西维因抗体库的构建及特异性克隆的筛选
抗人肺癌单链抗体噬菌体库的构建及特异性单链抗体的鉴定

抗人肺癌单链抗体噬菌体库的构建及特异性单链抗体的鉴定罗弋;庞华;李少林;曹辉;李淑杰;王树斌;樊春波【期刊名称】《基础医学与临床》【年(卷),期】2009(029)011【摘要】目的构建人源噬菌体抗体库,并从中筛选出抗肺癌人源单链抗体.方法提取肺癌患者癌旁淋巴结组织,通过RT-PCR扩增出重链可变区基因(VH)和轻链可变区基因(VL),再经剪切-重叠-延伸PCR(SOE-PCR)将VH和VL连接得到单链抗体(ScFv).将双酶切后的ScFv基因片段克隆入噬菌体表达载体pCANTAB5E,得到初级噬菌体抗体库.以肺腺癌细胞株A549为抗原对抗体库进行4轮筛选富集,鉴定抗体库性能.将得到的阳性克隆用IPTG诱导表达并进行检测.结果成功构建噬菌体单链抗体库.经筛选富集,噬菌体收获率得到增加,第4轮是第1轮的115倍.随机选取10个克隆,通过ELISA法检测到其中7个与A549细胞呈阳性反应,阳性率为70%.SDSPAGE及ELISA检测证实得到人源抗肺癌单链抗体.结论成功构建人源单链抗体噬菌体库,从中获得具有较高特异性的抗人肺癌单链抗体.【总页数】6页(P1155-1160)【作者】罗弋;庞华;李少林;曹辉;李淑杰;王树斌;樊春波【作者单位】重庆医科大学放射医学教研室,重庆,400016;重庆医科大学附属第一医院核医学科,重庆,400016;重庆医科大学放射医学教研室,重庆,400016;重庆医科大学放射医学教研室,重庆,400016;重庆医科大学放射医学教研室,重庆,400016;包头市中心医院肿瘤科,内蒙古包头,014040;重庆医科大学放射医学教研室,重庆,400016【正文语种】中文【中图分类】Q78【相关文献】1.抗脱氧雪腐镰刀菌烯醇噬菌体单链抗体库的构建与鉴定 [J], 周艳红;祭芳;丁衬衬;史建荣2.抗速灭威噬菌体单链抗体库的构建、筛选及鉴定 [J], 李铁军;李德全3.抗人β-淀粉样肽特异性单链抗体的噬菌体抗体库的构建与初步鉴定 [J], 段朝晖;毛越苹;罗晓红;李民友;劳伟思;王英;朱振宇4.鼠源抗AFB1噬菌体单链抗体库的构建与鉴定 [J], 裴世春;孙大庆;郭德军;宋薇;蒋琛5.全人源抗结肠癌噬菌体单链抗体库的构建及筛选鉴定 [J], 谢平丽;李官成;李艳东;周国华;李跃辉;郭锋杰;王甲甲因版权原因,仅展示原文概要,查看原文内容请购买。
西维因人工抗原的合成与鉴定

西维因人工抗原的合成与鉴定帅丽芳;赵勇;银涛;黄桂通;蒋昀;李代波【摘要】[目的]制备西维因农药小分子的抗体,为建立免疫分析方法提供保障.[方法]制备6-(1-萘氧基甲酰胺基)己酸及其活化酯,将其与生物大分子偶联得到人工免疫抗原和包被抗原.[结果]通过对CN H及其活化酯的偶联物进行质谱、核磁共振、红外线鉴定,证实合成的产物为目标人工标原.[结论]成功制备了西维因人工抗原,为西维因农药残留免疫分析的方法研究奠定了基础.%[Objective]To establish immunoassaymethod ,the antibody of carbary pesticide wasprepared .[Methods]6-(1-naphthyloxy methanamide)hexanoic acid was synthesized and activated ,and then coupled with the biological macro-molecules to obtain the artificial antigen .[Results]The synthesized product was confirmed as the target artificial antigen by massspectrometry ,nuclear magnetic resonance and infrared spectroscopy detection .[Conclusion]The artificial anti-gen was synthesized successfully ,and it laid the foundation for the preparation of carbaryl antibody and establishing immunemethod .【期刊名称】《山西农业大学学报(自然科学版)》【年(卷),期】2017(037)010【总页数】6页(P749-753,760)【关键词】西维因;人工抗原;合成【作者】帅丽芳;赵勇;银涛;黄桂通;蒋昀;李代波【作者单位】广州军区疾病预防控制中心 ,广东广州 510507;中山大学生命科学学院 ,广东广州 510006;广州军区疾病预防控制中心 ,广东广州 510507;广州军区疾病预防控制中心 ,广东广州 510507;广州军区疾病预防控制中心 ,广东广州510507;广州军区疾病预防控制中心 ,广东广州 510507【正文语种】中文【中图分类】S482.3+4农药是用来防治农作物虫害、病害及草害的化学物质,在农业生产上广泛使用,按化学组成可分为有机磷、氨基甲酸酯类等[1]。
【国家自然科学基金】_单链抗体库_基金支持热词逐年推荐_【万方软件创新助手】_20140731
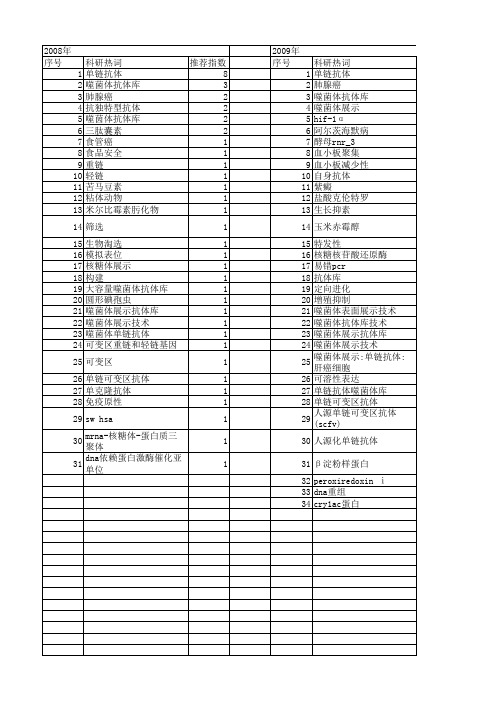
科研热词 推荐指数 单链抗体 8 噬菌体抗体库 3 肺腺癌 2 抗独特型抗体 2 噬茵体抗体库 2 三肽囊素 2 食管癌 1 食品安全 1 重链 1 轻链 1 苦马豆素 1 粘体动物 1 米尔比霉素肟化物 1 筛选 1 生物淘选 1 模拟表位 1 核糖体展示 1 构建 1 大容量噬菌体抗体库 1 圆形碘孢虫 1 噬菌体展示抗体库 1 噬菌体展示技术 1 噬菌体单链抗体 1 可变区重链和轻链基因 1 可变区 1 单链可变区抗体 1 单克隆抗体 1 免疫原性 1 sw hsa 1 mrna-核糖体-蛋白质三聚体 1 dna依赖蛋白激酶催化亚单位 1
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
科研热词 单链抗体 噬菌体展示 重组抗体 西维因 表达 胃癌 筛选 磁珠 生存素 时间分辨免疫分析 微囊藻毒素lr 大容量抗体库 噬菌体抗体库 噬菌体抗体 噬菌体展示技术 可溶性抗体 单链抗体库 兔源抗体库 人源性 亲水突变 亲和力成熟 乳腺癌 vegfr2
推荐指数 4 3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
科研热词 细菌噬菌体 噬菌体抗体库 单链抗体 随机突变pcr 重组系统 逆转录聚合酶链反应 蛋白相互作用 肽类 破伤风毒素 破伤风 狂犬病病毒g蛋白 流式细胞仪筛选 抗原抗体共表达 抗体库筛选 抗体库 抗体 宿主菌 定点突变pcr 噬菌体次级抗体库 噬菌体展示 周质空间 单链抗体(scfv) 单克隆 伏马菌素b1 中和活性
骆驼天然单域重链抗体库的构建与鉴定

骆驼天然单域重链抗体库的构建与鉴定周景明;翟圆君;耿玥;祁艳华;刘红亮;王爱萍【摘要】构建骆驼非免疫噬菌体抗体库,完成抗体库的鉴定.从未经免疫的健康骆驼的外周血淋巴细胞中提取总RNA后,特异性扩增骆驼重链抗体可变区(VHH)片段,将其与噬菌粒载体 pCANTAB5E连接获得重组子,多次电转感受态大肠埃希菌TG1后获得 VHH 抗体基因库,并分析其库容量及多样性.结果表明,经过吸附-洗脱的淘选过程筛选出能够与莱克多巴胺人工抗原特异性结合的噬菌体展示纳米抗体.构建的骆驼天然重链单域抗体库的库容量为107,多样性良好,独立克隆所占比例为90%,筛选出针对莱克多巴胺人工抗原的噬菌体颗粒.已成功构建骆驼天然噬菌体重链抗体库,并筛选出抗莱克多巴胺的噬菌体颗粒,为抗莱克多巴胺纳米抗体的表达和莱克多巴胺的免疫学检测奠定基础,并为后续淘选针对特定抗原的单域重链抗体奠定了基础.%A native camel variable heavy chain phage display antibody library was constructed and was au-thenticated.Total RNA was purified from peripheral blood lymphocytes of the healthy non-immune camel and the VHH coding sequence was amplified,then the PCR products were cloned into a phagemid vector pCANTAB5E.By electroporation of E.coli TG1,the primary library was obtained.The library capacity and diversity were analyzed.The phage display nano-antibodies specific binding ractopamine artificial antigens were selected by adsorption-elution panning.The results showed that a native camel single-domain antibody phage display library containing more than 107individual colonies and functional clones above 90% and having good diversity was obtained.And anti-ractopamine phage particles were selected.A native camel sin-gle-domain antibody phagedisplay library was constructed,and anti-ractopamine phage particles were se-lected.This would lay the foundation for expression of anti-ractopamine nanobody and immunological de-tection of ractopamine,which would be useful for generating VHHs with specific binding affinity.【期刊名称】《动物医学进展》【年(卷),期】2018(039)001【总页数】7页(P61-67)【关键词】纳米抗体;噬菌体抗体库;莱克多巴胺;骆驼;噬菌体展示【作者】周景明;翟圆君;耿玥;祁艳华;刘红亮;王爱萍【作者单位】郑州大学生命科学学院,河南郑州450001;郑州大学生命科学学院,河南郑州450001;郑州大学生命科学学院,河南郑州450001;郑州大学生命科学学院,河南郑州450001;郑州大学生命科学学院,河南郑州450001;郑州大学生命科学学院,河南郑州450001【正文语种】中文【中图分类】S852.4;Q7891993年,Hamers-Casterman C等发现骆驼免疫系统内存在一种仅由重链单一的折叠单元组成的抗体,将其称为单域重链抗体(variable domain of heavy-chain of heavy-chain antibody,VHH)或纳米抗体(nanobody,Nbs)[1]。
构建诱导型CRISPRCas9系统用于小鼠免疫细胞基因功能研究

Vol.41No.3Mar.2021上海交通大学学报(医学版)JOURNAL OF SHANGHAI JIAO TONG UNIVERSITY (MEDICAL SCIENCE)构建诱导型CRISPR/Cas9系统用于小鼠免疫细胞基因功能研究赵艳娜1*,邱荣2*,沈南1,唐元家11.上海交通大学医学院附属仁济医院风湿病科,上海市风湿病学研究所,上海200127;2.中国科学院上海营养与健康研究所,上海200031[摘要]目的·结合Dox 诱导型单链导向RNA (single guide RNA ,sgRNA )表达载体和Cas9转基因小鼠,构建诱导型CRISPR/Cas9系统用于小鼠免疫细胞基因功能研究。
方法·根据四环素诱导表达系统原理,基因合成U6-TetO-sgRNA 和EF1α-T2A-Puro-BFP-T2A-TetR 片段。
通过同源重组将2个片段组装进反转录病毒载体骨架,获得Dox 诱导型sgRNA 反转录病毒载体。
为了验证系统有效性,分离Cas9转基因小鼠骨髓细胞并诱导其向巨噬细胞方向分化。
设计对照(non-targeting control ,NC )组和实验组(靶向F4/80)的sgRNA ,利用反转录病毒感染细胞,分化条件设置添加Dox 组(Dox +)和不添加Dox 组(Dox -)。
通过流式细胞术和T7核酸内切酶Ⅰ(T7endonuclease Ⅰ,T7E Ⅰ)实验检测基因敲除效果。
结果·①成功构建Dox 诱导型sgRNA 反转录病毒表达载体和Cas9转基因小鼠。
②流式结果显示,在NC Dox -组、NC Dox +组和F4/80Dox -组中,几乎无F4/80阴性细胞群体;而在F4/80Dox +组中,F4/80阴性细胞群体高达50%。
③T 7E Ⅰ结果显示,在F4/80Dox -组中,DNA 条带完整,而在F4/80Dox +组中发生基因突变,DNA 条带被切开。
药学概论论文2000字

药学概论论文2000字对社会药学的初步认识摘要:药物与社会中的每一个人,男人与女人,老人与小孩,病人及其家庭以及社会医疗保障系统均有着密切的关系,同样药物与国家的预算,立法与执法管理也有着密切的关系,并与社会经济发展,科技术进步产生相互影响。
药物在使用过程中出现的不按医嘱使用,误用,滥用所引起的不良反应,中毒,药源性疾病以及药物非医学使用所带来的不仅是生物现象,更多是造成严重的社会问题:如药源性疾病造成的致畸、致聋、致残,吸毒引起的犯罪,酗酒引起的暴力和车祸伤人,吸烟造成的心血管疾病和呼吸系统疾病将消耗大量的卫生资源等,所以这些涉及社会的问题,仅从药和药学的角度考虑是无法解决的,必须与社会学、人文科学密切结合才能获得满意的解决,因此发展起了社会药学这门科学。
它是应用社会学观点和方法研究药学,是社会科学和自然科学相结合的边缘学科,是综合应用社会学与人文科学和基础与临床医药学以及自然科学等学科的最新知识来改善病人的药物治疗。
社会药学有社会学,当然也离不开药学。
药又分为传统药和现代药,传统药一般指各国历史上流传下来的,主要是动1物、植物和矿物药,最博大精深的应书中国的传统药,即中药。
早在商周时期,中医药学便有了萌芽,在已出图的殷商甲骨文中,便有大量疾病的名称。
大约成书于春秋战国时期的《黄帝内经》,是我国现存最早的医学典籍之一,东汉末期的药物学专著《神农本草经》,是我国现存最早的药物学专著,为中药学的发展奠定了一定基础,明代伟大医学家、药学家李时珍的《本草纲目》,载药1892种,附图1109幅,附方11000多首,该书综合了16世纪以前动物学、植物学、矿物和冶金学等多学科知识,其影响超出了本草学范围,并于17世纪末,先后以多种文字译本传至海外。
中药的应用涉及到各医学领域,如常见的感冒,则可用麻黄、桂枝等配伍;清热泻火则用石膏、知母、栀子等;风湿引起的各种疾病则用独活、防己,威灵仙等;安神理气用酸枣仁、柏子仁、陈皮、枳实等;另外还有补虚药,如人参、党参、当归、黄芪、地黄、沙参、麦冬等。
人源乳腺癌噬菌体抗体库的构建及初步筛选

MC _ 结合 活 性 癌 噬 菌 体抗 体 库 中 筛选 到 6 特 异 性 噬 菌体 抗 体 , 下一 步进 行 单 链 抗 体 的 株 为
乳 腺 癌放 射 性 核 素 显 影 及 治 疗 奠 定 了基 础 。
关 键词 : 腺 癌 ; 菌 体 抗 体 库 ; 乳 噬 筛选 ; 链 抗 体 单 中 图分 类 号 : 7 7 9 R 3 . 4 R 3 . ; 7 0 4 文献 标 识 码 : A 文 章 编 号 :6 18 4 (O O 0 —5 6O 1 7-3 8 2 1 ) 50 1一3
T ANG h — i , h o ln , S u b n LIS a —i PENG e e 1 Ji , t . a
.
( . e a t n o c l y, is P o l s s i l f Ne in Ne i n S c u n 6 1 0 , ln ; 1D p r me t f On o g F rt ep e p t i a g, i a g, i a 4 0 0 C ̄ a o Ho a o j j h i 2 De a t n f Nu l r d c e C l g f B s d c e C o g ig Me i l n v ri C o g ig 4 0 1 , hn ) . p rme t c a o e Me ii , ol e a i Me ii , h n qn d c iest h n q n 0 0 6 C i a n e o c n a U y,
EL S Re u t A h g n io y o . I A. s l s p a e a t d f 4 2× 1 wa b a n d a d s x a tv l n s a an tb e s a c r e l MCF 7 we e g i e b 0 s o t i e n i ci e co e g i s r a tc n e el - r an d fo t e S f i r r . o cu in S x a t o y bn i g t r a tc n e e l r to g y we e ie t id fo h ma iep a ea — r m h c v l a y C n lso i n i d i d n o b e s a c rc l mo e s r n l r d n i e r m u n z h g n b b f t o y l r r . i wo k p o ie st e b ssf rr d o u l e i g n n h r p o r a tc n e . i d i a y Th s b b r r v d s u h a i o a in ci ma i g a d t e a y f r b e s a c r d
抗体库的构建过程

抗体库的构建过程一、引言抗体库是指收集、保存和管理大量的抗体样本,可以应用于生物医学研究、药物研发等领域。
抗体库的构建过程包括样本采集、抗体制备、筛选和管理等多个环节。
本文将详细介绍抗体库的构建过程。
二、样本采集1.确定采集对象:根据研究目的和需求,确定采集对象。
常见的采集对象包括动物(如小鼠、大鼠、兔子等)、人类血清和细胞系等。
2.采集条件:在保证样本质量的前提下,选择合适的采集条件。
例如,在动物实验中应注意麻醉方式和剂量,避免对动物造成伤害;在人类血清采集中应注意消毒和针头选择等。
3.样本处理:对于不同类型的样本,需要进行不同的处理方法。
例如,对于血清样本需要离心分离血清,去除红细胞等。
三、抗体制备1.免疫原选择:根据研究目的和需求,选择合适的免疫原。
常见的免疫原包括蛋白质、多肽、细胞表面分子等。
2.免疫程序:根据免疫原的特性和抗体制备的需求,选择合适的免疫程序。
常见的免疫程序包括单次免疫、多次免疫等。
3.抗体纯化:通过蛋白A/G亲和层析、离子交换层析等方法纯化抗体。
四、筛选1.ELISA筛选:通过ELISA方法对抗体进行筛选。
将免疫原包被在96孔板上,加入抗体样本,然后加入二抗进行检测。
2.流式细胞术筛选:通过流式细胞术对抗体进行筛选。
将细胞表面分子作为免疫原进行免疫,然后用荧光标记的二抗进行检测。
五、管理1.样本存储:将制备好的抗体样本存储在低温冰箱或液氮罐中,保证其质量和稳定性。
2.信息管理:建立完整的信息管理系统,记录每个样本的来源、制备过程和特性等信息。
六、总结抗体库是生物医学领域中重要的资源之一,其构建过程需要经过样本采集、抗体制备、筛选和管理等多个环节。
在每个环节中,都需要注意细节,保证样本的质量和稳定性。
建立完整的信息管理系统,有助于提高抗体库的效率和可靠性。
VIGS实验方案

VIGS实验方案实验名称:VIGS(Virus-induced gene silencing)实验方案实验目的:通过病毒感染,引起基因静默,研究静默基因的功能和作用机制。
实验步骤:1.实验前准备-准备工作台、培养箱和所需设备材料。
-准备实验所需的植物材料(如拟南芥等)。
-配制所需的酶切酶、聚合酶、缓冲液等。
2.构建VIGS载体-选择合适的载体,并在该载体上插入目标基因的片段。
-将载体经酶切、连接、转化等步骤进行构建。
3.病毒感染- 将构建好的VIGS载体通过植物冠状病毒(例如Tobacco Rattle Virus,TRV)进行感染。
-制备病毒悬液,包括双病毒载体的混合物,通过注射或喷雾的方式进行感染。
4.病毒复制和扩增-在感染后的植物材料中,病毒会进行复制和扩增。
-收集感染后的植物部位,如叶片、茎等进行后续研究。
5.基因静默检测-取感染后的植物材料,进行总RNA的提取。
-利用逆转录酶将RNA转录为cDNA。
-进行PCR扩增,利用靶向基因的特异引物进行检测。
-利用凝胶电泳分析PCR产物,判断基因是否被成功静默。
6.静默基因功能研究-验证基因静默的效果和稳定性,可经过酶活测定、组织分析等方法进行。
-通过研究静默基因表型的变化,探讨其功能和作用机制。
7.数据分析和统计-根据实验结果,进行数据整理和统计分析。
-利用统计学方法,对数据进行可视化和解释。
-对结果进行讨论和对比研究。
实验注意事项:1.实验操作需要严格按照实验室安全操作规范进行,保证实验的安全进行。
2.构建VIGS载体时,在选择片段和选择载体时需严谨,确保成功插入目标基因。
3.病毒感染要注意避免交叉感染,用透明的手套和消毒工具进行操作。
4.实验过程中需要设置对照组,以确保实验结果的可靠性和准确性。
5.RNA提取和PCR扩增过程中,要严格遵循无污染的操作,以保障实验效果。
6.结果分析和统计时,要进行充分的数据量化和方法论的应用,保证分析结果准确。
基于CRISPR

第38卷第2期2024年3月山东理工大学学报(自然科学版)Journal of Shandong University of Technology(Natural Science Edition)Vol.38No.2Mar.2024收稿日期:20230301基金项目:江苏省高校 青蓝工程 项目(2023);江苏省高职院校青年教师企业实践项目(2021QYSJ063);江苏省精准诊疗药物创制工程研究中心(苏州大学)开放课题(SDGC2239)第一作者:谢钰珍,女,295842442@;通信作者:覃鸿妮,女,qinhn@文章编号:1672-6197(2024)02-0067-06基于CRISPR /Cas9基因编辑技术构建PD -L1-GFP 报告基因HT29细胞株谢钰珍1,覃鸿妮1,2,吴凡1,孙曙光1,孟丽君3(1.苏州工业园区服务外包职业学院生物科技学院,江苏苏州215123;2.江苏省精准诊疗药物创制工程研究中心(苏州大学),江苏苏州215000;3.苏州东岭生物技术有限公司,江苏苏州215123)摘要:利用CRISPR /Cas9基因编辑技术和同源重组技术在PD -L1基因的特定位置敲入绿色荧光蛋白(GFP )序列,构建含有PD -L1-GFP 报告基因的结肠癌细胞(HT29)稳定细胞株㊂根据CRISPR -Cas9靶点设计原则,针对PD -L1基因终止密码子设计两对sgRNA ,退火形成双链后连接至Lenti -V2空载质粒中,转化培养后提取质粒并测序验证㊂对于正确的Lenti -V2-sgRNA 重组质粒,T7E1酶切验证其基因编辑效率㊂根据靶点位置设计左同源臂+GFP +右同源臂序列合成Donor 片段,双酶切后连接至pUC19,重组载体转化扩增后同样进行质粒提取和测序验证㊂将验证成功的Lenti -V2-sgRNA 和pUC19-donor -GFP 共转染HT29细胞,荧光显微镜检测GFP 表达情况,菌落PCR 及基因测序验证GFP 报告基因的靶向插入效果㊂经酶切和测序鉴定,靶向编辑PD -L1的Cas9载体Lenti -V2-gRNA 和含GFP 基因的Donor 质粒pUC19-donor -GFP 构建成功㊂两个重组质粒转染HT29细胞后,显微观察㊁多克隆验证㊁单克隆筛选及鉴定结果显示,GFP 成功转入HT29细胞并进行了表达,经有限稀释法筛选出的单克隆细胞荧光均一,且与对照组相比,阳性细胞克隆的基因组PCR 均检测到特异条带,表明在PD -L1终止密码子前成功插入了GFP 片段,细胞株构建成功㊂通过基因编辑技术成功构建了稳定表达PD -L1-GFP 报告基因的HT29结肠癌细胞株,建立了一个可以直观在细胞上观察PD -L1是否表达以及表达的强弱程度的系统,为后期体内外筛选调控PD -L1的上游新靶点及药物奠定了基础㊂关键词:PD -L1;CRISPR /Cas9;报告基因;GFP 中图分类号:TB532.1;TB553文献标志码:AConstruction of HT29cell line with PD -L1-GFPreport gene by CRISPR /Cas9systemXIE Yuzhen 1,QIN Hongni 1,2,WU Fan 1,SUN Shuguang 1,MENG Lijun 3(1.School of Biotechnology,Suzhou Industrial Park Institute of Services Outsourcing,Suzhou 215123,China;2.Jiangsu Province Engineering Research Center of Precision Diagnostics and Therapeutics Development,Soochow University,Suzhou 215000,China;3.Suzhou Dongling Biotechnology Company Limited,Suzhou 215123,China)Abstract :Using CRISPR /Cas9and homologous recombination technology to tap green fluorescent protein (GFP)sequence at specific position of PD -L1gene,we constructed human colon cancer cell (HT29)㊀stable cell line containing PD-L1-GFP reporter gene.According to the design principle of CRISPR-Cas9target,two pairs of sgRNAs were designed for the stop codon of PD-L1gene.After annealing,the sgRNAs were connected to the Lenti-V2plasmid.The plasmid was extracted and verified by sequencing after amplification of the receptor cells.For the correct Lenti-V2-gRNA recombinant plasmid,the effi-ciency of gene editing was verified by T7E1digestion.According to the target location,Donor fragment was synthesized from left homologous arm+GFP+right homologous arm sequence,which was ligated to pUC19after double enzyme digestion.Plasmid extraction and sequencing were also performed after re-combinant vector transformation and amplification.HT29cells were co-transfected with Lenti-V2-gRNA and pUC19-donor-GFP,and the expression of GFP protein was detected by fluorescence microscopy. Finally,the targeted insertion effect of GFP reporter gene was verified by colony PCR and gene sequen-cing.The Cas9vector Lenti-V2-gRNA and Donor plasmid pUC19-donor-GFP containing PD-L1were successfully constructed by enzyme digestion and sequencing.After the two recombinant plasmids were transfected into HT29cells,the results of microscopic observation,polyclonal verification,monoclonal screening and identification showed that GFP was successfully transferred into HT29cells and expressed, and the monoclonal cells screened by limited dilution method had uniform fluorescence.Moreover,com-pared with the control group,specific bands were detected in the genomic PCR of the clones of positive cells,indicating that the GFP fragment was successfully inserted before the PD-L1stop codon,and the cell line was successfully constructed.HT29colon cancer cell line with stable expression of PD-L1-GFP reporter gene was successfully constructed by gene editing technology,and a system was established to di-rectly observe whether PD-L1is expressed and the degree of expression,which laid a foundation for sub-sequent in vitro and in vivo screening of upstream new targets and drugs regulating PD-L1. Keywords:PD-L1;CRISPR/Cas9;reporter gene;GFP㊀㊀PD1(programmed death-1,程序性死亡受体-1)为PDCD1基因编码的Ⅰ型跨膜糖蛋白,主要表达于活化的免疫细胞表面,如T细胞㊁B细胞㊁NK细胞以及单核细胞等㊂PD1是维持自身耐受性的重要因子,在生理条件下可通过TCR识别抗原,调节外周组织中T细胞的功能,从而应对和清除外源病菌及内源异常细胞㊂PD-L1(程序性死亡配体-1)也称CD274,是PD1的配体之一,是由CD274基因编码的一种细胞表面糖蛋白,多过度表达于肿瘤细胞表面[1]㊂作为重要的负性免疫调节因子,当T细胞表面PD1受体与肿瘤细胞表面表达的PD-L1配体结合后,可向细胞内传递调控信号,抑制T细胞活化与增殖,从而帮助肿瘤细胞逃脱宿主的免疫监视,因此,抑制PD1/PD-L1通路或者通过特异性靶点抑制PD-L1蛋白的表达,可有效增强肿瘤治疗效果㊂近年来的临床研究结果显示,由PD1/PD-L1通路介导下的免疫抑制在非小细胞肺癌㊁胃癌㊁乳腺癌㊁大肠癌等多种恶性肿瘤的发生发展中均具有重要作用,PD-L1在以上各种癌组织的表达明显升高,且PD-L1的阳性表达与肿瘤大小㊁转移情况㊁分化程度及远期生存率存在密切关系[2-3];此外,从诱导肿瘤细胞表面高表达PD-L1的相关因素来看,目前已报道的肿瘤细胞中调控PD-L1表达的相关信号通路主要有Akt3信号通路和MAPK信号通路㊁IFN-γ信号通路和AR信号通路[4],而且这些通路的某些抑制剂已被证明可调节PD-L1㊂例如:TGFβR-1抑制剂LY364947可明显降低TGFβR-1诱导的PD-L1mRNA和蛋白表达[5];突变转录因子FOXP3在PD-L1启动子区的结合位点后,胰腺癌细胞PD-L1的表达明显受到抑制[6];但关于结肠癌肿瘤细胞PD-L1表达的具体调控机制仍需进一步研究㊂CRISPR/Cas9系统是一种新型基因编辑技术,通过sgRNA介导核酸酶Cas9对基因组特定位点进行识别㊁切割,从而实现基因编辑,操作相对简便,基因编辑效率高㊂本文利用CRISPR/Cas9技术,在PD-L1基因终止密码子之前插入绿色荧光蛋白GFP基因,通过构建稳定表达PD-L1-GFP报告基因的结肠癌细胞株HT29,旨在建立一个可直观观察细胞上PD-L1是否表达以及表达强弱的系统,为进一步研究结肠癌细胞中PD-L1表达的可能调控机制提供帮助,为后期体内外筛选调控PD-L1的上游新靶点及药物奠定基础㊂86山东理工大学学报(自然科学版)2024年㊀1㊀材料与方法1.1㊀材料HT29细胞和293T细胞(苏州东岭生物技术有限公司保存),Lenti CRISPR V2质粒载体和pUC19载体(苏州东岭生物技术有限公司),胶回收试剂盒(Thermo),Stbl3感受态大肠杆菌(TransGen Biotech),DMEM培养基和D10培养基(HYclone), T4连接酶(Takara公司),DNA聚合酶(NEB)㊂1.2㊀方法1.2.1㊀sgRNA引物的设计与合成根据NCBI查询的PD-L1基因序列,使用CRISP在线设计工具(/E-CRISP/),根据靶点设计原则,在PD-L1基因终止密码子处设计sgRNA,并在序列正义链与反义链的5 端添加Esp3I(BsmBI)酶切位点,合成的具体序列如下:sgRNA-1:GAGGAGACGTAATCCAGCAT gRNA-1-F:CACCGAGGAGACGTAATCCAGCA gRNA-1-R:AAACATGCTGGATTACGTCTCCTC sgRNA-2:GTCTCCTCCAAATGTGTATC gRNA-2-F:CACCGTCTCCTCCAAATGTGTATC gRNA-2-R:AAACGATACACATTTGGAGGAGA为检测突变是否成功,设置了一对检测引物,扩增产物为包含sgRNA靶点的基因组DNA片段,检测引物具体序列如下:Test-F:TGGGGGACAAGCCATCCCAA Test-R:ATGATTTGCTTGGAGGCTCC1.2.2㊀LentiV2-gRNA重组质粒的构建及鉴定根据所设计序列合成单链sgRNA,经梯度降温PCR退火形成双链sgRNA㊂退火结束后,将得到的双链gRNA连接到已用Esp3I酶切回收后的线性Lenti-V2空载质粒中,连接产物转化Stbl3感受态细胞,37ħ培养过夜后挑取阳性单克隆进行测序验证,测序引物:LKO1-5(GACTATCATATGCTTAC-CGT)㊂针对序列正确的单克隆,提取其对应菌液中的质粒DNA,即可得到正确的Lenti-V2-sgRNA重组质粒㊂1.2.3㊀pUC19-Donor-GFP重组质粒的构建及鉴定设计左同源臂+GFP+右同源臂序列,序列两端添加酶切位点XbaI/BamHI,送公司合成Donor片段,将合成的Donor片段㊁pUC19分别用XhaI和BamHI双酶切后连接㊂取10μL连接产物转化至50μL Stbl3感受态大肠杆菌中,37ħ培养1h 后,均匀涂在含有Amp的LB平板上,过夜培养㊂次日挑取6个克隆于4mL含有Amp的LB培养基中, 37ħ培养16h㊂16h后取1mL菌液抽提质粒,并对质粒进行菌落PCR,判断选取的单克隆中是否含有目的片段㊂将验证正确的质粒送至测序,测序引物:M13-R(CAGGAAACAGCTATGACC),测序正确后冻存菌种㊂1.2.4㊀细胞培养和细胞转染从-80ħ冰箱中取出冻存的HT29细胞,复苏结束后留2mol/L的细胞量在10cm培养皿中培养,隔天进行传代培养㊂培养至第4天时将HT29转移至24孔板中的2孔(一孔转染,一孔阴性对照),每孔0.5ˑ106细胞㊂转染前1~2h更换新鲜培养基(0.9mL/孔),按Lenti-V2-sgRNA(0.6μg)㊁Donor-GFP(0.4μg)㊁1mg/mL PEI(3μL)㊁DMEM(97μL)配制转染体系,室温孵育30min后,将混合液加入到1孔中,另一个孔无须加入,轻轻摇匀后放入培养箱培养(37ħ,5%CO2)㊂1.2.5㊀多克隆效果验证提取转染后的HT29细胞基因组DNA,利用在PD-L1基因组以及GFP上分别设计的上下游引物,对其进行PCR鉴定,以检测其中是否含有在PD-L1位点成功插入GFP片段的目的细胞㊂引物序列: PD-L1-F(TTCAAATTTATCATTTATCA),GFP-R (CCGGACACGCTGAACTTGTG),PCR体系:模板DNA10ng㊁PD-L1-F和GFP-R各1μL,2X PrimeSTAR HS DNA Polymerase MIX10μL,总体积20μL㊂PCR扩增程序:98ħ预变性20s,98ħ变性10s,55ħ退火5s,72ħ延伸35s,共30个循环,68ħ彻底延伸2min㊂扩增的PCR产物经1%琼脂糖凝胶电泳分析㊂1.2.6㊀单克隆细胞筛选转染72h后,观察细胞生长状况并计数,采用有限稀释法,用D10(10%的FBS+DMEM)按每200μL一个细胞稀释到96孔板中,培养24h 后,显微镜观察荧光及细胞状态,并做好标记㊂继续培养,当上一步标记的单克隆细胞密度达到80%后,消化并收集细胞,用基因组DNA抽提试剂盒提取基因组,作为检测引物(TestF㊁R)的扩增模板,最后将扩增产物送至公司测序,以检测PD-L1-GFP 报告基因是否插入成功㊂96第2期㊀㊀㊀㊀㊀谢钰珍,等:基于CRISPR/Cas9基因编辑技术构建PD-L1-GFP报告基因HT29细胞株2㊀结果与分析2.1㊀Lenti -V2-gRNA 重组质粒构建结果重组质粒基因测序结果显示,在酶切位点之间插入的片段位置㊁方向以及序列与预期一致(图1),含有本实验所需要的两条sgRNA 序列,证明LentiV2-gRNA 重组质粒构建成功㊂2.2㊀两条sgRNA 切割效果验证结果为了验证所设计的两条sgRNA 的切割效果,将两种Lenti -V2-gRNA 重组质粒转入293T 细胞后提取基因组DNA,并对其进行T7E1酶切鉴定,结果显示1和2两种sgRNA 都能切下相应大小的条带(图2),证明Lenti -V2-sgR1和Lenti -V2-sgR2都具有切割效果,本实验随机使用其中1条,采用sgRNA2,即Lenti -V2-sgR2㊂图1㊀Lentiv2-gRNA重组质粒测序结果图2㊀T7E1酶切图2.3㊀pUC19-Donor -GFP 重组质粒构建结果以挑选的6个单克隆为模板,经菌落PCR 均能扩增出目的片段(图3),说明菌落中含有目标质粒㊂将阳性质粒进一步送至金唯智测序,从图4可以看出,第一行的序列为测序结果,第二行的序列是所需的模板序列,序列比对完全正确,pUC19-Donor -GFP 质粒构建成功㊂2.4㊀Lenti -V2-sgR2和pUC19-donor -GFP 细胞转染结果㊀㊀将Lenti -V2-sgR2和pUC19-donor -GFP 转染至HT29细胞,72h 后荧光显微镜下观察细胞状态㊂由图5可知,培养72h后可观察到少量细胞表达绿注:1-6为随机挑取的6个单克隆,M 为DNA marker㊂图3㊀pUC19-Donor -GFP 重组质粒转化质粒菌落PCR 结果色荧光,证明GFP 成功转入HT29并进行了表达㊂2.5㊀PD -L1-GFP 报告基因工程细胞株阳性单克隆筛选结果2.5.1㊀多克隆效果验证结果图6为多克隆PCR 鉴定结果,可以发现实验组在200bp 位置有目的条带,而对照组没有,证明GFP 插入到指定位点㊂2.5.2㊀单克隆细胞筛选结果为将上述验证的插入正确GFP 序列位点的细胞挑选出来,将多克隆细胞进行单克隆筛选,图7为荧光显微镜观察结果,可以看到细胞形成了单克隆并且具有均一的荧光,可以进行后续的验证㊂7山东理工大学学报(自然科学版)2024年㊀图4㊀pUC19-Donor重组质粒测序结果(a)细胞图(b)荧光图图5㊀HT29细胞荧光蛋白表达为验证上述挑选的单克隆中所需的序列存在,对其基因组PCR 之后送至金唯智测序,图8显示与正常HT29细胞基因组相比,敲入的实验组所挑的单克隆在终止密码子前插入了GFP 片段,证明细胞系构建成功㊂3㊀讨论PD1表达于活化的免疫细胞表面,如B淋巴细图6㊀多克隆验证琼脂糖凝胶电泳图图7㊀单克隆细胞荧光状态图胞㊁CD4+T 细胞等㊂PD -L1作为PD1的配体之一,在癌组织上可见异常高表达,如肾癌㊁肝癌㊁肺癌㊁乳腺癌及卵巢癌等;但在肿瘤邻近的正常组织中低水平表达,提示它参与肿瘤发生发展,并在削弱抗肿瘤免疫反应中发挥重要作用㊂PD1/PD -L1是负性共刺激分子,当肿瘤细胞表面的PD -L1与免疫细胞表面的PD1结合时,可抑制免疫细胞的增殖与活性甚至诱导其凋亡,使癌细胞成功逃脱免疫杀伤㊂近年来,PD -L1及其受体PD1信号通路一直是肿瘤免疫领域的热门研究对象,针对阻断PD1/PD -L1通路的单克隆抗体已然成为了肿瘤免疫治疗的明星产品,目前FDA 已经批准了五种PD -L1抑制剂㊂作为非特异性免疫治疗产品,PD -L1抑制剂的抗癌效应具有广谱性,且在不同癌症疾病中显示出了很好的治疗[7-11],但由于治疗过程中的原发性和获得性耐药,相当大比例的患者无法从中受益㊂相较于单药疗法,近年来越来越多的研究正向着PD1/PD -L1耐药机制研究以及联合用药靶点的筛选上转移[12]㊂在一些研究报道中,针对不同类型免疫检查点的联合疗法已被证明对几种肿瘤有效㊂例如:采用抗PD -1抑制剂抗体㊁抗CD137激动剂抗体和疫苗治疗的三联疗法可以显著增强胰腺导管腺癌的治疗效果[13-14];联合anti -PD -L1和anti -TIGIT 在临床上对转移性NSCLC 患者非常有效[15];增强ITCH 活17第2期㊀㊀㊀㊀㊀谢钰珍,等:基于CRISPR /Cas9基因编辑技术构建PD -L1-GFP 报告基因HT29细胞株性可促进PD -L1泛素化降解从而降低肿瘤细胞PD -L1表达,化合物AK087与MAPK 抑制剂联用可明显增强MAPK 靶向治疗黑色素瘤的效果[16]等㊂但对于其他大部分肿瘤来说,不同疗法治疗过程中肿瘤表面PD -L1的表达量变化情况及产生治疗抗性的关键机制,仍需进一步研究㊂注:斜体为GFP 片段,下划线为PDL1终止密码子,WT 为正常HT29测序序列,Knock -in clone 为实验组测序序列㊂图8 对照组和实验组序列对比图㊀㊀为了探究结肠癌细胞表面PD -L1表达的更多可能调控机制,本研究利用CRISPR /Cas9系统和同源重组的原理,通过两次转染将GFP 荧光基团成功插入到PD -L1基因终止密码子之前,成功构建了PD -L1-GFP 报告基因HT29细胞株,通过荧光信号直观观察细胞上PD -L1是否表达以及表达的强弱程度,为进一步研究结肠癌细胞中PD -L1表达的可能调控机制提供了材料,并为后期体内外筛选调控PD -L1的上游新靶点及药物奠定了基础㊂参考文献:[1]谢丽叶,付杰军,卢奕,等.PD1/PD -L1激活促进癌症发生㊁发展和转移的研究进展[J].肿瘤防治研究,2017,44(6):423-427.[2]车章洪.PD -1/PD -L1通路在肿瘤的发生发展过程中对T 细胞的活化作用研究进展[J].中国新药杂志,2017,26(16):1913-1917.[3]方宇,王海娟,李征洋,等.PD -L1和A2aR 在大肠癌中的表达及意义[J].河北医药,2021,43(3):345-348,352.[4]丰江舟,杨梦梦,朱彬彬,等.人PD -L1基因启动子克隆及其荧光素酶报告基因载体的构建[J].皖南医学院学报,2016,35(6):524-526.[5]柯佳,李卓伟,李超,等.TGF -β1对结肠癌SW620细胞及其PD -L1表达的影响[C]//中国解剖学会.中国解剖学会2019年年会论文文摘汇编.昆明:解剖学杂志编辑部,2019:182.[6]王秀超.胰腺癌肿瘤细胞PD -L1表达调控机制及联合干预实验研究[D].天津:天津医科大学,2017.[7]李青丽,唐瑶,李富丽,等.抗PD -L1抗体抑制小鼠非小细胞肺癌移植瘤的生长及其机制[J].肿瘤,2020,40(8):531-540.[8]孙晨,镡云辉,耿波,等.PD -1/PD -L1抑制剂在肾癌中的研究进展[J].国际外科学杂志,2020,47(9):639-643.[9]张丽娜,杨艳芳,姜战胜.PD -1/PD -L1抑制剂治疗三阴性乳腺癌的研究进展[J].中国肿瘤生物治疗杂志,2021,28(8):844-849.[10]李一鑫,路丹.PD -1/PD -L1抑制剂治疗晚期卵巢癌的研究进展[J].现代肿瘤医学,2021,29(15):2741-2744.[11]陈茜,周建英,黎扬斯.抗PD -1/PD -L1治疗在晚期难治性肺鳞癌中的疗效和安全性[J].循证医学,2015,15(4):213-216.[12]YUAN Y,ADAM A,ZHAO C,et al.Recent advancements in themechanisms underlying resistance to PD -1/PD -L1blockade immu-notherapy[J].Cancers(BaseⅠ),2021,13(4):663.[13]韩丽炘,黄玉,温娟娟.PD -1/PD -L1抑制剂联合化疗对比免疫单药治疗晚期NSCLC 疗效及安全性的Meta 分析[J].山西大同大学学报(自然科学版),2022,38(4):84-90.[14]汝继轩,吴川林,张占田,等.PD -1/PD -L1免疫检查点抑制剂治疗胰腺癌研究进展[J].中华胰腺病杂志,2021,21(2):143-147.[15]陈丽丽.EGFR 联合PD -L1在可切除或临界可切除胰腺癌中的预后评价作用[C]//第二届中国临床分子诊断大会.第二届中国临床分子诊断大会论文集.成都:出版者不详,2019:73.[16]YANG Z T,WANG Y,LIU S X,et al.Enhancing PD -L1degra-dation by ITCH during MAPK inhibitor therapy suppresses acquiredresistance[J].Cancer Discovery,2022,12(8):1942-1959.(编辑:姚佳良)27山东理工大学学报(自然科学版)2024年㊀。
免疫组学的研究进展

免疫组学的研究进展唐康侯永利王亚珍陈丽华(中国人民解放军空军军医大学基础医学院免疫学教研室,西安 710032)中图分类号R392.9 文献标志码 A 文章编号1000-484X(2024)01-0185-07[摘要]随着高通量测序技术、生物信息学等相关领域进展以及人类对免疫系统功能认识的逐步深入,免疫组学从最初解析B细胞受体(BCR)、T细胞受体(TCR)基因序列逐渐发展为解析和绘制宿主免疫系统和抗原的互作关系以及宿主免疫系统应答机制的全景图谱,主要包括抗原表位组学、免疫基因组学、免疫蛋白质组学、抗体组学和免疫信息学等方面的研究,并基于大量免疫学研究数据建立了ImmPort、VDJdb和IEDB等免疫学数据库,加速了新抗原表位的发现和免疫应答机制等研究。
免疫组学能够揭示免疫系统与疾病的关联,促进新型疫苗和免疫治疗策略开发,将有效推动个体化医疗和精准药物治疗。
近年免疫组与暴露组等的整合以及与人工智能的融合将对全面理解免疫系统对环境因素的响应和调节机制、解析疾病发生和发展的分子机制产生重大影响。
[关键词]免疫组;免疫组学;免疫信息学;人工智能Advances in immunomics researchTANG Kang, HOU Yongli, WANG Yazhen, CHEN Lihua. Department of Immunology, School of Basic Medicine,Air Force Medical University, Xi'an 710032, China[Abstract]With the progress of high-throughput sequencing technologies and bioinformatics, and deepening understanding of immune system,immunomics has evolved from initially deciphering gene sequences of B cell receptor (BCR)and T cell receptor (TCR) to unraveling and mapping interactions between host immune system and antigens, as well as panorama of host immune system response mechanisms, which now encompasses various research areas, such as antigen epitopeomics, immunogenomics, immunopro‐teomics, antibodyomics and immunoinformatics. Based on a large amount of immunological research data, immunological databases such as ImmPort, VDJdb and IEDB have been established to accelerate discovery of new antigen epitopes and study of immune response mechanisms. Immunomics has revealed the association between immune system and diseases, promoted the development of novel vac‐cines and immunotherapeutic strategies, and effectively drove the development of personalized medicine and precision medicine. In recent years, integration of immunome with exposome and fusion it with artificial intelligence will have a significant impact on compre‐hensively understanding immune system's response and regulatory mechanisms to environmental factors, as well as deciphering molecular mechanisms underlying disease occurrence and progression.[Key words]Immunome;Immunomics;Immunoinformatics;Artificial intelligence免疫组(immunome)是宿主免疫系统与抗原的互作关系以及宿主免疫系统应答机制的全景图谱,包括免疫系统的识别对象、识别受体以及参与免疫应答过程的其他分子[1-3]。
一种微生物基因数据库的构建方法及系统与流程

一种微生物基因数据库的构建方法及系统与流程微生物基因数据库的构建方法及系统与流程构建一个微生物基因数据库是进行微生物基因组学研究和分析的关键步骤之一。
本文将介绍一种常用的微生物基因数据库构建方法及相关的系统与流程。
方法一:基因组序列获取与预处理1. 选择目标微生物,获取其完整的基因组序列。
可以通过文献、公共数据库或合作伙伴提供的数据来获取。
2. 对获取的基因组序列进行预处理。
例如,去除低质量区域、过滤掉不完整的序列或重复的序列,并进行基因预测和注释。
方法二:数据库结构设计与构建1. 设计数据库结构。
根据微生物基因组的特点,设计相应的表结构,包括基因、蛋白质、转录组、调控元件等的信息表,以及相应的关系表。
2. 创建数据库。
使用关系型数据库管理系统(如MySQL、Oracle等)或非关系型数据库(如MongoDB)创建数据库,并创建相应的表结构。
方法三:数据导入与整合1. 根据预处理后的基因组序列,将基因、蛋白质、转录组等相关信息导入到相应的表中。
可以使用自己开发的脚本或专门的数据导入工具进行操作。
2. 整合外部数据。
整合其他公共数据库或相关资源中的数据,如基因序列、注释信息、代谢途径等,以丰富数据库内容。
方法四:数据库管理与维护1. 设置数据库的访问权限和安全措施,确保数据的安全性。
2. 定期更新数据库。
随着新的微生物基因组数据不断产生,需要定期更新数据库的内容,以确保数据的实时性。
3. 监测数据库性能。
定期对数据库进行性能监测和优化,以提高数据库的查询效率和响应速度。
系统与流程:1. 用户访问数据库平台。
提供一个用户友好的界面,允许用户进行数据查询、下载和分析。
2. 用户查询与分析。
用户根据需求选择适当的查询条件,在数据库中进行检索,并获取所需的微生物基因组数据。
3. 数据展示与分析结果。
将查询结果以表格、图形或其他形式展示给用户,方便用户对基因和相关信息进行分析和解读。
4. 数据下载与导出。
用户可以选择将查询的数据进行下载或导出,以便进行进一步的分析和研究。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
tmpaefrP mpic t no a a l e in f nio is T ev r bergo so g t h i ( L a d e lt o CR a l ai f r bergo so t de. h a a l e in f ih an v ) n i f o vi a b i l c
生 物工 程
食ቤተ መጻሕፍቲ ባይዱ研究与开发
F o sa c d De eo me t o d Re e rh An v lD n
21 0 2年 7月 第3 3卷第 7期
西维 因抗体库的构建及特异性克隆的筛选
王硕 , 刘昕煜 , 吴懿娜 , 杜欣军 , 朱超 , 张洁
( 品营养 与安全教育部重点 实验室 , 食 天津科 技大学 食 品工程 与生物技术学 院 , 天津 3 0 5 ) 0 4 7
i h bto a eo ei n i i n r t ft mmu e s r m e c e 5 0 n 6% r s e t ey Toa i h n e u r a h d 2 4 0 0 a d 2 e p ci l . tl v RNA se ta td fo t e wa xr c e r m h
分 别 由 18和 l2个 氨 基 酸 构 成 , 链 可 变 区分 别 由 17和 l2个氨 基 酸 构 成 。 0 1 重 1 1
关 键 词 : 维 因 ; 茵体 展 示 ; 组抗 体 西 噬 重
Co sr c i n o n t u to fPha e S Fv Li a y Ag i s r r la lc i n o pe i c Pha e Cl n s g c br r a n tCa ba y nd See to fS cf i g o e
单链抗体基 因。 经过 E o 和 Hi c RI n m酶切 后 , d 连入 1 7噬 菌体臂 , 构建 西维因特异性 的抗体库 , 体库原始 滴度 为 抗
1 4 1 f。 西 维 因一 V .  ̄ 0pu 以 7 O A为 配体 对抗 体 库 进 行 淘选 , 经过 4轮 淘 选 后 , 用 直接 E IA 对 获得 噬 茵 体 克 隆进 行 特 利 LS 异 性 检 测 , 维 因对 克 隆 A — 西 4 7和 A — 6的 抑 制 率分 别达 到 5 . %和 6 . 41 41 6 4%。 过 序 列 测 定 , 克 隆 的轻 链 可 变 区 经 2个
摘 要 : 计 算机 辅 助 下 , 计 并 合 成 西 维 因 半抗 原 , 载 体 蛋 白 K H连 接 后 免 疫 B t/ 鼠 , 过 5次 免 疫后 , 在 设 与 L a C小 b 经 最
高 效 价 和特 异性 抑 制 率分 别 达 到 2 4 0 5 0 0和 2 % ; 取 免 疫后 小鼠 脾 脏 总 R A, 6 提 N 纯化 m N R A后 反 转 录 获 得 c N 利 D A. 用 设计 的 兼 并 引物 获得 长 度 约 为 40b 0 p的轻 链 可 变 区和 重 链 可 变 区基 因 片段 ,重 叠 延 伸后 获得 长度 约 为 7 0 D的 8b
WA GS u, I i- u WUY— aD i-a , HUC a, H N e N h oLUXn y, in, UXn jn Z hoZ A GJ i ( e aoa r o o uri n a t M ns y f d ct n Taj nvrt o i c n eh o g, i j K y b rt y f od tt n d f y iir u a o, i i U i sy f ce e d cn l yTa i L o F N io a S e , t oE i nn e i S n a T o nn
i mmu ie ies le n n z d m c p e n a d mRNA sp rf d T ec wa u i e . i h DNA so tie yr v re ta s rp in a d u e s wa ba n d b e es r n c to n s d a i
3 0 5 , hn ) 0 4 7 C ia
A b t a t Ca b rl h p e , wh c wa d sg e u i g o ue s f r s n s nh sz d y h mia sr c : r a y a tn ih s e in d sn c mp tr ot e a d y te ie b c e c l wa s nh sz d meh ds y te ie t o ,wa o n c e t are r ti H . e l/ c r mmu ie sn h s c n e td wi c rirp oen KL h Th n Bab C mie wee i nz d u i g t e c n e t d p o u t s h i o n ce r d c a t e mmu o e .Afe r u d o mmu iai n h ih s i r a d h s e i c n gn t r5 o n s fi nz t ,t e h g e t tt n te p cf o e i
hayca (H w r a ot 0 pi l g . e e ig - hi a t oygn ( F) b u 7 0b a ev i V ) ee bu 4 0b n t T n h nl can n bd e s v ao t 8 p s hn ne h h t s e i e c w