松材线虫、松墨天牛和松树间的共生关系
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律松褐天牛(Ips subelongatus)是一种重要的松树害虫,广泛分布于东亚地区。
松褐天牛的幼虫和成虫都以松树为食,尤其是对松树幼树和病弱树的危害更为严重。
松褐天牛也是松材线虫病的传播媒介,加速了松树的死亡。
对松褐天牛在松材线虫病病死树上的垂直分布规律进行研究对于松树病害的防治具有重要意义。
天牛类昆虫对树木的危害在树干内部进行,因此其研究需要对树木的内部结构有较为深入的了解。
松褐天牛在松树上的垂直分布受到多种因素的影响,主要包括天牛自身的生物学特性,树木的生长情况,以及外界环境的影响等。
通过研究这些因素,可以更好地理解松褐天牛在松树上的垂直分布规律,从而为天牛类昆虫的防治提供科学依据。
了解松褐天牛的生物学特性对于研究其在松树上的垂直分布规律非常重要。
松褐天牛有着较高的繁殖力,成虫生活在松树干内,通过食用树木的组织和产卵来完成其繁殖过程。
研究发现松褐天牛在松树上的垂直分布规律与松树的生长情况密切相关。
松褐天牛在松木内部选择的营造区域位置个体发育多表现为有规律的垂直分布。
在一株松木上,松褐天牛的幼虫和成虫会选择特定的部位进行活动,例如在树干的上部、中部或下部。
通过对松褐天牛在松树上的垂直分布规律的研究,可以发现其对不同部位的生长情况有不同的偏好。
松树的生长情况对于松褐天牛在松树上的垂直分布也有着重要影响。
松褐天牛的生长和繁殖需要松木提供充足的养分和适宜的环境条件。
而树木的生长情况往往会受到外界环境的影响,例如土壤的肥沃程度、水分和光照等因素。
研究发现松褐天牛在松树上的垂直分布规律也需要考虑树木的生长情况。
一般来说,树木生长旺盛的部位会更容易受到松褐天牛的危害,因为这些部位提供了更充足的养分和生长条件。
通过对松褐天牛在松树上的垂直分布规律的研究,可以发现其在树木内部对生长情况的偏好,从而为预防和控制天牛类昆虫提供科学依据。
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律松褐天牛是一种松树的天敌,在松木枯死的树干上有一种松材线虫病则是它们的主要宿主,然而它们在这种环境下的垂直分布规律一直以来是一个研究的焦点。
本文将结合相关研究成果,探讨松褐天牛在松木枯死树干上的垂直分布规律。
首先我们需要了解一下松褐天牛和松材线虫病。
松褐天牛(Ips subelongatus Motschulsky)是一种松木的天敌,喜欢寻找松树已经感染松材线虫病并即将死亡的树干为活动区域,通过其嗅觉和视觉等感知系统定位病树,进而繁殖。
而松材线虫病是由松材线虫引起的松树内生病害,病树表现为树干树皮腐朽、树干树皮上有蓝色隧道、树脂外流或挂,距离病害点较近的树皮和树心也会褐化,并且挥发出一种芳香气味,这种病害会导致树木干枝枯死,严重的话整株树木会死亡。
而关于松褐天牛在松木枯死树干上的垂直分布规律,不同的研究者给出了不同的观点。
一些研究者认为松褐天牛在松木枯死树干上的垂直分布规律与其寄主-松材线虫病的分布规律有关。
研究表明,当松材线虫病初期发生时,松褐天牛会集中在感染区域附近,逐渐沿着树干向上移动。
随着病害的加重,松褐天牛的数量也会逐渐增多,直至树木死亡。
这种观点认为,松褐天牛的垂直分布规律受到松木病害的分布规律的影响,是一种被动的分布方式。
从目前的研究情况来看,对于松褐天牛在松木枯死树干上的垂直分布规律,还有待进一步的研究和探讨。
考虑到对松褐天牛垂直分布规律的研究可以为松木病害的监测和预防提供重要的理论依据,进一步深入研究这一课题具有重要的理论和实践意义。
松褐天牛在松木枯死树干上的垂直分布规律是一个复杂的问题,需要综合考虑寄主-松材线虫病的分布规律、松褐天牛的生物学特性等因素,结合实地观察和实验研究,加强对其动态分布规律的研究,以期为松木病害的监测和防控提供更为可靠的理论支持。
希望未来可以有更多的科研工作者投入到这一领域的研究中,为我国的森林健康管理提供更为科学的依据。
北京市东城区2023-2024学年高三二模考试生物试题(word版含答案)

2023北京东城高三二模生物2023.5本试卷共11页,共100分。
考试时长90分钟。
考生务必将答案答在答题卡上,在试卷上作答无效。
考试结束后,将本试卷和答题卡一并交回。
第一部分(选择题共30分)本部分共15题,每题2分,共30分。
在每题列出的四个选项中,选出最符合题目要求的一项。
的是1.组成植物细胞的各种化合物对植物体的生长发育有重要作用,下列相关叙述错误..A.自由水是植物细胞内良好的溶剂B.在叶绿素的合成中镁离子必不可少C.磷脂是构成植物细胞膜的主要成分D.糖原是植物细胞内重要的贮能物质2.H2O2溶液常用于伤口及环境消毒,会引起细菌等微生物细胞内蛋白质结构发生改变。
CAT是一种过氧化氢酶,可用于去除消毒后残余的H2O2。
关于CAT的叙述正确的是A.基本单位是氨基酸或脱氧核苷酸B.能提供分解H2O2所需的活化能C.去除残余H2O2的效果与反应温度、pH有关D.只能催化H2O2分解体现出CAT的专一性和高效性推断的是3.科研人员研究不同光照条件对柑橘生长的影响,部分检测结果见下表。
据此无法..光照强度叶色平均叶面积(cm2)净光合速率(μmolCO2m-2·s-1)强浅绿13.6 4.33中绿20.3 4.17弱深绿28.4 3.87A.三种光照条件下柑橘叶片均能将光能转化为NADPH和ATP中的化学能B.与强光条件相比,弱光下柑橘叶片叶绿素含量虽高但CO2吸收速率较低C.随光照强度减弱,平均叶面积增大可体现出柑橘对不同光照条件的适应D.光照强度增强主要提高了柑橘叶片的真光合速率并降低了呼吸速率4.紫杉醇为脂溶性抗肿瘤药物,可影响纺锤体的正常形成。
传统游离紫杉醇药物的注射液采用蓖麻油和乙醇作为溶剂,易引起严重过敏反应。
科研人员制备出一种紫杉醇脂质体药物,利用肝肿瘤模型鼠开展研究,结果如下表。
下列分析错误的是组别对照组脂质体药物组游离药物组药物浓度(mg•kg-1)020352035肿瘤重量(g) 1.3790.6960.3910.631死亡肿瘤抑制率(%)/49.471.650.1死亡A.紫杉醇能够通过抑制细胞的有丝分裂来实现抗肿瘤作用B.紫杉醇不能通过包在脂质体的两层磷脂分子之间进行运输C.浓度为35mg·kg-1的紫杉醇脂质体药物抗肿瘤效果明显D.与游离紫杉醇药物相比,紫杉醇脂质体药物对小鼠的副作用小5.牵牛花为虫媒、两性花,有白色、红色、蓝紫色等多种花色。
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律引言松褐天牛(Ips subelongatus Motschulsky)是一种重要的松树天敌,是松树林中较重要的害虫之一,在森林生态系统中具有重要的生物多样性和生态功能。
松褐天牛对松树的危害主要表现在在松树幼树、新树材和病树上产卵并导致树木死亡。
松材线虫病(Bursaphelenchus xylophilus)是松褐天牛传播的一种松树病害,在我国已成为林业的主要危害因素之一。
了解松褐天牛在松材线虫病病死树上的垂直分布规律对于控制其传播和减轻病害损失具有重要意义。
因此本文将对松褐天牛在松材线虫病病死树上的垂直分布规律进行研究。
材料与方法所研究的区域位于我国某地的松林区域,以不同程度的松材线虫病病死树为研究对象,使用拖曳式捕虫器(Lindgren Funnels)进行实验。
捕虫器分为三个高度(0.5米、1.5米、2.5米)进行设置。
每种高度分别设置10个捕虫器,每个捕虫器设置在不同的病死树上。
设置时间为3个月,每月进行一次数据的收集。
结果与讨论通过研究发现,在不同程度的松材线虫病病死树上,松褐天牛的垂直分布存在明显差异。
在0.5米高度处,松褐天牛数量较多,其次是1.5米高度,2.5米高度的松褐天牛数量较少。
这表明松褐天牛在松材线虫病病死树上的分布呈现出明显的垂直梯度,且其分布高度随着病死树的程度而有所不同。
在病死树的下部,由于枯死的木质部分较多,提供了更多的产卵和滋养的机会,因此0.5米高度处的数量最多。
2.5米处的数量最少,主要是由于距离地面较远,对于松褐天牛的产卵和滋养不利。
还发现不同程度的病死树上,松褐天牛的数量也有所不同,程度较轻的病死树上的松褐天牛数量较少,而程度较重的病死树上的松褐天牛数量较多。
这说明不同程度的病死树对于松褐天牛的吸引力也不同,程度较重的病死树对于松褐天牛的吸引力较大。
结论本研究结果表明,松褐天牛在松材线虫病病死树上的垂直分布呈现出明显的规律性。
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律
松褐天牛是一种木材害虫,常常在松林中活动。
它们喜欢在病死的松树上繁殖,因为
这些树木里面有松材线虫,是松褐天牛的主要食物来源。
然而,松褐天牛在病死树上的分
布规律并不是随机的,而是具有一定的空间结构性。
由于松褐天牛需要在树皮下寻找松材线虫,并用其产卵,因此它们通常会在垂直方向
上聚集在较浅的树皮处。
研究表明,松褐天牛成虫的分布范围通常是在树干基部到约2米
高度之间。
这个范围在不同松树的病死树上可能会有所变化,但总体趋势是相似的。
此外,在松树的树皮缝隙处,松褐天牛也有较高的分布密度。
这是因为在缝隙处,树
皮较薄,松褐天牛更容易进入树皮下寻找松材线虫。
因此,松树树皮缝隙处也是松褐天牛
的聚集区域之一。
在短期内,松褐天牛在病死树上的分布规律可能会受到环境和营养资源的影响而发生
一定的变化。
例如,如果树皮表面存在大量的腐菌,这些腐菌会分解松树树皮内部的养分,使得松褐天牛更倾向于在树干的上部寻找更多的营养来源。
但总体来说,在垂直方向上,
松褐天牛在树干基部到约2米高度之间的分布是相对稳定和规律的。
总的来说,松褐天牛在病死树上的垂直分布规律并不是随机的,而是受到树皮厚度、
树皮缝隙和松材线虫分布的影响。
对于研究和控制松褐天牛的繁殖,了解其分布规律是非
常重要的。
松墨天牛对寄主植物的适应性与松材线虫危害特性的研究的开题报告

松墨天牛对寄主植物的适应性与松材线虫危害特性的研究
的开题报告
一、研究背景
松墨天牛和松材线虫都是危害松树的主要害虫之一。
松墨天牛是一种木材性害虫,其幼虫和成虫均以松树为食,特别是以落叶松为主要食品来源,对森林和林木经济造
成了严重的危害。
而松材线虫则是一种致命的松树病原菌,它通过松墨天牛的传播侵
染松树,导致松树枯萎死亡,严重损害了林业资源和经济利益。
因此,对于松墨天牛
和松材线虫的研究具有重要的意义。
二、研究目的
本次研究旨在探究松墨天牛和松材线虫对寄主植物的适应性及其危害特性,为松树病虫害的防治提供科学依据。
三、研究内容和方法
1. 松墨天牛对寄主植物的适应性研究
通过野外调查和实验室试验,分析松墨天牛在不同寄主植物上的生存和繁殖能力。
采集不同地区的松墨天牛样本,进行形态学鉴定和分子生物学检测,并对其寄主选择
行为、产卵行为和幼虫发育状况进行观察和记录。
2. 松材线虫危害特性研究
以松墨天牛为传播媒介,通过采样、分离和鉴定,确定不同松树种及不同林分中松材线虫的分布和侵染情况。
利用组织学和分子生物学技术分析松材线虫在松树内的
生长和繁殖过程,研究其对松树的致病机制和危害特性。
同时,通过大量实验验证,
探究不同干扰因素对松墨天牛和松材线虫的影响及其生态适应策略。
四、研究意义
本研究的结果对于加强松树病虫害的防治、提高森林资源利用效益具有重要意义。
通过深入了解松墨天牛和松材线虫对寄主植物的适应性和危害特性,可以探讨有效的
防控措施,提高松树生长质量,保障林业经济的可持续发展。
松材线虫、松墨天牛和松树间的共生关系研究研究生课程论文大学论文

得分:_______ 南京林业大学研究生课程论文2015 ~20016 学年第二学期课程号:课程名称:论文题目:学科专业:森林保护学号:3150462姓名:王立超任课教师:黄麟二○一六年六月内容本文研究松材线虫、松墨天牛和真菌在松树中的共生关系实验材料和方法:(1)真菌小长喙霉在PDA平板上、25℃条件下培养一周,使用前在5℃条件下保存。
灰葡萄孢在10 mL的大麦籽粒上培养。
(2)线虫强毒虫株松材线虫T-4在25℃条件下在灰葡萄孢上培养,使用前在5℃条件下保存。
松材线虫悬浊液包括成虫和幼虫,悬浊液浓度为50000条/mL。
(3)昆虫松墨天牛在恒定条件下培养(25℃,16h光照、8h黑暗),让其繁殖10代,第七代到第十代成虫用来在赤松小苗上产卵,收集天牛所产的卵并将其放置在湿纸巾上,在一天之内孵化的幼虫用于实验。
(4)松枝2014年六月采伐两颗12年生(胸径分别为3.9和5.2cm)的赤松和两颗12年生的黑松(胸径分别为8.1和9.0cm),分别在实验1和实验3中使用。
2015年二月采伐3颗12年生的赤松用于实验2,胸径分别为7.4、7.6和8.4cm,2014年十月采伐一颗12年生的赤松用于实验4,其胸径为6.8cm。
树枝截为15cm一段,在树枝的两端用石蜡密封起来,每个实验组树枝的树木特定的每组树枝之间不存在显著差异(5)接种使用凿子和镊子在离树端13.7mm处从木质部分离树皮(面积22.3mm×17.3mm),用刻刀在漏出的木质部中间挖一个直径6-7mm圆形刻槽用以放置松墨天牛幼虫,一份带有灰葡萄孢或则没有灰葡萄孢的面积为5×5mm的PDA培养基放置在远离树端、接近圆形刻槽的位置。
用到划长16.8mm、最大宽度为14.3mm的三角形裂口,在漏出的韧皮部接种20uL的线虫悬浊液(1000条线虫)或者是无菌纯净水。
两个树皮伤口最小距离平均为13.7mm,两个伤口接种后覆盖原来的树皮,用封口膜包起来,即使不接种松墨天牛幼虫,也挖一个小刻槽,实验4中松树枝条不用接种松材线虫悬液或水,不会在树皮上形成倾斜伤口。
松材线虫的入侵危害与综合防治

松材线虫的入侵危害与综合防治杨钊(天津市河西区绿化管理所,天津300222)摘要:迄今为止,松材线虫已经成为园林绿化树种里危害最大的外来有害入侵生物之一,给我国的城市园林绿化建设以及森林生态防护体系建设造成了极为严重的危害。
松材线虫主要由松墨天牛等昆虫进行传播,其危害主要体现在对松树本身的生长造成树势衰弱、分泌物致使松树病、影响松林群落演替以及松林群落中的各个种群之间的关系等。
对于松材线虫,园林工作者应该本着“预防为主,防治结合,综合防治”的原则,应坚持IPM和SPMF思想,结合化学、物理、生物、天敌以及检验检疫技术等措施进行综合防治。
关键词:综合防治;外来有害生物;入侵;危害;松材线虫中图分类号:S763.18文献标识码:A文章编号:1005-7897(2020)14-0255-050前言松材线虫病是松树的一种危险性流行病,其病原为松材线虫(Bursaphelenchus xylophilus),隶属于垫刃线虫目、滑刃线虫科、伞滑刃线虫属。
该害虫主要以昆虫为传播媒介,其中,松墨天牛(Monochamus alternatus)又是最主要的媒介传播昆虫。
松材线虫可随被松墨天牛侵染过的原木整装调运而加快传播蔓延[1-2],因此松材线虫病又被称为“松树的癌症”[3]。
在松材线虫主要危害的松科植物中,松属树种有58种,非松属树种有13种。
1982年,我国于在南京中山陵首次发现该害虫,截至目前,已扩展至8个省(市、区),对我国松林资源、自然景观和生态环境造成了严重的破坏[4-5]。
国内外学者针对该害虫已展开了涉及症状分析、致病机理、风险评估以及检验检疫等旨在建立健全松材线虫综合防控体系的相关探索。
关于松材线虫的防治方法,目前大多集中在物理、化学、生物防治以及出入境检验检疫与各项规范性营林措施等方面。
这些从不同领域与角度开展实施的防治方法,目前已取得了一定的成效,可作为进一步深入研究的基础[6]。
然而,在松材线虫的整体防治体系中,由于缺乏可持续控制和综合调控防治思想,导致很多防治方法过分依赖于某一种单独的防治方式(或几种简单防治方式的叠加),比如单一的物理防治或化学防治,这很可能对整个松林生态系统造成严重损害,导致生态位不平衡化现象,进而影响松林生态系统生态功能的正常发挥。
松材线虫病及其媒介昆虫松墨天牛综合防治

松材线虫病及其媒介昆虫松墨天牛综合防治耿晓萍松材线虫病主要有松材线虫进入松树枝中,引发松树枯萎,松材线虫并具有极强的传播性会导致松林大面积枯死,产量减少,最终造成社会经济损失。
而松墨天牛是一种以松树计生的害虫,可以在松树树杆之内进行寄生,导致树木逐渐衰弱,最终死亡。
松墨天牛是传播松材线虫病的主要媒介,同时也能够在较短的时间内进行扩散,因此通过抑制媒介昆虫松墨天牛能够在一定程度上减少松材线虫病的发生。
一、松材线虫病发病机理松材线虫病及其媒介昆虫松墨天牛在进行综合防治的过程中,主要由于松墨天牛携带松材线虫进行传播,或是以人为的方式对感染病害的树木进行运输,从而实现不同区域之间的有效传播,松材线虫主要进入松木的内部,并且以寄生的方式大量繁殖,进一步阻止松石分泌,使得松木大量失水,最终导致植物死亡。
通过研究我们发现存在松材线虫寄生的松树会出现细胞结构的变化,同时松树体内的酶活力也会产生改变,植物激素含量也会随之变化。
23度左右是松材线虫并扩散的适宜温度区间。
二、松材线虫病防治技术1、预防措施通过调查我们可以发现松材线虫病发病主要集中在每年的七月份至九月份,因此我们可以在每年的三四月份以及十月份对于比较常见的疫区进行检查,对于并使树木的整体数量进行全面的统计,只有这样才能够针对松材线虫病的治理方式提出与之相对应的处理依据。
同时在对人为活动较多的区域进行定点巡查,对于没有发生松材线虫病的松木可以设置监测点,通过捕杀中间宿主,从而防止松材线虫病的再次发生,对于一些死亡的松树也可以进行采样鉴定,最终预防松材线虫病的再次发生。
2、治理措施在对松材线虫病进行检疫的过程中,我们可以采取以下方法,首先通过直观检验的方式,对于林间树木的生长发育情况以及媒介昆虫松墨天牛的危害情况进行检测,通过解剖的方式,对于疑似患病的树木进行观察,检测是否存在质量减轻的情况以及是否具备松墨天牛的蛀道,同时采用漏斗进行分离检验,对于坏死处进行加水浸泡,并对沉淀物进行鉴定是否存在松材线虫。
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律松褐天牛(Ips typographus)是一种重要的松木干蠹害虫,它主要危害松树。
与此松材线虫病(MPB)是一种由线虫引起的松树疾病,它同样对松树造成了严重的危害。
在松树死亡后,松褐天牛会趁机寄生于死去的树干中。
研究松褐天牛在松材线虫病病死树上的垂直分布规律,对于有效地控制松褐天牛的危害具有重要意义。
本文将探讨松褐天牛在松材线虫病病死树上的垂直分布规律,并对相关的研究成果进行分析和总结。
一、松褐天牛在松材线虫病病死树干中的生活史松褐天牛主要以松树为寄主,它的寿命大约为一年。
在松树受害后,松褐天牛会在松树的干部附近产卵,然后幼虫在树干内进行孵化和生长。
幼虫口器发达,可以溜进松树的材料中,并在其中进行取食和寄生。
在取食过程中,幼虫会在松树的内部形成许多长而半径适中的孔道。
这些孔道对于松树的营养和生长起到了非常不利的作用。
当松树患上了松材线虫病后,其树干会逐渐枯死。
松褐天牛却会在这些病死树干中寻找到了新的生存机会。
在松树枯死后,松褐天牛仍会在枯死的树干中进行产卵,并大量繁衍后代。
这样一来,松褐天牛对枯死树干的侵害会进一步加剧,从而加速树干的腐朽和崩溃。
研究发现,松褐天牛在松材线虫病病死树上的垂直分布是不均匀的,它们主要集中分布在树干的下部。
这一现象可以通过多种因素进行解释。
松褐天牛在寄生时主要依赖于树木的营养组织进行生长和发育。
而这些营养组织主要位于树干的下部部分,因此松褐天牛更倾向于选择树干下部作为产卵和取食的场所。
树干下部相对于树冠来说更加阴暗潮湿,这样的环境更有利于松褐天牛的生长和繁殖。
树干下部的环境对于天敌的侵袭也更加不利,这样一来,松褐天牛更容易避免天敌的威胁。
值得注意的是,尽管松褐天牛更倾向于分布在树干的下部,但它们对整个树干的侵害却是全面的。
一旦松褐天牛寄生于树干中,它们就会在树干的各个部分进行产卵和取食,从而导致树干内部的大量孔道形成。
这些孔道会影响树干内部的结构和稳固性,最终导致树干的崩溃和倒塌。
林业常见病虫害防治及综合防治

林业常见病虫害防治及综合防治林业作为生态系统的重要组成部分,对于维持生态平衡、提供生态服务、促进经济发展等方面都具有不可替代的作用。
然而,病虫害的侵袭一直是林业发展面临的严峻挑战。
了解和掌握常见病虫害的防治方法,实施综合防治策略,对于保护森林资源、维护生态安全至关重要。
一、常见的林业病虫害1、病害(1)松材线虫病:这是一种毁灭性的病害,主要危害松属树种。
松材线虫通过松墨天牛等媒介昆虫传播,染病松树会迅速死亡,且传播速度极快。
(2)杨树溃疡病:在杨树种植区较为常见,通常在树干上形成溃疡斑,严重时可导致树木生长不良甚至死亡。
2、虫害(1)美国白蛾:具有食性杂、繁殖量大、适应性强等特点,能危害多种阔叶树,严重时可将树叶全部吃光。
(2)松毛虫:是我国松林的主要害虫之一,幼虫取食松针,大发生时可将成片松林针叶吃光,造成松树生长不良甚至死亡。
二、病虫害的发生原因1、树种单一人工林往往树种单一,生态系统相对脆弱,缺乏生物多样性,这为病虫害的滋生和传播提供了有利条件。
2、气候异常气候变化,如温度升高、降水不均等,可能导致病虫害的发生周期和危害程度发生变化。
3、检疫漏洞检疫工作不到位,使得外来病虫害传入并扩散,给本地林业造成严重威胁。
4、人为活动不合理的采伐、抚育措施,以及林内垃圾的堆积等人为活动,破坏了森林的生态平衡,增加了病虫害发生的风险。
三、常见病虫害的防治方法1、物理防治(1)人工捕杀:对于个体较大、活动范围有限的害虫,如天牛、金龟子等,可以通过人工捕捉的方式进行防治。
(2)诱捕:利用害虫的趋光性、趋化性等特性,设置黑光灯、性诱剂等诱捕装置,将害虫集中诱杀。
(3)阻隔:在树干上涂抹粘虫胶、绑塑料布等,阻止害虫上树或下树,从而达到防治的目的。
2、化学防治(1)药剂喷雾:选择合适的农药,按照规定的浓度和方法进行喷雾,直接杀灭病虫害。
(2)树干注药:对于一些蛀干害虫,如天牛,可以在树干上打孔注药,使药剂通过树干内部的输导组织到达害虫危害部位,起到杀虫作用。
2024届山东省德州市高三一模生物试题
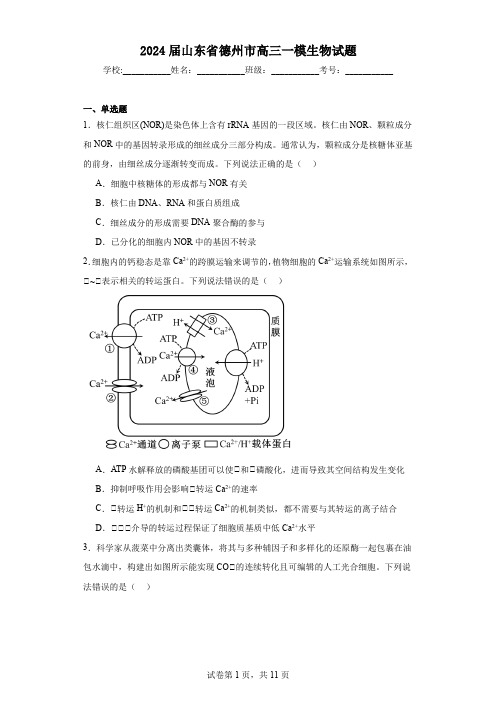
2024届山东省德州市高三一模生物试题学校:___________姓名:___________班级:___________考号:___________一、单选题1.核仁组织区(NOR)是染色体上含有rRNA基因的一段区域。
核仁由NOR、颗粒成分和NOR中的基因转录形成的细丝成分三部分构成。
通常认为,颗粒成分是核糖体亚基的前身,由细丝成分逐渐转变而成。
下列说法正确的是()A.细胞中核糖体的形成都与NOR有关B.核仁由DNA、RNA和蛋白质组成C.细丝成分的形成需要DNA聚合酶的参与D.已分化的细胞内NOR中的基因不转录2.细胞内的钙稳态是靠Ca2+的跨膜运输来调节的,植物细胞的Ca2+运输系统如图所示,①~①表示相关的转运蛋白。
下列说法错误的是()A.ATP水解释放的磷酸基团可以使①和①磷酸化,进而导致其空间结构发生变化B.抑制呼吸作用会影响①转运Ca2+的速率C.①转运H+的机制和①①转运Ca2+的机制类似,都不需要与其转运的离子结合D.①①①介导的转运过程保证了细胞质基质中低Ca2+水平3.科学家从菠菜中分离出类囊体,将其与多种辅因子和多样化的还原酶一起包裹在油包水滴中,构建出如图所示能实现CO①的连续转化且可编辑的人工光合细胞。
下列说法错误的是()A.人工光合细胞膜应该由单层磷脂分子组成B.需持续加入多种辅因子为CO①转化提供能量和还原剂C.通过改变还原酶的种类能实现可定制的CO①转化D.与菠菜叶肉细胞相比,人工光合细胞更有利于有机物积累4.姐妹染色单体通过粘连蛋白相互黏附而不能分离。
分离酶(SEP)可水解粘连蛋白,其活性受核基因编码的两种蛋白调控:SCR蛋白与SEP结合抑制其活性,而APC蛋白可催化SCR蛋白水解。
下列说法错误的是()A.SCR蛋白基因和APC蛋白基因可能在分裂间期表达B.SEP的功能异常可能导致子细胞染色体数目异常C.APC蛋白失活会导致姐妹染色单体不能正常分离D.SCR蛋白基因和APC蛋白基因的表达说明细胞发生了分化5.果蝇的有眼与无眼、正常翅与裂翅分别由基因D/d、F/f控制,已知这两对基因中只有一对位于X染色体上,且某一种基因型的个体存在胚胎致死现象。
浅谈松材线虫病防治技术

浅谈松材线虫病防治技术摘要概述松材线虫病发生的原理,松材线虫病发病规律、和松材线虫病防治。
关键词松材线虫病;发病规律;防治由于松材线虫病其危害的严重性和防治的难度而受到全世界各国的重视。
松材线虫病自1982年传入我国以来,造成了极大的生态破坏和经济损失。
现在国内已有15个省(区、市)的193个县(市、区)相距发生疫情。
这也给我国松材线虫病防治工作提出了新的挑战。
1病发的原理松材线虫病的发生与流行必须有松树、线虫、天牛、温湿度,四个因素存在。
只有四者共存才能形成病害,并流行为害。
研究表明,松材线虫主要在病树枝干内繁殖,针叶、球果和树皮中则很少发现线虫。
松树感线虫后,松脂减少并在短时间内停止流动,这就给天牛成活繁殖创造了良好的生存条件。
天牛在松树上产卵后,初孵幼虫先在韧皮部取食,并进入木质部钻蛀取食。
到春天,天牛化蛹时,木材中的线虫向蛹室聚集,待天牛羽化时携带大量的松材线虫从病死树内羽化逸出。
松褐天牛羽化后,必需在健康松树的嫩枝上补充营养,以一年生枝条为多。
这时由于嫩枝被天牛咬啃后有伤口,线虫就很快从天牛体内逸出,从伤口进入松树,然后线虫在松树体内大最繁殖。
据我们引诱到的松褐天牛分离有65%的天牛成虫携带线虫,携带500~999条线虫的占25%,1000~4999条的占20%。
而且松褐天牛有多次交尾、多次产卵的习性,这样天牛就反复接种线虫到松树上,当线虫在树体内温度适合时,仅需3~4天即可繁殖一代。
2病发的规律研究松材线虫主要依靠松褐天牛完成其侵染循环。
松褐天牛成虫从松材线虫寄主树中羽化脱离时携带了大量的松材线虫至新的寄主体内。
松材线虫(病原)、松褐天牛(传播媒介)和松树(寄主)三者之间这种生物学联系就构成了松材线虫病的侵染循环。
循环大致如下:5~6月时,寄生在死松树体内的松褐天牛开始化蛹,此时蛹室中大量的松材线虫。
在松褐天牛成虫从寄主树中飞出之前,大量的松材线虫通过附着在松褐天牛成虫身体各部位上。
松墨天牛携带松材线虫传播的化学生态学机制

松墨天牛携带松材线虫传播的化学生态学机制郑雅楠;杨忠岐;王小艺【摘要】松材线虫(Bursa phelenchus ylo ph ilus)在中国主要借助媒介昆虫松墨天牛(Monochamus alternatus)进行传播.通过识别寄主树木和媒介天牛释放的化学物质信号,松材线虫的4龄扩散型幼虫(JⅣ)进出松墨天牛体内,侵染健康寄主.本文综述了媒介昆虫天牛携带松材线虫进行传播的化学生态学研究进展.在松材线虫传播的过程中,脂肪酸类物质、挥发性萜烯类物质、碳氢化合物和CO2发挥重要的调控作用;除此之外,松材线虫体内的化学信号物质也决定其是否能够脱离媒介昆虫,从而成功地侵入新的健康寄主.【期刊名称】《植物保护》【年(卷),期】2014(040)001【总页数】5页(P12-15,29)【关键词】松材线虫;松墨天牛;媒介昆虫;化学信号【作者】郑雅楠;杨忠岐;王小艺【作者单位】中国林业科学研究院森林生态环境与保护研究所,国家林业局森林保护学重点实验室,北京 100091;沈阳农业大学林学院,沈阳110866;中国林业科学研究院森林生态环境与保护研究所,国家林业局森林保护学重点实验室,北京 100091;中国林业科学研究院森林生态环境与保护研究所,国家林业局森林保护学重点实验室,北京 100091【正文语种】中文【中图分类】S763.7松材线虫[Bursaphelenchus xylophilus (Steiner et Buhrer)]是引起松材线虫病(松树萎蔫病)的病原物。
该病被称为松树的“癌症”,对松林可造成毁灭性的危害。
然而,松材线虫却不能自行传播,必须借助媒介昆虫天牛进行传播扩散[1]。
松材线虫侵染感病的松树,在其上完成生活史,4龄幼虫能分化成繁殖型或分散型。
繁殖型幼虫用阿拉伯数字表示(如J2, J3, J4),分散型幼虫用罗马字符表示(如JⅢ,JⅣ)。
当食物和含水量充足时,松材线虫蜕皮为成虫,并通过分化成繁殖型来快速繁殖[2-3]。
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律松褐天牛(Ips subelongatus)是一种寄生于松树的天牛科昆虫,是松树上的一种重要害虫。
松材线虫病是由松材线虫引起的疾病,其以发病快、传播速度快、破坏力大而著称。
松褐天牛和松材线虫病的垂直分布规律一直是研究者们关注的焦点之一。
本文将分析松褐天牛在松材线虫病病死树上的垂直分布规律,以期为防治和管理提供科学依据。
一、松褐天牛和松材线虫病的基本特点松褐天牛是一种寄主植物为松树的害虫,对松树造成严重的破坏。
成虫体长3-4mm,呈褐色,寿命较长,约有一年左右。
幼虫为松树的内皮虫,主要在松树内皮部分活动,并损害松树的绿色部分。
松褐天牛不仅直接损害松树,同时也是松材线虫的传播媒介。
松材线虫病是一种由松材线虫引起的疾病,其以发病快、传播速度快、破坏力大而著称。
松材线虫主要寄生于松树的内皮部分,引起松树内皮和木质部的损害,严重影响松树的生长和发育。
松材线虫是由一种寄生虫引起的,因此在它的生长发育过程中需要有一定的宿主。
而松褐天牛则是其主要的传播媒介之一。
1. 林冠层在研究中发现,松褐天牛主要分布在病死松树的林冠层部分。
病死树的林冠层是其最主要的生活环境,也是其产卵和繁殖的地方。
林冠层对于松褐天牛来说是一个相对理想的环境,其温度适宜,空气流通性好,适合松褐天牛的生长和繁殖。
2. 松树干部松褐天牛在松材线虫病病死树上主要分布在松树的干部。
松褐天牛主要是通过寿食、产卵等方式在松树的干部上生活和繁殖的。
松树的干部是其最主要的生活环境,也是其产卵和寿食的地方。
松褐天牛在松树的干部上的分布是其主要的分布规律之一。
3. 树皮下部1. 对松树的损害2. 对森林生态系统的影响松褐天牛在松材线虫病病死树上的垂直分布规律也对森林生态系统的影响是非常大的。
松褐天牛是松树上的一种重要害虫,对松树的生长和发育造成了很大的破坏。
这种破坏不仅仅是对松树个体的影响,同时也对整个森林生态系统产生了影响。
松褐天牛在松材线虫病病死树上的垂直分布规律

松褐天牛在松材线虫病病死树上的垂直分布规律松褐天牛是松木林中的一种害虫,它对松树的危害主要表现在向幼树和病弱树群中产卵孳生的突发性爆发,也可以对成年树造成程度不同的危害。
而松材线虫病是松木上的一种疾病,它由松材线虫引起,极大地影响了森林的健康。
松褐天牛在松材线虫病病死树上的垂直分布规律对于森林病虫害的监测与防治有着重要的意义。
松褐天牛在松材线虫病病死树上的垂直分布规律是指在病死的树上,松褐天牛的分布位置与高度、树轮的厚度、树干的直径等因素之间的关系。
由于松褐天牛的生活习性和生长发育需要,它会选择不同的位置进行活动,而这些位置的选择往往与它们的生活史和生长环境息息相关。
树干的高度与松褐天牛的垂直分布规律密不可分。
根据研究资料显示,松褐天牛在松树的树干上的垂直分布区域主要集中在树干的中下部分,即从树干底端到离地面1.5米高度之间。
而在这个高度范围内,松褐天牛的数量会随着树干的高度逐渐增加,表现出明显的正相关关系。
这是因为松褐天牛产卵需要选择适宜的地方,而这个高度范围内的条件最为适宜,所以松褐天牛的数量也会相对较多。
树轮的厚度对于松褐天牛的垂直分布规律也有一定的影响。
研究表明,树轮的厚度越大,松褐天牛的数量也会越多。
这是因为树轮的厚度代表了树木生长的年限和生长环境,而这个因素会直接影响到松褐天牛的生活史和生长发育。
在树轮的厚度较大的树木上,松褐天牛的生存条件会更为舒适,因此它们也会选择这样的树木进行生长繁殖。
松褐天牛在松材线虫病病死树上的垂直分布规律受到树干高度、树轮厚度和树干直径等因素的影响。
为了有效地控制松褐天牛的数量,森林管理者需要充分了解这些因素对于害虫分布的影响,制定相应的防治策略。
只有这样,才能更好地保护森林资源,维护生态平衡。
松墨天牛防治技术

松墨天牛防治技术松墨天牛是我国松树的重要蛀干害虫,也是松材线虫病(Bursaphelench us xylophilus)的主要媒介昆虫。
拉丁学名:Monochamus alternatus Hope异名:Monochammus tesserula Wirte英文名:Pine longicom beetle;Pine sawyer分类地位:鞘翅目(Coleoptera),天牛科(Cerambycidae),沟胫天牛亚科 (Lamiinae)。
中文别名:松褐天牛、松天牛分布:中国北京、河北、上海、江苏、浙江、福建、广东、广西、湖南、四川、西藏、陕西、河南、台湾;国外分布于越南、老挝、朝鲜、日本。
寄主:主要有马尾松、冷杉、云杉、红松、黑松、雪松、落叶松、油松、华山松、柏、杉。
危害:成虫补充营养,啃食嫩枝皮,造成寄主衰弱,幼虫大量钻蛀树势衰弱或新伐倒的树干,引起成片松树枯死。
此外,成虫还是松材线虫病的主要传播媒介,一旦传入,将会造成松林大面积死亡。
形态特征成虫:体长10~30mm,宽4.5~9.5mm,橙黄色至赤褐色。
每个鞘翅上有5条纵纹,由方形或长方形黑色及灰白色绒毛斑点相间组成,斑纹特殊,故与其他种易于区别。
头部额区刻点细密,头顶处较粗糙并略有皱纹。
口器为下口式,侧观其额面与体轴近似垂直。
触角栗色或称棕褐色,雄虫触角比雌虫的长。
第三节比柄节长约一倍,并略长于第四节,柄节上具封闭式端疤。
雄虫第一、二节全部及第三节基半部,具稀疏的灰白色绒毛,触角全长超过体长的1.5倍。
雌虫除末端一、二节外,其余各节除端处留有深色环以外,均具稀疏的灰白色绒毛,触角长度近似于体长或仅超过体长的1/3。
前胸宽胜于长,刻点粗糙多皱纹,中瘤不显著,两侧具一对较大的刺突,其端处稍呈圆锥形。
前胸背板上有二条明显的橙黄色纵带与三条黑褐色纵带间隔排列。
前足基节窝为封闭式,中足胫节处侧缘端处具一斜沟。
小盾片上密被橙黄色绒毛。
翅基部有颗粒状突起和粗大的刻点,呈半规则排列,翅面上具方形或长方形黑色、褐锈色及灰白色绒毛,组成大小不等的斑块,镶嵌成不规则的五条纵纹。
松材线虫与松墨天牛

松材线虫与松墨天牛作者:毕役来源:《课外语文》2012年第12期松材线虫是一种非常细小的蠕虫,体长仅有14—16微米。
可是,这个小虫却是世界各国严密防范的入侵生物,因为它就是松材线虫病的病原。
松材线虫病又名松枯萎病,是影响最严重的森林病害,危害的对象主要是松属树种,如马尾松、黑松、白皮松和红松等,也包括部分非松属的树木,如一些落叶松属和云杉属的树种。
患病的树木水分运输、光合作用、呼吸和蒸腾逐渐停顿,树皮变得干枯,针叶开始一束束地褪去绿色,变为黄褐色乃至红褐色,逐渐萎蔫,最后整株都会枯死。
在我国,松材线虫病于1982年首见于江苏南京,现在已有疫区近100个,直接、间接损失达上千亿元。
松材线虫原产于北美洲,它的远距离扩散主要是依靠潜藏在患病的树苗、木材以及用患病树木所制成的木制品中,靠人为带入一个新环境的。
松材线虫无足无翅、步履维艰,它在新环境中是如何实现大规模的入侵的呢?原来,它每到一地,就与其他一些物种建立“利益共同体”,以“多兵种联合作战”的方式进行协同入侵,而与松材线虫“狼狈为奸”的最主要接应者就是松墨天牛。
松墨天牛也叫松褐天牛,在我国分布较广,寄主植物主要有马尾松、黑松等松属植物以及冷杉、云杉、栎、苹果等树种。
它的幼虫是一种重要的蛀干害虫,能够钻蛀松树枝干的韧皮部及木质部,影响水分、养分在树体内的传输,致使树木生长衰弱,有的甚至枯死。
不过,这些还不是它的全部罪恶。
松墨天牛的成虫还能作为传播媒介,携带松材线虫,导致松材线虫病的扩散蔓延。
当松墨天牛化蛹过冬时,大量的松材线虫幼虫便已聚集在其蛹室周围,成为最有资格搭乘松墨天牛“航班”的“旅客”。
一旦松墨天牛羽化外出后,每只松墨天牛的身上都搭载着成千上万只松材线虫,最多可达将近30万只!借助松墨天牛一对能够飞翔的翅膀,松材线虫可以向森林大举入侵。
当然,松墨天牛也不是甘愿“助人为乐”的奉献者,它与松材线虫是为了共同的利益而走到一起的。
被松材线虫侵入后衰弱、濒死或枯死的松树,就不会再分泌大量的香油树脂,而松墨天牛的幼虫最怕被香油树脂包裹而死。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
得分:_______ 南京林业大学研究生课程论文2015 ~20016 学年第二学期课程号:课程名称:论文题目:学科专业:森林保护学号:********名:***任课教师:**二○一六年六月内容本文研究松材线虫、松墨天牛和真菌在松树中的共生关系实验材料和方法:(1)真菌小长喙霉在PDA平板上、25℃条件下培养一周,使用前在5℃条件下保存。
灰葡萄孢在10 mL的大麦籽粒上培养。
(2)线虫强毒虫株松材线虫T-4在25℃条件下在灰葡萄孢上培养,使用前在5℃条件下保存。
松材线虫悬浊液包括成虫和幼虫,悬浊液浓度为50000条/mL。
(3)昆虫松墨天牛在恒定条件下培养(25℃,16h光照、8h黑暗),让其繁殖10代,第七代到第十代成虫用来在赤松小苗上产卵,收集天牛所产的卵并将其放置在湿纸巾上,在一天之内孵化的幼虫用于实验。
(4)松枝2014年六月采伐两颗12年生(胸径分别为3.9和5.2cm)的赤松和两颗12年生的黑松(胸径分别为8.1和9.0cm),分别在实验1和实验3中使用。
2015年二月采伐3颗12年生的赤松用于实验2,胸径分别为7.4、7.6和8.4cm,2014年十月采伐一颗12年生的赤松用于实验4,其胸径为6.8cm。
树枝截为15cm一段,在树枝的两端用石蜡密封起来,每个实验组树枝的树木特定的每组树枝之间不存在显著差异(5)接种使用凿子和镊子在离树端13.7mm处从木质部分离树皮(面积22.3mm×17.3mm),用刻刀在漏出的木质部中间挖一个直径6-7mm圆形刻槽用以放置松墨天牛幼虫,一份带有灰葡萄孢或则没有灰葡萄孢的面积为5×5mm的PDA培养基放置在远离树端、接近圆形刻槽的位置。
用到划长16.8mm、最大宽度为14.3mm的三角形裂口,在漏出的韧皮部接种20uL的线虫悬浊液(1000条线虫)或者是无菌纯净水。
两个树皮伤口最小距离平均为13.7mm,两个伤口接种后覆盖原来的树皮,用封口膜包起来,即使不接种松墨天牛幼虫,也挖一个小刻槽,实验4中松树枝条不用接种松材线虫悬液或水,不会在树皮上形成倾斜伤口。
实验一、为了确定松墨天牛幼虫和小长喙霉对松材线虫种群生长的影响,一组赤松枝条接种松材线虫、松墨天牛和小喙长霉。
第二组枝条接种松材线虫、松墨天牛和PDA,第三组枝条接种松材线虫和小喙长霉,第四组枝条接种松材线虫和PDA。
他们被垂直放置使线虫远离容器底部。
25℃,16h光照、8h黑暗,相对湿度100%条件下常温培养,用纯净水保持湿度。
接种18天后,从松枝上剥离树皮,从枝条上取幼虫和幼虫粪便,记录韧皮部和木质部上蓝变面积确定小喙长霉的繁殖情况,从纵向韧皮部上剥离四份之一到二分之一的树皮,将树皮外层剥离,收集里面树皮以获得韧皮部样品,再将里层树皮的韧皮部剪切为1-2mm大小的树皮块,使用一把15mm宽的刀片,从接种松墨天牛幼苗和松材线虫的地方包括髓取木质部样品,记录木质部样品真菌繁殖情况,粪便、韧皮部和木质部分别在25℃下使用贝尔曼漏斗法分离48-72h,计算分离线虫浓度,材料在60℃下烘5-12d,称重,线虫密度表示每克材料干重中松材线虫的数量。
分别使用电子天平和游标卡尺测量幼虫体重和头壳宽度,分别精确到0.1mg和0.05mm,根据头壳宽度确定幼虫龄期:0.585–1.170,1.260–1.935, 1.980–3.150和3.150–4.365 mm分别为一龄、二龄、三龄和四龄虫。
实验二、确定赤松中韧皮部、木质部和天牛幼虫蛀屑线虫密度的不同,在2015年2月26日将刚孵化的幼虫、1000条松材线虫和一份接有小长喙霉的PDA接种在10条直径相似的上述赤松枝条上,2015年3月26日分别将韧皮部、木质部和幼虫蛀屑用以分离线虫,确定线虫密度,实验三、确定松材线虫种群和小喙长霉对松墨天牛生长发育的影响,四组黑松枝条接种松材线虫、松墨天牛和小喙长霉三个物种组成的四种组合。
方法和实验一方法相同,培养条件相同。
每天观察松枝,记录松墨天牛羽化情况,分性别记录成虫数量,体重和鞘翅长分别精确到0.1mg和0.05mm。
没有松墨天牛出现的松枝在接种153-164d后分离,称量幼虫的鲜重和记录头宽、颜色、肠内排泄物的有无以及幼虫所在的位置。
实验四、为确定松墨天牛幼虫对小喙长霉繁殖的影响,6个赤松枝条接种小喙长霉和松墨天牛幼虫,接种22d后,剥离松枝树皮,幼虫蛀屑和幼虫取样,测量幼虫体重和头壳宽度文章结构设计思路实验一为研究赤松枝条中松墨天牛幼虫和小喙长霉菌对松材线虫种群的影响,实验选取年龄和粗度相似的赤松为材料,分四组实验进行,一组赤松实验材料只接种松材线虫、一组赤松材料接种松材线虫和松墨天牛,一组赤松材料接种松材线虫和小喙长霉菌,最后一组赤松实验材料接种松材线虫、松墨天牛和小喙长霉菌,四组实验之间彼此形成对照,通过测量不同接种组合、不同部位的松材线虫密度和松墨天牛的体重和头宽指标来确定三者之间的相互影响。
实验二为研究韧皮部、木质部和天牛幼虫粪便中线虫密度的不同,刚孵化的松墨天牛、松材线虫和小喙长霉菌接种在10个粗细相似的赤松枝条上,按照实验一发法在各部位取样,测定松材线虫密度。
实验三为研究松材线虫种群和小喙长霉菌对松墨天牛生长发育的影响,在四组黑松实验材料上按照实验一中接种方法接种松材线虫、松墨天牛和小喙长霉菌三个物种的组合。
每天观察记录松墨天牛成虫出现情况,测定体重和鞘翅长度指标。
在接种后153-164天后,解剖没有松墨天牛出现的松树枝条,称量记录活天牛幼虫的头宽、身体颜色、排泄物状态和其在松树枝条内所在的位置。
实验四研究松墨天牛对小喙长霉菌影响,6个赤松枝条接种松墨天牛和小灰长霉菌,24天后,在接种位置上剥离树皮,从松树枝条上取下幼虫和幼虫粪便,取整个松树枝条四分之一到二分之一的树皮(包括天牛取食过的树皮,不包括石蜡覆盖的树皮)去除外层树皮,韧皮部切成1-2cm宽的小块,使用手钻在接种松墨天牛和小喙长霉菌的位置取韧皮部包括髓,松墨天牛粪便、韧皮部和木质部放在聚乙烯袋子里,-20℃下保存,两天后,在真空状态下冻干。
测定麦角固醇含量。
结果和结论实验一在接种18天后,解剖松树枝条,在树皮下发现20只活的天牛幼虫和4只死的天牛幼虫,活天牛幼虫包括1个一龄幼虫、2个二龄幼虫和17个三龄幼虫,平均体重99.2mg,平均头壳宽2.23mm,在幼虫肠道内发现粪便。
解剖实验表明在单独接种松材线虫的11个松树枝条中有6个松树枝条可观察到小喙长霉菌,在接种松材线虫和人松墨天牛的12个松树枝条中有9个松树枝条可观察到小灰长霉菌,在单独接种小灰长霉菌的所有松树枝条上的木质部和韧皮部均有真菌分布。
在未接种松墨天牛幼虫的松树枝条,木质部和韧皮部的分别分离到线虫量为2.2g和2.9g,在接种松墨天牛幼虫的松树枝条上,木质部、韧皮部和粪便中分别分离到3.6g、3.2g和0.29g松材线虫。
平均线虫密度随接种位置、松墨天牛幼虫及真菌的有无而不同,在未接种真菌的松树枝条木质部的松材线虫密度小于其他接种位置的松材线虫密度,小于接种真菌的松树枝条。
未接种松墨天牛幼虫的松树枝条线虫密度大于接种松墨天牛幼虫的松树枝条线虫密度。
双因素方差分析表明,在接种松墨天牛幼虫的受接种位置和真菌有无影响,但不受接种位置和真菌互作影响,松材线虫密度在粪便中最大,其次为木质部,最小为韧皮部。
实验二在接种刚孵化的松墨天牛幼虫的赤松枝条中存在天牛粪便,解剖发现9个活的天牛和1个死亡的天牛幼虫,活天牛幼虫平均体重为118.4mg,头壳宽2.19mm,表明活天牛为三龄幼虫,死亡天牛头壳宽为1.35mm,说明此为二龄幼虫,解剖实验发现在10个松树枝条的韧皮部存和5个松树枝条的部分木质部存在真菌。
韧皮部、木质部和粪便线虫密度为5.3g、3.3g和0.52g。
双因素方差分析表明,线虫密度受接种位置影响,但不受松树枝条影响,松材线虫密度最大为天牛粪便、其次为韧皮部和木质部。
实验三在接种幼虫后的71天到167天期间出现23只雌天牛和30只雄天牛,在接种后153-164天后解剖松树枝条发现在木质部蛹室内发现11只活天牛幼虫和9只死亡天牛幼虫,活天牛幼虫平均体重为982.3mg,头壳宽为3.89mm,体色为黄白色到黄色,表明他们是四龄幼虫。
方差分析表明成虫的体重不受真菌接种实验、线虫接种、性别、树皮面积和彼此之间互作关系影响。
方差分析也表明天牛鞘翅长度受真菌和松材线虫互作关系、真菌、松材线虫和树皮面积三者互作关系的影响,但是不受真菌、线虫和树皮面积的影响。
雌天牛的鞘翅比雄天牛鞘翅长。
鞘翅长度与树皮面积呈负相关关系。
方差分析显示接种天牛幼虫后,天牛成虫出现时间不受真菌、线虫、树皮面积和天牛性别以及彼此之间互作关系的影响。
实验四在接种22天后,在松树枝条发现6只活天牛幼虫,平均体重171.2mg,平均头壳宽为2.4mm,表明他们为三龄幼虫。
天牛粪便的麦角固醇含量最大,其次为韧皮部和木质部,方差分析显示三者之间存在极显著差异。
创新性指导意义实验过程控制单因素变量,实验测定的指标多样,可以从不同角度反映实验结果,实验分析多样化,实验数据多次筛选,层层分析,增加数据可靠性。
不足之处实验中只考虑线虫取食小喙长霉菌,没有考虑线虫取食松树枝条活细胞的实验四研究松墨天牛对小喙长霉菌影响,实验中在6个赤松枝条上均接种松墨天牛幼虫和小喙长霉菌,但未在赤松枝条上单独接种小喙长霉菌作为对照。
ReferencesAkbulut S, Linit MJ (1999) Flight performance of Monochamus carolinensis (Coleoptera: Cerambycidae) with respect to nematode phoresis and beetle characteristics. EnvironEntomol 28:1014–1020.Arakawa Y, Togashi K (2002) Newly discovered transmission pathway of Bursaphelenchus xylophilus from males of the beetle Monochamus alternatus to Pinus densiflora trees via oviposition wounds. J Nematol 34:396–404Carlile MJ, Watkinson SC, Gooday GW (2001) The fungi, 2nd edn. Academic Press, New York Dowding P (1970) Colonization of freshly bared pine sapwood surfaces by staining fungi. Trans Br Mycol Soc 55:399–412Edwards OR, Linit MJ (1992) Transmission of Bursaphelenchus xylophilus through oviposition wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). J Nematol 24:133–139 Haack RA, Slansky F (1987) Nutritionalecology of wood-feeding Coleoptera, Lepidoptera, and Hymenoptera. In: Slansky F, Rodriguez JG (eds) Nutritional ecology of insects, mites,spiders, and related invertebrates. Wiley, New York, pp 449–486Ichihara Y, Fukuda K, Suzuki K (2000) Early symptom development and histological changes associated with migration of Bursaphelenchus xylophilus in seedling tissues of Pinusthunbergii. Plant Dis 84:675–680Kanzaki N, Futai K (2001) Life history of Bursaphelenchus conicaudatus (Nematoda: Apehelenchoididae) in relation to the yellowspotted longicorn beetle, Psachothea hilaris(Coleoptera: Cerambycidae). Nematology 3:473–479Kanzaki N, Futai K (2002) Life history of Rhabdontolaimus psacotheae (Diplogasterida: Diplogasteroididae) and its habitat segregation from Bursaphelenchus conicaudatus(Aphelenchida: Aphelenchoididae). Jpn J Nematol 32:60–68Kaya HK (1984) Nematode parasites of bark beetles. In: Nickle WR (ed) Plant and insect nematodes. Marcel Dekker, New York, pp 727–754Kobayashi T, Sasaki K, Mamiya Y (1974) Fungi associated with Bursaphelenchus lignicolus, the pine wood nematode (I). J Jpn Soc 56:136–145 (in Japanese with English summary) Kojima K, Katagiri K (1964) On the larval instar and changes of its composition of Monochamus alternatus Hope. J Jpn Soc 46:307–310 (in Japanese with English summary)Kuroda K, Ito S (1992) Migration speed of pine wood nematodes and activities of other microbes during the development of pine-wilt disease in Pinus thunbergii. J Jpn Soc 74:383–389 (in Japanese with English summary)Maehara N, Futai K (2000) Population changes of the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), on fungi growing in pine-branch segments. ApplEntomol Zool 35:413–417Mamiya Y (1983) Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu Rev Phytopathol 21:201–220Mamiya Y, Enda N (1972) Transmission of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae) by Monochamus alternatus (Coleoptera: Cerambycidae). Nematologica 18:159–162Massey CL (1974) Biology and taxonomy of nematode parasites and associates of bark beetles in the United States, Agricultural Handbook 446. USDA Forestry Service, Washington Matsunaga K, Togashi K (2009) Seasonal change in susceptibility of Pinus densiflora to Bursaphelenchus xylophilus infection, determined from the number of nematodes passing through branch sections. Nematology 11:409–418Ochi K (1975) Ecological studies on the Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae). II. Size of the first instar larva. In: Transactions of the 86thannual meeting of the Japanese Forestry Society. pp 323–324 (in Japanese)Oku H, Shiraishi T, Chikamatsu K (1989) Active defense as a mechanism of resistance in pine against pine wilt disease. Ann Phytopathol Soc Jpn 55:603–608Rutherford TA, Webster JM (1987) Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Can J Res 17:1050–1059Rühm W (1956) Die Nematoden der Ipiden. Parasitologische Schriftenreihe 6:1–435Seitz LM, Mohr HE, Burroughs R, Sauer DB (1977) Ergosterol as an indicator of fungal invasion in grains. Cereals Chem 54:1207–1217Son JA, Komatsu M, Matsushita N, Hogetsu T (2010) Migration of pine wood nematodes in the tissues of Pinus thunbergii. J Res 15:186–193Sudhaus W, von Lieven AF (2003) A phylogenetic classification and catalogue of the Diplogastridae (Secernentea, Nematoda). J Nematode Morphol System 6:43–89Togashi K (1991) Different developments of overwintered larvae of Monochamus alternatus (Coleoptera: Cerambycidae) under a constant temperature. Jpn J Entomol 59:149–154 Togashi K (1997) Lifetime fecundity and body size of Monochamus alternatus (Coleoptera: Cerambycidae) at a constant temperature. Jpn J Entomol 65:458–470Togashi K (2014) Effects of larval food shortage on diapause induction and adult traits in Taiwanese Monochamus alternatus alternatus. Entomol Exp Appl 151:34–42Togashi K, Matsunaga K (2003) Between-isolate difference in dispersal ability of Bursaphelenchus xylophilus and the vulnerability to inhibition by Pinus densiflora.Nematology 5:559–564Togashi K, Sekizuka H (1982) Influence of the pine wood nematode, Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae), on longevity of its vector, Monochamus alternatus(Coleoptera: Cerambycidae). Appl Entomol Zool 17:160–165Wingfield MJ, Blanchette RA (1983) The pine-wood nematode, Bursaphelenchus xylophilus in Minnesota and Wisconsin: insect associates and transmission studies. Can J Res 13:1068–1076Wingfield MJ, Blanchette RA, Nicholls TH (1984) Is the pine wood nematode an important pathogen in the United States? J Forestry 82:232–235Zhao LL, Lu M, Niu HT, Fang GF, Zhang SA, Sun JH (2013) A native fungal symbiont facilitates the prevalence and development of an invasive pathogen-native vector symbiosis. Ecology 94:2817–2826Zhao LL, Mota M, Vieira P, Butcher RA, Sun JH (2014) Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol 30:299–308。