真核生物DNA复制过程讲义
分子生物学 第3章 DNA复制
DNA helicase (DNA解旋酶)
利用ATP供能,解开DNA双链, 可随复制叉 的伸展向前移动
大肠杆菌中解旋酶的种类
种 类
DnaA DnaB DnaC
功 能
辨认起始点,并结合到复制起始部位 解开DNA双链 运送和协同DnaB
single-stranded binding protein (SSB, 单链结合蛋白)
是一类调节DNA分子的超螺旋水平,可改变DNA拓扑性 质的酶。对DNA分子的作用是既能水解、又能连接磷酸 二酯键。 • 拓扑异构酶 I: 切开DNA双链中的一股,使DNA在解链旋 转中不打结,DNA变为松弛状态再封闭切口。 同转录有 关 • 拓扑异构酶 II: 能切断DNA双链,使螺旋松弛。在ATP参 与下,松弛的DNA进入负超螺旋,再连接断端。同复制
3´→5´外切酶活性: 切除错配的核苷酸
5'
3' C T T C A G G A G A A G T C C G G C G 5'
3'
DNA ligase
连接DNA链3-OH末端和相邻DNA链5-P末端,形成磷 反应需要ATP。
酸二酯键,从而把两段相邻的DNA链连接成完整的链。
二、 DNA复制的过程
E. Coli DNA在15N-标记的营养液中生
长多代,使DNA双链充分标记
将15N-标记
细胞在
14N中
细胞在
14N中复
细胞在
14N中复
的E.Coli 加入14N 培 养液中
万有引力
复制1 次
制第2次
制第3次
单林娜 制作
11
DNA半保留复制的生物学意义:
DNA的半保留复制表明DNA在代谢上的稳定性,
DNA的复制PPT课件
•Molecular Biology Course
(CsCl gradient centrifuge)
N15
DNA
N14
Semi-ConservationReplication
–第三阶段为DNA复制的终止阶段。DNA复制的整个 过程中需要30多种酶及蛋白质分子参加,我们将 在DNA复制的各个阶段着重介绍它们的作用。
•Molecular Biology Course
•Molecular Biology
Course (二)、复制的起始阶段
1、复制的起点 2、复制的方向 3、复制的速度 4、DNA复制起始引发体的形成及所参与的酶和 蛋白质
1、DNA半保留复制的机理 2、DNA的半不连续复制
•Molecular Biology
Course
1、DNA半保留复制的机理
Semi-Conservation Replication
DNA作为遗传物质的基本特点就是在细胞分裂前进行准 确的自我复制,使DNA的量成倍增加,这是细胞分裂的 物质基础。
当用缺乏糖苷酶的大肠杆菌变异株(ung-进行 实验时,尿嘧啶不再被切除。)
此时,新合成的DNA有一半放射性标记出现于岗 崎片断中,另一股直接进入大的片断。由此可 见,当DNA复制时,一条链是连续的,另一条链 是不连续的,因此称为半不连续复制(semidiscontinuous replication) 。
二、复制的起始阶段
•Molecular Biology Course
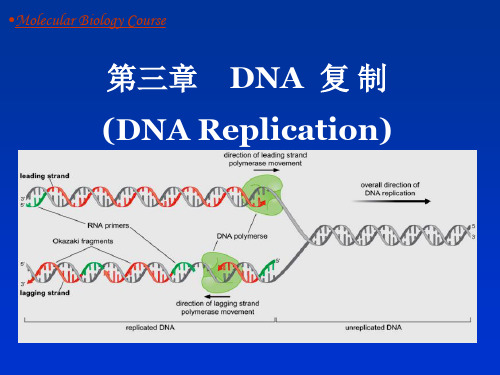
复制叉( replication fork ):DNA分子中正在进行 复制的分叉部位。它由两条亲代链及在其上新合成的子 链构成。
dna的复制ppt课件

针对DNA复制机制的研究有助 于深入了解衰老的机制,并开发 新的抗衰老治疗方法,延长人类
寿命和提高健康水平。
06
DNA复制的应用
基因克隆和基因组学
基因克隆
通过复制特定的DNA片断,科学家 可以克隆出特定的基因,用于研究基 因的功能、表达和调控机制。
基因组学
通过大规模复制DNA,科学家可以测 定全部基因组的序列,从而研究基因 组的组成、结构和功能,推动人类对 生命本质的理解。
科学研究与应用
DNA复制是生物学、遗传学和分子生 物学等领域的重要研究对象,对于理 解生命本质、疾病治疗和生物技术应 用等方面具有重要意义。
02
DNA复制的机制
DNA解旋酶
01
02
03
功能
DNA解旋酶能够解开 DNA双螺旋结构,暴露出 单链的DNA模板,为复制 进程做准备。
类型
DNA解旋酶分为原核生物 和真核生物两种类型,它 们在结构、功能和作用方 式上有所不同。
胞膜上的受体结合,发挥调节作用。
DNA复制
02
在细胞生长和分裂进程中,DNA复制是一个重要的环节,它决
定了细胞的遗传信息和表型特征。
生长因子对DNA复制的调控
03
生长因子通过调节细胞内信号转导途径,影响DNA复制的起始
和进程,从而调控细胞的生长和分裂。
基因表达对DNA复制的调控
01
基因表达
基因表达是指基因经过转录和翻译, 合成具有功能的蛋白质的进程,是基 因发挥生物学效应的基础。
变积累。
DNA复制进程中出现特殊可能导 致基因扩增、基因缺失或染色体 特殊,从而促进肿瘤的产生和发
展。
针对DNA复制机制的靶向治疗是 肿瘤治疗的新方向之一,旨在抑 制肿瘤细胞的DNA复制或修复进 程,从而抑制肿瘤的生长和扩散
DNA复制过程ppt课件

4)PolⅢ为不对称二聚体, 一个作用于前导链, 另一个作用于随从链(loop)。
5′ 3′
领头链 (leading strand)
顺着解链方向生成的子链,其复制是 连续进行的,得到一条连续的子链。
3'
5' 解 链 方 向 3 '
5'
3'
5'
随从链 (lagging strand)
参与原核生物复制起始的主要成分
DnaA蛋白
辨认起始点
DnaB蛋白(解螺旋酶) 解开DNA双链
DnaC蛋白
协助DnaB蛋白
DnaG蛋白(引物酶) 催化形成RNA引物
SSB
稳定解开的单链DNA
拓朴异构酶
理顺DNA链
oriC
大肠杆菌的复制起始点
2.复制的延长
1)在DNA聚合酶Ⅲ的作用下 2)前导链合成1000-2000个核苷酸后 3)随从链开始合成,岗崎片段的长度
PRA:复制蛋白A
2.RNA引物的合成
DNA聚合酶α能识别起始位点,并且以核 糖核苷三磷酸为底物(NTP),以解开的一 段DNA为模板,合成一个短链RNA(8~10 个核苷酸)引物。然后从引发酶的活性转变为 DNA 聚 合 酶 α 的 活 性 , RNA 引 物 的 3’-OH 末端为合成新的DNA单链的起点,以dNTP 为原料,延长引物大约15-30个脱氧核苷酸。
DNA复制过程
(一)原核生物DNA复制过程 1.复制的起始
DNA复制的起始就是要解开双链和生成引物。 (1)DNA解成单链
由拓扑异构酶松弛超螺旋,解螺旋酶 解开双链,SSB结合到单链上使其稳定。
复制起始的解链需要多种蛋白质参与。这 些蛋白质与复制起始点的特有序列结合, 促使其邻近的D来自A解链。3.DNA链的延伸
真核生物DNA复制过程ppt课件

19
前复制复合物在G1期形成而在S期被激活
复制基因(replicator)是指DNA复制起始 所必需的全部DNA序列
真核细胞DNA复制的起始分两步进行,即复 制基因的选择和复制起点的激活
20
前复制复合物(pre-RC)的形成
复制起点识别复合物(origin recognition complex, ORC)识别并结合复制基因
7
DNA聚合酶/转换
➢ 前导链:出现在引发后期 ➢ 后随链:发生于每个冈崎片段合成之际 ➢ 发生DNA聚合酶转换的原因是Pol 不具备
持续合成能力 ➢ DNA聚合酶转换的关键蛋白是RFC
8
真核DNA聚 合酶转换和 后随链合成
9
三、真核生物DNA合成后立即组装成核小体
亲代DNA
5
3
5 3
引物 核小体
PCNA
激活DNA聚合酶和RFC的ATPase活性
RFC
有依赖DNA的ATPase活性,结合于引物-模板链,激活DNA聚合酶,
促使PCNA结合于引物-模板链
Pol /引发酶 合成RNA-DNA引物
Pol /
DNA复制,核苷酸切除修复,碱基切除修复
FEN1
核酸酶,切除RNA引物
RNAse H
核酸酶,切除RNA引物
ORC至少募集两种解旋酶加载 蛋白Cdc6和Cdt1
三种蛋白质一起募集真核细胞 解旋酶Mcm2-7
21
pre-RC的激活 和组装真核 DNA复制叉
22
CDK控制pre-RC的形成和激活
真核细胞通过依赖细胞周期蛋白的蛋白激 酶(cyclin-dependent kinases,CDK)严格 控制pre-RC的形成和激活
中国海洋大学生物化学课件第二十三章 DNA的复制-讲义

第二十三章 DNA的复制目的和要求:了解DNA的半保留、半不连续复制、复制的起点和方式;重点掌握DNA复制相关酶和蛋白质;掌握大肠杆菌DNA复制过程,特别是起始过程和调节;真核生物DN复制的特点及与原核生物的异同;了解真核生物端粒复制DNA生物合成概述:核酸的生物合成是模板指导下进行的,模板有DNA和RNA.因此,DNA合成分为DNA为模板的DNA复制和RNA为模板的逆转录;DNA合成主要原料为四种脱氧核苷三磷酸,酶系和辅助因子的参与;本章主要讲解DNA复制,以原来DNA分子为模板合成出具有相同分子的过程,自我复制。
一. DNA的半保留复制1958年,Meselson&Stahl氯化铯梯度离心实验,多种真核和原核生物做了类似实验,证明DNA的复制为半保留复制,但由于实验研究中的DNA均为提取的片断,反映复制前和复制后的状态1963年,Cairns用放射自显影的方法观察到完整的正在复制的大肠杆菌染色体DNADNA的半保留复制:以双链DNA分子的每一条链为模板,按照碱基配对的原则,合成出两个与原DNA 分子碱基顺序完全一样的新DNA分子,其中每个子代分子的一条链来自亲代DNA,另一条链则是新合成的,这种复制方式为半保留复制二. DNA复制的起点和方式复制子:基因组能独立进行复制的单位,包含控制复制的起点和复制的终点.单一复制子和多复制子复制的起点:特殊的碱基序列能与控制复制开始的蛋白质或酶特异识别,结合,复制开始复制叉:复制开始,两条链解开,形成的叉形结构.复制终点:原核生物环状DNA分子具有特殊的碱基序列,终止复制;真核生物不同复制叉相遇,复制终止。
复制方向和复制方式:复制方向有单向和多向;复制方式多种多样。
三. DNA的半不连续复制随着复制叉的延伸,DNA的两条母链分别合成与其互补的子链,目前分离到的催化DNA形成的聚合酶的合成方向都是5’→3’,DNA在复制时如何同时作为模板合成其互补链?半不连续复制当DNA复制时,一条链是连续的,另一条链是不连续的,因此称半不连续复制;前导链;滞后链四.DNA复制有关的酶和蛋白质㈠拓扑异构酶功能:DNA超螺旋松弛,消除解链产生的扭曲张力TopI:单链蛋白质,广泛存在于原核和真核生物转录区,在转录过程中发挥作用切开超螺旋DNA的两条链中的一条,切口末端反超螺旋转动,闭合。
真核生物DNA复制和起始调控资料

二、前DNA复制复合体的组装过程
三、DNA复制起始点识别复合 体的构成和组装
ORC:由六个蛋白(或亚基)构成, 分别为Orc1、Orc2、Orc3、Orc4、 Orc5和Orc6。Orc蛋白在结构和功能上 均相当保守。
经分析表明,芽殖酵母的ORC仅分布 在有限的DNA复制起始位点上。与启 动子相关的转录调节因子,可能影响 ORC与DNA复制起始位点结合,进而 影响DNA复制的起始。ATP的结合与 水解也显著影响ORC的功能。
结构致密的异染色质区的DNA较晚 复制。染色质重构因子也被认为影 响DNA复制。局部核小体结构也会 影响DNA复制起始调控。
第四节 DNA复制起点的激活 与G1/5期检控点
一、DNA复制起始的激活
DNA复制起始的激活也称为 “复制起始点触发”,需要 更多的因子组装到复制起始 点上。这个过程可以分为两 个步骤:
六、Mcm2-7复合体与前DNA 复制复合体组装
MCM蛋白在DNA复制起始过程中起主 要作用。
第三节 前DNA复制复合体 组装过程的调节
前DNA复制复合体组装过程的调节机制主要 是由周期性依赖性的CDK激酶活性进行直接 或间接驱动的。
Cdk(cyclin dependent kinase)激酶活性在 真核生物DNA复制起始调控中具有双重作用。 一方面,当细胞进入S期,Cdk激酶活性增高, 激活DNA复制的起始;另一方面,Cdk激酶 活性阻止同一周期中S、G2和M期中DNA的 多重复制。此外,Cdk激酶还控制特定起始 点复制的起始时间。
一个周期内发生多重复制。在酵母细胞中,调 节Cdc6的表达水平是保证“在一个细胞周期中 DNA只能复制一次”的重要机制之一。
五、Cdt1蛋白与前DNA复 制复合体组装
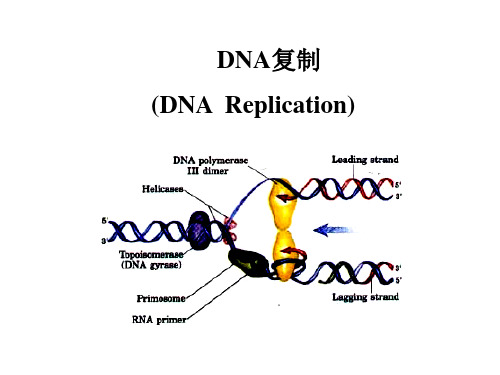
DNA复制 (DNA Replication)

processivity).
聚合酶的第3个组成部分是由5个亚基构成的一个所
谓的γ复合体。γ复合体又称夹子装载因子(clamp
loader),催化滑动夹打开,并将其结合在引物-
模板上。介导β亚基与模板-引物双螺旋的结合。 最后,两个拷贝的τ亚基使核心聚合酶形成二聚体。
3.在复制叉处先导链和后随链的合成同时进行 RNA引物合成以后,DNA的延伸过程便开始了。先 导链被连续合成,而后随链是不连续合成的。在复
包括oriC区域中的GATC,都被甲基化。
新复制的GATC位点呈半甲基化状态,即旧链是甲基
化的,但新链尚未被甲基化。
新复制、半甲基化的oriC可被SeqA蛋白识别,
SeqA与半甲基化的GATC紧密结合。SeqA的结合出现
了两个结果:首先它极大地降低了与之结合的GATC 序列的甲基化速率;其次阻止了DnaA蛋白与复制起 点的结合。当SeqA偶尔从GATC位点上脱离时,序列 即被DNA甲基转移酶完全甲基化,防止了SeqA的重新
第二节:DNA的复制起点和复制方式
一、复制起点与复制子
Replicon: 作为一个单位进行复制的任何一段DNA
序列。 它含有一个复制起点,有时还含有一个复制
终点。
Origin :是复制子起始复制的一段DNA序列。
作为一个单位进行复制的任何一段DNA序列。 复
制子的复制通常从一个固定的位点开始的,这种起
复制叉不起作用。例如,TerB阻断顺时针方向复制叉,TerA
阻止逆时针方向复制叉。终止区这样的安排可确保两个从相 反方向进入ter区的复制叉总能相遇,当一个复制叉遇到另 一个复制叉时,DNA复制就完成了。
四、复制起始调控
大肠杆菌染色体DNA的 oriC含有11个拷贝的GATC序
真核生物DNA的复制
counterpart, the dimeric sliding clamp.
真核生物DNA的复制
1. 真核与原核生物在复制过程上的相同之处: -半不连续复制; -具有相似的起始、延伸等过程。
真核生物DNA的复制
❖ 定义: 在一系列酶的作用下,将DNA分子中受损伤
部分切除掉,并以完整链的一条链为模板,合成 切去的部分,使DNA恢复正常结构的过程。
❖ 包括:
碱基切除修复(base-excision repair) 核苷酸切除修复(nucleotide-excision repair)
真核生物DNA的 复制
两个pol分别催化前导 链和滞后链的合成
真核生物DNA的复制
1. 端粒(telomere)
真核生物线性染色体末端的特殊结构,由许 多成串短的重复顺序组成,具有稳定染色体末 端结构的功能。
四膜虫 TTGGGG(仅列一条链的序列)
人
TTAGGG
2. 端粒酶(telomerase)
含有RNA链的逆转录酶,它以所含RNA为模 板来合成 DNA端粒结构。
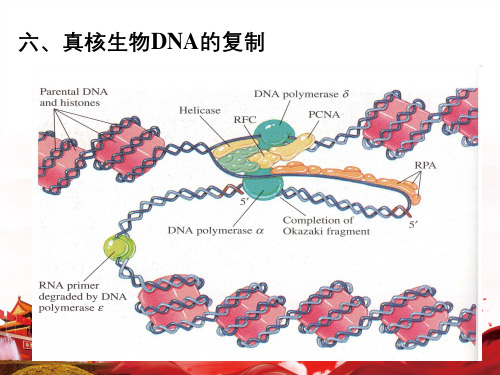
六、真核生物DNA的复制
(一)参与复制的酶及蛋白质因子
1. DNA聚合酶:pol 、、、 、
位置 亚基数目 3’→5’ 外切酶 引物合成酶 持续合成能力 准确性
α (I) 细胞核
4 + 中等 高
(IV) 细胞核
1 低 低
功能
引物合成 修复
γ (M) 线粒体
2 + 高 高 线粒体DNA 合成
δ (III) 细胞核
高中生物必修二DNA的复制课件

04
原料
脱氧核苷酸是合成新链的基本原料, 包括脱氧核糖、磷酸和含氮碱基。
DNA复制的特点
半保留复制
半不连续复制
双向复制
精确性高
DNA复制后,每条新链 都保留了原来模板链的
一半序列。
DNA复制时,前导链连 续合成,后随链不连续
合成。
DNA复制时,两条链同 时进行复制,形成两个
子代DNA分子。
DNA聚合酶具有校对功 能,能够保证复制的精
02
DNA复制是生物体生长、发育和 繁殖的基础,是遗传信息传递的 关键环节。
DNA复制的过程
DNA解旋
DNA双链在解旋酶的作用下解 开,形成单链模板。
校对与修复
合成过程中存在校对和修复机 制,确保新合成的子链准确性 。
DNA复制起始
DNA复制起始于特定的起始点 ,由特定的起始蛋白激活。
合成子链
DNA聚合酶按照单链模板的顺 序,合成新的子链。
高中生物必修二DNA的复制课件
• DNA复制的概述 • DNA复制的条件和特点 • DNA复制的过程和机制
• DNA复制的调控 • DNA复制的错误与修复 • DNA复制的应用与展望
01 DNA复制的概述
DNA复制的定义
01
DNA复制是指DNA双链在细胞分 裂前,通过复制的方式,将遗传 信息传递给下一代的过程。
考古研究等,促进相关领域的发展。
THANKS 感谢观看
DNA复制的研究展望
探索DNA复制的分子机制
01
深入研究DNA复制的分子机制,了解DNA复制过程中各种蛋白
质的作用和相互调控关系。
开发新型DNA复制技术
02
探索新的DNA复制技术,提高DNA复制的速度、准确性和效率
DNA复制过程解析
多起点、单方向(真核) 多起点、双方向(真核)
3. 复制方式
(1)从新起始( de novo initiation )或复制叉式( replication fork )
即先导链是从新开始合成的,DNA在复制原点解开成单 链状态,分别作为模板,各自合成其互补链,则出现两个叉 子状结构,称为从新起始。
● 真核生物(Eukaryote) : 染色体上有多个复制起点,即 一个genome中有多个复制单位,真核生物基因组DNA为多复 制子DNA。
Replication fork
● 复制起点富含AT, “呼吸作用”十分 明显,是许多酶的 结合位点。长度一 般为几十--几百bp。
AT rich
2. 复制方向 ● 复制叉(Replication fork):
The multi protein (30±) structure that assembles at replicating fork to undertake synthesis of DNA
DNA 复制发生时期:细胞周期的S期
一. DNA的半保留复制 (Semi-Conservation Replication)
● 转入正常的以14N为碳源的培养基中;
● 提取各代DNA,进行CsCl密度梯度离心,分析各代 DNA的浮力密度。
CsCL密度梯度离心 浮力密度
N15-DNA
1.742g/ml
N15-N14-DNA 1.717g/ml
N14-DNA
1.710g/ml
15N标记实验
二、DNA复制的原点、方向和方式
线粒体和叶绿体DNA的复制方式为D-环复制。
(3)共价延伸方式(covalence elongation)或滚环复制 (rolling circle replication) 某些环状DNA分子复制时,在复制原点处以某种方式
分子生物学 第三章 DNA的复制PPT课件

复制过程中首先两条链间的氢键破裂并使双链
解旋和分开,然后以每条链为模板,按碱基互
补配对原则(A:T,G:C),由DNA聚合酶催化合
成新的互补链,结果由一条链成为互补的两条
链,这样新形成的两个DNA分子与原来的DNA
分子的碱基序列完全相同。在此过程中,每个
子代DNA的一条链来自亲代DNA,另一条链则
是新合成的。这种复制方式称此过程中,每个
37
(三)DNA复制的终止
❖ 过去认为,DNA一旦复制开始,就会将该 DNA分子全部复制完毕,才终止其DNA复制。 但最近的实验表明,在DNA上也存在着复制 终止位点,DNA复制将在复制终止位点处终 止,并不一定等全部DNA合成完毕。
Meselson等证明DNA的半保留复制
6
复制起点和复制子
❖ DNA复制在生物细胞中要从DNA分子上特定
位置开始。这个特定的位置就称为复制起点
(Origin of replication),用ori表示。DNA复制 从起点开始双向进行直到终点为止,每一个 这样的DNA单位称为复制子或复制单元 (replicon)。
12
Okazaki fragment 1968 Reiji Okazaki
3‘ (semi-discontinuous replication ! )
5‘
3‘
5‘
DNA replication in Okazaki fragment 1kb At least one strand of DNA replication in Okazaki fragment 1kb13
26
❖ 为什么需要有RNA引物来引发DNA复制呢? 这可能尽量减少DNA复制起始处的突变有关。 DNA复制开始处的几个核苷酸最容易出现差 错,因此,用RNA引物即使出现差错最后也 要被DNA聚合酶Ⅰ切除,提高了DNA复制的 准确性。RNA引物形成后,由DNA聚合酶Ⅲ 催化将第一个脱氧核苷酸按碱基互补原则加 在RNA引物3&#点
DNA分子的复制--公开课PPT课件

9
练习:结合所学知识回答下列问题:
(1)从图中可看出DNA复制的方式是 __半__保__留__复__制___。 (2)A和B是DNA分子复制过程需要的酶, 其中B能将单个的脱氧核苷酸连接成 脱氧核苷酸链,从而形成子链;
则A是____解__旋_____酶, B是_____D_N__A_聚__合_____酶。 (3)图示过程在绿色植物叶肉细胞 中进行的场所细有胞__核__、__线__粒__体,、叶绿体 进行的时间为_有__丝__分__裂__的__间__期____。
位素磷分别是: 31P , 31P 和32P 。
⑷、上述实验结果证明了DNA的复制方式
是 半保留复制 。
23
2020/1/15
26
10
二、对DNA分子复制的推测
半保留复制
全保留复制
11
如果要你来设计实验,你认为最基本的思路是什么?
区分亲代和子代的DNA,
标记亲代DNA链,然后观察它们在子代DNA中如何分 布
12
2020/1/15
13
问题1:如果要在实验中直观地区别、“标识” 母链或子链,可以采取什么办法?
同位素标记法
问题2:可以用什么元素的同位素(放射性)
6
一、DNA分子复制的过程(P54) :
解旋: 解旋酶催化 模板 边解旋边复制
合成子链:以母链为模板进行碱基配对 (在DNA聚合酶的催化下,利用游离的脱 氧核苷酸合成脱氧核苷酸链)
母链(旧链) 形成子代DNA: 组成
子链(新链)
方式:半保留复制
7
8
一、DNA复制的过程
思考及小组讨论解决以下问题:
5
一、DNA复制的过程
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA聚合酶/转换
前导链:出现在引发后期 后随链:发生于每个冈崎片段合成之际
端粒酶以自己的RNA组分作为模板,以染色体的3’端 ssDNA(后随链模板)为引物,将端粒序列添加于染色体 的3 端。这些新合成的DNA为单链
端粒酶催化作用的爬行和端粒的延长过程
五、真核生物染色体DNA在每个细胞周 期中只能复制一次
真核所有染色体DNA复制仅仅出现在细胞 周期的S期,而且只能复制一次
组成: 端粒酶RNA (human telomerase RNA, hTR) 端粒酶协同蛋白(human telomerase associated protein 1, hTP1) 端粒酶逆转录酶(human telomerase reverse transcriptase, hTRT)
端粒酶(telomerase)是一种核糖核蛋白(RNP),由 RNA和蛋白质组成
ORC至少募集两种解旋酶加载 蛋白Cdc6和Cdt1
三种蛋白质一起募集真核细胞 解旋酶Mcm2-7
pre-RC的激活 和组装真核 DNA复制叉
CDK控制pre-RC的形成和激活
真核细胞通过依赖细胞周期蛋白的蛋白激 酶(cyclin-dependent kinases,CDK)严格 控制pre-RC的形成和激活
蛋白激酶通过磷酸化激活或抑制各种复制因子 而实施调控作用
一、真核生物复制的起始与原核基本相似
真核生物每个染色体有多个起始点,是多复制 子复制。复制有时序性,即复制子以分组方式 激活而不是同步启动
复制的起始需要DNA-pol α(引物酶活性)、 pol δ和pol ε参与。还需解螺旋酶活性、拓扑 酶和复制因子(replication factor, RF)
Pol /
DNA复制,核苷酸切除修复,碱基切除修复
FEN1
核酸酶,切除RNA引物
RNAse H
核酸酶,切除RNA引物
DNA连接酶Ⅰ 连接冈崎片段
DNA解旋酶 DNA双螺旋解链,参与组装引发体
TolⅠ和TolⅡ 拓扑异构酶,去除负超螺旋(使解旋酶容易解旋),去除复制叉前方产生的正 超螺旋
二、真核生物复制的延长发生DNA聚合酶转换
发生DNA聚合酶转换的原因是Pol 不具备 持续合成能力
DNA聚合酶转换的关键蛋白是RFC
真核DNA聚 合酶转换和 后随链合成
三、真核生物DNA合成后立即组装成核小体
亲代DNA
5
3
5 3
引物 核小体
3 5
前导链
后随链
3 5
切除引物的两种机制
线性DNA复制一次端粒缩短
5
3
3
5
5
3
3
5
5
3
3
5
+
5
3
3
端粒(telomeres)由富含TG的重复序列组成 人的端粒重复序列为5’-(TnGn)x-3 这些重复序列多为双链,但每个染色体的3’
端比5端长,形成单链ssDNA。这一特殊结构 可解决染色体末端复制问题
端粒酶(telomerase)
真核DNA复制叉主要蛋白质的功能
蛋白质
功能
RPA
单链DNA结合蛋白,激活DNA聚合酶,使解旋酶容易结合DNA
PCNA
激活DNA聚合酶和RFC的ATPase活性
RFC
有依赖DNA的ATPase活性,结合于引物-模板链,激活DNA聚合酶,
促使PCNA结合于引物-模板链
Pol /引发酶 合成RNA-DNA引物
Cdk功能: 激活pre-RC,以起始DNA复制 抑制形成新的pre-RC
六、真核生物线粒体DNA按D环方式复 制
D-环复制(D-loop replication)是线粒 体DNA的复制形式
DNA-pol γ dNTP
前复制复合物在G1期形成而在S期被激活
复制基因(replicator)是指DNA复制起始 所必需的全部DNA序列
真核细胞DNA复制的起始分两步进行,即复 制基因的选择和复制起点的激活
前复制复合物(pre-RC)的形成
复制起点识别复合物(origin recognition complex, ORC)识别并结合复制基因
真核生物 DNA复制过
程
真核生物与原核生物DNA复制的差异: 真核生物复制子多、冈崎片段短、复制叉前进速度慢等 DNA复制从引发进入延伸阶段发生DNA聚合酶转换 切除冈崎片段RNA引物的是核酸酶RNAse H和FEN1等
DNA合成期
G2
S
M 哺乳动物的 细胞周期
G1
细胞能否分裂,决定于进入S期及M期这两个关 键点。G1→S及G2→M的调节,与蛋白激酶活 性有关