第四章 细胞质膜
第四章 细胞质膜
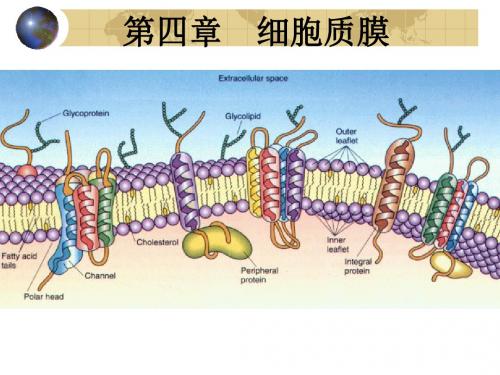
影响中枢神经系统,甚至危及生命。
糖脂
其极性头部共价结合到鞘氨醇上的糖分子或寡糖链。 普遍存在于原核和真核细胞的质膜上,约占膜脂总 量的5%以下,但神经细胞膜上糖脂含量较高,约 5-10%。 最简单的糖脂是半乳糖脑苷脂;最复杂的糖脂是神 经节苷脂。家族性白痴病和神经节苷脂的异常降解 有关。
糖脂,是鞘氨醇的衍生物
膜的流动性低于一定的阈值时,许多酶的活动和跨
膜运输将停止.
流动性过高,又会造成膜的溶解。
(二)细胞膜的不对称性
膜内外两层组分和功能的差异,称膜的不对称性。 (二)细胞膜的不对称性 样品经冰冻断裂处理后,各断面命名为:ES,细胞外表面;
EF ,细胞外小页断面; PS ,原生质表面; PF ,原生质小
甘油磷脂
(3-磷酸甘油的衍生)
2、鞘脂
主要在高尔基体合成。 是含有鞘氨醇骨架的两性脂,具有一条 长链脂肪酸,一条烃链 ,和一个头部。 根据头部基团的不同,可分鞘磷脂、脑 苷脂以及神经节苷脂三种;其中,鞘磷 脂中含有磷酸基团,脑磷脂和神经节苷 脂中含糖残基而不含磷酸基团。
鞘 磷 脂 p56
鞘磷脂(sphingomyelin)是最普通的鞘脂,其极 性头是磷酰胆碱或磷酰乙醇胺。 在高等动物的组织中含量极为丰富,细胞内鞘磷脂 的水解有赖于磷脂酶,磷脂酶的先天性缺损便导致 鞘磷脂在组织内堆积,可造成肝、脾肿大,严重者
白比较容易纯化、分析。
红细胞经过低渗处理,质膜破裂,内容物释放,留下一 个保持原形的壳,称为红细胞影(血影)。红细胞是研究膜 骨架的理想材料。
(二)红细胞质膜蛋白及膜骨架
SDS聚丙烯酰胺凝胶电泳分析:血影的蛋白成 分包括:血影蛋白、锚蛋白、带3蛋白、带4.1 蛋白、肌动蛋白和血型糖蛋白。
第4章-细胞质膜(翟中和第四版)

6197–202.
Simons K, Ikonen E (June 1997). "Functional rafts in cell membranes". Nature,387 (6633): 569–72.
精选可编辑ppt
13
脂筏结构模型
脂筏模型(lipid rafts):脂质双分子层不是一个完全 均匀的二维流体,膜中富含胆固醇和鞘磷脂的微区, 其中聚集一些特定的蛋白质区。 特点:这些区域比膜的其他部分厚,更有秩序且较 少 流动性。其周围是流动性较高的液态区。 功能:参与信号转导、受体介导的内吞作用以及胆固 醇代谢运输等。脂笩功能的紊乱涉及多种疾病的发生。
帽现象(capping)
Figure 10-35 Molecular Biology of the Cell (© Garland Science 2008)
精选可编辑ppt
41
(二)膜蛋白的流动性
• 膜蛋白流动性受 多种因素限制: 如紧密连接、细 胞骨架的影响等
精选可编辑ppt
42
(三)膜脂和膜蛋白运动速率的检测
翟中和 王喜忠 丁明孝 主编
细胞生物学(第4版)
第4章 细胞质膜
1
细胞表面是复合的结构体系与多功能的体系,细胞膜是细胞表
面的核心结构。
精选可编辑ppt
2
基本概念
• 细胞质膜(plasma membrane)又称质膜,曾称 细胞膜(cell membrane),是围绕在细胞最外层 ,由脂质、蛋白质和糖类组成的生物膜。
• 脂筏基本结构成分, 还是很多重要生物活 性分子的前体化合物
Figure 10-4 Molecular Biology of the Cell (© Garland Science 2008)
细胞生物学 名词解释 第四章 细胞质膜

lipid anchored protein
通过与之共价连接的脂分子插入膜的脂双分子层中,从而锚定在细胞质膜上。与脂肪酸结合的脂锚定蛋白分布在质膜内测,与糖脂结合的脂锚定蛋白分布在质膜外侧,GPI脂锚定膜蛋白分布在质膜外侧。
研究膜蛋白的分析技术
低温电镜、X射线晶体衍射技术、电镜三维重构
研究膜蛋白的难处
1、表达量低
2、分离纯化困难
3、难以形成三维晶体
细胞膜流动性பைடு நூலகம்
胆固醇含量减低,脂肪酸链短,不饱和度高,相变温度低,PC/SM比例高,流动性大(胆固醇即可增强流动性,又可降低流动性),由此可得流动性与胆固醇含量、脂肪酸链长短、饱和程度、PC/SM比值有关。
研究膜蛋白流动性(侧向运动)的实验
荧光抗体免疫标记细胞融合:用抗鼠细胞质膜蛋白的荧光抗体(绿色荧光)和抗人细胞质膜蛋白的荧光抗体(红色荧光)分别标记小鼠和人的细胞表面,然后用灭活的仙台病毒介导两种细胞融合,10min后,不同颜色的荧光开始在融合细胞的表面扩散,40min后分辨不出融合细胞表面的绿色荧光和红色荧光区域,两种荧光均匀的分布在融合细胞表面。这一实验证明膜蛋白在脂膜上的运动。
带3蛋白
band3 protein
是红细胞质膜Cl-/HCO3-阴离子运输的载体蛋白,带3蛋白的N末端伸向细胞质膜基质面折叠成不连续的水不溶性的区域,为膜支架蛋白提供结合位点,多次跨膜蛋白,内在膜蛋白
细胞质膜的功能
1、为细胞的生命活动提供相对稳定的内环境,2、选择性的物质运输,3、提供细胞识别位点,4、为多种酶提供结合位点;5、介导细胞与细胞,细胞与胞外基质之间的连接;6、质膜参与形成具有不同功能的细胞表面特化结构;7、膜蛋白的异常与某些遗传病、恶性肿瘤、自身免疫病甚至神经退行性疾病有关,很多膜蛋白可作为疾病治疗的药物靶标
第4章 细胞质膜 (西安农林科技大学)

第四章细胞质膜•细胞质膜是围绕细胞最外层的一层由脂质和蛋白质组成的生物膜4.1 细胞膜的结构模型与基本成分膜。
•质膜提供给细胞一个相对稳定的内环境①。
在物质运输①③,能量转换③,信息传递④⑤,细胞连接和黏着⑤等方面有重要作用。
用•真核细胞内部存在由膜构建的各种细胞器,它们有与质膜结构类似的生物膜②。
Singer 和Nicolson 的流动镶嵌模型是膜生物学的核心理论。
膜的流动性(流动):膜脂处于流动状态,膜脂和膜蛋白均处于动态运动中,可侧向运动。
膜蛋白分布的不对称性(镶嵌):膜蛋白分布的不对称性(镶嵌)膜蛋白以多种方式镶嵌于脂双分子层内、贯穿其中或附着在脂层表面。
三明治模型和流动镶嵌模型。
电镜超薄切片显示的细胞膜生物膜模型:A,流动镶嵌;B,生物膜结构示意100的质膜凹陷区也富含•膜泡窖:50~100nm 的质膜凹陷区。
也富含胆固醇、鞘脂和糖基磷脂酰肌醇脂锚定膜蛋白。
含有窖蛋白。
膜泡窖:transient confinement 瞬时封闭,caveolae 膜泡窖,caveolin 窖蛋白,nanodomain 超微区,GPI 糖基磷脂酰肌醇,fluid lipid 流动脂。
脂筏形成机制:蛋白亚基聚集导致与其连接的相对稳定的膜脂微区域聚集。
ligand 配体,lectin 凝集素,oligomerization 低聚化,adapter 接头蛋白,scaffold 支架蛋白。
3. 生物膜可看成是蛋白质在双分子层中的二维溶液;但存在脂筏等结构功能单位;4. 生物膜在三维空间上可弯曲、折叠、延伸等改变,处于动态变化之中。
4.1.2膜脂:膜脂为两性分子兼具亲水性和疏水性。
■膜脂主要有三种:磷脂、糖脂和胆固醇。
●磷脂:甘油磷脂和鞘氨醇磷脂。
约占膜脂的50%以上。
神经系统含鞘磷脂较丰富。
主要特征:•一个个极性头、两个非极性尾(脂肪酸链)。
•脂肪酸碳链为偶数,16,18或20个碳原子。
•常含有不饱和脂肪酸(如油酸)。
第四章细胞质膜-2010-课件

冰冻蚀刻技术 揭示的膜结构
12
其他的细胞膜结构模型
是对流动镶嵌模型的充实 ,完善和补充
液晶态模型 板块镶嵌模型 脂筏模型
脂筏模型 脂筏区域富含胆固醇
生物膜结构的特征
1 磷脂双分子层是组成生物膜的基本结构成分,尚未 发现膜结构中起组织作用的蛋白;
2 蛋白分子以不同方式镶嵌在脂双层分子中或结合 在其表面,膜蛋白是赋予生物膜功能的主要决定者;
How Proteins Associate with the membrane
①、②整合蛋白;③、④脂锚定蛋白;⑤、⑥外周蛋白
外周蛋白
• 外周蛋白靠离子键或其它较弱的键与膜表面蛋白 或脂分子结合,改变溶液的离子强度、提高温度 就可以从膜上分离下来。
• 一个蛋白可以由多个亚基构成,有的亚基为跨膜 蛋白,有的则结合在膜的外部。
Hale Waihona Puke 一、生物膜的结构模型1890s,E.Overton 发现脂溶性物质容易透过 细胞, 推测细胞膜由连续的脂类物质组成。
• 1925年, 荷兰的两位科学家E.Gorter和 F.Grendel根据对红细胞的研究,提出质膜 的脂双层(lipid bilayer)结构
电镜下的红细胞
实验结果证明∶红细胞膜脂与表面 积之比约为1.8~2.2∶1。
精品
第四章细胞质膜-2010
细胞质膜的结构模型 生物膜基本特征与功能 膜骨架
第一节 细胞质膜的结构模型
细 胞 膜 (cell membrane), 又 称 质 膜 (plasma membrane)指围绕在细胞最外层, 由脂类和蛋白质组 成的薄膜.
细胞内膜 生物膜(biomembrane) 。 细胞膜的结构模型(流动镶嵌模型) 膜脂——生物膜的基本组成成分 膜蛋白
040细胞质膜:
①
②
③ ③④脂锚定膜蛋白 ③④脂锚定膜蛋白
⑤ ⑤⑥外周膜蛋白 ⑤⑥外周膜蛋白
①②膜整合蛋白 ①②膜整合蛋白
膜蛋白类型
外在膜蛋白 或外周膜蛋白 extrinsic or peripheral membrane protein
内在膜蛋白 或整合膜蛋白 intrinsic or integral membrane protein
血影蛋白 锚定蛋白 带3蛋白 蛋白 带4.1蛋白 蛋白 肌动蛋白 血型糖蛋白
1 膜脂分子的运动
第四章 细胞质膜
第二节 细胞膜基本特征
影响膜脂流动性的因素
胆固醇 胆固醇的含量增加会降低膜的流动性。 胆固醇的含量增加会降低膜的流动性。
脂肪酸链饱和度 脂肪酸链所含双键越多越不饱和,流动 脂肪酸链所含双键越多越不饱和, 性增加。 性增加。 脂肪酸链链长 卵磷脂/ 卵磷脂/鞘磷脂 长链脂肪酸相变温度高,膜流动性降低。 长链脂肪酸相变温度高,膜流动性降低。 比例高则膜流动性增加, 比例高则膜流动性增加,是因为鞘磷脂 粘度高于卵磷脂。 粘度高于卵磷脂。 温度、酸碱度、离子强度等。 温度、酸碱度、离子强度等。
第四章 细胞质膜
第一节 细胞膜结构膜
Phospholipids
第四章 细胞质膜
第一节 细胞膜结构膜
磷脂类型
-CH2-CH-N+3 I COOH 磷酯酰丝氨酸
-CH2-CH2-N+3 O C O CH2 O C O CH O H 2C O P O OCH2 CH2 CH3
磷脂酰乙醇胺: 磷脂酰乙醇胺: 脑磷脂
EF
PF
第四章 细胞质膜
第二节 细胞膜基本特征
1 膜脂的不对称性 同一种脂分子在膜的脂双层中呈不均匀分布, 同一种脂分子在膜的脂双层中呈不均匀分布,质膜的内 外两侧分布的磷脂的含量比例也不同。 外两侧分布的磷脂的含量比例也不同。
细胞生物学第四章细胞质膜

最丰富和最具特性的脂。它有一个极性的头部 和一个疏水的尾部。
(internal membrane), 习惯上把细胞所有膜 结构统称为 生物膜 (biomembrane)。
细胞生物学第四章细胞质膜
虽然细胞很早就在光镜下被发现,但其是否有明确的边界结构,尚 未可知,直到电镜发明发现质膜的超微结构,但人们并不惊奇,因 为在此之前已侦知。
1、关于膜的化学组成的早期研究: 18世纪90年代,Overton 用植物的根毛作实验,
第四章、细胞质膜
知识要点: 1 、掌握几种膜分子结构模型学说,并评价之。 2 、了解膜结构的组成成分和组成方式。 3 、理解质膜流动性和不对称性两大特点。 4、了解红细胞膜骨架的组成和功能。
细胞生物学第四章细胞质膜
第一节、细胞膜结构模型与成分 一、细胞膜结构
膜(membrane)是细胞 的重要结构, 包括细 胞质膜 (plasma membrane)、内膜
高尔基体膜)、细胞类型(肌细胞、肝细胞)、生物类型(动物、植物 和原核生物)的不同而不同。
一般而言ห้องสมุดไป่ตู้膜脂占50%,蛋白质占40%,碳水化合物约1-10%
细胞生物学第四章细胞质膜
1、膜脂
膜脂都具有双亲性,这种性质使生物膜具有屏 障作用,大多数水溶性物质不能自由通过,只 允许亲脂性物质通过。
膜脂是生物膜的基本组成成分, 主要有三大类 型:磷脂、糖脂、胆固醇。
质膜的片层结构模型
细胞生物学第四章细胞质膜
5、单位膜模型(unit membrane model)
1959年,J.D.Robertson在电子显微镜下发现细胞膜是类似 铁轨结构,两条暗线被一条明亮的带隔开,显示暗—明— 暗三层,总厚度为7.5nm,中间层为3.5nm,内外两层各为 2nm。并推测:暗层是蛋白质,透明层是脂,并建议将这种 结构称为单位膜。
第四章 细胞质膜 南开大学细胞生物学课件

鞘氨醇 神经酰胺
神经鞘磷脂,SM 脑苷脂A
神经节苷脂A
ABO 血 型 糖
3. 固醇
胆固醇及其类似物统称固醇。胆固醇是中性脂类,存在于动 物细胞和少数植物细胞质膜上,含量不超过膜脂的1/3。胆固 醇包括三部分:羟基团代表极性的头部,非极性的甾环结构和 一个非极性的碳氢尾部。胆固醇在调节膜的流动性、加强膜的 稳定性、降低水溶物质的通透性方面都起重要作用。
(3)生物膜可以看成是蛋白质在双层脂分子中的二维溶液。 即膜的流动性。
(4)在细胞生长和分裂等整个生命活动中,生物膜在三维空 间上可出现弯曲、折叠、延伸等改变,处于不断的动态变化中。
二、膜脂
通过对血影的分析,一般情况下: 脂 类 占 40% 蛋白质 占 50% 糖 类 占 1-10%
膜中脂类和蛋白质的含量变化与膜的功能有关。膜中含蛋白 质越多,膜的功能越复杂;所含的蛋白质种类和数量越少,膜 的功能越简单。如神经髓鞘,它的作用是起绝缘作用,所以蛋 白质的含量显著低于脂类。
(二)膜脂的不对称性
膜脂的不对称性是指同一种膜脂分 子在膜的脂双层中呈不均匀分布。 糖脂的分布表现出完全不对称性
(三)膜蛋白的不对称性
膜蛋白的不对称性是指每种膜蛋白分子在质膜上都 具有确定的方向性;
所有膜蛋白在质膜上都呈不对称分布,膜蛋白的不 对称性是生物膜完成复杂的在时间与空间上有序的各种 生理功能的保证。
2. 内在膜蛋白
(intrinsic membrane protein)
又称整合膜蛋白(integral membrane protein)。 整合膜蛋白均为双性分子,非极性区插在脂双层分子之间,极性区则朝 向膜的表面,它们通过很强的疏水或亲水作用力同膜脂牢固结合,一般不易 分离开来,只有用去垢剂(detergent)使膜崩解后才可分离出来。 内在膜蛋白占整个膜蛋白的70%~80%。估计人类基因组中,1/4~1/3基因 编码的蛋白为内在膜蛋白。
细胞质膜优秀课件

4.1.5 细胞膜的功能
细胞生物学
细胞膜与细胞表面
• 为细胞的生命活动提供相对稳定的内环境 • 物质和能量的交换 • 细胞内外信息跨膜传递 • 提供识别和结合位点 • 介导细胞与细胞、细胞与基质间的连接
• 参与形成具有不同功能的细胞表面特化结 构
细胞生物学
细胞膜与细胞表面
• 第三节 膜骨架
膜糖脂的功能
可以提高膜的稳定性,增强膜蛋白对细胞 外基质中蛋白酶的抗性,帮助膜蛋白进行正确 的折叠和维持正确的三维构型。
参与细胞的信号识别、细胞的粘着。 有些糖蛋白中的糖基还帮助新合成蛋白质 进行正确的运输和定位。
例如 ABO血型抗原,它是一种糖脂, 其寡糖部分 具有决定抗原特异性的作用。
A型: 红细胞膜脂寡糖链的末端是N -乙酰半乳糖胺 (GalNAc)
4.1.4 膜的不对称性(membrane asymmetry)
细胞质膜的不对称性是指细胞质膜脂双层 中各种成分不是均匀分布的,包括种类和数 量的不均匀。
■ 不对称性的表现 ● 膜脂的不对称性; ● 膜蛋白的不对称; ● 膜糖脂的不对称 。
不对称性:生物膜各膜面的名称细胞膜与细胞表面
细胞生物学
• ES (extro’cytoplasmic surface)细胞外表面
●人、鼠细胞融合实验
细胞生物学
细胞膜与细胞表面
人-鼠细胞融合实验
小鼠-人细胞融合过程中膜蛋白的侧向扩细散胞膜与细胞表面
细胞生物学
● 淋巴细胞的成斑和成帽反应 细胞膜与细胞表面 细胞通生物过学抗体交联膜蛋白分子聚集成斑(patching)、成 帽(capping)的现象也是证明膜蛋白在膜平面侧向扩散 的例子。
细胞生物学
细胞膜与细胞表面
细胞生物学第四章复习

第四章:细胞质膜一、名词解释:1、细胞质膜:由脂质、蛋白质、糖类组成的生物膜2、生物膜:细胞内的膜系统和细胞质膜的统称3、去垢剂:一端亲水、一端疏水的两性小分子4、膜骨架:是指细胞质膜下与膜蛋白相连的由纤维蛋白组成的网架结构二、基本知识:1、细胞质膜的作用:质膜在结构上作为细胞的界膜,使细胞具有相对稳定的内环境,同时又是细胞与环境之间的物质运输、能量转换和信息传递之间发挥重要作用。
2、细胞质膜模型的提出历程:三明治式(蛋白质—脂质-蛋白质)单位膜模型流动镶嵌模型脂筏模型3、目前对生物膜结构的认识磷脂分子在水相中具有自发封闭的性质蛋白质分子以不同方式镶嵌在脂双层分子中或结合在其表面生物膜可看成是蛋白质在双层脂分子中的二维溶液细胞生命活动中,生物膜处于不断的动态变化中4、膜脂成分:甘油磷脂、鞘脂、固醇运动方式:沿膜平面的侧向运动脂分子围绕轴心的自旋运动脂分子尾部摆动双层脂分子之间的翻转运动5、膜蛋白外在膜蛋白内在膜蛋白(胞质外、跨膜和胞质内结构域)成分A:脂肪酸结合到膜蛋白N 端锚定膜蛋白B:烃链结合到膜蛋白C 端C:通过糖脂锚定在细胞质膜上6、去垢剂离子型去垢剂(作用比较剧烈,使蛋白质变性)非离子型去垢剂(作用比较温和)7、膜的基本特性:①膜的流动性脂肪酸链越短、不饱和程度越高,流动性越大膜脂的流动性温度对膜脂的运动有明显影响胆固醇对膜的流动性起着双重调节作用膜蛋白的流动性(用荧光漂白恢复技术能推算膜脂和膜蛋白的扩散速度)②膜的不对称性细胞质膜各膜面的名称质膜的细胞外表面(ES)质膜的原生质表面(PS)细胞外小叶断裂面(EF)原生质小叶断裂面(PF)膜脂的不对称性(糖脂分布细胞质膜外侧、细胞器膜内侧)膜蛋白的不对称性8、膜骨架与红细胞成熟的红细胞没有细胞器,质膜是它的惟一结构,并且易于提纯和分离,是研究膜和膜骨架的最好材料。
9、膜的基本功能为细胞的生命活动提供较为稳定的内部环境参与物质运输与能量传递为许多酶提供结合位点,使酶促反应高效有序进行。
细胞生物学-4第四章质膜

七、细胞质膜的功能
为细胞的生命活动提供相对稳定的内环境; 选择性的物质运输,包括代谢底物的输入与代谢
产物的排除,其中伴随着能量的传递; 提供细胞识别位点,并完成细胞内外信息跨膜传递; 为多种酶提供结合位点,使酶促反应高效而有序地进行; 介导细胞与细胞、细胞与基质之间的连接; 质膜参与形成具有不同功能的细胞表面特化结构。
脂质体的应用
研究膜脂与膜蛋白及其生物学性质; 脂质体中裹入DNA可用于基因转移; 在临床治疗中,脂质体作为药物或酶等载体
三、膜蛋白
基本类型 内在膜蛋白与膜脂结合的方式 外在膜蛋白与膜脂结合的方式 去垢剂(detergent)
基本类型
外在(外周)膜蛋白(extrinsic/peripheral membrane proteins )
膜的不对称性
细胞质膜各部分的名称 膜脂与糖脂的不对称性
糖脂仅存在于质膜的ES面,是完成其生理功能的结构基础 膜蛋白与糖蛋白的不对称性
膜蛋白的不对称性是指每种膜蛋白分子在细胞膜上都 具有明确的方向性; 糖蛋白糖残基均分布在质膜的ES面(GO+3HBH4 labeling); 膜蛋白的不对称性是生物膜完成复杂的在时间与空间上 有序的各种生理功能的保证。
糖脂:糖脂普遍存在于原核和真核细胞的质膜上(5%以下),神经细 胞糖脂含量较高;
胆固醇:胆固醇存在于真核细胞膜上(30%以下),细菌质膜不含有 胆固醇,但某些细菌的膜脂中含有甘油脂等中性脂类。
运动方式
沿膜平面的侧向运动(基本运动方式),其扩散 系数为10-8cm2/s;相当于2μm/s
脂分子围绕轴心的自旋运动; 脂分子尾部的摆动; 双层脂分子之间的翻转运动,发生频率还不到
第四章 细胞质膜 (Cell membrance)
第四章 细胞质膜
The basic structural unit of biological membranes is a phospholipid bilayer
● Phospholipids-the most abundant lipids in cell membranes ● Each phospholipid has a hydrophilic (“waterloving”) head and one or two hydrophobic (“water-hating”) hydrocarbon tails
the plasma membrane ● All of the plasma membrane and the internal membranes
are composed of lipids and proteins, share a common general
structure--lipid bilayer (脂双层) penetrated by proteins ● Lipid bilayer is selective permeable(选择透性)-semipermeability
Cell membranes acts as selective barriers
Figure
Cell membranes acts as selective barriers (selective permeable).
Membranes serve as barriers between two compartments-either between the inside and the outside of the cell (A),or between two intracellular compartments (B). In either case the membrane prevents molecules on one side from mixing with those on the other side
第四章 细胞质膜

膜脂— 二、膜脂—生物膜的基本组成成分
磷脂、 胆固醇三种类型 (一)成分:主要包括磷脂、糖脂 胆固醇三种类型。 成分:主要包括磷脂 糖脂和胆固醇三种类型。 磷脂(phospholipid):膜脂的基本成分(50%以上); 1、 磷脂 ( phospholipid) 分为二类:甘油磷脂和鞘磷脂,其中甘油磷脂又包括: 磷脂酰胆碱(卵磷脂)、磷脂酰丝氨酸、磷脂酰乙醇胺 (脑磷脂)和磷脂酰肌醇等,磷脂的主要特征:
根据Fluid-mosaic model: 根据 :
1. 2.
细胞膜由流动的双脂层和嵌在其中的蛋白质组成。 细胞膜由流动的双脂层和嵌在其中的蛋白质组成。 流动的双脂层和嵌在其中的蛋白质组成 磷脂分子以疏水性尾部相对, 磷脂分子以疏水性尾部相对,极性头部朝向水相组成生 物膜骨架; 物膜骨架;
3.
蛋白质或嵌在双脂层表面,或嵌在其内部, 蛋白质或嵌在双脂层表面,或嵌在其内部,或横跨整个 双脂层,表现出分布的不对称性 不对称性。 双脂层,表现出分布的不对称性。
甘油磷脂
以甘油 甘油为骨架的磷脂类,在骨架上结合两个脂肪酸 两个脂肪酸链一 甘油 两个脂肪酸 个磷酸基团 磷酸基团,胆碱、乙醇胺、丝氨酸或肌醇等分子靠磷 磷酸基团 酸基团连接到脂tidylcholine,PC,旧称卵磷脂 磷脂酰丝氨酸phosphatidylserine,PS 磷脂酰乙醇胺phosphatidylethanolamine,PE,旧称脑磷脂 磷脂酰肌醇phosphatidylinositol,PI 双磷脂酰甘油Diphosphatidylglycerol, DPG,旧称心磷脂
具有一个极性头部和两个非极性尾部(心磷脂除外) ① 具有一个极性头部和两个非极性尾部(心磷脂除外); 脂肪酸碳链碳原子为偶数,大多数由16 18或20个组成 16, 个组成; ② 脂肪酸碳链碳原子为偶数,大多数由16,18或20个组成; 含有饱和脂肪酸(如软脂酸)和不饱和脂肪酸(如油酸) ③ 含有饱和脂肪酸(如软脂酸)和不饱和脂肪酸(如油酸);
第四章 细胞质膜

2. 三明治模型
• J. Danielli & H.
Davson 1935 发现质 膜的表面张力比油- 水界面的张力低得多, 提出三明治模型(蛋 白质-脂类-蛋白质)。
3. 单位膜模型
• JD. Robertson 1959
根据电镜观察提出单 位 膜 模 型 。 厚 约 7.5 nm。
5. 脂筏 ( lipid raft )
• 生物膜上富含鞘磷脂和胆固醇的相对有 序的微小区域,与生物膜某些特定功能 的发挥有关。
细胞膜的脂筏模型示意图
病毒出芽过程中细胞质膜的动态变化
二、膜脂
(一)、成分
(二)、膜脂的运动方式
(三)、脂质体
(一)、成分
1. 甘油磷脂 2. 糖脂
3.胆固醇
膜脂的基本类型
• 胆固醇起束尾(与磷脂尾部结合)和疏开(将磷脂分
子隔开)的双重作用,但通常胆固醇起到防止膜脂由
液相变成固相以保证膜脂处于流动状态的作用。
膜脂的流动性主要由脂分子本身的性质决定。
(二)、膜蛋白的流动性
• 荧光抗体免疫标记实验:两种荧光标记→细胞融合→对
半→均匀→成斑→成帽 • 膜蛋白在脂双层二维溶液中的运动是自发的热运动 • 细胞骨架既限制膜蛋白的运动,也影响其周围的膜脂的
电的极性头形成离子键,或带负电的氨基酸残基通过 Ca2+、
Mg2+ 等阳离子与带负电的磷脂极性头相互作用。
• 某些膜蛋白在细胞质基质一侧的半胱氨酸残基上共价结
合脂肪酸分子 , 插入脂双层之间 , 进一步加强膜蛋白与脂
双层的结合力,还有少数蛋白与糖脂共价结合。
内在膜蛋白与膜脂结合方式示意图
2) 跨膜结构域与膜脂的作用方式
第四章细胞质膜

2. 围绕轴心的自旋运动:膜脂分子围绕与膜平面垂直的轴进行快速旋转。 3.脂分子尾部的摆动:膜脂分子围绕与膜平面垂直的轴进行左右摆动。 4.双层脂分子之间的翻转运动:膜脂分子从脂双层的一层翻转到另一
层 。是在翻转酶(flippase)催化下完成发生频率不到脂分子侧向交换频率的 10-10。但在内质网膜上,新合成的磷脂分子翻转运动发生频率很高。
2.脂质体的应用
研究生物膜的特性研究膜脂与膜蛋白及
其生物学性质;
转基因脂质体中裹入DNA可用于基因转移;
制备药物在临床治疗中,脂质体作为药物或
酶等载体。
三、膜蛋白
占核基因组编码蛋白质的30%。 1、基本类型 (1)外在(外周)膜蛋白(extrinsic/peripheral membrane protein ):水溶性蛋白,靠离子键或其它弱键与膜内表面的 蛋白质分子或脂分子极性头部非共价结合,改变溶液的离子 强度、提高温度就可从膜上分离下来。 (2)内在(整合)膜蛋白(intrinsic/ integral membrane protein):整合蛋白为跨膜蛋白,水不溶性蛋白。是两性分 子,形成跨膜螺旋,跨膜结构域为1至多个疏水的α螺旋, 与膜结合紧密,需用去垢剂使膜崩解后才能从膜上洗涤下来。
小窝蛋白
糖基磷脂酰肌醇锚定
细胞膜穴样内陷
过渡限制区
微结构域
二、膜脂——生物膜的基本组成成分
膜脂的成分:膜脂主要包括磷脂、糖脂 和胆固醇三种。
膜脂的运动方式 脂质体(liposome)
(一)膜脂成分
1.磷脂: 膜脂的基本成分(50%以上),包括甘油磷脂和鞘磷脂二 类。 主要特征:①具有一个极性头部和两个非极性的尾部(心磷 脂除外);②脂肪酸碳链碳原子为偶数,多数碳链由16、18 或20个组成;③含饱和脂肪酸(如软脂酸)及不饱和脂肪酸 (如油酸);
细胞生物学-第四章 细胞质膜

水溶性蛋白-脂质-去污剂复合体
可溶性脂质去污剂混合胶粒
第二节 生物膜基本特征与功能
一.膜的流动性 (一)膜脂的流动性
膜的流动性是生物膜的基本特征之一,也是细胞进行 生命活动的必须条件。流动性包括膜脂的流动性和膜蛋白的 流动性。
典型的磷脂分子有1个极性的头部和2个疏水的碳氢链尾
部,当一条烃链中含有一个或几个顺式排列的双键(不饱和 的),在双键部分可形成弯曲。烃链的长度及饱和状态的不 同,影响膜的流动性。
翻转运动
弯曲 弯曲的碳氢链
饱和的碳氢链
胆固醇
(三)脂质体 (1)膜脂都是兼性分子,其分子结构中含有亲水 和疏水两部分。两条脂肪酸链几乎彼此平行,分子 的两端,形成所谓极性的头部和非极性的尾部,头 部具有亲水性而尾部具有疏水性。
(2)在水溶液中会自动形成双分子层结构,即亲水
第一节 细胞质膜的结构模型
一、生物膜的结构模型 (一)片层结构模型
1925年荷兰的2位科学家Gorter和Grendel用有机 溶剂抽提人的红细胞膜的膜脂成分并测定膜脂单层分 子在水面的铺展面积,发现它为红细胞表面积的二倍, 提 示 了 质 膜 是 由 双 层 脂 分 子 构 成 的 。 1935 年 , Daniell&Davson:发现质膜的表面张力比油-水界面的 表面张力低得多,已知脂滴表面如果吸附有蛋白质成 分则表面张力降低,因此推测,质膜中含有蛋白质成 分。从测定膜的表面张力得出细胞膜的“三明治结构 型”,即蛋白质—脂质—蛋白质。
不足之处:(1)单位膜为一种静态单一的结构无
法说明膜的动态结构的变化,(2)各种膜的功能不 一样,而单位膜模型显示不出来,(3)各种不同细 胞和同一细胞中的不同部分膜的厚度实际上并不都 是7.5nm。
第四章 细胞质膜

第四章细胞质膜第一节细胞质膜的结构模型与基本成分一细胞质膜的结构模型发现过程:1925年,E.Gorter和F.Grendel通过对红细胞质膜的研究揭示了质膜是由双层脂分子构成的。
之后,Davson和Danielli推测出质膜中含有蛋白质成分,并提出三明治式模型—蛋白质-脂质-蛋白质1959年,Robertson提出单位膜模型,并大胆推测所有的生物膜都是由蛋白质-脂质-蛋白质的单位膜构成。
这一推测得到了X射线分析结果的支持Singer和Nicolson在1972年提出生物膜的流动镶嵌模型,该模型得到了各种实验结果的支持。
1975年Uuwin和Henderson首次报道了质膜蛋白菌紫红质的三维结构,他是一个跨膜蛋白镶嵌在膜上。
流动镶嵌模型强调:1,膜的流动性,即膜蛋白和膜脂都可以流动;2.膜蛋白分布的不对称性,有的在膜表面,有的镶嵌,有的跨膜。
Simons在1988年提出了脂筏模型。
脂筏模型—在生物膜上胆固醇,鞘磷脂等富集区域形成相对有序的脂相,如同漂浮在脂双层上的脂筏一样载着执行某些特定生物学功能的各种膜蛋白。
生物膜结构归纳总结:1.具有极性头部和非极性尾部的磷脂分子在水相中具有自发形成封闭的膜系统的性质,磷脂分子以疏水性的尾部相对,极性头部朝向水相形成脂双分子层,每层磷脂分子称为一层小叶。
2.蛋白质分子以不同的方式镶嵌在脂双层分子中或结合在其表面,蛋白质的类型,蛋白质分布的不对称性及其与脂分子的协同作用赋予生物膜各自的特性与功能。
3.生物膜可看成是蛋白质在双层脂分子之中的二维溶液。
膜蛋白与膜脂之间,膜蛋白与膜蛋白之间及其与膜两侧其他生物大分子的复杂的相互作用,在不同成度上限制了膜蛋白和膜脂的流动性。
4.在细胞生长和分裂整个生命活动中,生物膜在三维空间上可出现弯曲,折叠,延申等改变,处于不断的动态变化中。
二、膜脂(一)成分1.甘油磷脂构成了膜脂的基本成分,占整个膜脂的一半以上。
是3-磷酸甘油的衍生物.主要在内质网合成主要特征:1,具有一个与磷酸基团相结合的极性头部和两个非极性的尾部,但是存在于线粒体和某些细菌质膜上的心磷脂除外,它有4个尾部。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
3. 胆固醇(cholesterol)
• 主要存在真核细胞膜上,但植物细胞膜中含量较少, 其功能是:调节膜的流动性,增加膜的稳定性,降低 水溶性物质的通透性。胆固醇的结构如下图:
Cholesterol in a lipid bilayer. Schematic drawing of a cholesterol molecule interacting with two phospholipid molecules in one leaflet of a lipid bilayer.
(三)脂质体
• 脂质体(lipsome)是根据磷脂分子可在水相中形成
稳定的脂双层的趋势而制备的人工膜。
(a)水溶液中的磷脂分子团;(b)球形脂质体;(c)平面脂
质体膜;(d)用于疾病治疗的脂质体的示意图
脂质体的应用
研究膜脂与膜蛋白及其生物学性质;
脂质体中裹入DNA可用于基因转移;
在临床治疗中,脂质体作为药物或酶等载体
第四章 细胞质膜
第一节 细胞质膜的结构模型
第二节
第三节
细胞膜基本特征与功能
膜骨架
第一节 细胞质膜的结构模型
一、细胞膜(cell membrane):又称质膜 (plasma membrane),是围绕在细胞表面, 由脂质和蛋白质组成的生物膜。由于它的 存在,使细胞成为一个相对独立的结构, 并通过它使细胞与环境之间能够进行物质、 能量的交换及信息的传递。 对于细胞膜结构和功能的认识,有一个由表 及里、不断深入的有趣历程,产生了几个 结构模型。
(1)甘油磷脂
• 以甘油为骨架的磷脂类,在骨架上结合两个脂肪酸链一
个磷酸基团,胆碱、乙醇胺、丝氨酸或肌醇等分子籍磷
酸基团连接到脂分子上。主要类型有: ① 磷脂酰胆碱 phosphatidylcholine,PC,旧称卵磷脂 ② 磷脂酰丝氨酸 phosphatidylserine,PS ③ 磷脂酰乙醇胺 phosphatidylethanolamine , PE ,旧 称脑磷脂 ④ 磷脂酰肌醇phosphatidylinositol,PI
受到配体时,对脂筏的亲和力低,当结合配体,
发生寡聚化时就会转移到脂筏中。
质膜微囊(Caveolae)
• 质膜微囊是50 年代日本科学家Yamada发现的, 他在电 镜下观察到上皮细胞质膜含有小孔结构, 外貌呈瓶形, 命名为Caveolae (按希腊字原意即为小孔的意思) 。 Caveolae 的分布较广, 尤其是上皮细胞, 内皮细胞, 肌 肉细胞(包括心肌和骨骼肌)等. 它是一种有微囊素 Caveolin 包被的脂质微区. • 质膜微囊在多种细胞生理功能方面发挥重要作用,它 参与大分子物质的跨膜运送及内吞和“胞饮”作用, 也参与胞浆合成膜蛋白的极化运输及信号传递过程。
(一)三明治模型 1. 细胞膜脂双层概念的提出
1925年两位荷兰科学家E.Gorter和F.Grendel分离 纯化了红细胞,从一定数量的红细胞中抽提脂类, 按Langmuir的方法进行展层,并比较展层后的脂 单层的面积和根据体积所推算的总面积, Gorter 和Grendel发现提取的脂铺展后所测的面积同实际
• 脂筏最初可能在高尔基体上形成,然后转移 到细胞膜上。有些脂筏可在不同程度上与膜 下细胞骨架蛋白交联。 • 目前已发现几种不同类型的脂筏,它们在细 胞的信号转导、物质的跨膜运输和蛋白质分 选等过程中均发挥重要作用。
脂筏模型强调细胞膜并不是绝对均匀的流动体系, 而是有分区的,有相对静止的“筏子”。 从脂筏的角度来看,膜蛋白可以分为三类: 1. 存在于脂筏中的蛋白质; 2. 存在于脂筏之外无序液相的蛋白质; 3. 介于两者之间的蛋白质,如某些蛋白在没有接
二、 膜蛋白
膜蛋白是膜功能的主要体现者。核基因组编码的蛋白 质中 30%左右的为膜蛋白。根据与膜脂的结合方式以及 在膜中的位置的不同,膜蛋白分为内在蛋白( integral protein )、外周蛋白( peripheral protein )和脂锚定 蛋白(lipid-anchored protein)。
The parts of a phospholipid molecule. Phosphatidylcholine, represented schematically (A), in formula (B), as a space-filling model (C), and as a symbol (D). The kink due to the cis-double bond is exaggerated in these drawings for emphasis.
(三)脂筏模型 (lipid rafts model)
1997 年由 K.Simons 等人提出,在以甘油磷脂为 主体的生物膜上,胆固醇和鞘磷脂等形成相对有序 的微结构域,大小约55-300nm 左右,就像漂浮在 脂双层上的“脂筏”一样,承载着执行某些特定生 物学功能的某些蛋白质。“脂筏” 是膜上蛋白质停 泊的平台,也是生物膜不易被去圬剂溶解的部分。
(2)鞘磷脂
• 鞘磷脂( sphingomyelin , SM )在脑和神经细胞膜 中含量特别丰富。以鞘胺醇(sphingoine)为骨架,
与一条脂肪酸链组成疏水尾部,亲水头部也含胆碱与
磷酸结合。原核细胞、植物中没有鞘磷脂。
2. 糖脂
•是含糖而不含磷酸的脂类,在膜中约占脂总量的5%
以下,在神经细胞膜上糖脂含量较高,约占 5-10 %。
• 目前对生物膜结构的认识可归纳如下:
• (1)具有偶极性的磷脂分子在水相中具有自发形成封闭膜 系统的性质。尚未发现在生物膜结构中起组织作用的蛋白
。但在脂筏中存在一些有助于其结构相对稳定的功能蛋白
• (2)蛋白分子以不同的方式镶嵌在脂双层分子中火结合在 其表面。 • (3)生物膜可看作是在脂双层分子中嵌有蛋白质的二维溶 液。膜中及其两侧的分子间存在着相互作用。
二、膜脂
(一)成分
膜脂主要包括磷脂、糖脂和胆固醇三种类型。
1.磷脂:包括甘油磷脂和鞘磷脂,是构成膜的主要成分。 磷脂分子的主要特征是: a.具有一个极性头和两个非极性的尾(脂肪酸链),线 粒体内膜上的心磷脂具有4个非16,18或20个碳原子 组成;
c.除饱和脂肪酸外,常含有不饱和脂肪酸(如油酸)。
蛋白与膜的结合方式 ①、②整合蛋白;③、④脂锚定蛋白;⑤、⑥外周蛋白
(一)内在蛋白(integral proteins)
内在蛋白又称为整合蛋白,以不同程度嵌入脂双层的内部 ,有的为全跨膜蛋白(tansmembrane proteins)。膜蛋白为
两性分子。它与膜结合非常紧密,只有用去垢剂(detergent)
A typical single-pass transmembrane protein. Note that the polypeptide chain traverses the lipid bilayer as a right-handed a helix and that the oligosaccharide chains and disulfide bonds are all on the noncytosolic surface of the membrane. Disulfide bonds do not form between the sulphydryl groups in the cytoplasmic domain of the protein because the reducing environment in the cytosol maintains these groups in their reduced (-SH) form.
测量的红细胞的表面积之比约为1.8~2.2∶1,为
了解释这一结果,他们提出红细胞膜的基本结构 是脂双层结构的概念。
Langmuir 的研究工作
2. 随后,人们发现质膜的表面张力比油-水界面的表 面张力低得多,因此推测质膜中含有蛋白质成分,并提 出“蛋白质-脂质-蛋白质”的三明治质膜结构模型。 3. 1959年,Robertson 用超薄切片技术获得了清晰 的细胞膜照片,显示暗-明-暗三层结构,这就为三明治 模型提供了证据,并提出了单位膜模型,认为所有的生 物膜都是由“蛋白质-脂质-蛋白质”的单位膜构成。单 位膜模型是三明治模型发展。
sugar that forms its head group is uncharged. A ganglioside (B) always contains one or more negatively charged sialic acid residues (also called N-acetylneuraminic acid, or NANA), whose structure is shown in (C). Whereas in bacteria and plants almost all glycolipids are derived from glycerol, as are most phospholipids, in animal cells they are almost always produced from sphingosine, an amino alcohol derived from serine, as is the case for the phospholipid sphingomyelin. Gal = galactose; Glc = glucose, GalNAc = N-acetylgalactos-amine; these three sugars are uncharged.
(二)膜脂的运动方式
1. 沿膜平面的侧向运动(基本运动方式) , 其扩散速 率为10-8cm2/s;
2.脂分子围绕轴心的自旋运动;
3.脂分子尾部的摆动;
4.双层脂分子之间的翻转运动,发生频率还不到脂分