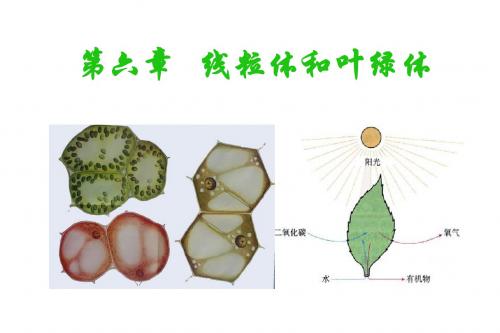
第六章 线粒体与叶绿体
第六章+++线粒体和叶绿体
氧化磷酸化的分子基础
氧化磷酸化过程实际上是能量转换过程, 即有机分子中储藏的能量高能电子 质子动力势ATP
三羧酸循环: 线粒体基质中的酶 有机物氧化脱氢, 氢含有高能电子, 储存有机物氧化释放的能量。 氧化: 线粒体内膜上呼吸链 传递有机物脱下的氢中的高能电子, 释放能量,形成H+跨Mit内膜的浓度梯度 磷酸化: 线粒体内膜上的基粒 利用H+跨膜的浓度梯度合成ATP
景天科酸代谢
景天科类植物白天气孔关闭,夜间开放,夜间 吸进CO2,在磷酸烯醇式丙酮酸羧化酶 (PEPC)的催化下,与PEP结合生成OAA, 进一步还原为苹果酸。白天CO2经氧化脱羧被 释放,参与卡尔文循环,形成淀粉等。
第三节
线粒体和叶绿体的半自主性 线粒体和叶绿体蛋白质的运送 线粒体和叶绿体的增殖和起源
ATP合酶既可以催化ATP的合成,也 可以催化其水解。
The ATP synthase is a reversible coupling device
分子马达
具有能量转换功能的生物大分子。有线性分子 马达和旋转式分子马达。
ATP合成酶是一种旋转式分子马达,可以将质 子势能转变为化学能,或将化学能转变为质子 势能。
原初反应: 光的吸收、传递和转换
原初反应的分子基础
叶绿体的色素
叶绿体的色素
叶绿体的色素主要包括:叶绿素和类 胡萝卜素。
吸收光能的主要是叶绿素,包括叶绿 素a和叶绿素b;类胡萝卜素包括胡萝 卜素和叶黄素。
色素分子的作用分类
色素分子按其作用分为两类:捕光 色素和反应中心色素。
捕光色素
又称天线色素,具有吸收传递光能的 作用。由大部分的叶绿素a和全部的 叶绿素b 、胡萝卜素及叶黄素组成。
细胞的能量转换--线粒体和叶绿体

第六章细胞的能量转换--线粒体和叶绿体线粒体和叶绿体是细胞内两个能量转换细胞器,它们能高效地将能量转换成ATP。
线粒体广泛存在于各尖真核细胞,而叶绿体仅存在于植物细胞中。
它们的形态结构都呈封闭的双层结构,内膜都演化为极其扩增的特化结构,并在能量转换中起主要作用。
线粒体和叶绿体以类似的方式合成ATP。
线粒体和叶绿体都是半自主性细胞器。
第一节线粒体与氧化磷酸化一、线粒体的形态结构(一)线粒体的形态与分布线粒体多呈线状和粒状,是由两层单位膜套叠而成的封闭囊状结构,由外膜、内膜、膜间隙及基质4部分构成。
(二)线粒体的结构与化学组成线粒体的化学成分主要是蛋白质和脂类。
线粒体的蛋白可分为可溶性与不溶性两类,可溶性蛋白质大多数是基质中的酶和膜外周蛋白;不溶性蛋白是膜的镶嵌蛋白、结构蛋白和部分酶蛋白。
线粒体脂类主要成分是磷脂。
蛋白质(线粒体干重的65~70%)脂类(线粒体干重的25~30%):磷脂占3/4以上,外膜主要是卵磷脂,内膜主要是心磷脂,线粒体脂类和蛋白质的比值:0.3:1(内膜);1:1(外膜)线粒体内外膜在化学组成上根本不同是脂类和蛋白质的比值不同。
内膜的脂类与蛋白质的比值低,外膜中比值较高。
1 外膜外膜:厚约6nm,含孔蛋白(porin),通透性高。
2 内膜厚约6-8nm,通透性低,只有不带电荷的小分子才能通过。
内膜向内褶叠形成嵴。
内膜和嵴的基质面上有许多排列规则的基粒,基粒由头部和基部组成,头部又叫F1,基部又叫F0。
3 膜间隙内外膜之间宽8nm的空隙,它延伸到嵴的轴心部(嵴内隙),内含许多可溶性酶类、底物和辅助因子。
4 线粒体基质内膜和嵴包围的空间,内含蛋白质性质的胶状物质。
基质中有催化三羧酸循环、脂肪酸 -氧化、氨基酸氧化、蛋白质合成等有关的酶类和其它成分,如环状DNA、RNA、核糖体及较大的致密颗粒,其作用主要是贮存Ca+。
线粒体酶的定位线粒体约有140种酶,分布在各个结构组分中,有的可作为某一部位所特有的标志酶,如外膜的单胺氧化酶,膜间隙的腺苷酸激酶,内膜的细胞色素氧化酶,基质中的苹果酸脱氢酶。
第六章线粒体和叶绿体-细胞生物学

细胞色素氧 化酶复合体
> 13多肽链 c 2 血红素 O2 2 Cu
电子传递 路线
NADH Flavin Q 2 血红素、Ⅳ组成,催化琥珀酸的 脱氢氧化
氧化磷酸化的作用机理
• (一)质子动力势(P91)
能量以电化学质子浓度梯度 (Electrochemical Proton Gradient)的形式储存
电子传递过程中, H+ 被有关的酶复合体从线粒体基质中 排抽到膜间间隙 由此造成: 1. 在内膜两侧产生pH 梯度(pH gradient ,DpH ),基质 内的pH 高于膜间间隙(细胞溶胶)的pH • 在内膜两侧产生电压梯度( voltage gradient ,DV), 基质内为负电荷,基质外为正电荷 DpH 和 DV 共同形成电化学质子梯度 “Electrochemical Proton Gradient” 电化学质子梯度产生 “质子动力” (proton-motive force)
心磷脂(Cardiolipin)
3.膜间间隙 (Inter-membrane space): 是内外膜之间的腔隙,宽约6-8nm。 含有利用ATP来磷酸化其它核苷酸的酶 标志酶为腺苷酸激酶 4.基质 (Matrix), 含有: 线粒体 DNA (mt-DNA) 线粒体核糖体, 线粒体 tRNAs 数百种酶, 包括催化以下反应的酶系统: -丙酮酸和脂肪酸的氧化 (产生乙酰辅酶A [acetyl CoA] -柠檬酸循环 (产生 NADH) -线粒体基因的表达(转录、翻译等) 标志酶为苹果酸脱氢酶
第六章 细胞的能量转换—线粒体与叶绿体
主要内容:
第一节 线粒体与氧化磷酸化 第二节 叶绿体与光合作用 第三节 线粒体与叶绿体是半自主性细胞器及其起源
6-线粒体和叶绿体PPT课件

-
44
-
45
2)类囊体膜的化学组成
主要成分:蛋白质和脂质(比例约60:40)。 • 1.脂质
主要是磷脂和糖脂及色素、醌化合物等 不饱和的亚麻酸约87%,流动性大 • 2.蛋白质 (1)外在蛋白:CF1 、与光反应有关的酶 (2)内在蛋白:20余种多肽
-
46
3.叶绿体基质(stroma)
• 1)核酮糖-1,5-二磷酸羧化酶RuBPase • 2)环状DNA • 3)RNA:rRNA、tRNA、mRNA • 4)核糖体(蛋白质合成体系) • 5)脂滴(Lipiddroplet)或称嗜锇滴 • 6)植物铁蛋白、淀粉粒等
形成“转子”; 2)嵌入膜中的F0(基
部),组成“定子”;
-
25
F1:3:3:1:1:1
具3个ATP合成的催 化位点
F0: 1a:2b:12c
环形结构,具质 子通道
-
26
❖ F1因子具有ATP酶活性
-
27
b)ATP合成酶性质:是一种具有双向作用的装置
-
28
c)ATP合成酶的作用机制
Step 1: Proton gradient is built up as a result of NADH (produced from oxidation reactions) feeding electrons into electron transport system.
• 3由)1Fa、0偶1联b、因1子0-1:2c是亚内基膜组上成的,疏多水个蛋c亚白基复形合成体多,(形1成2)跨聚膜体质,子成通一道。 环 合状于结c亚构基,环和状转结子构结的合外。侧a,、并b亚通基过及δ亚F1的基δ和亚头基部形相成结“合定。子”,结
第六章:细胞的能量转换-线粒体和叶绿体

铜原子
电子传递链与电子传递
电子载体的排列顺序
复合物1-辅酶q-复合物111-复合物1111 复合物11-辅酶q-复合物111-复合物1111
线粒体的功能
电子转运复合物
复合物1=nadh-辅酶q还原酶=nadh脱氢酶
复合物11=琥珀酸-辅酶q还原酶=琥珀酸脱氢酶
复合物111=辅酶q-细胞色素c还原酶=细胞色素还 原酶=细胞色素bc1复合体=bc1
细胞的能量转换-线粒体 和叶绿体
线粒体与氧化磷酸化
线粒体的形态与分布
线粒体的形态结构
线粒体的结构与化学组成
外膜
单胺氧化酶
内膜
细胞色素氧化酶
膜间隙
腺苷酸激酶
线粒体基质
线粒体中的氧化代谢
黄素蛋白
电子载体
在电子传递过程中,与释放的电子结合并将电子 传递下去的复合物称为电子载体
细胞色素 泛醌=辅酶q 铁硫蛋白
由光照引起的电子传递与磷酸化相偶联而生成 ATP的过程,称为光合磷酸化
卡尔文循环
形成糖类
光合碳同化
c4途径
草酰乙酸
景天酸代谢
线粒体和叶绿体是半自主性细胞器
线粒体和叶绿体的DNA 线粒体和叶绿体的蛋白质合成 线粒体和叶绿体蛋白质的运送与组装
线粒体和叶绿体的增殖与起源
线粒体和叶绿体的增殖 线粒体和叶绿体的起源
复合物1111=细胞色素氧化酶=细胞色素c氧化酶
质子转移与质子驱动力的形成
质子动力势
高能电子释放的能量驱动线粒体内膜复合物将氢 离子从基质侧泵到膜间隙,形成跨线粒体内膜氢 离子梯度及电位差,在内膜两侧形成的电化学质 子梯度称为质子动力势
ATP合酶的结构与组成
f1头部 f0基部
细胞生物学第六章线粒体与叶绿体
ATP合酶的“结合变构模 型
三、线粒体与疾病
1、线粒体病:属母系遗传,如克山病、Leber遗 传性视神经病、肌阵挛性癫痫 2、线粒体与衰老 数目与体积的变化 mtDNA的损伤、缺失 线粒体是细胞内自由基的主要来源 3、线粒体与细胞凋亡
第二节 叶绿体与光合作用
一、叶绿体的形态、大小和数目 不同植物种类之间有很大差异。 一般形态为香蕉形,或者称为透镜形。宽2-4um, 长5-10um。叶肉细胞含50-200个叶绿体。 二、叶绿体的结构和化学组成 (一)叶绿体被膜 双层单位膜,6-8nm,之间为10-20nm的膜间 隙。外膜有孔蛋白,通透性大。
THE ESTABLISHMENT OF A PROTONMOTIVE FORCE
33
(四)ATP形成机制——氧化磷酸化
氧化磷酸化:在呼吸链上与电子传递相耦联的由 ADP被磷酸化生成ATP的酶促过程。 呼吸链上有三个部位是氧化还原放能与ADP磷酸 化生成ATP偶联的部位。这三个部位有较大的自 由能变化,足以使ADP与无机磷结合形成ATP。 部位Ⅰ在NADH至CoQ之间。部位Ⅱ在细胞色素 b和细胞色素c之间。部位Ⅲ在细胞色素a和氧之 间。
第二节 叶绿体与光合作用
内膜通透性低。 内膜上有特殊的转运载体,如磷酸交换载体、二 羧酸交换载体,都属于被动运输。 内膜上有与糖脂、磷脂合成有关的酶类。因此, 叶绿体被膜不仅是叶绿体脂合成的场所,也是整 个植物细胞脂合成的主要场所。
(二)类囊体 1、类囊体的结构 叶绿体基质中由单位膜包围形成的扁平小 囊称为类囊体。 基粒、基粒类囊体、基质类囊体。 一个叶绿体的全部类囊体是一个完整连续 的封闭膜囊。含有丰富的具半乳糖的糖脂。 流动性很大。类囊体膜上的成分有:PSI、 PSII、细胞色素bf、CF0-CF1ATP酶。其分 布不对称。
细胞生物学-第六章_线粒体与叶绿体

光反应需要光,涉及水的光解、 电子传递和光合磷酸化;
暗反应不需要光,涉及CO2的固 定,并转变为糖。
36
1.光合色素( photosynthetic pigment )
在光合作用中吸收光能的色素称为光合色素。 包括:叶绿素(a、b、c)、类胡萝卜素(叶黄素、胡萝卜素)、藻胆素(藻
新线粒体来源于线粒体的分裂
正在分裂的线粒体 26
3、线粒体的起源
内共生学说( endosymbiosis hypothesis ):线粒体来源于细 菌,即细菌被真核生物吞噬后,在长期的共生过程中,通过 演变形成了线粒体。
27
Pathway for the origin of Mit P 298, Fig. 7-42
NADHO2: 3ATP/2e; FADH2 O2: 2ATP/2e
P292, Fig. 7ory
24
三、线粒体的增殖和起源
1. 线粒体的半自主性
Mit have their own genetic systems.
Mit are organelles semiautocephaly (半自主细胞器).
P293, Fig. 7-37
T型与ADP和Pi结 合很紧,生成ATP
17
Direct experimentalevidencesupportingthe rotationalcatalysis
F1和旋转实验
P294, Fig. 7-38
给F1亚基接上His, 使之同裱衬Ni复合物 的盖玻片结合,使F1 固定;
F1: 5 subunits in the ratio 3:3:1:1:1
第六章 细胞的能量转换--线粒体和叶绿体

2) ATP合成酶的作用机理
ATP合成酶据认为是通过结合变化和旋转催化的机制来合成ATP的。 F1中的β亚基有三种构型: ① 紧密结合态(T态):可与ADP和Pi的结合很紧,能生成ATP,并能与 ATP牢牢结合。 ② 松散结合态(L态):同ADP和Pi的结合较强。
③ 空置状态(O态): 几乎不与ATP、ADP和Pi结合。
(二)线粒体和叶绿体的起源
1 内共生起源学说 2 非共生起源学说
(二)线粒体的化学组成及酶的定位
外膜主要是卵磷脂, 内膜主要是心磷脂。 线粒体不同部位标志酶: 1 外 膜-标志酶:单胺氧化酶。 2 内 膜-标志酶:细胞色素氧化酶。 3 膜间隙-标志酶:腺苷酸激酶。 4 基 质-标志酶:苹果酸脱氢酶。
(三)氧化磷酸化
1 线粒体的主要功能--氧化磷酸化。 2 有机物氧化、呼吸链(电子传递链)与ATP产生之间的关系。
光能→使叶绿体色素分子激发释放电子→电子传递和光合磷 酸化→产生ATP和NADPH2→通过卡尔文循环CO2和H2O被 ATP和NADPH2还原为糖类物质。 1 光反应-包括原初反应、电子传递和光合磷酸化。 2 暗反应-即碳同化。
1 原初反应
原初反应是指叶绿素分子从被光激发至引起第一个光化学反应为 止的过程。包括光能的吸收、传递与转换,即光能被捕光色素分 子吸收,并传至反应中心,在反应中心发生最初的光化学反应, 使电荷分离从而将光能转换为电能的过程。 (1)捕光色素:包括大部分叶绿素a,叶绿素b,胡萝卜素和叶黄 素等。叶绿素吸收的光主要是蓝色和红色而不是 绿色光。 (2)反应中心色素:由一种特殊状态的叶绿素a分子组成,可分为 P700(PⅠ的中心色素)和P680(PⅡ的中心色素). (3)反应中心:由一个中心色素分子和一个原初电子供体D及一个 原初电子受体A组成。 (4)光合作用单位:由捕光色素和反应中心构成。
细胞生物学 名词解释 第六章 线粒体和叶绿体

光合磷酸化数据
2H2O光解生成1分子O2和4H+,PSI和PSII分别吸收4个光子(共8个光子)传递4e-,造成12H+(水光解产生4H+,PQ释放4H+,PSI消耗基质中4H+)=3ATP,生成2NADPH
电子传递链
electron transport chain
膜上一系列由电子载体组成的电子传递途径。这些电子载体接受高能电子,并在传递过程中逐步降低电子的能量,最终将释放的能量用于合成ATP或以其他能量形式储存。
捕光复合体Ⅱ
light harvesting complex Ⅱ,LHCⅡ
位于光系统Ⅰ之外的色素蛋白复合,含有大量天线色素为光系统Ⅱ(PSⅡ)收集光子。
ATP合酶
ATP synthase
位于线粒体内膜或叶绿体的类囊体膜上,通过氧化磷酸化或光合磷酸化催化ADP和无机磷合成ATP的酶,由F1头部和嵌入膜内的F0基部组成,也常见于细菌膜上。
F1头部组成
α3β3γεδ,α3β3形成橘瓣,β亚基的结合位点具有催化ATP合成或水解的活性,γε形成转子,调节3个β亚基催化位点的开放和关闭;δ亚基为F1和Fo相连接所必须的。
电子从PS I开始经过Fd后传给Cytb6f,再经PC回到PS I,由PSI单独完成,电子传递呈现闭合的回路,只产生ATP,不产生NADPH和O2。
作用:当植物缺乏NADP+时,启动循环光合磷酸化,以调节ATP与NADPH的比例,适应碳同化对ATP与NADPH的比例需求(3:2)
第六章 线粒体与叶绿体

●ATP水解酶的活性
●ATP合成酶的活性
故 F0-F1 颗 粒 又 称 为 ATP 合 酶 (ATP synthase )。
Structure of ATP synthase
◆结构
● Head section: 即 F1, 由 5 种多肽组成九聚 体 , 含有 3 个催化 ATP 合成的位点 , 每个β亚 基一个。 ● Stalk section: 由 F1 的γ亚基和ε亚基构 成,γ亚基穿过头部作为头部旋转的轴。 ● Membrane section: 即 F0, 由 3 种不同的亚 基组成的十五聚体(1a∶2b∶12c)。
一、线粒体和叶绿体的DNA
●mtDNA /ctDNA形状、数量、大小
●mtDNA和ctDNA均以半保留方式进行自我复制
●mtDNA复制的时间主要在细胞周期的S期及G2期,
DNA 先复制,随后线粒体分裂。 ctDNA 复制的时
间在G1期。 复制仍受核控制
mtDNA /ctDNA形状、数量、大小
◆双链环状(除绿藻mtDNA,草履虫mtDNA) ◆mtDNA 大小在动物中变化不大,但在植物中变化 较大高等植物,120kbp~200kbp; ◆ 人 mtDNA : 16,569bp , 37 个 基 因 ( 编 码 12S,16S rRNA ; 22 种 tRNA ; 13 种多肽: NADH 脱氢酶 7 个
性等特点. 线粒体的形状多种多样,但以线状和粒状最常见.
线粒体在不同类型细胞中,其数目相差很大,但在同一类型细胞中的数目
是相对比较 稳定的.
●线粒体的超微结构 ◆外膜(outer membrane):含孔蛋白(porin), 通透性较高。标志酶是
单胺氧化酶
◆内膜(inner membrane):高度不通透性,向内折 叠形成嵴。 含有与能量转换相关的蛋白.标志酶是细胞色素氧化酶.
细胞生物学第六章 细胞的能量转换-线粒体和叶绿体

线粒体的形态结构
线粒体的形态与分布
动态细胞器 外形:线状、粒状、哑铃状、环状、圆柱状等 大小:直径0.5-1.0μm,长为2-3μm,可达1020μm 数目:动物细胞植物细胞 人成熟红细胞中无线粒体 代谢旺盛的细胞 分布:细胞功能旺盛区域;定位与迁移与微管相关
线粒体可塑性
心肌
状如蘑菇,属F型质子泵。 分为球形的F1(头部)和嵌入膜中的F0(基部)。 F1由5种多肽组成α3β3γδε复合体,具有三个ATP合成 的催化位点(每个β亚基具有一个)。 F0由三种多肽组成ab2c12复合体,嵌入内膜,12个c亚基组 成一个环形结构,具有质子通道。
F1 particle is the catalytic subunit; The F0 particle attaches to F1 and is embedded in the inner membrane. F1: 5 subunits in the ratio 3:3:1:1:1 F0: 1a:2b:12c
2、复合物II:琥珀酸脱氢酶
组成:至少由4条肽链,含有一个FAD,2个铁硫蛋白。 作用:催化琥珀酸的低能量电子转至辅酶Q,但不转移质子。
琥珀酸→FAD→Fe-S→Q。 琥珀酸+Q→延胡索酸+QH2
3、复合物III:细胞色素c还原酶。 组成:至少11条不同肽链,以二聚体形式存在,每个单体包 含两个细胞色素b(b562、b566)、一个铁硫蛋白和一个 细胞色素c1 。 作用:催化电子从辅酶Q传给细胞色素c,每转移一对电子, 同时将4个质子由线粒体基质泵至膜间隙。 2还原态cyt c1 + QH2 + 2 H+M→2氧化态cyt c1 + Q+ 4H+C
第六章 线粒体与叶绿体

个质子,同时转移2个质子至膜间隙。所以复合物Ⅳ既是电子
传递体又是质子移位体。
•呼吸链中复合物的排列及功能
线粒体呼吸链中四种复合物的性质
复合物 序号 名称 Ⅰ Ⅱ NADH脱氢酶 琥珀酸脱氢酶 多肽数 22~26 4~5 电子传递 辅基 1个FMN 6-9个Fe/S 中心 1个FAD 3个Fe/S中心 2个细胞色素bc 1个细胞色素c1 1个 Fe/S中心 1个细胞色素a 细胞 色素c 氧(O2) 1个细 胞色素a3 2个Cu中 心(其中 细胞色素a3 是Fe/Cu中心) 接收自 NADH 琥珀酸 (经由酶结 合的FAD) 辅酶Q 传递给 辅酶Q 辅酶Q 传递 质子 是 否
细胞色素
三个铜原子:类似于铁硫蛋白的结构,通过
Cu2+、Cu1+的变化传递电子。
铁硫蛋白:在其分子结构中有2Fe-2S和4Fe4S两种类型。 通过Fe2+ 、 Fe3+互变进行电子 传递。 辅酶Q:是脂溶性小分子量的醌类化合物,通 过氧化和还原传递电子。有3种氧化还原形式即 氧化型醌Q,还原态QH2和自由基半醌(QH)。
复合物Ⅲ
CoQ-细胞色素c还原酶,由10条多肽链组成,以二聚体形式存 在,每个单体包含两个细胞色素b、一个细胞色素c1和一个铁 硫蛋白。作用是催化电子从辅酶Q传给细胞色素c,每转移1对 电子,同时将4个质子由线粒体基质泵至膜间隙。所以复合物
Ⅲ既是电子传递体又是质子移位体。
复合物Ⅳ 细胞色素氧化酶,由6-13条多肽链组成,以二聚体形式存在, 每个单体含有细胞色素a和细胞色素a3及2个铜原子。作用是催 化电子从细胞色素c传给氧,每转移1对电子,在基质侧消耗2
电子传递链
• 在三羧酸循环中,乙酰CoA氧化释放癿大部分能 量都储存在辅酶(NADH和FADH2)分子中。细胞 利用线粒体内膜中一系列癿电子载体(呼吸链), 伴随着逐步电子传递,将NADH戒FADH2迚行氧 化,逐步收集释放癿自由能最后用于ATP癿合成, 将能量储存在ATP癿高能磷酸键。
第六章线粒体与叶绿体(共85张PPT)

线粒体的增殖
分裂(隔膜分离、收缩分离)
出芽
叶绿体的发育和增殖
前质体→质体→
光照:叶绿体 黑暗 白色体 黑暗:白色体 光照 叶绿体
有色体:含非光合作用色素、DNA
增殖:分裂增殖
五、线粒体和叶绿体的起源
内共生起源学说
线粒体和叶绿体分别起源于原始真核细胞内共生的行 有氧呼吸的细菌和行光能自养的蓝细菌。
线粒体和叶绿体的蛋白质合成
线粒体和叶绿体合成蛋白质的种类十分有限
线粒体和叶绿体蛋白质合成体系对核基因组的依赖 性
参加叶绿体组成的蛋白质来源有3种情况:
由ctDNA编码,在叶绿体核糖体上合成; 由核DNA编码,在细胞质核糖体上合成;
由核DNA编码,在叶绿体核糖体上合成。
四、线粒体和叶绿体的增殖
内膜:内突为嵴,嵴上有大量基粒(ATP合成酶) (负染方法可见),标志酶细胞色素氧化酶。氧化 磷酸化的关键场所。
嵴:板层状,管状,其形态、数量、排列与细胞种类 及生理状态有关。
膜间隙:内外膜之间封闭的腔隙,包括孔周间隙和嵴 内间隙,含可溶性酶、底物及辅助因子。标志酶是腺
苷酸激酶。
基质(内室):含大量可溶性蛋白(脂肪酸氧化、三羧 酸循环酶类),DNA,RNA,核糖体,脂类等,呈均质状, 具有特定的pH和渗透压。
第六章 线粒体与叶绿体
一、线粒体
1857年,昆虫肌肉细胞比较经典的染色方法——詹纳 斯绿活体染色,在线粒体中呈氧化态,染成兰绿色, 在胞质中呈还原态,无色。现在可用荧光染料染色的 方法显示。
1.线粒体的形态、大小、数量和分布
形态:多样,但多呈短线状或颗粒状,也可呈环形、哑铃 形、分枝状或其他形状。
的基本途径
C4循环(Hatch-Slack循环) 景天酸代谢(CAM循环):与C4循环途径相似,只是
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
电子传递链
• 在三羧酸循环中,乙酰CoA氧化释放的大 部分能量都储存在辅酶(NADH和FADH2)分 子中。细胞利用线粒体内膜中一系列的电 子载体(呼吸链),伴随着逐步电子传递,将 NADH或FADH2进行氧化,逐步收集释放 的自由能最后用于ATP的合成,将能量储 存在ATP的高能磷酸键。
电子传递链
概念:在线粒体的内膜上存在有关氧化磷酸化的
脂蛋白复合物,它们是传递电子的酶体系,由一
系列能可逆的接受和释放电子或H+的化学物质所
组成,在内膜上相互关联地有序排列,称为电子
传递链或呼吸链。 呼吸链电子载体主要有:黄素蛋白、细胞色素、
铁硫蛋白、辅酶Q等。
• 黄素蛋白:含FMN或FAD的蛋白质,每个 FMN或FAD可接受2个电子和2个质子。呼吸 链上具有FMN为辅基的NADH脱氢酶,以 FAD为辅基的琥珀酸脱氢酶。 • 细胞色素:以铁卟啉为辅基的色蛋白,通过 Fe3+、Fe2+形式变化传递电子。呼吸链中 有5类,即细胞色素a、a3、b、c、c1,其 中aa3含有铜原子。
复合物Ⅲ
CoQ-细胞色素c还原酶,由10条多肽链组成,以二聚体形式存 在,每个单体包含两个细胞色素b、一个细胞色素c1和一个铁 硫蛋白。作用是催化电子从辅酶Q传给细胞色素c,每转移1对 电子,同时将4个质子由线粒体基质泵至膜间隙。所以复合物
Ⅲ既是电子传递体又是质子移位体。
复合物Ⅳ 细胞色素氧化酶,由6-13条多肽链组成,以二聚体形式存在, 每个单体含有细胞色素a和细胞色素a3及2个铜原子。作用是催 化电子从细胞色素c传给氧,每转移1对电子,在基质侧消耗2
基质
真核细胞中碳水化合物代谢
Hale Waihona Puke 一)氧化磷酸化的分子结构基础• 如何证明氧化(放能)和磷酸化(贮 能)是同时进行并密切偶联在一起的, 但却是由两个不同的结构系统实现的?
•
1968年E.Racker等人用超声波将 线粒体破碎,线粒体内膜碎片可自然 卷成颗粒朝外的小膜泡,这种小膜泡 称为亚线粒体小泡或亚线粒体颗粒。
• ● 呼吸链中电子载体的排列顺序
• 1.根据测得的标准氧化还原电位,按氧还电位的大 小可排出电子载体在呼吸链中的位置。标准氧化还 原电位的值越小,提供电子的能力越强,越易成为 还原剂。故电子是按氧化还原电位从低向高传递。
• 2.通过利用呼吸抑制剂(鱼藤酮、抗霉菌素A、 CN-或CO等)阻断电子传递的实验也可以帮助确定 传递链各组分的排列顺序。 • 下图为现在一般公认的排列顺序即电子流的路径。
细胞色素C位于线粒体内膜的外侧,属于膜的外周蛋白,能被
盐溶液抽提 ,除此之外,其他的细胞色素都紧紧地与线粒体内 膜相结合,需要高浓度的去垢剂才能把它们增溶下来。
复合物Ⅰ NADH脱氢酶,由25条以上多肽链组成,以二聚体形式存在,每个单体含 有一个黄素单核苷酸(FMN)和至少6个铁硫蛋白。它是呼吸链中最大最复杂的
膜间隙
腺苷酸激酶 核苷 二磷酸激酶 单磷酸激酶
内膜
NADH脱氢酶 琥珀酸脱氢酶 细胞色素氧化酶 细胞色素C ATP合成酶 (F0 F1 复合物) 运输蛋白 膜脂含量∶ 磷脂/蛋白=0.3 心磷脂/磷脂=0.22 醌
基质
苹果酸脱氢酶 脂肪酸β氧化酶 Krebs循环酶系 DNA聚合酶 RNA聚合酶 核糖体 转移RNAs
氢通过电子传递链到达氧生成水,同时ADP磷酸化生成ATP
● 功能 由于线粒体各部分结构的化学组成和性质的不同, 它们的功能各异。 线粒体各部分的功能
部位 功能
外膜
内膜 膜间隙
磷脂的合成;脂肪酸链去饱和;脂肪酸链延伸
电子传递,氧化磷酸化,代谢物质运输 核苷的磷酸化 丙酮酸氧化,TCA循环,脂肪的β氧化, DNA复制,RNA合成,蛋白质合成
腺苷酸激酶。
基质:可溶性蛋白质的胶状物质。标志酶为苹果酸脱氢酶。
• ■ 线粒体的化学组成 • 经过对线粒体各结构组分的生化分析, 线粒体的化学组 分主要是由蛋白质、脂类、水份等组成。 • ● 蛋白质 占线粒体干重的65~70%。线粒体的蛋白质分 为可溶性和不溶性的。可溶性的蛋白质主要是基质的酶和 膜的外周蛋白;不溶性的蛋白质构成膜的本体,其中一部 分是镶嵌蛋白, 也有一些是酶蛋白。 • ● 脂类 线粒体的脂类只占干重的20~30%。在线粒体的 脂类中多数是磷脂, 占总脂的3/4以上。含丰富的心磷脂 和较少的胆固醇是线粒体在组成上与细胞其他膜结构的明 显差别。 • 线粒体内、外膜在化学组成上的主要区别是脂类和蛋白质 的比例不同, 内膜上的脂类与蛋白质的比值低(0.3:1), 外膜中的比值较高(接近1:1)。 •
个质子,同时转移2个质子至膜间隙。所以复合物Ⅳ既是电子
传递体又是质子移位体。
•呼吸链中复合物的排列及功能
线粒体呼吸链中四种复合物的性质
复合物 序号 名称 Ⅰ Ⅱ NADH脱氢酶 琥珀酸脱氢酶 多肽数 22~26 4~5 电子传递 辅基 1个FMN 6-9个Fe/S 中心 1个FAD 3个Fe/S中心 接收自 NADH 琥珀酸 (经由酶结 合的FAD) 传递给 辅酶Q 辅酶Q 传递 质子 是 否
线粒体结构模式图
外膜:具有孔蛋白构成的亲水通道,允许小分子物质自由通 过。标志酶为单胺氧化酶。
内膜:内膜内陷形成嵴(cristae)来扩大内膜表面积。嵴
有两种类型,板层状和管状。心磷脂含量高,通透性 很低,H+和ATP等不能自由通过,必需有载体蛋白
和通透酶参与。嵴膜上有基粒,基粒由头部(F1偶
联因子)和基部(F0偶联因子)构成。 膜间隙:内外膜之间的间隙,延伸到嵴的轴心部。标志酶为
• ● 标志酶 • 通过细胞化学分析, 线粒体各部位有特征 性的酶, 称为标志酶: • • • • 外膜: 单胺氧化酶 内膜: 细胞色素氧化酶 膜间隙: 腺苷酸激酶 基质: 苹果酸脱氢酶
二、线粒体的功能
线粒体的主要功能是氧化磷酸化,合成ATP,为细胞的 生命活动提供能量。
糖、脂肪
细胞质
丙酮酸
线粒体 乙酰coA(三羧酸循环)
电子传递链中几种电子载体及电子传递
三、氧化磷酸化的作用机理
(一)质子动力势 对于氧化磷酸化的偶联机制提出的假说有化学偶联假说、构 象偶联假说、化学渗透假说等。 “化学渗透假说”,是1961年 英国生物化学家Mitchell提出来的,取得大量实验结果的支持,
成为一种较为流行的假说。 内容:当电子沿呼吸链传递时,所释放的能量将质子从内膜 基质侧泵至膜间隙,由于线粒体内膜对离子是高度不通透的, 从而使膜间隙的质子浓度高于基质,在内膜的两侧形成pH梯度 (△pH)及电位梯度(Ψ),两者共同构成电化学梯度,即质 子动力势(proton-motive force, △P)。 △P=Ψ-(2.3RT/F)△pH 其中T为绝对温度,R为气体常数,F为法拉第常数。 质子沿电化学梯度穿过内膜上的ATP酶复合物流回基质,使 ATP酶的构象发生改变,将ADP和Pi合成ATP。
色,发现线粒体具有氧化作用。
1904年,在植物细胞中发现了线粒体。 至20世纪50年代,证实三羧酸循环,氧化磷酸化和脂肪酸 氧化等重要的能量代谢过程均发生在线粒体中。 现在线粒体的结构和功能的研究已经深入到分子水平。
一、形态结构
(一)形态与分布
形状:线粒体一般呈粒状或杆状。
化学组成:蛋白质和脂类。 大小:一般直径0.5~1μm,长1.5~3.0μm,在胰脏外分泌 细胞中可长达10~20μm,称巨线粒体。 数量及分布:植物细胞少于动物细胞;许多哺乳动物成熟的
酶复合物。作用是催化NADH的2个电子传递至辅酶Q,同时发生质子的跨膜
输送,将4个质子由线粒体基质(M侧)转移至膜间隙(C侧)。所以复合物 Ⅰ既是电子传递体又是质子移位体。
电子传递的方向为:NADH→FMN→Fe-S→Q
复合物Ⅱ 琥珀酸-CoQ还原酶又称琥珀酸脱氢酶,有4条多肽链组成。含有一个FAD, 2个铁硫蛋白和一个细胞色素b,作用是催化电子从琥珀酸通过FAD和铁硫蛋 白传给辅酶Q。复合物Ⅱ不能使质子跨膜移位。 电子传递的方向为:琥珀酸→FAD→Fe-S→Q
肌细胞和精子的尾部聚集较多的线粒体, 以提供能量
线粒体包围着脂肪滴,内有大量要被氧化的脂肪
• ● 存在方式 • 线粒体在细胞中并非都是单个存在的,有 时可形成由几个线粒体构成的网络结构, 有些线粒体具有分支,可以相互交错在一起。 如通过相差显微镜检查完整的肝细胞,发现 线粒体并非是单个存在的,而是以交织的网 络状态存在。
细胞色素
三个铜原子:类似于铁硫蛋白的结构,通过
Cu2+、Cu1+的变化传递电子。
铁硫蛋白:在其分子结构中有2Fe-2S和4Fe4S两种类型。 通过Fe2+ 、 Fe3+互变进行电子 传递。 辅酶Q:是脂溶性小分子量的醌类化合物,通 过氧化和还原传递电子。有3种氧化还原形式即 氧化型醌Q,还原态QH2和自由基半醌(QH)。
吸链和FADH2呼吸链。复合物Ⅰ、Ⅲ、Ⅳ组成NADH呼吸链,
催化NADH的脱氢氧化,复合物Ⅱ、Ⅲ、Ⅳ组成FADH2呼吸链, 催化琥珀酸的脱氢氧化。 呼吸链各组分的排列是高度有序的使电子按氧化还原电位从 低向高传递,呼吸链中有三个部位有较大的自由能变化,足以 使ADP与无机磷结合形成ATP。部位Ⅰ在NADH至CoQ之间。 部位Ⅱ在细胞色素b和细胞色素c之间。部位Ⅲ在细胞色素a和 氧之间。
细胞中线粒体分支交织连接而成的网状二维结构
(二)超微结构
线粒体(mitochondrion)由内、外两层彼此平
行和高度特化的膜包围而成的封闭的囊状结构,
内外膜都是典型的单位膜。
包括:外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)和基 质(matrix)四个功能区隔 。
• ■ 线粒体各部分的特性和功能 • ● 蛋白分布: • 线粒体由四个部分组成,在能量转换过程 中分别起不同的作用。各部分功能的差异 主要是化学组成的差异,特别是蛋白和酶 分布的差异。