗传图谱和挖掘性状相关候选基因的一种方法ꎮ近年来ꎬ为解析高粱抗旱性的遗传基础ꎬXin等[58]研究354份甜高粱在两种不同干旱处理下的株高性状ꎬ并将基于株高的平均生产力㊁干旱指数和胁迫耐受指数作为表型数据ꎬ结合甜高粱再测序获得的6186个SNPsꎬ使用三种不同的数量性状遗传模型进行GWAS分析ꎬ结果表明ꎬ在GLM㊁MLM和FarmCPU下分别检测到49㊁5个和25个耐旱相关的遗传位点ꎬ发现2个耐干旱的候选基因ꎬ其中ꎬSb08g019720.1基因与Athali ̄anaEFMTF基因同源ꎬ而Sb01g037050.1基因与玉米bZIPTF基因同源ꎮ高奇[59]用401份甜高粱材料进行干旱胁迫处理ꎬ并通过三个与干旱相关的性状筛选出耐旱评价指标并作为表型数据ꎬ利用高粱全基因组SNP标记ꎬ对三个性状耐旱指数进行全基因组关联分析ꎬ检测到两个株高性状基因可能是耐旱候选基因ꎮ赵晓倩[43]对259份高粱的7个苗期耐旱性相关性状进行全基因组关联分析ꎬ检测到102个显著的SNP位点ꎬ筛选出7个抗旱候选基因ꎮ通过全基因组关联分析揭示耐旱候选基因可为后续基因功能验证和高粱耐旱分子机制研究奠定基础ꎮ5㊀提高高粱抗旱性的方法5.1㊀传统育种方法传统的育种方法包括杂交育种㊁回交育种㊁系统选育㊁混合选育等ꎬ其中较为常见的是杂交育种ꎮ杨伟等[60]通过母本不育系7501A和父本恢复系RHMC386进行组配杂交ꎬ选育出优质抗旱高粱新品种潞杂9号ꎮ杨婷婷等[61]研究发现ꎬ以不育系SX605A为母本㊁以恢复系SX870为父本杂交育成高粱品种晋杂31号ꎮ其选育过程中ꎬ亲本都是通过杂交再连续多代自交得到的稳定品种ꎬ都具有很强的抗旱性ꎬ通过该方法可以提高选育品种的抗旱性ꎮ李继洪等[62]同样用不育系亲凡A为母本㊁以恢复系苏丹草黑壳3号为父本杂交选育出抗性强的品种吉草3号ꎮ由此可见ꎬ选育抗旱性强的不育系和恢复系是提高高粱杂交种耐旱性的重要途径ꎮ5.2㊀施加外源物质通过对高粱施加外源营养元素㊁生长调节剂以及进行种子引发等都可以提高其抗旱性ꎮAhmed等[63]发现硅营养对高粱的生长和生理参数有显著影响ꎬ通过在干旱胁迫条件下对高粱进行施加硅营养处理ꎬ可以提高耐旱基因型品种(系)的叶片水势㊁叶面积指数㊁蒸腾速率和SPAD值ꎬ同时在硅处理下净同化和相对生长量表现出最大值ꎮ张瑞栋等[64]分别用聚乙二醇(PEG)㊁KCl㊁CaCl2和水杨酸(SA)对高粱种子进行引发处理ꎬ显示其可以促进干旱胁迫下种子萌发率ꎬ促进胚根和胚芽的伸长ꎮ其原因可能是引发处理提高了胚芽内抗氧化酶活性ꎬ同时促进糖代谢ꎬ增加脯氨酸含量ꎬ解决了干旱胁迫下发芽率低㊁胚根胚芽生长受抑制的问题ꎬ进而提高高粱萌发期的抗旱性ꎮTounekti等[65]的研究也得到一致的结果ꎮKamali等[66]研究发现ꎬ使用固氮菌和丛枝菌根真菌(AMF)的高粱比不使用的受干旱胁迫程度较轻ꎬ固氮菌和丛枝菌根真菌可以减少高粱电解质渗漏和丙二醛含量ꎬ通过提高花青素㊁类胡萝卜素㊁黄酮㊁生长素(IAA)等物质含量和抗氧化酶活性来缓解干旱胁迫的影响ꎮKamali等[67]同时也发现细菌和丛枝菌根真菌也可以通过增加光合色素㊁可溶性蛋白等物质含量提高高粱渗透调节能力ꎬ继而应对干旱胁迫环境ꎮShehab等[68]研究发现ꎬ脱落酸(ABA)和茉莉酸甲酯(MeJA)可以减轻干旱胁迫引起的负面效应ꎬ降低干旱胁迫下高粱中氰化氢(HCN)的含量ꎮ植物生长调节剂(PGRs)改善高粱抗旱性归因于可溶性蛋白㊁丙二醛㊁活性氧㊁过氧化氢等的积累减少ꎬ光合参数的改善以及抗氧化酶活性的变化ꎬ进而提高其抗旱能力ꎬ特别在甜高粱中尤为明显ꎮ5.3㊀转基因方法植物抗旱性是受多基因控制的数量性状ꎬ其中包括参与调控植物活性氧㊁可溶性糖㊁抗氧化酶㊁叶绿素和ABA信号转导等生理生化过程的基961㊀第1期㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀王晓东ꎬ等:高粱抗旱性研究进展。
孔雀草的研究进展

孔雀草的研究进展霍雅楠;王文;祁智;毛惠平【摘要】孔雀草菊科万寿菊属一年生草本植物。
由于其优良的观赏特性,在园林绿化中应用广泛。
孔雀草一般大量应用于花坛、花境和庭院。
文章总结了孔雀草在遗传育种、栽培、抗逆性、化感及应用等方面的研究进展,为孔雀草的进一步研究提供参考,并对其未来的研究提出了展望。
【期刊名称】《北方农业学报》【年(卷),期】2017(045)004【总页数】4页(P123-126)【关键词】孔雀草;遗传;育种;栽培;抗逆性;化感作用【作者】霍雅楠;王文;祁智;毛惠平【作者单位】[1]内蒙古大学生命科学院,内蒙古呼和浩特010010;[2]呼和浩特市园林管理局,内蒙古呼和浩特010010;[1]内蒙古大学生命科学院,内蒙古呼和浩特010010;[1]内蒙古大学生命科学院,内蒙古呼和浩特010010;【正文语种】中文【中图分类】S681.9孔雀草(Tagetes patura L.)菊科万寿菊属一年生草本植物。
别名法兰西菊、小万寿菊,原产于北美洲墨西哥,现广泛分布于世界各地。
株高20~100 cm,茎直立,近基部分枝,分枝斜展开。
叶对生或互生,羽状全裂,头状花序单生,径约4cm。
舌状花金黄色或橙色,基部带有红褐斑[1]。
花较万寿菊小而多,花期5—9月。
喜阳光,但在半荫处栽植也能开花。
对土壤要求不严,耐移植,生长迅速,栽培管理简单[2]。
孔雀草园林用途广泛,普遍应用于花坛花镜庭院。
此外,孔雀草还是良好的中药、驱虫药、杀虫剂和着色剂。
由于目前孔雀草广泛应用于园林景观设计,因此,对孔雀草的研究也主要集中在遗传育种、栽培技术、抗逆性及化感作用等方面。
为了更好地开展对孔雀草的研究和开发利用,现将有关孔雀草的研究情况与应用进展综述如下。
1 孔雀草的遗传特性为了了解孔雀草种质间的亲缘关系,付巧娟等[3]通过18个形态特征对40份孔雀草种质进行了遗传变异分析、主成分分析和聚类分析。
结果表明,孔雀草具有丰富多样的表型,种质间表型性状变异程度高。
百里香属(ThymusL.)植物的分类与抗旱性研究的开题报告

百里香属(ThymusL.)植物的分类与抗旱性研究的
开题报告
一、研究背景和意义
百里香属(ThymusL.)是一种常见的草本植物,其具有香气浓郁的香味和丰富的药用价值,被广泛应用于调味、保健和药物等领域。
在我国,百里香属植物主要分布于北方干旱半干旱区域,因其具有较强的抗旱性,在干旱气候条件下仍能生长繁殖,成为了荒漠化地区进行生态修复、绿化建设的理想材料。
因此,对百里香属植物的分类和抗旱性研究具有重要的理论和应用价值。
二、研究目的
本次研究的目的是对百里香属植物进行系统分类和抗旱性分析,具体包括以下几个方面内容:
1.对百里香属植物进行形态学和生物学特征的观察和记录,结合现有分类标准进行分类鉴定;
2.利用生理生化测试方法,评估百里香属植物的抗旱性能力,比较不同品种或种间的抗旱性差异;
3.探究百里香属植物在缺水胁迫下的生理响应机制,深入了解其抗旱适应策略;
4.通过实验研究,探索提高百里香属植物抗旱性的途径,为干旱地区的生态修复和绿化建设提供新的思路和方法。
三、预期成果
本次研究的预期成果主要包括:
1.明确百里香属植物的系统分类和种间区别,为其更好的利用和保护提供科学依据;
2.深入了解百里香属植物的抗旱机制和适应策略,为荒漠化地区进行生态修复和绿化建设提供理论基础;
3.发现并研究提高百里香属植物抗旱性的途径,为农业生产和环境保护提供技术支持和指导。
几种野生菊科植物含油量和抗旱性初探

不显著, 蒙山葛首的细胞膜透性最弱, 苦卖菜细胞膜透性最强; 种植物的含油量则呈相反的趋势, 3 蒙山 葛筐 叶片的含油量最高, 且与苦卖菜 和蒲公英的粗脂肪含量差异显著。 故3种植物中蒙山葛筐是在北方干 旱地区发展成为野生能源植物栽培的最具潜力的对象。 关键词: 野生菊科; 含油量; 杭旱性; 能源植物
天津农学院青年重点科技项目。 作者简介: 李建科 ( 197 一) ,男, 3缓慢, 但是抗旱性较强。
南
方
农
业
第1卷第2期
Vol. I No .2
2007 年3 月
M ar. 200 7
South Chi na Agr icul tur e
工 、 }
! r .
n “
J 惑 胭 即 黔 戳
蒙山葛芭
蒲公英
苦卖菜
图3 3 种野生菊科植物粗脂肪含量的比较
从图3能看出, 3种植物叶片中, 蒙山葛首的平均粗 脂肪含量最高, 平均为2 .46%, 1 其次是蒲公英, 最小是 苦卖菜, 平均只有1 8%。 0 显然, 3种材料的含油量是蒙 山葛首> 蒲公英> 苦卖菜。 经差异显著性分析, 5 %和9 %的置信度区间内, 在9 9 蒲公英、 苦卖菜的平均粗脂肪含量差异性不显著, 而蒙 蒙山葛芭 蒲公英 苦卖菜 山葛首与蒲公英、 苦卖菜的平均粗脂肪含量差异性极显 著。 总的来说, 蒙山葛芭的粗脂肪含量最高, 且与苦卖 菜和蒲公英的平均粗脂肪含量差异极显著。
全球性的干旱日 趋明显, 水资源短缺继续加剧, 土
壤有效水含量逐年减少1 2 水分不足是限制农业发展的 , 1 ]。 重要因子, 水资源短缺以及土壤盐渍化是目 前制约农业 生产的一个全球性问题。 在我国, 0 1年华北、 仅20 西北
干旱胁迫下四种矿区生态修复植物的抗旱性研究

摘
要: 以八 宝景天 、 沙打 旺、 紫花苜蓿和早熟禾 4种典 型干旱 区草本植物 为试材 , 采 用盆栽
控 水法进行 干旱胁迫 , 测定 了 4种植物 叶片相 对含水量 、 相对 电导率、 叶绿素含量 、 脯氨酸( P r o ) 含 量、 丙二醛( MD A ) 含量和可溶性糖含 量 6个抗旱指标 的变化 ; 运用隶属 函数值 法进行植 物抗旱性 综合评价 , 并采用灰 色关联度 法对 6 项抗旱指标与植物抗旱 性的关联度进行 分析 , 以研 究不 同植
物在干旱胁迫下的抗旱机制 。结果表明 : 4 种 植物 综合 抗旱性 顺序 为: 八 宝景 天>早 熟禾> 紫花 苜蓿> 沙打 旺; 各项抗旱指标与植 物抗 旱隶属 函数均值 关联度 大小顺序为 : 叶绿 素含 量( 0 . 9 7 2 7 ) 、
叶片相对含 水量 ( O . 9 3 8 7 ) 、 可溶性糖含量 ( O . 8 3 1 4 ) 、 丙二醛含 量( O . 7 9 4 0 ) 、 相对 电导率( O . 6 3 2 0 ) 、 脯 氨酸含量 ( 0 . 4 5 2 8 ) , 即叶绿素含量 、 叶片相 对含 水量和 可溶性糖含量对干旱环境最为敏 感 , 可作为
随着人类对矿产资源需求 的增加 , 各种 开挖 行为导 致大规模的土地和植被破坏 以及严 重 的水 土流失 , 极大 地破坏 了原有的生态 系统[ 1 _ 2 ] 。如何使废 弃矿 山恢复植
被, 已成为生态环境建设 中亟需 解决 的问题 。缺水 和植
属 函数值法对植物进行抗旱性评 价 , 具 有科学性和可靠 性, 利用灰 色关 联度 分 析法 进行 抗 旱 指标 间 关联 性分 析, 避免 主观差异 , 具有可行性和准确性_ l 】 , 以期为矿区 生态修 复抗旱植物选择提供理论基础 和技术依据 。
香根草种质资源抗旱性的变异研究

任 编 辑
1 材料与方 法
11 供 试 材 料 .
收 稿 日期 :2 0 — 2 2 0 80 — 6 基 金项 目 :国 家科 技 部 攻 关 项 目 (0 4 A 2 B 7 4 、广 东省 20B 52 0—) 科 技 攻 关 项 目 (0 5 2 2 12 ) 及 珠 海 市 科 技 计 划 项 目 20 1 0 0 0 0 3
地位 。本试验 对来 自国内外 不 同 国家 和地 区香根草 种
质资源 耐旱性 能 的变异情况 进行测 定 和分析 ,力 图筛 选 出抗 旱 陛能 比较好 的生 态型 品系应 用于具 体 的生 产 实践 ,也为将来 我 国进一步 培育具 有 自主知 识产权 的
优 良香根 草品种提供 原始 材料 。
重 要 的现 实 意 义 。香 根 草 ( ei r i no e) 又 V t ei z i d s v a a i
所在实验 室对香 根草 种质 资源 的分 布 区域 、物候
特 征 、生态特 点 以及 生 物学特性 等进行 了比较详 实的
调查 和研 究 ,同时搜 集 国 内外香 根草 种质 资源 ,建 立 了香 根 草种质 资源 圃 。供 试草 种为来 自该种 质资 源 圃 不 同生 态型 的香根 草 。各供 试香 根草生 态 型的名称及
用 D S 3 7型 电 导 率仪 ( 海 虹 益 仪 器 有 限公 司) D 一0 上
通 讯作 者
维普资讯
业 畜 2年4 第9 与 牧 0第 0 期总1 8 4 期
C0EUUU AYY×M
可溶性糖 含量 用蒽 酮 比色法 进行测 定 。 1 抗旱性 的综合评 定文条件 ,是 一种非 常优 秀的 水土保 持和 环境恢 复植物 , 目前 已经 广泛地 应用 在我 国南 方地 区的水土保 持 与国土绿 化 中。在香 根草 的推
紫花苜蓿对干旱胁迫适应性的研究进展

紫花苜蓿对干旱胁迫适应性的研究进展韩德梁1,2,王彦荣1(1.兰州大学草地农业科技学院甘肃草原生态研究所,甘肃兰州730020;2.贵州大学生命科学学院,贵州贵阳550025)摘要:苜蓿是抗旱且需水较多的豆科牧草,水分供应不足会使其受到干旱胁迫的危害,从而导致农艺性状,如草产量、种子产量、叶片形态和根系特征等,均发生改变。
同时,苜蓿体内也发生一系列生理生化反应来消除或降低干旱胁迫的伤害作用,即阻止、减少或抵消干旱胁迫诱导的生理生化过程。
本研究综述和讨论了苜蓿的特征特性及其对干旱胁迫的响应、抗旱性评价和缓解干旱胁迫的途径等研究进展。
关键词:紫花苜蓿;干旱胁迫;适应性中图分类号:S551.703.4 文献标识码:A 文章编号:100425759(2005)06200072073 紫花苜蓿(Medicago sativ a )简称苜蓿,是世界分布最广的豆科牧草,也是我国种植面积最大的人工牧草,达210万hm 2,在世界苜蓿生产大国中居第5位[1]。
目前,人们已充分认识到紫花苜蓿不仅在发展畜牧业生产、建植人工草地方面有着显著的经济效益,而且在建设和保护草地生态环境,绿化、美化生活环境等方面同样具有潜在的经济效益和重要的社会效益。
干旱是影响牧草分布及产量的重要因素之一,已成为限制苜蓿大面积扩展的重要原因。
世界上约有1/3的地区属于干旱和半干旱区,我国干旱半干旱地区约占国土面积的1/2[2]。
旱作条件下,牧草生长所需的水分主要依靠自然降水,即使有灌溉条件的地区,也往往由于灌水量不足或不及时等原因使牧草受到干旱的威胁。
干旱对牧草造成的损失在所有的非生物胁迫中占首位,仅次于生物胁迫病虫害造成的损失[3]。
在我国实施“退耕还草”等一系列生态环境建设的良好机遇下,大力发展“朝阳产业”中素有“牧草之王”的苜蓿新产业,无疑具有广阔的前景。
因此,深入开展苜蓿抗旱性研究,科学选育适宜广大干旱、半干旱地区种植的优良苜蓿品种,对于水资源的合理利用和生态环境的改善均有重要意义。
西藏地区披碱草属植物研究进展

西藏地区披碱草属植物研究进展作者:唐希明来源:《南方农业·中旬》2019年第11期摘要披碱草属牧草是西藏地区天然草地重要的牧草品种之一。
基于此,综述了西藏地区披碱草属植物的种质资源、生理生化特征及其在牧草生产和生态恢复方面的应用,并提出了未来西藏地区披碱草属植物的研究重点和研究方向。
关键词披碱草属;人工草场;植被恢复;乡土物种;西藏自治区中图分类号:F302.1 文献标志码:B DOI:10.19415/ki.1673-890x.2019.32.067披碱草属(Elymus L.)植物是禾本科(Poaceae)多年生草本植物,分布范围广泛,从海滩到海拔5 200 m以上的喜马拉雅山区均有分布[1-2]。
披碱草属植物中多数物种是高寒草原和草甸的建群种和优势种,是维持高寒地区生态平衡的重要植物种群之一,具有较高的饲用价值。
同时,它还是麦类作物的野生近缘种遗传资源,具有麦类作物缺乏的优质、高产、抗虫、抗病和抗逆等基因,是丰富麦类作物和牧草遗传多样性的基因资源库。
此外,披碱草属植物在防风固沙、水土保持等方面也发挥着重要的作用[3-4]。
西藏天然草地野生牧草种质资源非常丰富,有饲用植物2 672种,占西藏地区草地植物种总数量的84.26%[1]。
因此,综述西藏地区披碱草属植物的研究现状,是开展种植资源保护和开发的重要前提,对于提高与优化西藏地区人工草场管理和退化草地生态系统的恢复具有重要的参考价值。
1 西藏地区披碱草属植物种质资源概述在我国,披碱草属植物有12种和1个变种,西藏地区分布的披碱草属植物有5种,分别为披碱草(Elymus dahuricus)、老芒麦(E. sibiricus)、垂穗披碱草(E. nutans Griseb)、黑紫披碱草(E. atratus)和麦宾草(E. tangutorum),广泛分布在西藏各地,是西藏高寒草原中具有代表性的草种[2,5]。
2 西藏地区披碱草属植物生理生化方面的研究2.1 披碱草属植物生物学特性研究2.1.1 种子萌发特性秦爱琼等[6]在野外试验条件下,对巴青县、索县和当雄县采集的6个野生披碱草类群进行研究发现,雄披碱草种子萌发率最高为90.3%,可作为野生资源进行驯化。
4种地被植物的抗旱性评价

随着 世界 环境 的 不断 恶化 , 美化环 境 越 来越 受 到关 注 , 地被 植物 作 为美化 环境 的 组成 部 分成 为人 们研 究 的~ 个 重 要 内容 , 资源匮 乏引起 的地被 植物 灌溉 也因此 受到广泛 关 水
注 , 找和 使 用 抗旱 植 被 成 为 园林 绿化 工 作 的重 点 , 此 , 寻 因
wekw s e u se tbl> e u y r u O ho o oo jp nc s He eo al ^删 a a d m ca i S d m bi m> p ip gg n a o iu > m rcls l S p e d i K y o d g u d oe a td u r i a c ; m rhn i a a o e r s r n c vr l ;r  ̄t s t e c pe e s e vl t n w o pn o e sn o ve u i
多年生宿根草本 , 茎圆柱形 、 粗壮 , 生或对 生, 叶轮 具波状齿
多 年 生常 绿 草 本 , 直 立 , 对 生 或 3 4枚 轮 生 , 伞 花序 顶 生 茎 叶 - 聚 根茎细长 , 叶丛生 , 花茎自叶 丛中抽 出, 直立 , 总状花序 , 顶生 o ho ooo jp nc s 多年生草本植物 , e i ggn ao i p u 多年 生 草 本 植 物 , 下 具 根 状 茎 和 肉 质 肥大 的纺 锤 状 块 根 , 基 生 螺 旋状 聚 伞 花 序 地 叶 He e o l ,擅 Ⅱ m rc i j s
S du s e a l e m pe tbi e S du e m rd m iu
111 供 试植 物 。 地被植 物 ( 1 。 .. 4种 表 ) 11 仪 器 设 备 。 分 速 测 仪 、 土盒 、 箱 、 分 之 一 天 .. 2 水 取 烘 干
西藏地区高原林木抗寒抗旱性研究进展

㊀2020年6月J o u r n a l o fG r e e nS c i e n c e a n dT e c h n o l o g y第12期收稿日期:2020G05G21作者简介:黄㊀勇(1982-),男,工程师,研究方向为林业工程.西藏地区高原林木抗寒抗旱性研究进展黄勇(西藏自治区那曲市林业和草原局,西藏那曲852000)摘要:指出了西藏地区具有独特的自然环境特征,海拔较高,温度较地,对当地林木种植起着重要主导作用.介绍了西藏地区独特的自然气候特征以及现有的树种种植情况,并针对该地林木抗旱性,主要从生物膜系统㊁渗透调节物质以及抗氧化酶层面进行了探讨,对高原地区林木种植提供一定参考.关键词:西藏地区;高原林木;抗寒性;生物膜系统;抗氧化酶中图分类号:Q 948.117㊀㊀㊀㊀㊀㊀文献标识码:A㊀文章编号:1674G9944(2020)12G0036G021㊀引言西藏高原地区海拔高于4000m ,具有湖泊㊁高山㊁冰川等多种地貌,日照时间较长㊁太阳光照较为强烈,昼夜之间具有较大的温差,这一独特气候对西藏地区渗透调节物质性[1].2㊀西藏地区林木种植情况西藏地区四季ȡ0ħ积温为800~1300ħ,平均日照比率为52%~67%,具有6000M J /m 2太阳辐射能量,5~9月为降雨集中地区,降雨量达到全年80%,林木ȡ10ħ最适生长积温为16~60ħ,目前种植有30余种木本植物,已经成活20余种[2],见表1.表1㊀目前西藏地区林木树种种植情况林木名称学名科名生长习性小叶硬叶柳S a l i x s c l e r o p h y l l a v a r .t i b e t i c a 杨柳科落叶灌木墨竹藏川杨P o pu l u s s z e c h u a n i c a v a r .t i b e t i c a M o Z h u 杨柳科落叶乔木小叶金露梅P o t e n t i l l a p a r v i fo l i a 蔷薇科落叶灌木银白杨P o p u l u s a l b a 杨柳科落叶乔木左旋柳S a l i x p a r a p l e s i a 杨柳科落叶乔木青海云杉P i c e a c r a s s i fo l i a 松科常绿乔木江孜沙棘H i p p o ph a e r h a m n o i d e s s u b s p .g ya n t s e n s i s 胡颓子科落叶灌木粗刺锦鸡儿C a r a g a n a c r a s s i s p i n a 豆科落叶灌木墨竹柳A l i x m a i z h o k u n g g a r e n s i s 杨柳科常绿乔木变色锦鸡儿C a r a g a n a v e r s i c o l o r 豆科落叶灌木近硬叶柳S a l i x s c l e r o p h yl l o i d e s 杨柳科落叶灌木目前西藏地区林木种植类型包括杨柳科㊁蔷薇科㊁松科㊁豆科等,所种植的林木包括墨竹藏川杨㊁银白杨㊁江孜沙棘㊁小叶硬叶柳㊁小叶金露梅㊁墨竹柳㊁墨竹藏川杨等,一般分布在西藏地区中海拔相对较低㊁环境相对较好㊁温湿较小的地区,与当地气候具有一定联系,对该地林木抗寒性研究具有重要意义[3].3㊀西藏地区林木抗旱性研究在西藏地区独特的高原环境之下,该地植被多为多年生草本植物,由此一般针对灌木与草本植物进行抗旱性研究,其中研究较为细致的林木类型为披碱草属(E l ym u s )㊁早熟禾属(P o a )禾本科植物类型[4].该树种是西藏地区重要的牧草来源之一,在3000m 以上地带较为常见,属于高寒草甸中的优势种植树种[5].西藏地区植物林木植物抗旱性研究过程中与其生理生化关系层面的研究具有一定联系,林木抗旱性研究与渗透调节物质之间具有一定联系,在现代分子生物学技术快速发展背景下,人们逐渐从分子生物学领域层面进行林木抗旱性研究[6].3.1㊀植物抗寒性与膜系统之间的关系林木冻害容易发生的部位在质膜部位,在细胞膜上会发生大量的生理生化反应,通过电导仪法的运用能够看到,冻害现象能够增加植物细胞膜的透性,由此会使得细胞中出现可溶性物质大量外渗现象,不利于植物代谢.西藏高原地区垂穗披碱草㊁矮嵩草㊁苔草3种植物生长过程中低温半致死温度(L T 50)会随着气温的变化而产生相应变化,三者抗寒能力排序依次为垂穗披碱草㊁矮嵩草㊁苔草.在温度降低与胁迫时间增长的情况下,梭罗草㊁早熟禾㊁垂穗披碱草质膜透性也会随着而增长,其中抗寒能力最强的植物是青海草地早熟禾,其胁迫时间增长幅度最小,细胞膜呈较为稳定的状态[7].林木膜损伤强度可以通过丙二醛(M D A )量的多少进行反映,随着质膜透性提升,植物体内M D A 含量也会随之提升,其中较为典型的植被类型为青藏高原黑褐苔草[8].3.2㊀林木抗寒性与抗氧化酶关系研究在较低的环境下,林木中自由基㊁活性氧利用率会下降,林木之中会积累较多的活性氧,破坏了林木中活性氧的产生与清除之间的平衡状态,不利于林木的自然生长,较为严重情况下甚至会导致林木的死亡.但是西藏地区林木在长期生长过程中,林木体内已经形成了一种抗氧化防御系统,为了减少较低温度对林木的不良影63㊀黄㊀勇:西藏地区高原林木抗寒抗旱性研究进展自然与生态响,植物体内会清除活性氧自由基[9].一般由过氧化物酶(P O D)㊁谷胱甘肽过氧化酶㊁超氧化物歧化酶(S O D)㊁过氧化氢酶(C A T)㊁谷胱甘肽还原酶共同构成酶防御系统,目前国内在抗氧化酶的研究中主要集中在C A T㊁S O D㊁P O D3个层面.在低温环境下,能够提升绵头雪莲花抗氧化酶活性,减少对幼苗的损害,避免或者减少由于低温胁迫而导致的膜脂过氧化现象,以此提升林木的抗旱性[10].西藏高原中,同样一种林木由于所处的海拔不同,植物体内抗氧化酶活性也具有一定差异,唐古特山莨菪林木生长过程中,低温环境会损害其细胞膜,在海拔增加的情况下,此种损害程度也在随之而增加.随着海拔的增加,莨菪叶片中C A T㊁S O D活性也会随之而提升[11].3.3㊀林木抗寒性与渗透物质调节之间的关系西藏地区林木渗透调节所依赖的重要物质为可溶性蛋白质㊁可溶性糖㊁游离脯氨酸.在林木受到低温胁迫情况下,以上渗透调节物质浓度会随之增加,细胞冰点也会随之而降低,以此使得植物细胞具有良好的保水能力,在较低的温度环境之下也能够生存.在低温胁迫时,田苜蓿种子㊁青藏扁蓿豆中脯氨酸和可溶性糖能够随之而增加,由此提升了青藏扁蓿豆抗寒性,可见青藏扁蓿豆具有较强的抗旱能力,可以在高寒地区进行推广种植[12].4㊀结论与展望本文对青藏高原地区进行了林木抗寒抗旱性研究,分析了西藏地区林木抗寒抗旱性与生物膜系统㊁渗透调节物质以及抗氧化酶之间的关系,并介绍该地已有的树种类型,西藏地区四季ȡ0ħ积温为800~1300ħ,平均日照比率为52%~67%,林木种植中具有30余种木本植物,已经成活20余种[2],包括杨柳科㊁蔷薇科㊁松科㊁豆科等,所种植的林木包括墨竹藏川杨㊁银白杨㊁江孜沙棘㊁小叶硬叶柳㊁小叶金露梅㊁墨竹柳㊁墨竹藏川杨等,一般分布在西藏地区中海拔相对较低㊁环境相对较好㊁温湿较小的地带[13].进行西藏地区林木抗旱性研究,林木抗旱性研究与渗透调节物质之间具有一定联系,从分子生物学领域层面进行林木抗旱性研究,研究植物抗寒性与膜系统之间的关系,电导仪法分析可知冻害现象能够增加植物细胞膜的透性,会使得细胞中出现可溶性物质大量外渗现象[14].研究林木抗寒性与抗氧化酶关系,低温环境下,林木中自由基㊁活性氧利用率会下降,林木之中会积累较多的活性氧,破坏了林木中活性氧的产生与清除之间的平衡状态,西藏地区林木体内形成了一种抗氧化防御系统,植物体内会清除活性氧自由基.研究林木抗寒性与渗透物质调节之间关系,西藏地区林木渗透调节所依赖物质包括可溶性蛋白质㊁可溶性糖㊁游离脯氨酸[15].在林木受到低温胁迫情况下,细胞冰点会降低,植物细胞具有良好的保水能力,使得在较低温度环境生存.应当利用本文的研究成果挖掘适合当地自然气候的树种,在保护当地自然环境的同时,还能够有效挖掘当地名贵的藏药材,通过树种研究开发当地林木种植中巨大的社会经济价值,以此达到良好的环境效益与社会效益.参考文献:[1]王小军,武紫娟.林木抗旱性机理研究进展[J].天津农业科学,2017,23(12):13~15.[2]路之娟,张永清,张㊀楚,等.不同基因型苦荞苗期抗旱性综合评价及指标筛选[J].中国农业科学,2017(17).[3]旺㊀加,陈㊀彪,尼㊀珍,等.西藏自治区高原地区350例急性胰腺炎回顾性临床研究[J].中华消化杂志,2019,39(1):50~52.[4]尼玛玉珍,德㊀央,史建玲,等.西藏不同海拔地区藏族人群高原性心脏病的超声心动图指标分析[J].中华医学超声杂志(电子版),2019,16(4):306~310.[5]王保海,翟㊀卿,张亚玲,等.青藏高原林木主要害虫发生与绿色防控研究[J].西南农业学报,2019,32(8):1805~1809.[6]曾俊霞.高原种苗造林和抚育技术要点浅析[J].农家科技(下旬刊),2018(7):204.[7]潘㊀昕,邱㊀权,李吉跃,等.基于叶片解剖结构对青藏高原25种灌木的抗旱性评价[J].华南农业大学学报,2015(2):61~68.[8]邱㊀权,潘㊀昕,李吉跃,等.青藏高原20种灌木抗旱形态和生理特征[J].植物生态学报,2014,38(6):562~575.[9]吕志刚.黄土高原抗旱造林技术分析及新技术的应用[J].北京农业,2014(36):109~109,110.[10]邱㊀权,潘㊀昕,李吉跃,等.青藏高原20种灌木生长时期根系特征及抗旱性初探[J].中南林业科技大学学报,2014(3):29~37.[11]黄瑞敏,潘㊀刚,周㊀晔,等.西藏核桃引种到内地后坚果品质变化研究[J].中国油脂,2019,44(5):150~154.[12]邓㊀磊,关晋宏,高万里,等.黄土区辽东栎群落细根生物量对物种多样性及气候的响应[J].西北林学院学报,2018,33(5):16~22.[13]祝友朋,韩长志.核桃细菌性黑斑病菌分泌蛋白的预测及生物信息学分析[C]//中国植物病理学会.2018年学术年会论文集.北京:中国农业科学技术出版社,2018.[14]黄艳丽,李占斌,苏㊀辉,等.人工林对黄土高原小流域上下游不同坡面土壤水分的影响[J].农业工程学报,2018,34(15):108~116.[15]张保刚.省林业工作站:举办高原藏区基层林业站技术扶贫第一期培训班[J].绿色天府,2018(6):26.73。
蒙草抗旱措施

蒙草抗旱措施引言干旱是一个严重影响农作物产量和生态系统健康的自然灾害。
在干旱地区,寻找有效的抗旱措施至关重要。
蒙草(Mongolian Leek)是一种生长于蒙古高原地区的耐旱植物,它具有出色的抗旱能力。
本文将介绍蒙草的一些关键特点及其在抗旱方面的应用措施。
蒙草介绍蒙草是一种多年生草本植物,属于百合科植物。
它的地下部分主要由块茎和根系组成,地上部分为长叶葱状,具有特殊的水分调节机制。
蒙草能够生长在干旱、高寒和贫瘠的土壤中,适应能力强,抗旱性能突出,因此被广泛应用于抗旱和生态恢复领域。
蒙草抗旱措施1. 水分调节机制蒙草具有特殊的水分调节机制,包括: - 厚而多层次的叶片:蒙草的叶片覆盖层非常丰富,形成一种“气候屏障”,减少水分的蒸发。
- 延缓蒸腾作用:蒙草的叶片表面有一层厚厚的枯皮,减缓叶片的蒸腾作用,降低水分的蒸发速率。
- 深入地下水层:蒙草的根系发达,能够深入地下水层,获取更多的地下水资源。
2. 土壤保水能力蒙草能够改善土壤的保水能力,主要包括: - 增加土壤有机质含量:蒙草的茎叶体积庞大,能够产生大量的有机质,提高土壤的保水能力。
- 促进土壤微生物活动:蒙草的根系分泌出一些物质,可以促进土壤中有益微生物的生长,提高土壤的保水能力。
- 遮荫保湿:蒙草茂密的叶片可以遮荫土壤表面,减少日照和风蚀,降低土壤水分的蒸发率。
3. 生态系统恢复蒙草不仅可以抵御干旱,还能促进生态系统的恢复,具体体现在以下几个方面:- 防风固沙:蒙草的根系可以有效地固定沙土,防止风沙的侵蚀。
- 改善土壤肥力:蒙草的有机质能够分解,形成肥沃的土壤,为其他植物的生长提供良好的土壤环境。
- 促进物种多样性:蒙草的生长可以为其他植物提供保护和遮荫,增加生态系统的物种多样性。
同时,蒙草还能为动物提供食物和栖息地。
结论蒙草作为一种具有出色抗旱能力的植物,是干旱地区抗旱措施的重要组成部分。
其特殊的水分调节机制和能够改善土壤保水能力的特点,使其成为一种理想的植物选择。
凤仙花种子的水分调节与抗旱性

凤仙花种子的水分调节与抗旱性凤仙花(Scientific name:Impatiens balsamina L. )是一种常见的观赏植物,以其艳丽的花朵和易种植的特性受到广泛喜爱。
凤仙花属于早熟草本植物,其种子在生长过程中对水分的调节和抗旱性具有重要意义。
本文将重点探讨凤仙花种子的水分调节机制和抗旱性方面的研究进展。
植物种子对于生存和繁殖的成功至关重要,其中一个重要的因素就是对水分的调节。
凤仙花种子具有良好的耐旱性,其水分调节机制主要表现在以下几个方面:首先,凤仙花种子具有较低的种子含水率。
研究发现,凤仙花种子在成熟之后,种子含水率仅为8-10%左右,远低于一些其他植物种子。
这种低含水率可以降低种子内部的水分活性,减少水分流失的风险,有效延长种子的保存寿命。
其次,凤仙花种子具有较高的抗脱水能力。
由于生活环境的不确定性,凤仙花种子的抗脱水能力成为其适应干旱条件的关键特征之一。
研究表明,凤仙花种子表皮细胞具有丰富的脂肪体,这些脂肪体可以在脱水过程中释放油脂来保持种子的稳定状态,降低水分流失。
此外,凤仙花种子还具有较强的吸湿能力。
在湿度较高的环境中,凤仙花种子表皮细胞可以吸收周围的水分,增加种子含水率,从而提高种子的萌发率和存活率。
凤仙花种子的抗旱性研究主要集中在以下几个方面:首先,凤仙花种子的抗旱性与种子贮藏蛋白相关。
研究发现,凤仙花种子中的贮藏蛋白在干旱条件下能够被分解,产生有利于种子保护和抵抗干旱的物质,从而提高种子的抗旱能力。
其次,凤仙花种子的抗旱性与内源激素水平调节相关。
激素在植物的逆境适应中发挥重要作用,研究发现,凤仙花种子中的激素含量在干旱条件下有所增加,特别是脱落酸(abscisic acid,ABA)的含量显著上升,这一过程可以促进种子进入休眠状态,减少水分流失,从而提高种子的抗旱性。
此外,一些环境因素也可以影响凤仙花种子的抗旱性。
例如,研究发现温度和光照条件对凤仙花种子的抗旱性有明显影响。
抗旱育种植物有哪些

抗旱育种植物有哪些抗旱育种是一项重要的农业科研领域,旨在培育出能够在干旱条件下生长良好的植物品种。
以下是一些常见的抗旱育种植物:1. 玉米(Zea mays):玉米是一种非常重要的食物作物,也是耐旱性强的植物之一。
通过选育出对干旱条件适应性强的耐旱玉米品种,可以提高玉米的产量和抵抗干旱的能力。
2. 大豆(Glycine max):大豆是一种富含蛋白质和植物油的作物,对干旱条件的适应性较强。
通过选育出耐旱大豆品种,可以提高大豆的产量和质量。
3. 小麦(Triticum):小麦是一种重要的粮食作物,也是一种相对耐旱的植物。
通过选育出耐旱小麦品种,可以提高小麦的产量和抵抗干旱的能力。
4. 水稻(Oryza sativa):水稻是全球最重要的粮食作物之一。
尽管水稻对水分的需求较大,但通过选育出对干旱条件适应性强的耐旱水稻品种,可以提高水稻的产量和抗旱能力。
5. 马铃薯(Solanum tuberosum):马铃薯属于块茎类作物,对干旱条件的适应性较强。
通过选育出耐旱马铃薯品种,可以提高马铃薯的产量和抵抗干旱的能力。
6. 红豆杉(Taxus chinensis):红豆杉是一种重要的林木和药用植物,对干旱条件的适应性较强。
红豆杉的干旱适应性可以通过选育和改良来提高。
7. 参(Panax):参属于药用植物,具有很高的商业价值。
一些耐旱性较强的参品种,如云南参、东北参等,对干旱条件有较强的适应能力。
8. 苜蓿(Medicago sativa):苜蓿是一种常见的饲料植物,对干旱条件的适应性较强。
通过选育出耐旱苜蓿品种,可以提高饲料产量和质量。
9. 沙棘(Hippophae rhamnoides):沙棘是一种适应性强的灌木植物,对干旱条件和贫瘠土壤有很强的耐受力,是一种理想的抗旱植物。
10. 紫花苜蓿(Trifolium pratense):紫花苜蓿是一种草本植物,适应性强,对干旱条件的适应能力较强,广泛用于牧草和土壤改良。
植物抗旱性研究进展及发展趋势

植物抗旱性研究进展及发展趋势户连荣1,郎南军2,郑科2 (1.西南林学院,云南昆明650224;2.云南省林业科学院,云南昆明650204)摘要 从植物干旱的种类、植物对水分胁迫的生理反应、抗旱机理、植物水分胁迫的研究方法等几个方面,探讨植物抗旱研究的进展、存在问题及发展趋势。
关键词 植物;抗旱机理;水分胁迫;进展;趋势中图分类号 Q948.112+.3 文献标识码 A 文章编号 0517-6611(2008)07-02652-03R esearch P rogress on the Drought R esistance of P lants and Its Developm ent T rendHU Lian2rong et al (S outhwest F orestry C ollege,K unm ing,Y unnan650224)Abstract T he research progress,the existing problems and the developm ent trend of the drought resistance of plants were discussed from such aspects as the kinds of plant drought,the physiological response of plants to water stress,the drought2resistant m echanism,the research m eth od on water stress in plants.K ey w ords Plant;Drought2resistance m echanism;W ater stress;Progrss;T rend 干旱是一个长期存在的世界性难题,全球干旱半干旱地区约占陆地面积的35%,遍及世界60多个国家和地区。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
草本植物抗旱性研究导师:董智教授姓名:彭志芳学号:20137101专业班级:13级水保2班E-mail:pzhf520@草本植物抗旱性研究彭志芳(山东农业大学林学院,山东泰安 271001)1文献检索概述基于研究课题“草本植物抗旱性研究”,特分别以“草本植物”和“抗旱性研究“为关键词,通过集中国知网、维普数据库、道客巴巴等于一体的百度学术进行了检索,检索结果如图1.1所示,显示“草本植物”从1958年开始出现相关研究,2008年达到最热,至今共有5939篇相关论文。
“草本植物”研究进程中,夏汉平、赵学勇、张洪江、徐海量、曹广民、李英年、刘国彬等前辈贡献了诸多优秀研究成果,他们推动并引领着草本植物学科的发展与进步。
其中中国农业科学院蔬菜花卉研究所、中国科学院植物研究所、中国科学院西北高原生物研究所、中国科学院沈阳应用生态研究所、中国科学院新疆生态与地理研究所、武汉市蔬菜科学研究所、青海大学地质工程系、中国科学院研究生院、娄底职业技术学院南校区10所研究机构在“草本植物”领域成果斐然,共有275篇相关论文。
图1.1“草本植物”研究走势图(资料来源:百度学术,)图1.2 “抗旱性研究”研究走势(资料来源:百度学术,)如图1.2所示“抗旱性研究”从1981年开始出现相关研究,2010年达到最热,至今共有567篇相关论文。
随着研究的不断深入,出现了越来越多与“抗旱性研究”相关的研究点,形成了庞大的研究网络,以下图1.3是高相关的研究点及其研究走势。
然相关文献浩如烟海,今研究即筛选其中极具代表的经典文献进行概述展开本文的研究。
图1.3 “抗旱性研究”关联研究(资料来源:百度学术,)2 草本植物抗旱性概述2.1草本植物草本(Herb),是一类植物的总称,但并非植物学科分类中的一个单元。
人们通常将草本植物称作“草”,而将木本植物称为“树”,但是也偶尔有例外,比如竹就属于草本植物,但人们经常将其看做是一种树。
草本植物多数在生长季节终了时,其整体部分死亡,包括一年生和二年生的草本植物,如水稻,萝卜等。
多年生草本植物的地上部分每年死去,而地下部分的根、根状茎及鳞茎等能生活多年,如天竺葵等。
草本植物有一年生、二年生和多年生的习性,有时会随地理纬度及栽培习惯的改变而变异,如小麦和大麦在秋播时为二年生草本,在春播时则成为一年生草本;又如棉花及蓖麻在江浙一带为一年生草本,而在低纬度的南方可长成多年生草本。
草本植物与木本植物最显著的区别在于他们茎的支持力量,草本植物的支持力量为膨压。
生长特点是生长周期短,受短日照影响,会长不大就开花,在盛夏来临前会开花、结种子,完成生命周期。
代表植物有秋播喜冷凉气候的草花(如紫罗兰、二月兰、瓜叶菊等)。
多年生草本植物的根一般比较粗壮,有的还长着块根、块茎、球茎、鳞茎等器官、冬天,地面上的部分仍安静地睡觉,到第二年气候转暖,它们又发芽生长。
这样一年一年地生长,地下的根或茎也许会渐渐地肥大起来,也许还会分枝。
这就给我们提供了识别它们年龄的依据;可以从它的地下部分分枝的多少,茎或根的大小,长短、粗细来推测它的年龄。
多年生草本能生活二年以上。
有些植物的地下部分为多年生,如宿根或根茎、鳞茎、块根等变态器官,而地上部分每年死亡,待第二年春又从地下部分长出新枝,开花结实,如藕、洋葱、芋、甘薯、大丽菊等;另外有一些植物的地上和地下部分都为多年生的,经开花、结实后,地上部分仍不枯死,并能多次结实,如万年青、麦门冬等。
草本植物有很多用处,所有重要的粮食都是草本植物,如小麦、粟米、玉米、大麦、高粱等,可以供人类食用。
自然界的很多动物,以及猪牛马羊等各类家畜也都吃草本植物,所以草本植物是食物链中的重要组成部分。
一些草本植物在日常生活处处可见,可以用来在编制和制作用具,东南亚还有竹子造的房屋。
大自然中的很多草本植物不但是动物的食物,而且还能制造大量氧气,防止水土流失。
很多草本植物是中药材的重要来源。
草本植物和木本植物最显著的区别在于他们茎的结构,草本植物的茎为“草质茎”,茎中密布很多相对细小的维管束,充斥维管束之间的是大量的薄壁细胞,在茎的最外层是坚韧的机械组织。
草本植物的维管束也与木本植物不同,维管束中的木质部分布在外侧而韧皮部则分布在内侧,这是与木本植物完全相反的,另外草本植物的维管束不具有形成层,不能不断生长,因而树会逐年变粗而草和竹子就没有这样的本领。
相比于木质茎,草质茎是更进化的特征。
2.2抗旱性陆生植物最常遭受的环境协迫是缺水,当植物耗水大于吸水时,就使组织内水分亏缺。
过度水分亏缺的现象,称为干旱(drought)。
旱害(drought injury)则是指土壤水分缺乏或大气相对湿度过低对植物的危害。
植物抵抗旱害的能力称为抗旱性(drought resistance)。
中国西北、华北地区干旱缺水是影响农林生产的重要因子,南方各省虽然雨量充沛,但由于各月分布不均,也时有干旱危害。
2.2.1干旱类型(1)大气干旱是指空气过度干燥,相对湿度过低,常伴随高温和干风。
这时植物蒸腾过强,根系吸水补偿不了失水,从而受到危害。
中国西北、华北地区常有大气干旱发生。
(2)土壤干旱是指土壤中没有或只有少量的有效水,这将会影响植物吸水,使其水分亏缺引起永久萎蔫。
(3)理干旱土壤水分并不缺乏,只是因为土温过低、土壤溶液浓度过高或积累有毒物质等原因,妨碍根系吸水,造成植物体内水分平衡失调,从而使植物受到的干旱危害。
大气干旱如持续时间较长,必然导致土壤干旱,所以这两种干旱常同时发生。
在自然条件下,干旱常伴随着高温,所以,干旱的伤害可能包括脱水伤害(狭义的旱害)和高温伤害(热害)。
2.2.2干旱伤害植物的机理干旱对植株最直观的影响是引起叶片、幼茎的萎蔫。
萎蔫可分为暂时萎蔫和永久萎蔫,两者根本差别在于前者只是叶肉细胞临时水分失调,而后者原生质发生了脱水。
原生质脱水是旱害的核心,由此可带来一系列生理生化变化并危及植物的生命。
(1)改变膜的结构及透性:当植物细胞失水时,原生质膜的透性增加,大量的无机离子和氨基酸、可溶性糖等小分子被动向组织外渗漏。
细胞溶质渗漏的原因是脱水破坏了如图2.1所示的原生质膜脂类双分子层的排列所致。
正常状态下的膜内脂类分子靠磷脂极性同水分子相互连接,所以膜内必须有一定的束缚水时才能保持这种膜脂分子的双层排列。
而干旱使得细胞严重脱水,膜脂分子结构即发生紊乱,膜因而收缩出现空隙和龟裂,引起膜透性改变。
图2.1原生质膜脂类双分子层(2)破坏了正常代谢过程:细胞脱水对代谢破坏的特点是抑制合成代谢而加强了分解代谢,即干旱使合成酶活性降低或失活而使水解酶活性加强。
对光合作用的影响是水分不足使光合作用显著下降,直至趋于停止。
番茄叶片水势低于-0.7MPa时,光合作用开始下降,当水势达到-1.4MPa时,光合作用几乎为零。
干旱使光合作用受抑制的原因是多方面的,主要由于:水分亏缺后造成气孔关闭,CO2扩散的阻力增加;叶绿体片层膜体系结构改变,光系统Ⅱ活性减弱甚至丧失,光合磷酸化解偶联、叶绿素合成速度减慢、光合酶活性降低、水解加强、糖类积累等这些都是导致光合作用下降的因素。
(3)对呼吸作用的影响:干旱对呼吸作用的影响较复杂,一般呼吸速率随水势的下降而缓慢降低。
有时水分亏缺会使呼吸短时间上升,而后下降,这是因为开始时呼吸基质增多的缘故。
若缺水时淀粉酶活性增加,使淀粉水解为糖,可暂时增加呼吸基质。
但到水分亏缺严重时,呼吸又会大大降低。
如马铃薯叶的水势下降至-1.4MPa时,呼吸速率可下降30%左右。
(4)脯氨酸积累:干旱时植物体内的蛋白质分解加速,合成减少,这与蛋白质合成酶的钝化和能源(ATP)的减少有关。
如玉米水分亏缺3小时后,ATP含量减少40%。
蛋白质分解则加速了叶片衰老和死亡,当复水后蛋白质合成迅速地恢复。
所以植物经干旱后,在灌溉与降雨时适当增施氮肥有利于蛋白质合成,补偿干旱的有害影响。
与蛋白质分解相联系的是,干旱时植物体内游离氨基酸特别是脯氨酸含量增高,可增加达数十倍甚至上百倍之多。
因此脯氨酸含量常用作抗旱的生理指标,也可用于鉴定植物遭受干旱的程度。
(5)使核酸减少:随着细胞脱水,其DNA和RNA含量减少。
主要原因是干旱促使RNA酶活性增加,使RNA分解加快,而DNA和RNA的合成代谢则减弱。
当玉米芽鞘组织失水时,细胞内多聚核糖体解离成单体,失去了合成蛋白质(酶)的功能。
因此有人认为,干旱之所以引起植物衰老甚至死亡,是同核酸代谢受到破坏有直接关系的。
干旱时细胞分裂素含量降低,脱落酸含量增加,这两种激素对RNA酶活性有相反的效应,前者降低RNA酶活性,后者提高RNA酶活性。
此外水分的分配异常干旱时植物组织间按水势大小竞争水分。
一般幼叶向老叶吸水,促使老叶枯萎死亡。
有些蒸腾强烈的幼叶向分生组织和其它幼嫩组织夺水,影响这些组织的物质运输。
例如禾谷类作物穗分化时遇旱,则小穗和小花数减少;灌浆时缺水,影响到物质运输和积累,籽粒就不饱满。
对于其它植物,也常由此造成落花落果,影响产量。
(6)机械性损伤 :上述的旱害多属破坏正常代谢,一般不至于造成细胞或器官的立即损伤或死亡。
而干旱对细胞的机械性损伤可能会使植株立即死亡。
细胞干旱脱水时,液泡收缩,对原生质产生一种向内的拉力,使原生质与其相连的细胞壁同时向内收缩,在细胞壁上形成很多折叠,损伤原生质的结构。
如果此时细胞骤然吸水复原,可引起细胞质、壁不协调膨胀把粘在细胞壁上的原生质撕破,导致细胞死亡。
干旱引起的伤害可由图2.2所示。
图2.2 干旱引起的植物伤害机理示意图2.2.3抗旱性的机理抗旱性是植物对旱害的一种适应,通过生理生化的适应变化减少干旱对植物所产生的危害。
通常农作物的抗旱性主要表现在形态与生理两方面。
形态结构特征是抗旱性强的作物往往根系发达,伸入土层较深,能更有效地利用土壤水分。
根冠比大可作为选择抗旱品种的形态指标。
抗旱作物叶片的细胞体积小,这可减少失水时细胞收缩产生的机械伤害。
抗旱作物的维管束发达,叶脉致密,单位面积气孔数目多,这不仅加强蒸腾作用和水分传导,而且有利于根系的吸水。
有的作物品种在干旱时叶片卷成筒状,以减少蒸腾损失。
不同植物可通过不同形态特征适应干旱环境。
生理生化特征方面是保持细胞有很高的亲水能力,防止细胞严重脱水,这是生理性抗旱的基础。
最关键的是在干旱条件下,水解酶类如RNA酶、蛋白酶、脂酶等保持稳定,减少生物大分子分解,这样既可保持原生质体,尤其是质膜不受破坏,又可使细胞内有较高的粘性与弹性,通过粘性来提高细胞保水能力,同时弹性增高又可防止细胞失水时的机械损伤。
原生质结构的稳定可使细胞代谢不至发生紊乱异常,使光合作用与呼吸作用在干旱下仍维持较高水平。
植物保水能力或抗脱水能力是抗旱性的重要指标。
另外脯氨酸、甜菜碱和脱落酸等物质积累变化也是衡量植物抗旱能力的重要特征。