百合MDHAR基因的克隆与表达分析
兰州百合蔗糖代谢关键酶基因的克隆及原核表达

兰州百合蔗糖代谢关键酶基因的克隆及原核表达淀粉含量是决定百合种球质量的最关键因素。
蔗糖作为百合鳞茎中可溶性碳水化合物的主要形态,可在蔗糖合成酶(Sucrose Synthase,SuSy)和可溶性酸性转化酶(Soluble Acid Invertase,SAI)调控下参与淀粉合成。
为探究百合SuSy和SAI的基因功能及其在蔗糖-淀粉代谢途径中的调节机理,本研究首先克隆得到兰州百合(Lilium davidiivar.unicolor)两个SuSy基因SuSy1和SuSy2的全长cDNA序列,并对其序列进行了生物信息学分析,将其过表达载体和RNAi载体通过农杆菌介导转化兰州百合。
而后构建了兰州百合SuSy及SAI基因的原核表达载体,并在适宜条件诱导融合蛋白表达,为进一步研究百合SuSy和SAI基因功能奠定了基础。
主要研究结果如下:1.SuSy1和SuSy2基因cDNA全长克隆及生物信息学分析以兰州百合扦插鳞片的总RNA为模板,采用RT-PCR及RACE法成功克隆获得SuSy 基因SuSy1和SuSy2全长cDNA序列,其长度分别为2,877 bp和2,865 bp.序列分析结果显示,基因SuSy1和SuSy2的同源性为85.73%,与同科植物同源性为65%-88%,分别编码807和810个氨基酸,其蛋白等电点均为6.10.结构域分析表明,SuSy1和SuSy2均编码含有188个氨基酸的保守结构域,其中包含一个糖基转移酶结构域,与糖基转化酶家族GT1有很高的相似性。
2.SuSy基因转化兰州百合研究利用农杆菌介导转化法,将带有兰州百合SuSy1和SuSy2基因的过表达载体和RNAi载体导入兰州百合。
为优化遗传转化过程,进行了兰州百合小鳞片对潮霉素(Hyg)和头孢霉素(Cef)浓度敏感试验,并对影响转化的各因素进行筛选。
获得最适条件为:20 mg L-1 Hyg为抗性芽筛选的适宜浓度;300 mg L-1 Cef为最适抑菌浓度;预培养2 d,农杆菌菌液培养至OD600为0.6,侵染鳞片20 min,重悬液和共培养基中加入终浓度为100 μM的乙酿丁香酮(AS),共培养3 d.最终获得了 3株带有基因SuSy2片段的RNAi载体的转基因抗性苗,目前正处于筛选阶段。
百合花的基因组和遗传特性研究

百合花的基因组和遗传特性研究百合花(Lilium spp.)是一类受欢迎的花卉,其美丽的花朵和独特的芳香吸引了人们的注意。
百合花具有多样化的颜色、花型和芳香特性,这些特征是由基因组和遗传机制决定的。
近年来,人们对百合花的基因组和遗传特性进行了广泛的研究,旨在深入了解其进化历史、增强品种改良和保护自然遗传资源。
首先,百合花的基因组研究是了解其基因组结构和功能的关键。
基因组是生物体内包含全部遗传信息的DNA的集合。
通过对百合花基因组的测序和分析,研究人员可以识别出其中的基因,了解其功能和组织表达模式。
目前已经对百合花多个种类的基因组进行了测序和比较研究。
这些研究揭示了百合花的基因组大小、染色体数目和结构,为进一步的遗传和表达研究提供了基础。
在百合花的遗传研究中,人们关注的主要是其颜色、花型和芳香特性的遗传机制。
百合花的颜色有红色、粉色、黄色等多个色彩变异。
通过遗传分析,研究人员发现这些颜色变异是由于不同色素合成基因的转录调控和功能变异所导致的。
同时,百合花的花型也呈现出丰富的变异,如单瓣、重瓣和斑点等。
这些花型的变异主要是由于MADS-box基因家族的表达水平和功能变化所致。
此外,百合花丰富的芳香特性也受到基因的调控。
通过遗传和分子生物学实验,研究人员鉴定了与百合花芳香相关的基因,如芳香酶基因和芳香物质合成酶基因。
除了基因组和遗传特性的研究,人们还关注百合花的进化和栽培历史。
通过比较不同种类百合花的基因组序列,可以追溯其进化历史和亲缘关系。
研究证明,百合花起源于早白垩世,随着地理隔离和自然选择的作用,逐渐形成了今天的多样化物种。
此外,人们对百合花的栽培历史也进行了研究。
百合花在人类历史长河中有着悠久的栽培与应用历史,研究人员通过对遗传标记的分析,可以追溯不同品种百合花的起源和遗传背景。
百合花的基因组和遗传特性研究为我们深入了解百合花的进化机制、品种改良和保护自然资源提供了重要的科学依据。
基于对百合花基因组的测序和功能分析,我们可以根据需要进行基因工程和遗传改良,以培育具有特定花色或芳香特性的新品种。
《2024年‘西伯利亚’百合LiCMK基因克隆及功能分析》范文

《‘西伯利亚’百合LiCMK基因克隆及功能分析》篇一范文字体:西伯利亚百合LiCMK基因克隆及功能分析一、引言西伯利亚百合(Lilium sibiricum)是一种广泛分布于我国北方及西伯利亚地区的美丽花卉,因其高雅的花形和香气被人们所喜爱。
近年来,随着分子生物学和基因组学技术的飞速发展,越来越多的研究者开始关注于西伯利亚百合的基因克隆及功能分析。
本文以LiCMK基因为研究对象,详细介绍其克隆过程及功能分析,为后续研究提供基础资料。
二、材料与方法1. 材料本实验所用西伯利亚百合样品采集自某植物园,并经过DNA 提取试剂盒进行基因组DNA的提取。
2. 方法(1)基因克隆首先,通过PCR技术扩增LiCMK基因的cDNA序列。
然后,将PCR产物进行纯化、连接至克隆载体,并转化至大肠杆菌中。
最后,通过蓝白斑筛选和测序验证,获得LiCMK基因的克隆序列。
(2)功能分析采用生物信息学方法,对LiCMK基因的序列进行注释和预测其功能。
同时,通过构建LiCMK基因的过表达和敲除载体,进行转基因植物的培育和表型分析,以探究其生物学功能。
三、结果与分析1. LiCMK基因的克隆结果通过PCR扩增及克隆载体连接转化等步骤,成功获得LiCMK基因的克隆序列。
测序结果显示,该序列与NCBI数据库中的已知序列高度相似,表明成功克隆了LiCMK基因。
2. LiCMK基因的功能分析(1)生物信息学分析通过生物信息学方法对LiCMK基因进行注释,发现其编码一个细胞膜蛋白。
同时,利用多种在线工具预测其功能,发现该基因可能参与植物的生长和发育过程。
(2)转基因植物培育及表型分析构建LiCMK基因的过表达和敲除载体,并转化至拟南芥中。
经过培育和表型分析,发现过表达LiCMK基因的拟南芥植株生长更为茂盛,而敲除LiCMK基因的拟南芥植株则表现出生长迟缓、叶片发黄等表型。
这表明LiCMK基因在西伯利亚百合中具有促进植物生长的重要功能。
四、讨论与展望本实验成功克隆了西伯利亚百合的LiCMK基因,并对其功能进行了初步分析。
百合花发育相关MADS box基因的克隆

百合花发育相关MADS box基因的克隆
张云;刘青林;欧阳青;蔡文启
【期刊名称】《园艺学报》
【年(卷),期】2004(31)3
【摘要】利用RT PCR的方法 ,从百合中克隆得到了 3个接近全长的花发育相关MADSbox基因片段Lf MADS1~ 3。
其中LfMADS1和LfMADS3以往未见报道 ,分别与多种作物的AG、SEP同源基因具有较高的同源性 ;LfMADS2与已发表的LMADS2对应区段的氨基酸同源性高达 98 99% 。
【总页数】5页(P332-336)
【关键词】百合;花发育;MADS;box基因;克隆
【作者】张云;刘青林;欧阳青;蔡文启
【作者单位】中国农业大学观赏园艺与园林系;中国科学院微生物研究所
【正文语种】中文
【中图分类】S682.2
【相关文献】
1.蜻蜓凤梨花发育相关B类MADS-box基因克隆及表达分析 [J], 张俊芳;徐立;李志英
2.无核荔枝花发育相关MADS-box基因的克隆及结构分析 [J], 禤维言;李明芳;郑学勤
3.钙影响花生胚发育相关MADS box基因的克隆与初步鉴定 [J], 张君诚;张新文;
宋育红;刘思衡;庄伟建
4.2个水稻花发育相关MADS-box基因的全序列cDNA克隆及结构分析 [J], 陈锐;高之桢;詹树萱;孙崇荣;曹凯鸣
5.柿花发育相关的MADS-box基因克隆与表达 [J], 丁燕;韩振海;许雪峰;李天忠因版权原因,仅展示原文概要,查看原文内容请购买。
百合分子生物学研究进展

百合分子生物学研究进展徐雷锋摘要百合具有很高的观赏价值,是世界著名的切花之一。
本文从百合DNA分子标记及其初步应用、转基因和百合基因克隆等3方面综述了国内外在百合分子物学方面所取得的研究进展,并对分子生物学在百合应用上存在的问题与前景进行了讨论。
关键词百合DNA分子标记基因克隆转基因分子生物学Advance in Study on Lilium Molecular BiologyAbstract Lilium has high ornamental value, is one of world famous cut-flowers. A review was given in this paper with advance in study on Lilium molecular biology , such as DNA molecular marker ,gene cloning gene engineering. The questions and prospect of molecular biology research on Lilium were also discussed.Key words Lilium DNA molecular marker gene cloning gene engineering molecular biology百合(Lilium)是百合科百合属多年生草本球根植物,是世界重要的鲜切花之一,有2 000 余年栽培历史,具有较高的经济和观赏价值。
加强百合育种具有很高的经济效益和社会效益。
长期以来其品质的改良都是依赖于传统的遗传育种方法。
植物分子生物学的迅速发展,为改良植物品质和性状提供了全新的途径,利用转基因的手段改良百合性状和品质变得切实可行[1]。
目前,在百合分子生物学方面的研究主要包括百合DNA分子标记的研究和应用、利用转基因技术育种及百合相关的基因克隆等。
DHAR基因植物表达双元载体的构建及转化百合研究

DHAR基因植物表达双元载体的构建及转化百合研究张栋;冯凤娟;马锋旺;张燕子【期刊名称】《西北农业学报》【年(卷),期】2009(018)001【摘要】将质粒pSB166中包含ED35s肩动子,Omega元件及TNOS终止子的一段核苷酸序列定向克隆到质粒pCAMBIA1303,构建了中间表达载体,将DHAR 基因与pCSB定向连接,构建了DHAR基因植物表达载体pCSB-DHAR.进而以百合叶片为受体,通过根癌农杆菌介导的方法将DHAR基因转入百合,对转基因百合进行PCR检测,初步表明DHAR基因已整合到百合基因组中.【总页数】5页(P225-229)【作者】张栋;冯凤娟;马锋旺;张燕子【作者单位】西北农林科技大学,园艺学院,陕西杨凌,712100;西北农林科技大学,园艺学院,陕西杨凌,712100;西北农林科技大学,园艺学院,陕西杨凌,712100;西北农林科技大学,园艺学院,陕西杨凌,712100【正文语种】中文【中图分类】Q786;S682.2【相关文献】1.新型HMW-GS基因1Dy12.1~(*t)植物双元表达载体的构建及其遗传转化 [J], 张艳贞;王轲;张静;晏月明2.种子中特异表达IPT基因的植物双元载体构建及转基因水稻获得 [J], 张红心;王桂兰;陈超;赵璞;马春红3.APX基因植物表达双元载体的构建及转化剑麻 [J], 覃海燕;徐洪伟;高建明;鹿志伟;莫廷辉;易克贤4.HA标签的水稻水孔蛋白OsPIP2;7基因植物双元表达载体构建及转基因水稻的获得 [J], 王晓静; 孙林静; 孙玥; 张融雪; 李军玲; 闫双勇; 苏京平5.人α-干扰素2b基因转化番茄、番木瓜的研究(Ⅰ)──—人α-干扰素2b基因植物表达载体构建方法的研究 [J], 周鹏;赵志英;郑学勤因版权原因,仅展示原文概要,查看原文内容请购买。
百合MDHAR基因的克隆与表达分析

s a e i h y h mo o y wi t e l n s h r d h g l o l g t o h rp a t .RT— h PCR a a y i s o d t a x e so f l DHAR g n o l e d t c e n n l ss h we h te pr s i n o M L e ec ud b ee td i
重要 的 自由基 清 除剂 , 通 过参 与 一 系列 的氧 化还 是
原 反 应 而 发 挥 抗 氧 化 剂 的 功 能 。 单 脱 氢 抗 坏 血 酸 还 原 酶 ( o o ey rac ra eu t e MD A ) 抗 m n dh d0 sobt rd ca , H R 在 e s
第4 业
科
学
Vo . 146. . No 9
S p., e 20 10
SCI ENTI A
S LVAE I
S NI I CAE
百 合 MD A H R基 因 的 克 隆 与 表 达 分 析
陈 莉 辛 海 波 李 晓艳 李 晓 昕 义 鸣放
坏 血酸 一谷胱 甘肽 循 环 中 , 以将 氧 化 型抗 坏 血酸 可 ( D A) M H 还原 为还 原 型 抗 坏 血 酸 ( s , A A) 在维 持植 物体 内抗 坏血 酸 的正常代 谢和植 物体 氧化还原 平衡 方 面起着 重要 作用 ( h ne a.2 0 ; 0 5 。 C e t 1 0 3 2 0 ) , 百合 ( iu 作 为 主要 的切 花 和盆 花 材 料 , Ll m) i 在 国内外 花卉市 场上 占有重 要地位 。百合 性喜冷 凉湿 润气 候 , 越夏 的百 合 经常 出现 生 长停 滞 、 植株 低 矮 、 花 朵败 育等现 象 , 重影 响 了切 花质量 , 严 并造成 百合 种球 退化 。夏 季高 温 已成 为 限制 国内百 合周 年生产
百合花香合成相关基因LiMCS 的克隆、定位和表达特性研究

西北农业学报 2022,31(8):981-989A c t aA gr i c u l t u r a eB o r e a l i -o c c i d e n t a l i sS i n i c a 网络出版日期:2022-04-08 d o i :10.7606/j.i s s n .1004-1389.2022.08.005网络出版地址:h t t ps ://k n s .c n k i .n e t /k c m s /d e t a i l /61.1220.S .20220407.1656.002.h t m l 百合花香合成相关基因L i M C S 的克隆、定位和表达特性研究收稿日期:2021-08-20 修回日期:2021-10-15基金项目:国家重点研发计划(2018Y F D 1000400);国家自然科学基金(31201645);北京市自然科学基金(6172006)㊂第一作者:张 茜,女,硕士研究生,研究方向为百合花香分子调节机制㊂E -m a i l :z h a n gx i 0522@126.c o m 通信作者:胡增辉,男,教授,研究方向为植物生理生态㊂E -m a i l :b u a h u z e n gh u i @163.c o m 张 茜1,2,罗景琳2,王浩楠1,2,李迎迎1,2,冷平生1,2,胡增辉1,2(1.北京林木分子设计育种高精尖创新中心,北京 102206;2.北京农学院园林学院,北京 102206)摘 要 百合花香中含有大量的萜类化合物,2-C -甲基-D -赤藻糖醇-2,4-环二磷酸合成酶(M C S )是合成单萜的甲基赤藓糖醇磷酸(M E P )途径中的关键酶之一㊂为了进一步探究M C S 在百合花香合成中的作用,克隆了 西伯利亚 百合L i M C S 基因,进行生物信息学分析,使用亚细胞定位确定该基因编码蛋白的位置,根据荧光定量P C R 确定了该基因的时空表达模式㊂结果表明,L i M C S 基因开放阅读框为669b p,编码222个氨基酸,与盾叶薯蓣的相似度最高㊂L i M C S 蛋白定位于烟草叶表皮细胞的叶绿体中㊂L i M C S 基因在 西伯利亚 百合花的半开期表达量最高,花蕾期最低;在盛开期,该基因在花瓣中表达量最高,在其他部位表达量较低㊂结果证明L i M C S 可能参与百合花香单萜类物质合成,这为后续利用基因工程手段调控百合花香释放奠定了理论基础㊂关键词 西伯利亚 百合;L i M C S ;基因克隆;表达分析中图分类号 S 682 文献标志码 A 文章编号 1004-1389(2022)08-0981-09释放花香是开花植物的重要特性㊂花香不仅在促进植物传粉和抵御外来侵害[1-2]等方面发挥着重要作用,而且作为植物重要的观赏性状之一,是评价植物观赏价值和经济价值的重要指标㊂花香由一系列低分子量次生代谢挥发物组成,主要包括萜烯类㊁脂肪族类㊁苯环类/苯丙烷类及一些含硫㊁含氮类等化合物[3-4]㊂深入了解花香合成调控机制能为开展植物花香育种及揭示花香生理生态功能提供理论支撑㊂百合是世界著名的五大切花之一,因其气味芳香浓郁而具有极高的观赏和经济价值㊂百合花香的主要成分为萜烯类化合物,包括芳樟醇㊁罗勒烯和月桂烯等[5-6]㊂笔者的前期研究发现,不同品系百合的花香也存在明显差别[7],如浓香型的东方百合香气过于浓郁,而淡香型的亚洲百合的香味很弱,因此培育芳香适宜的百合新品种成为新的育种趋势㊂研究表明,萜烯合成途径活化水平不同是导致东方百合(浓香型)和亚洲百合(淡香型)香味差异的一个重要原因[8]㊂但目前对于百合萜烯合成调控机制尚未清楚解析㊂萜类化合物是由异戊二烯(C 5)基本单元组成,C 5单元的合成前体物质为异戊二烯焦磷酸(i s o p e n t e n y l p y r o p h o s ph a t e ,I P P )㊂I P P 的合成主要包括两条途径,分别是在细胞质中以倍半萜合成为主的甲羟戊酸(MV A )途径和在质体中以单萜合成为主的甲基赤藓糖醇磷酸(M E P )途径[9]㊂由于单萜类化合物是百合花香的主要成分,因此对M E P 途径关键酶基因的功能解析成为实现百合花香合成调控的前提㊂近年来,对于M E P 途径关键酶基因的研究越来越多,在金鱼草(A n t i r r h i n u m m a j u s )[10]㊁百合㊁薰衣草(L a v a n -d u l aa n g u s t i fo l i a )[11]等植物中多个关键基因被克隆出来,并且部分基因也已被证明在单萜合成中起关键作用㊂在百合中,L i T P S ㊁L i D X S ㊁L i D X R ㊁L i M C T 等M E P 途径基因已被克隆[12-14],表达模式和功能也逐渐被解析,但尚有多个基因未被鉴定㊂在M E P 途径中,2-C -甲基赤藓糖醇-2,4-环焦磷酸合成酶(M C S )催化二磷酸胞苷-2-C -甲基-D -赤藻糖醇-2-磷酸(C D P -M E P )生成2-C -甲基-D -赤藻糖醇-2,4-环二磷酸(M E P -c P P ),是合成单萜的关键步骤㊂该基因已在长春花(C a t h a r a n t h u s r o s e u s )[15]㊁盾叶薯蓣(D i -Copyright ©博看网. All Rights Reserved.o s c o r e a z i n g i b e r e n s i s)[16]㊁青蒿(A r t e m i s i a a n-n u a)[17]等植物中相继被克隆出来,且在长春花㊁盾叶薯蓣和青蒿中都已被证明其对萜类物质合成具有重要作用㊂但M C S基因在百合中尚未被克隆,对其表达模式和功能还不清楚㊂本研究选用东方百合 西伯利亚 为材料,克隆L i M C S基因,通过生物信息学㊁荧光定量P C R (q R T-P C R)及亚细胞定位等,预测其特征及功能,并明确其表达模式,为揭示L i M C S基因在 西伯利亚 百合单萜类物质合成中的作用和功能奠定基础㊂1材料与方法1.1试验材料本试验选用的植物材料为 西伯利亚 百合(L i l i u m S i b e r i a )实生苗,种球购自北京荷景良苑贸易有限公司,种植于北京农学院东大地(40ʎ5'24ᵡN,116ʎ17'55ᵡE)单坡面温室中㊂试验选取3株无病虫害㊁生长健壮㊁均高100c m的植株,在不同的花期及在盛花期的不同器官和组织进行采样㊂采样分为花蕾期㊁半开期㊁盛开期和衰败期4个时期,在盛花期采集根㊁茎㊁叶㊁外花被片㊁内花被片㊁花药㊁子房㊁花柱㊁花丝的样品,每组样品设置3个重复㊂置于液氮中速冻,-80ħ保存㊂1.2 西伯利亚 百合总R N A的提取及第一条链的合成将样品在灭菌及用液氮遇冷过的研钵中进行研磨,直至样品研磨成粉末状㊂选用T r a n s Z o l U PP l u sR N A K i t试剂盒(T r a n s G e nB i o t e c h,中国北京)提取 西伯利亚 百合的总R N A㊂用于基因克隆与q R T-P C R的c D N A第一条链合成分别选用T r a n s S c r i p t O n e-S t e p g D N A R e m o v a l a n d c D N AS y n t h e s i s S u p e rM i x(T r a n s-G e n B i o t e c h,中国北京)与E v o M-M L V R T P r e m i x f o r q P C R试剂盒(A c c u r a t eB i o l o g y,中国湖南),按照说明书进行,反应完成后将其置于-20ħ冰箱中保存㊂1.3L i M C S基因的克隆及验证基于前期测得 西伯利亚 百合转录组数据[6],从中调取M C S基因,通过S n a p G e n e设计全长扩增引物(表1)㊂P C R扩增使用制备好的c D N A为模板进行,P C R反应体系为:上下游引物各1μL,c D N A模板1μL,2ˑA8F a s t H i F i P C R M a s t e rM i x12.5μL,d d H2O9.5μL,共25μL的体系㊂P C R反应条件为:95ħ预变性3 m i n,95ħ变性10s,55ħ退火15s,72ħ延伸20s,进行35个循环,最后72ħ延伸5m i n㊂将扩增产物利用琼脂糖凝胶电泳实验进行验证㊂随后使用50μL体系进行P C R扩增,将产物使用E a s y P u r e Q u i c k G e lE x t r a c t i o n K i t(T r a n s G e n B i o t e c h,中国北京)进行回收纯化,纯化后的D N A片段置于-20ħ冰箱中保存,备用㊂将得到的目的片段连接至P c l o n e007载体上,转化至大肠杆菌D H5α(E s c h e r i c h i ac o l i)菌株,挑取阳性克隆,扩大培养后,用菌液P C R的方式进行初步验证,选取符合要求的菌液送至华大基因公司进行测序㊂1.4L i M C S生信分析使用生物信息学技术对 西伯利亚 百合L i M C S相关生物信息进行分析预测(表2)㊂1.5L i M C S的亚细胞定位将L i M C S终止子去除后,使用N i m b l eC l o-n i n g[18]的通用接头序列设计引物G-L i M C S-F㊁G-L i M C S-R(表1),以L i M C S质粒为模板以50μL体系进行P C R扩增,同时使用2ˑS e a m L e s s M i x酶切载体p N C-G r e e n-S u b N,将两个产物分表1 西伯利亚 百合L i M C S基因克隆㊁亚细胞定位和表达分析引物序列T a b l e1P r i m e r s f o r g e n e c l o n i n g,s u b c e l l u l a r l o c a l i z a t i o na n d e x p r e s s i o na n a l y s i s o f L i M C S引物名称P r i m e r n a m e引物序列(5'ң3')P r i m e r s e q u e n c e(5'ң3') L i M C S-F A T G G C T T C T T C T C T C T C C T TL i M C S-R T C A C T T C C T C A T C A G A A G A A C A A CG-L i M C S-F A G T G G T C T C T G T C C A G T C C T A T G G C T T C T T C T C T C T C C T TG-L i M C S-R G G T C T C A G C A G A C C A C A A G T C T T C C T C A T C A G A A G A A C A A CL i M C S-Q-F G T T G A T C C T C C A G A A G C C G A A GL i M C S-Q-R A G T G T G A G C A G C G A T A C T A C G Aβ-A c t i n-F C G G T G T C T G G A T T G G A G G G T C Aβ-A c t i n-R C T T C C T G T G G A C G A T G G C T G G A㊃289㊃西北农业学报31卷Copyright©博看网. All Rights Reserved.表2 西伯利亚 百合L i M C S基因生物信息预测软件及网站T a b l e2S o f t w a r e a n dw e b s i t e f o r p r e d i c t i n g t h e b i o l o g i c a l i n f o r m a t i o no f L i M C S生物信息B i o l o g i c a l i n f o r m a t i o n相关软件及网站R e l a t e d s o f t w a r e a n dw e b s i t e开放阅读框O p e n r e a d i n g f r a m e h t t p://w w w.n c b i.n l m.n i h.g o v/O r f f i n d e r蛋白序列比对P r o t e i n s e q u e n c e a l i g n m e n t h t t p s://b l a s t.n c b i.n l m.n i h.g o v/B l a s t.c g i和D N AMA N 进化树构建E v o l u t i o n a r y t r e e c o n s t r u c t i o n M E G A7.0理化性质预测P r e d i c t i o no f p h y s i c a l a n d c h e m i c a l p r o p e r t i e s h t t p s://w e b.e x p a s y.o r g/p r o t p a r a m二级结构预测P r e d i c t i o no f s e c o n d a r y s t r u c t u r e N P S A-P R A B I三级结构预测T h e p r e d i c t i o no f t e r t i a r y s t r u c t u r e h t t p s://s w i s s m o d e l.e x p a s y.o r g信号肽预测P r o t e i n s i g n a l p e p t i d e p r e d i c t i o n h t t p s://w w w.d e t a i b i i o.c o m/t o o l s/s i g n a l-p e p t i d e.h t l 跨膜信息预测T r a n s m e m b r a n e p r e d i c t i o n h t t p s://w w w.d e t a i b i i o.c o m/t o o l s/t r a n s m e m b r a n e.h t m l 磷酸化位点预测P r e d i c t i o no f p r o t e i n p h o s p h o r y l a t i o n s i t e s N e t P h o s3.1S e r v e r别回收纯化后进行同源克隆,构建p N C-G r e e n-S u b N-L i M C S载体,送华大基因测序㊂测序结果比对成功后将其转入农杆菌感受态G V3101中,使用注射法侵染烟草叶片,在激光共聚焦显微镜下进行观察㊂1.6L i M C S的时空表达分析根据克隆得到的L i M C S基因,利用S n a p-G e n e设计该基因的荧光扩增引物(表1)㊂以90~100n g用于q R T-P C R的c D N A作为反应模板配制反应溶液,再加入上下游引物各0.4μL,2ˑS Y B RG r e e nP r o T a q H SP r e m i x10μL,最后使用R N a s e f r e ew a t e r定容至至20μL,随后将配制好并混匀的反应溶液置于荧光P C R仪(B i o-R a d-i Q5)中进行q R T-P C R分析㊂将得到的数据依据相对表达量公式2-ΔΔC t并使用i Q5㊁M i-c r o s o f tE x c e l和S P S S等软件对L i M C S基因在 西伯利亚 百合中时空表达的试验结果进行计算分析㊂2结果与分析2.1 西伯利亚 百合L i M C S的克隆从全长L i M C S基因的电泳图(图1)可以看出,克隆的片段为500~750b p,随后将其产物回收纯化,与P c l o n e007载体链接并转化到D H5α上,经菌落P C R验证后送往华大基因公司进行测序,将结果与转录组中的基因进行序列比对,以此确定L i M C S的最终序列㊂2.2L i M C S核苷酸和氨基酸序列分析2.2.1L i M C S核苷酸序列及其编码蛋白保守结构分析使用N C B I网站中的O p e n R e a d i n g F r a m eF i n d e r功能,确定L i M C S最大的开放阅读框为669b p,利用D N AMA N软件将其翻译成氨基酸序列(图2)㊂L i M C S蛋白是由222个氨基酸构成,属于2-C-甲基-D-赤藻糖醇-2,4-环二磷酸合酶(2-C-m e t h y l-D-e r y t h r i t o l-2,4-c y c l o-d i p h o s p h a t e s y n t h a s e(M E C D P))超家族(图3)㊂M.D L2000D N A M a r k e r;1.L i M C S基因全长电泳M.D L2000D N A M a r k e r;1.F u l l l e n g t ho f L i M C S g e n e图1 西伯利亚 百合L i M C S基因全长电泳扩增F i g.1A g a r o s e g e l f o r a m p l i f i c a t i o no ff u l l l e ng t ho f L i M C S g e n e2.2.2 L i M C S氨基酸序列同源性比对及系统进化树构建分析使用N C B I中的B l a s t和D N A-MA N软件将L i M C S与其他植物M C S氨基酸序列的同源性进行比对,发现其与盾叶薯蓣D i M C S 的相似度极高,与芝麻(S e s a m u m i n d i c u m) S e M C S㊁海枣(P h o e n i x d a c t y l i f e r a)P h M C S㊁毛果杨(P o p u l u s t r i c h o c a r p a)P o M C S㊁椰子(C o c o s n u c i f e r a)C o M C S㊁油棕(E l a e i s g u i n e e n s i s)E l-a M C S㊁香蕉(M u s a a c u m i n a t a)M u M C S㊁荸荠(E l e o c h a r i sd u l c i s)E l e M C S等植物M C S蛋白的相似度较高(图4)㊂为了进一步研究 西伯利亚 百合L i M C S与其他植物的M C S同源基因的系㊃389㊃8期张茜等:百合花香合成相关基因L i M C S的克隆㊁定位和表达特性研究Copyright©博看网. All Rights Reserved.统进化关系,使用M E G A 7.0软件构建L i M C S 与其他植物的M C S 氨基酸序列的进化树(图5),发现其与盾叶薯蓣同源性最高并形成一个小亚群,和荸荠㊁香蕉㊁海枣㊁椰子和油棕等植物的亲缘性较近㊂图2 西伯利亚 百合L i M C S 基因核苷酸序列以及其氨基酸序列F i g .2 T h e n u c l e o t i d e a n da m i n o a c i d s e qu e n c e o f L i M C S g e ne 图3 西伯利亚 百合L i M C S 蛋白保守结构F i g.3 T h e c o n s e r v e dd o m a i n s o fL i M C S 2.2.3 西伯利亚 百合L i M C S 蛋白理化性质利用E x P A S y -P r o t P a r a m 在线工具对L i M C S 蛋白的理化性质进行预测,该基因共编码222个氨基酸,其中含量最高的为亮氨酸(L e u ),占13.5%;其次是丙氨酸(A l a )㊁脯氨酸(P r o)和丝氨酸(S e r ),分别为11.3%㊁11.3%和9.0%,其正电残基数(A s p +G l u )与负点残基数(A r g +L y s )分别为21和23,为亲水性不稳定蛋白(表3)㊂2.2.4 L i M C S 二级和三级结构分析 利用N P S A -P R A B I 在线软件对L i M C S 蛋白二级结构进行预测,发现其由70个α-螺旋(a l ph ah e l i x ,H h )㊁28个延伸链(e x t e n d e ds t r a n d ,E e )㊁9个β-转角(b e t a t u r n ,T t )及115个无规卷曲(r a n d o mc o i l ,C c)组成(图6)㊂由二级结构卷曲折叠形成L i M C S 的三级结构模型,以2p m p .1.A (拟南芥异戊二烯生物合成途径中2-C -甲基-D -赤藓糖醇2,4-环二磷酸合酶的结构)为模板,二者相似度高达89.31%,相似度大于60%被认为非常准确,其中序列覆盖率为72%,C -β相互作用值为2.54(图7)㊂2.2.5 L i M C S 蛋白信号肽和跨膜结构预测分析利用信号肽预测网站对L i M C S 进行蛋白质信号肽的预测,发现该蛋白不含信号肽(图8)㊂随后,又使用跨膜信息预测网站预测该蛋白的跨膜结构,发现该蛋白不含跨膜结构(图9)㊂由以上结果推断,该蛋白不属于信号蛋白㊂㊃489㊃西 北 农 业 学 报31卷Copyright ©博看网. All Rights Reserved.黑色.同源性=100%;粉色.同源性ȡ75%;蓝色.同源性ȡ50%;D i M C S .A S L 70633.1;S e M C S .X P _011073285.1;P h M C S .X P _008805558.1;P o M C S .X P _002304519.1;C o M C S .K A G 1362373.1;E l a M C S .X P _010933967.1;M u M C S .X P _009397837.1;E l e M C S .QMU 23681.1B l a c k .T h eh o m o l o g y =100%;P i n k .T h eh o m o l o g y ȡ75%;B l u e .T h eh o m o l o g y ȡ50%;D i MC S .A S L 70633.1;S e M C S .X P _011073285.1;P h M C S .X P _008805558.1;P o M C S .X P _002304519.1;C o M C S .K A G 1362373.1;E l a M C S .X P _010933967.1;M u M C S .X P _009397837.1;E l e M C S .QMU 23681.1图4 L i M C S 基因编码的氨基酸序列与其他植物同源性比对F i g .4 T h e h o m o l o g y c o m p a r i s o nb e t w e e na m i n o a c i d s e q u e n c e s e n c o d e db y Li M C S g e n e a n dM C S i no t h e r p l a n ts 图5 西伯利亚 百合L i M C S 基因编码氨基酸序列的系统进化树F i g .5 P h y l o g e n e t i c t r e e o f a m i n o a c i d s e q u e n c e e n c o d e db y Li M C S g e n e 表3 西伯利亚 百合L i M C S 蛋白理化性质分析T a b l e 3 P h y s i c a l a n d c h e m i c a l p r o pe r t i e s o fL i M C S p r o t e i n 蛋白名称P r o t e i n n a m e 分子质量/uM o l e c u l a r m a s s 总原子数T o t a l n u m b e r o f a t o m s分子式F o r m u l a理论等电点T h e o r e t i c a l pI 脂肪系数A l i p h a t i c i n d e x 不稳定指数I n s t a b i l i t y i n d e x 亲水系数G R A V Y L i M C S23636.423365C 1052H 1703N 303O 299S 88.5100.2753.19-0.048㊃589㊃8期张 茜等:百合花香合成相关基因L i M C S 的克隆㊁定位和表达特性研究Copyright ©博看网. All Rights Reserved.蓝色为α-螺旋;红色为延伸链;绿色为β-转角;紫色为无规卷曲T h eb l u e i s α-h e l i x ;R e d i s e x t e n d e d s t r a n d ;G r e e n i s b e t a t u r n ;P u r pl e i s r a n d o mc o i l 图6 西伯利亚 百合L i M C S 蛋白质二级结构的预测F i g .6 P r e d i c t i o no f s e c o n d a r ys t r u c t u r e o fL i M CS 图7 西伯利亚 百合L i M C S 蛋白质三级结构的预测F i g .7 T h e p r e d i c t i o no f t e r t i a r y st r u c t u r e o fL i M CS 图8 西伯利亚 百合L i M C S 蛋白质信号肽预测F i g .8 P r o t e i n s i g n a l p e pt i d e p r e d i c t i o no fL i M CS 图9 西伯利亚 百合L i M C S 蛋白质跨膜螺旋预测F i g.9 T r a n s m e m b r a n e p r e d i c t i o no fL i M C S 2.2.6 L i M C S 蛋白磷酸化位点预测分析 通过在线软件N e t P h o s 3.1S e r v e r 对 西伯利亚 百合L i M C S 蛋白磷酸化位点进行预测,发现该蛋白可能含有20个丝氨酸㊁4个苏氨酸和1个酪氨酸(图10)㊂图10 西伯利亚 百合L i M C S 蛋白磷酸化位点预测F i g .10 P r e d i c t i o no f p r o t e i n p h o s p h o r yl a t i o n s i t e s o fL i M C S 2.3 L i M C S 亚细胞定位分析图11显示L i M C S 亚细胞定位的结果,通过激光共聚焦显微镜观察发现融合蛋白与叶绿体荧光场重合,说明其蛋白特异性定位于叶绿体中㊂2.4 L i M C S 时空表达模式分析L i M C S 在百合中的时空表达模式显示(图12),L i M C S 的表达量随着百合花的发育整体呈现先升高后降低的规律㊂在半开期L i M C S 基因的表达量达到顶峰,其次为盛开期与衰败期,花蕾期表达量最低,仅为半开期的1/3㊂西伯利亚 百合L i M C S 基因在不同器官和组织中的表达量也不相同(图13)㊂L i M C S 基因在花器官中的表达总和远远大于根㊁茎㊁叶,占总量的4/5,而花瓣中的表达量占总量的1/2㊂L i M C S 在外瓣中表达量最高,内瓣其次,叶中表达量仅次于花瓣㊂3 讨论单萜类化合物是西伯利亚 百合花香的主要㊃689㊃西 北 农 业 学 报31卷Copyright ©博看网. All Rights Reserved.从左到右依次为目标蛋白荧光通道;明场;叶绿体荧光通道;叠加场T h e r e a r e t h e t a r g e t p r o t e i n f l u o r e s c e n c e c h a n n e l ;B r i g h t f i e l d ;C h l o r o p l a s t f l u o r e s c e n c e c h a n n e l a n dm e r g e d f r o ml e f t t o r i gh t 图11 西伯利亚 百合L i M C S 亚细胞定位F i g.11 S u b c e l l u l a r l o c a l i z a t i o no fL i M CS 不同小写字母表示差异显著(P <0.05)D i f f e r e n t l o w e r c a s e l e t t e r s i n d i c a t e s i gn i f i c a n t d i f f e r e n c e s (P <0.05)图12 西伯利亚 百合L i M C S 在不同花期的表达分析F i g .12 E x p r e s s i o na n a l y s i s a t d i f f e r e n t f l o w e r i n g s t a ge s of L i M CS 不同小写字母表示差异显著(P <0.05)D i f f e r e n t l o w e r c a s e l e t t e r s i n d i c a t e s i gn i f i c a n t d i f f e r e n c e s (P <0.05)图13 西伯利亚 百合盛开期L i M C S 在不同器官和组织的表达分析F i g .13 E x p r e s s i o na n a l y s i s i nd i f f e r e n t p l a n t o r g a n s a n d t i s s u e s o f L i M C S i n t h e f l o w e r i n g s t a ge 组成物质㊂M C S 是植物单萜物质合成途径中催化第五步反应所需的酶,该途径的产物可以催化生成单萜类化合物的合成前体物质I P P ㊂因此,M C S 基因的克隆及其功能的挖掘,能为 西伯利亚 百合花香单萜类挥发物的合成及调控机制的研究提供依据㊂通过对一系列结构基因的克隆,不断深入花香调节机制的研究,从而实现花香的人为调控及后续的育种工作㊂本研究克隆出了西伯利亚 百合中的L i M C S 基因,并通过生物信息学技术对其编码蛋白的理化性质㊁结构功能㊃789㊃8期张 茜等:百合花香合成相关基因L i M C S 的克隆㊁定位和表达特性研究Copyright ©博看网. All Rights Reserved.进行了预测和分析,同时明确了其表达位置及表达模式㊂通过氨基酸序列比对分析,得知L i M C S与盾叶薯蓣d z M C S具有较高的同源性㊂在盾叶薯蓣中,通过构建d z M C S基因的大肠杆菌表达载体,最终加强了大肠杆菌的M E P途径,尤其促进了胡萝卜素的合成与积累[16]㊂同时,L i M C S具有保守结构域,表明该基因编码的蛋白是具有生物功能的蛋白,为L i M C S基因的功能研究提供了理论依据㊂L i M C S基因编码的蛋白质定位于烟草叶片表皮细胞的叶绿体中,这与单萜在质体中经M E P途径合成相一致㊂ 西伯利亚 百合花香由花蕾期开始积累,在盛开期达到顶峰[19],而L i M C S基因主要在半开期表达量最高,推测该基因作为M E P途径上游基因,在半开期进行了大量的转录积累,逐步合成下游单萜产物,最终在盛花期花香得到大量释放㊂该基因主要在花瓣中发挥作用,这与 西伯利亚 百合M E P途径中上游L i D X S基因㊁L i D X R基因和下游单萜合酶L i T P S基因的表达模式基本吻合[12-13]㊂目前, M C S基因在部分植物中已有研究㊂长春花M C S 基因的过表达,增加了单萜吲哚生物碱的含量,这表明增加M C S基因的表达将有助于代谢向下游流动[15]㊂将青蒿A a M C S基因在拟南芥中超表达,类胡萝卜素㊁叶绿素a和叶绿素b含量显著增加[17]㊂这些研究都表明M C S基因在萜烯类物质合成中起重要作用㊂L i M C S基因的克隆不仅扩充了百合的基因库,为今后花香调控相关的基因工程研究提供目的基因,还为萜烯类合成释放分子机制的深入探究奠定了基础㊂但本研究仅阐述了该基因相关的理化性质和表达模式,缺乏关于L i M C S基因调节 西伯利亚 百合花香合成机制的研究,后续将会使用病毒诱导的基因沉默(V I G S)㊁瞬时过表达㊁启动子克隆等分子生物学技术对L i M C S基因的功能展开更深入的探究,最终明确L i M C S 基因在 西伯利亚 百合花香合成中的作用机制㊂参考文献R e f e r e n c e:[1] MA J E T I CCJ,R A G U S O R A,A S HMA N T L.T h e s w e e ts m e l l o f s u c c e s s:f l o r a l s c e n t a f f e c t s p o l l i n a t o r a t t r a c t i o n a n ds e e d f i t n e s s i n H e s p e r i sm a t r o n a l i s[J].F u n c t i o n a lE c o l o-g y,2009,23(3):480-487.[2] K A T T ET,T A N K H,S UZ HH,e t a l.F l o r a l f r a g r a n c e s i nt w o c l o s e l y r e l a t e df r u i t f l y o r c h i d s,B u l b o p h y l l u m h o r t o-r u m a n d B.m a c r a n t h o i d e s(O r c h i d a c e a e):a s s o r t m e n t so fp h e n y l b u t a n o i d s t o a t t r a c t t e p h r i t i d f r u i t f l y m a l e s[J].A p-p l i e dE n t o m o l o g y a n dZ o o l o g y,2020,55(1):55-64.[3] D U D A R E V A N,P I C H E R S K Y E.B i o c h e m i c a l a n dm o l e c u-l a r g e n e t i c a s p e c t so f f l o r a ls c e n t s[J].P l a n tP h y s i o l o g y, 2000,122(3):627-633.[4] K AM I N A G A Y,S C H N E P PJ,P E E LG,e t a l.P l a n t p h e n y-l a c e t a l d e h y d e s y n t h a s e i s a b i f u n c t i o n a l h o m o t e t r a m e r i c e n-z y m e t h a t c a t a l y z e s p h e n y l a l a n i n ed e c a r b o x y l a t i o na n do x i-d a t i o n[J].T h eJ o u r n a l o f B i o l o g i c a l C he m i s t r y,2006,281(33):23357-23366.[5]李天娇,胡增辉,郑健,等. 西伯利亚 百合单萜释放与L i-m T P S表达日变化动态分析[J].西北农业学报,2016, 25(5):763-769.L IT J,HU Z H,Z H E N G J,e ta l.D i u r n a lc h a n g e so f m o n o t e r p e n e s e m i s s i o n a n d L i-m T P S e x p r e s s i o n i n L i l i u ms i b e r i a [J].A c t aA g r i c u l t u r a eB o r e a l i-O c c i d e n t a l i s S i n i-c a,2016,25(5):763-769.[6]唐彪,胡增辉,冷平生.不同花期 西伯利亚 百合花瓣单萜合成途径转录组分析[J].植物科学学报,2018,36(2): 252-263.T A N GB,HUZH,L E N GPS H.T r a n s c r i p t o m e a n a l y s i s o f t h em o n o t e r p e n eb i o s y n t h e s i s p a t h w a y i n p e t a l so f L i l i u mS i b e r i a a td i f f e r e n tf l o w e r i n g s t a g e s[J].P l a n tS c i e n c e J o u r n a l,2018,36(2):252-263.[7]张辉秀,胡增辉,冷平生,等.不同品种百合花挥发性成分定性与定量分析[J].中国农业科学,2013,46(4):790-799.Z H A N G H X,HUZH,L E N GPS H,e t a l.Q u a l i t a t i v e a n d q u a n t i t a t i v e a n a l y s i s o f f l o r a l v o l a t i l e c o m p o n e n t s f r o md i f-f e r e n tv a r i e t i e so f L i l i u m s p p.[J].S c i e n t i a Ag r i c u l t u r aS i n i c a,2013,46(4):790-799.[8]唐彪,胡增辉,冷平生,等.浓香型和淡香型百合单萜合酶基因差异表达[J].北京农学院学报,2016(2):88-94.T A N GB,HU Z H,L E N G PS H,e ta l.T h ee x p r e s s i o no f m o n o t e r p e n es y n t h a s e g e n e si n L i l i u m w i t h s t r o n g a n d l i g h t f l o r a l f r a g r a n c e[J].J o u r n a l o f B e i j i n g A g r i c u l t u r a lC o l l e g e,2016(2):88-94.[9] D U D A R E V A N,N E G R E F.P r a c t i c a la p p l i c a t i o n so fr e-s e a r c hi n t ot h er e g u l a t i o no f p l a n tv o l a t i l ee m i s s i o n[J].C u r r e n tO p i n i o n i nP l a n tB i o l o g y,2005,8(1):113-118.[10] D U D A R E V A N,MA R T I ND,K I S H C M,e t a l.(E)-b a t e-o c i m e n e a n d m y r c e n es y n t h a s e g e n e so f f l o r a ls c e n tb i o-s y n t h e s i s i ns n a p d r a g o n:f u n c t i o na n de x p r e s s i o no f t h r e et e r p e n e s y n t h a s e g e n e s o f a n e w t e r p e n e s y n t h a s e s u b f a m i-l y[J].P l a n tC e l l,2003,15(5):1227-1241.[11]S A R K E RL,A D A LA M,L I A N GP,e t a l.R N A-s e q i n t h ed i s c o ve r y of a s p a r s e l y e x p r e s s e d s c e n t-d e t e r m i n i n gm o n o t e r p e n e s y n t h a s e i n l a v e n d e r(l a v a n d u l a)[J].P l a n-t a,2018,249:271-291.[12] Z HA N G T X,S U N M,L IL L,e ta l.M o l e c u l a rc l o n i n ga n de x p r e s s i o na n a l y s i so fa m o n o t e r p e n es y n t h a s e g e n ei n v o l v e d i n f l o r a l s c e n t p r o d u c t i o n i n l i l y(L i l i u m S i b e r i-a )[J].R u s s i a nJ o u r n a lo f P l a n tP h y s i o l o g y,2017,64(4):600-607.[13] Z HA N G T X,S U N M,G U O Y H,e ta l.O v e r e x p r e s s i o no f L i D X S a n d L i D X R f r o m L i l y(L i l i u m S i b e r i a )e n-h a n c e s t h e t e r p e n o i d c o n t e n t i n t o b a c c o f l o w e r s[J].F r o n-t i e r s i nP l a n t S c i e n c e,2018(9):909.[14]马波,杨珺,李迎迎,等. 西伯利亚 百合L i M C T基因的克隆及表达特性分析[J].分子植物育种,2020,18(18): 5933-5942.MAB,Y A N G J,L IY Y,e ta l.C l o n i n g a n de x p r e s s i o nc h a r a c t e r i s t i c a n a l y s i s o f L i M C T g e n e i n L i l i u m S i b e r i a[J].M o l e c u l a rP l a n tB r e e d i n g,2020,18(18):5933-5942.[15] B U R L A T V,O U D I N A,C O U R T O I SM,e t a l.C o-e x p r e s-s i o no f t h r e eM E P p a t h w a yg e n e s a n d g e r a n i o l10-h y d r o x-㊃889㊃西北农业学报31卷Copyright©博看网. All Rights Reserved.y l as e i n i n t e r n a l p h l o e m p a r e n c h y m ao f C a t h a r a n t h u s r o -s e u s i m p l i c a t e sm u l t i c e l l u l a r t r a n s l o c a t i o no f i n t e r m e d i a t e s d u r i n g t h eb i o s y n t h e s i so f m o n o t e r pe n ei n d o l ea l k a l o i d s a n d i s o pr e n o i d -d e r i v e d [J ].T h eP l a n tJ o u r n a l ,2004,38:131-141.[16] 王润发.盾叶薯蓣D X R 基因和MD S 基因的克隆与功能验证[D ].湖北武昌:湖北大学,2014.WA N G R F .C l o n i n g a n dc h a r a c t e r i z a t i o no f g e n e se n c o -d i n g D X R g e n e a n d MD S g e n e f r o m D i o s c o r e a z i n gi b e r e n -s i s C .H W r i g h t [D ].W u c h a n g H u b e i :H u b e iU n i v e r s i t y,2014.[17] 张 曼.青蒿A a C MK ㊁A a M C T ㊁A a M C S 基因的克隆与功能分析[D ].重庆:西南大学,2016.Z HA N G M.M o l e c u l a r c l o n i n g an dc h a r a c t e r i z a t i o no f t h e 2-C -m e t h y l -D -e r y t h r i t o l 4-p h o s p h a t e c y t i d y l yl t r a n s f e r a s e g e n e ,4-(c y t i d i n e5'-d i p h o s p h o )-2-C -m e t h y l e r y-t h r i t o lk i -n a s e g e n e ,2-C -m e t h y l e r y t h r i t o l -2,4-c y c l o d i p h o s p h a t e s yn -t h a s e g e n ef r o m A r t e m i s i a a n n u a L [D ].C h o n g q i n g:S o u t h w e s tU n i v e r s i t y,2016.[18] Y A NP ,Z H E N G Y ,S H E N W ,e t a l .N i m b l e c l o n i n g:a s i m -p l e ,v e r s a t i l e ,a n d e f f i c i e n t s y s t e m f o r s t a n d a r d i z e dm o l e c u l a r c l o -n i n g [J ].F r o n tB i o e n g B i o t e c h n o l o g y ,2020,7:460.[19] 张辉秀,冷平生,胡增辉,等. 西伯利亚 百合花香随开花进程变化及日变化规律[J ].园艺学报,2013,40(4):693-702.Z HA N G H X ,L E N G P S H ,HU Z H ,e ta l .T h ef l o r a ls c e n t e m i t t e d f r o m L i l i u m S i b e r i a a t d i f f e r e n t f l o w e r i n g s t a ge s a n dd i u r n a l v a r i a t i o n [J ].A c t a H o r t i c u l t u r a eS i n i -c a ,2013,40(4):693-702.C l o n i n g ,L o c a l i z a t i o na n dE x pr e s s i o nC h a r a c t e r i s t i c s o f F l o w e r F r a g a n c e S yn t h e s i sR e l a t e dG e n e L i M C S i n L i l i u m Z H A N G X i 1,2,L U OJ i n g l i n 2,WA N G H a o n a n 1,2,L IY i n g y i n g 1,2,L E N GP i n g s h e n g 1,2a n dHUZ e n gh u i 1,2(1.B e i j i n g A d v a n c e d I n n o v a t i o nC e n t e r f o rT r e eB r e e d i n g b y M o l e c u l a rD e s i g n ,B e i j i n g102206,C h i n a ;2.C o l l e g e o fL a n d s c a p eA r c h i t e c t u r e ,B e i j i n g U n i v e r s i t y o fA g r i c u l t u r e ,B e i j i n g102206,C h i n a )A b s t r a c t T h e f l o r a l v o l a t i l e s o f L i l i u m a r e c o m p o s e do f a l a r g en u m b e ro f t e r p e n o i d s ,a m o n g wh i c h 2-C -m e t h y l -D -e r y t h r i t o l -2,4-c y c l o p y r o p h o s p h a t es y n t h a s e (M C S )i so n eo f t h ek e y e n z ym e s i nt h e m o n o t e r p e n e s y n t h e s i s 2-C -m e t h y l -D -e r y t h r i t o l 4-p h o s p h a t e p a t h w a y .I no r d e r t o f u r t h e r e x pl o r e t h e -r e g u l a t i o nm e c h a n i s mo fM C So n l i l y f l o r a l f r a g r a n c e s y n t h e s i s ,w e c l o n e d t h e L i M C S g e n eo f L i l i u m S i b e r i a .S u b s e q u e n t l y ,t h eb i o i n f o r m a t i c s a n a l ys i sw a s c a r r i e do u t ,a n d t h e s u b c e l l u l a r l o c a t i o nw a s u s e d t od e t e r m i n e t h e l o c a t i o no f t h e p r o t e i ne n c o d e db y t h e g e n e ,a n d f i n a l l y t h e s p a t i o t e m po r a l e x -p r e s s i o n p a t t e r no f t h e g e n ew a s d e t e r m i n e d a c c o r d i n g to t h e q u a n t i t a t i v e r e a l -t i m eP C R.T h e r e s u l t s s h o w e d t h a t t h e o p e n r e a d i n g f r a m e o f t h e L i M C S g e n ew a s 669b p ,e n c o d i n g 222a m i n o a c i d s ,a n d h a d t h eh i g h e s t s i m i l a r i t y w i t h D i o s c o r e az i n g i b e r e n s i s .L i M C S p r o t e i nw a s l o c a t e d i n t h e c h l o r o p l a s t s o f t o b a c c o l e a f e p i d e r m a l c e l l s .T h e e x p r e s s i o n l e v e l o f L i M C S g e n ew a sh i g h e s t i n t h e e a r l y f l o w e r i n gs t a g e a n d l o w e s t i n t h e b u d s t a g e .D u r i n g f l o w e r i n g ,t h e e x p r e s s i o no f t h i s g e n ew a s h i g h e s tm a i n l y i n p e t a l s ,b u t l o wi no t h e r p a r t s .T h er e s u l t s p r o v e dt h a tL i M C S m a y b e i n v o l v e d i nt h es yn t h e s i so f m o n o t e r p e n o i d so f l i l i u mf r a g r a n c e ,w h i c h l a i da t h e o r e t i c a l f o u n d a t i o n f o r t h e s u b s e qu e n tu s eo f g e -n e t i c e n g i n e e r i n g m e t h o d s t o r e g u l a t e t h e r e l e a s e o f l i l y f r a gr a n c e .K e y w o r d s L i l i u m S i b e r i a ;L i M C S ;G e n e c l o n i n g ;E x p r e s s i o na n a l y s i s R e c e i v e d 2021-08-20 R e t u r n e d 2021-10-15F o u n d a t i o n i t e m T h e N a t i o n a l K e y R e s e a r c h a n d D e v e l o p m e n t P r o gr a m o f C h i n a (N o .2018Y F D 1000400);t h eN a t i o n a lN a t u r a lS c i e n c eF o u n d a t i o no fC h i n a (N o .31201645);t h eB e i j i n g N a t u r a l S c i e n c eF o u n d a t i o n (N o .6172006).F i r s t a u t h o r Z HA NG X i ,f e m a l e ,m a s t e rs t u d e n t .R e s e a r c ha r e a :r e g u l a t i o n m e c h a n i s m o f l i l y fr a -g r a n c e .E -m a i l :Z h a n gx i 0522@126.c o m C o r r e s p o n d i n g a u t h o r HU Z e n g h u i ,m a l e ,p r o f e s s o r .R e s e a r c ha r e a :p l a n t p h y s i o l o g y a n de c o l o g y .E -m a i l :b u a h u z e n gh u i @163.c o m (责任编辑:潘学燕 R e s p o n s i b l e e d i t o r :P A NX u e ya n )㊃989㊃8期张 茜等:百合花香合成相关基因L i M C S 的克隆㊁定位和表达特性研究Copyright ©博看网. All Rights Reserved.。
亚洲百合DNA的提取及RAPD-PCR反应体系的优化

亚洲百合DNA的提取及RAPD-PCR反应体系的优化
赵琛
【期刊名称】《中国农学通报》
【年(卷),期】2007(23)2
【摘要】为了促进亚洲百合分子水平的研究,以亚洲百合幼嫩叶片为材料,采用改良CTAB法提取亚洲百合基因组DNA,得到的DNA质量较高,适合于RAPD-PCR分析。
通过单因素优化得到在25μl反应体系中,亚洲百合RAPD分析的最佳反应体系:160μmol/LdNTPs,0.3μmol/L随机引物,TaqDNA聚合酶1.5U,Mg2+浓度2.0mmol/L。
【总页数】4页(P89-92)
【关键词】CTAB;亚洲百合;DNA提取;RAPD—PCR
【作者】赵琛
【作者单位】广东仲恺农业技术学院实习农场
【正文语种】中文
【中图分类】Q523
【相关文献】
1.卷丹百合RAPD-PCR反应体系的单因素逐项优化研究 [J], 韩凌;雷家军
2.卷丹百合RAPD-PCR反应体系的正交优化 [J], 韩凌;雷家军
3.蜡梅花瓣基因组DNA提取及RAPD-PCR反应体系优化 [J], 叶丽娟;禄久幸;杨秋生
4.猕猴桃模板DNA的提取及RAPD-PCR最佳反应体系的建立 [J], 陈万秋;李思光;罗玉萍;陈少风
5.黄瓜DNA提取及其RAPD-PCR反应体系的优化 [J], 孙敏;乔爱民;王和勇;曾建军
因版权原因,仅展示原文概要,查看原文内容请购买。
百合MDHAR基因的克隆与表达分析

百合MDHAR基因的克隆与表达分析陈莉;辛海波;李晓艳;李晓昕;义鸣放【期刊名称】《林业科学》【年(卷),期】2010(046)009【摘要】@@ 抗坏血酸(ascorbic acid,AsA)作为植物细胞内重要的自由基清除剂,是通过参与一系列的氧化还原反应而发挥抗氧化剂的功能.单脱氢抗坏血酸还原酶(monodehydroascorbate reductase,MDHAR)在抗坏血酸-谷胱甘肽循环中,可以将氧化型抗坏血酸(MDHA)还原为还原型抗坏血酸(AsA),在维持植物体内抗坏血酸的正常代谢和植物体氧化还原平衡方面起着重要作用(Chen et al.,2003;2005).【总页数】4页(P178-181)【作者】陈莉;辛海波;李晓艳;李晓昕;义鸣放【作者单位】中国农业大学农学与生物技术学院,北京,100193;中国农业大学农学与生物技术学院,北京,100193;中国农业大学农学与生物技术学院,北京,100193;中国农业大学农学与生物技术学院,北京,100193;中国农业大学农学与生物技术学院,北京,100193【正文语种】中文【中图分类】S718.46;Q943.2【相关文献】1.百合VAL2基因的克隆与表达分析 [J], 彭梦笛; 杨捷; 蒙娟; 贾桂霞; 何恒斌2.东方百合'索邦'组蛋白去乙酰化酶基因LoSorHDA1的克隆与表达分析 [J], 蒙娟;杨捷; 彭梦笛; 贾桂霞; 何恒斌3.百合LoNHX2基因的克隆与表达分析 [J], 王安琪;薛晓霞;左杨;张克中;崔金腾4.东方百合'索邦'几丁质酶基因的克隆与表达分析 [J], 崔祺;吴凡;詹鹏;汪莲娟;韩蕊莲;吴昀5.百合LbCAT和LbGPX基因的克隆与表达分析 [J], 迟博文;刘立;谢天;阎俊卉;赵子贤;文锦芬因版权原因,仅展示原文概要,查看原文内容请购买。
百合花发育相关MADSbox基因的克隆

园 艺 学 报 2004,31(3):332~336A cta Horticulturae Sinica百合花发育相关MADS box基因的克隆张 云1 刘青林1* 欧阳青2 蔡文启2(1中国农业大学观赏园艺与园林系,北京100094;2中国科学院微生物研究所,北京100080)摘 要:利用RT PCR的方法,从百合中克隆得到了3个接近全长的花发育相关MADS box基因片段Lf MA DS1~3。
其中Lf MA DS1和Lf MA DS3以往未见报道,分别与多种作物的A G、SE P同源基因具有较高的同源性;Lf MADS2与已发表的LMADS2对应区段的氨基酸同源性高达98 99%,推测可能是同一个基因。
关键词:百合;花发育;M A DS box基因;克隆中图分类号:S682 2 文献标识码:A 文章编号:0513 353X(2004)03 0332 05C loning o f Flo wer Development Associated MAD S bo x G ene Fragments in Lilium Zhang Yun1,Liu Qinglin1*,Ouyang Qing2,and Cai Wenqi2(1Dep ar tment of Or namental H or ticulture and L andscape A r chitecture,China A gr icultur al Univer sity,Beij ing100094, China;2I nstitute of M icrobiology,Chinese A cademy of Sciences,Beij ing100080,China)Abstract:Lf MADS1-3,three MADS box cDNA frag ments associated w ith flower development w ere cloned from L ilium using a RT PCR based cloning method.Lf M ADS1and Lf MADS3are new M ADS box genes in lilies,which have not been reported before.They show ed hig h sequence homolog y w ith A G and SEP homologs,respectively.The deduced amino acid sequence of Lf M ADS2shared 98 99%identity w ith the reported LM ADS2,w hich sug gested that they mig ht be the same gene.Key words:Lilium;Flower development;MADS box g ene;Cloning百合花姿态优美、芳香宜人,但花型较为单一,野生的百合原种均为单瓣花,目前国内市场上能够见到的也只有单瓣百合品种。
《2024年‘西伯利亚’百合LiCMK基因克隆及功能分析》范文

《‘西伯利亚’百合LiCMK基因克隆及功能分析》篇一西伯利亚百合LiCMK基因克隆及功能分析一、引言西伯利亚百合,以其独特的生长环境与美丽花朵闻名于世。
随着现代生物学技术的不断发展,基因克隆和功能分析已成为植物遗传学研究的重要领域。
本文以西伯利亚百合为研究对象,详细介绍其LiCMK基因的克隆过程以及对其功能的分析。
二、材料与方法1. 材料本实验以西伯利亚百合为研究对象,收集其基因组DNA和mRNA样本。
2. 方法(1)基因克隆:通过PCR技术扩增LiCMK基因,将PCR 产物与载体连接,转化至大肠杆菌中,筛选阳性克隆。
(2)功能分析:采用生物信息学方法对LiCMK基因进行序列分析,预测其功能;通过构建过表达和沉默载体,分析LiCMK 基因在植物生长、发育及抗逆等方面的作用。
三、实验结果1. LiCMK基因克隆通过PCR技术成功扩增出LiCMK基因,将其与载体连接后转化至大肠杆菌中,经过筛选得到阳性克隆。
对阳性克隆进行测序,确认序列正确无误。
2. LiCMK基因序列分析通过生物信息学方法对LiCMK基因进行序列分析,发现其编码一个蛋白激酶。
进一步分析该蛋白的结构和功能域,预测其在植物生长、发育及抗逆等方面可能发挥重要作用。
3. LiCMK基因功能分析(1)过表达载体构建及转基因植株分析:构建LiCMK基因过表达载体,转化西伯利亚百合,获得转基因植株。
通过对转基因植株的生长、发育及抗逆性等方面进行分析,发现LiCMK基因在植物生长和抗逆方面具有重要作用。
(2)沉默载体构建及功能验证:构建LiCMK基因沉默载体,转化西伯利亚百合,获得沉默植株。
与野生型植株相比,沉默植株表现出明显的生长和发育异常,进一步证实了LiCMK基因在植物生长和发育中的重要作用。
四、讨论本实验成功克隆了西伯利亚百合的LiCMK基因,并通过生物信息学方法和转基因技术对其功能进行了分析。
结果表明,LiCMK基因编码一个蛋白激酶,在植物生长、发育及抗逆等方面具有重要作用。
百合分子生物学研究进展
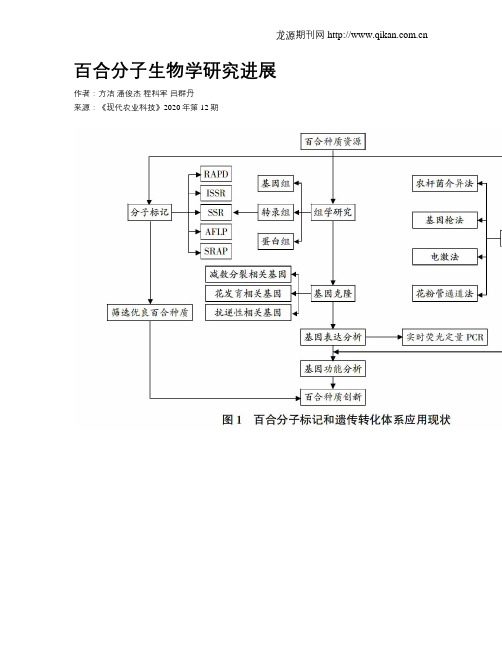
百合分子生物学研究进展作者:方洁潘俊杰程科军吕群丹来源:《现代农业科技》2020年第12期摘要 ; ;综述了百合分子生物学的研究进展,主要包括4个方面,即百合基因组学、转录组学和蛋白组学的研究;百合遗传多样性分析和5种分子标记(RAPD、ISSR、SSR、AFLP、SRAP)的开发和利用;百合遗传转化体系的构建,包括农杆菌介导法、基因枪法、电激法和花粉管介导法;百合减数分裂基因、花发育基因及抗逆性相关基因的克隆和表达分析。
最后指出了当前研究存在的问题,并展望了进一步研究的方向。
关键词 ; ;百合;分子标记;遗传转化;基因克隆中图分类号 ; ;S644.1 ; ; ; ;文献标识码 ; ;A文章编号 ; 1007-5739(2020)12-0069-05 ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; 開放科学(资源服务)标识码(OSID)A ;Review ;on ;Molecular ;Biology ;of ;LilyFANG Jie ; ;PAN Jun-jie ; ;CHENG Ke-jun ; ;LV Qun-dan *(Lishui Institute of Agriculture and Forestry Sciences, Lishui Zhejiang 323000)Abstract ; ;This paper reviewed the following four aspects in lily molecular biology: research progresses of lily genomics, transcriptomics and proteomics; the development and utilization of five types of molecular markers including RAPD, ISSR, SSR, AFLP and SRAP in lily genetic diversity analysis; construction of lily transformation system including Agrobacterium mediated method, gene gun bombardment method, electroporation method and pollen tube pathway method; cloning and expression analysis of genes related to meiosis, flower development and stressresistance. Finally, the current research defects and prospects of lily molecular biology were summarized.Key words ; ;lily; molecular marker; genetic transfomation; gene cloning百合属(Lilium)植物属于百合科(Liliaeeae)多年生球根植物,集药用、食用及观赏于一体,具有极大的商业价值。
《2024年‘西伯利亚’百合LiCMK基因克隆及功能分析》范文

《‘西伯利亚’百合LiCMK基因克隆及功能分析》篇一西伯利亚百合LiCMK基因克隆及功能分析一、引言西伯利亚百合,作为一种具有独特魅力的花卉,其基因组的研究对于植物学、遗传学以及分子生物学等领域具有重要的意义。
近年来,随着基因克隆技术的不断发展,越来越多的研究者开始关注于植物基因的克隆与功能分析。
本篇论文将主要探讨西伯利亚百合中LiCMK基因的克隆及其功能分析,为后续的基因工程应用提供理论依据。
二、材料与方法1. 材料本实验所使用的西伯利亚百合样本采自我国北方地区。
此外,实验所需的各种酶、载体、试剂盒等均来自可靠的生物科技公司。
2. 方法(1)基因克隆:采用PCR技术从西伯利亚百合的cDNA中扩增出LiCMK基因片段,并将其克隆至表达载体中。
(2)功能分析:通过生物信息学手段对LiCMK基因进行序列分析,预测其编码的蛋白质功能;并通过构建转基因植物,观察LiCMK基因在植物生长发育过程中的作用。
三、实验结果1. LiCMK基因的克隆通过PCR技术成功地从西伯利亚百合的cDNA中扩增出LiCMK基因片段,经过序列测定验证了其正确性。
将该基因片段克隆至表达载体中,构建了重组表达载体。
2. LiCMK基因的序列分析通过生物信息学手段对LiCMK基因进行序列分析,发现该基因编码一个具有跨膜结构的蛋白质。
进一步分析表明,该蛋白质具有多种功能域,可能参与植物的生长、发育和代谢等过程。
3. LiCMK基因的功能分析通过构建转基因植物,观察到LiCMK基因在植物生长发育过程中具有一定的作用。
具体表现为,过表达LiCMK基因的植物在生长速度、抗逆性以及代谢等方面均表现出明显的优势。
这表明LiCMK基因具有重要生物学功能。
四、讨论本实验成功克隆了西伯利亚百合中的LiCMK基因,并对其进行了功能分析。
结果表明,LiCMK基因在植物生长发育过程中具有重要作用,可能参与植物的生长、发育和代谢等过程。
此外,过表达LiCMK基因的植物在生长速度、抗逆性以及代谢等方面均表现出明显的优势,这为后续的基因工程应用提供了重要的理论依据。
百合查尔酮合成酶基因克隆及其转化烟草的花色表达分析

百合查尔酮合成酶基因克隆及其转化烟草的花色表达分析陈洁;安利清;王涛;姚娜;李潞滨;杨凯【期刊名称】《西北植物学报》【年(卷),期】2012(32)8【摘要】以‘西伯利亚’百合为试材,利用PCR技术克隆了查尔酮合成酶基因(CHS),构建了CHS基因的正义和反义植物表达载体,采用农杆菌介导法转化烟草叶盘,获得了转正义CHS基因的本明烟草18株,转反义CHS基因的普通烟草21株,总转化率为26.0%.高效液相色谱法(HPLC)检测结果显示,正义CHS转基因的本明烟草类黄酮含量升高14.0%~59.7%,反义CHS转基因的普通烟草类黄酮含量降低44.5%~76.4%.花色观察结果显示,正义转基因烟草的花瓣颜色未见变化,反义转基因烟草部分植株的花瓣颜色变浅.研究表明,CHS基因遗传转化是进行花色调控的有效手段之一.%The chalcone synthase gene (.CHS) was cloned from Lilium oriental hybrid 'Siberia' by PCR method. Sense and antisense CHS were constructed into plant expression vectors pCAMBIAl304,and then transformed into tobacco by agrobacterium mediated approach. 18 plants of sense CHS transgenic Nicoti-ana benthamiana and 21 plants of antisense CHS transgenic Nicotiana tabacum were obtained and the total transform ratio was 26. 0%. High Performance Liquid Chromatography (HPLC) suggested that CHS expression accumulation amplified by 14.0%~59. 7% in AT. benthamiana transformed with sense gene, and the accumulation declined by 44. 5% - 76. 4% in N. tabacum transformed with antisence gene compared to the wild type. The petals of the transgenic N.benthamiana had no color changes, but the petal color in some of the transgenic N. tabacum plants was faded. The results indicated that the transformation of CHS gene'was an effective method to flower color regulation.【总页数】7页(P1511-1517)【作者】陈洁;安利清;王涛;姚娜;李潞滨;杨凯【作者单位】中国林业科学研究院林业研究所林木遗传育种国家重点实验室,北京100091;北京农学院农业应用新技术北京市重点实验室,北京102206;北京农学院农业应用新技术北京市重点实验室,北京102206;3北京林业大学园林学院,北京100083;中国林业科学研究院林业研究所林木遗传育种国家重点实验室,北京100091;中国林业科学研究院林业研究所林木遗传育种国家重点实验室,北京100091;北京农学院农业应用新技术北京市重点实验室,北京102206【正文语种】中文【中图分类】Q789【相关文献】1.罗布麻查尔酮合成酶基因克隆及序列分析 [J], 李苗;李国旗2.链霉菌查尔酮合成酶基因克隆及原核表达载体构建 [J], 肖静;吕玉红;李园园;陈雪梅;彭廷文;周春伶;曾令荣;保玉心;凌锌3.嫁接陆地棉查尔酮合成酶与查尔酮异构基因的克隆及r表达分析 [J], 宋成攀;夏松波;王孝刚;张教海;秦鸿德;张友昌;冯常辉;别墅4.观赏植物花色形成关键酶查尔酮合成酶研究进展 [J], 马喆5.观赏植物花色形成关键酶查尔酮合成酶研究进展 [J], 马喆;因版权原因,仅展示原文概要,查看原文内容请购买。
《2024年‘西伯利亚’百合LiCMK基因克隆及功能分析》范文

《‘西伯利亚’百合LiCMK基因克隆及功能分析》篇一西伯利亚百合LiCMK基因克隆及功能分析一、引言近年来,植物基因组学与功能基因组学研究领域迅速发展,基因克隆和功能分析已成为探究植物基因的重要手段。
其中,百合作为一种具有观赏价值的植物,其基因组研究具有较高的科研价值。
本篇论文旨在研究西伯利亚百合(Lilium siberianum)的LiCMK基因的克隆与功能分析,以期为后续百合基因编辑和遗传改良提供理论基础。
二、材料与方法1. 材料实验所用的西伯利亚百合品种为栽培型百合品种,实验过程中所用试剂和仪器均符合分子生物学实验要求。
2. 方法(1)基因克隆:通过PCR技术,以西伯利亚百合cDNA为模板,扩增LiCMK基因。
(2)生物信息学分析:利用生物信息学软件对LiCMK基因进行序列分析、结构预测及功能注释。
(3)功能验证:通过转基因技术,将LiCMK基因转入模式植物中,观察其表达对植物生长及抗逆性的影响。
三、实验结果1. LiCMK基因克隆通过PCR技术成功扩增出LiCMK基因的cDNA序列,序列比对结果表明,该基因与已报道的百合基因高度相似。
2. 生物信息学分析(1)序列分析:LiCMK基因编码的蛋白质具有典型的蛋白激酶结构域,属于蛋白激酶家族成员。
(2)结构预测:通过生物信息学软件预测,LiCMK基因编码的蛋白质具有较高的稳定性,可能参与细胞内的多种生物过程。
(3)功能注释:通过数据库检索,发现LiCMK基因与植物生长、抗逆性等过程密切相关。
3. 功能验证将LiCMK基因转入模式植物中,观察其表达对植物生长及抗逆性的影响。
结果显示,LiCMK基因的表达能够显著促进植物生长,提高植物的抗逆性。
四、讨论本实验成功克隆了西伯利亚百合的LiCMK基因,并通过生物信息学分析和功能验证,证实了该基因在植物生长及抗逆性方面的重要作用。
这为后续百合基因编辑和遗传改良提供了重要的理论基础。
然而,LiCMK基因的具体作用机制仍需进一步研究。
百合单萜合成酶基因的克隆与序列分析

百合单萜合成酶基因的克隆与序列分析李天娇;冷平生;杨凯;郑健;胡增辉【期刊名称】《北京农学院学报》【年(卷),期】2014(29)3【摘要】单萜化合物是‘西伯利亚’百合花香的主要成分,为了研究百合单萜化合物的合成与代谢机理,以百合的花瓣为材料,根据GenBank发表的单萜合成酶基因的保守区设计1对简并引物,通过RT-PCR与RACE相结合的方法,克隆得到一个单萜合成酶基因的cDNA序列,命名为Li-mTPS2.该基因全长为1 766 bp,包含1 608 bp的基因开放阅读框(ORF),45 bp的5'UTR和113 bp的3'UTR,编码535个氨基酸,含有萜烯合成酶(TPS)保守序列DDxxD以及TPSb亚家族共有的RRx8W基序.氨基酸同源性分析表明该基因所编码蛋白的氨基酸序列与小苍兰芳樟醇合成酶、姜花单萜合成酶、六出花月桂烯合成酶的同源性分别为55%,53%,51%.【总页数】5页(P6-10)【作者】李天娇;冷平生;杨凯;郑健;胡增辉【作者单位】北京农学院园林学院,北京102206;北京农学院园林学院,北京102206;北京农学院植物科学技术学院,北京102206;北京农学院园林学院,北京102206;北京农学院园林学院,北京102206【正文语种】中文【中图分类】S682.2【相关文献】1.岷江百合单萜合酶基因克隆与表达分析 [J], 李路路;王欢;孙明;张启翔2.赤桉单萜合成酶基因的分子克隆与生物信息学分析 [J], 陈鸿鹏;吴志华;尚秀华;高灵;谢耀坚3.茶树单萜合成酶CsTPS基因的克隆及表达分析 [J], 王鹏杰;陈丹;俞滢;林浥;曹红利;杨国一;陈笛;叶乃兴4.艾草(Artemisia argyi)单萜合成酶基因的克隆及序列分析 [J], 刘雷;罗英;陶红;姜立春;徐应文;刘群;黄坤5.香蕉叶片颗粒结合性淀粉合成酶Ⅰ和可溶性淀粉合成酶Ⅲ基因的克隆与序列分析[J], 匡云波;赖钟雄因版权原因,仅展示原文概要,查看原文内容请购买。
《2024年‘西伯利亚’百合LiCMK基因克隆及功能分析》范文

《‘西伯利亚’百合LiCMK基因克隆及功能分析》篇一范文字体:西伯利亚百合LiCMK基因克隆及功能分析一、引言西伯利亚百合(Lilium sibiricum)是一种具有重要观赏价值的植物,其生长环境及植物本身的遗传特征使它在花卉种植及生物医药研究中具有重要的地位。
其中,对特定基因的研究成为了解析西伯利亚百合各种特性的关键步骤。
本研究中,我们将深入讨论西伯利亚百合的LiCMK基因的克隆及功能分析。
二、材料与方法(一)材料本实验使用的西伯利亚百合由我实验室自主种植,并在适当的生长条件下进行取样。
(二)方法1. 基因克隆:通过PCR技术,从西伯利亚百合的基因组DNA中扩增出LiCMK基因。
2. 序列分析:利用生物信息学软件对LiCMK基因的序列进行分析,包括开放阅读框(ORF)预测、蛋白结构预测等。
3. 功能分析:通过构建过表达和敲除的转基因植物,分析LiCMK基因在植物生长发育过程中的功能。
三、LiCMK基因克隆及序列分析(一)基因克隆我们成功地从西伯利亚百合中克隆出了LiCMK基因,其PCR产物经测序验证与预期序列一致。
(二)序列分析通过生物信息学软件分析,我们发现LiCMK基因具有完整的开放阅读框,编码的蛋白质具有典型的结构域和功能域。
此外,我们还预测了该蛋白的三维结构,为后续的功能研究提供了基础。
四、LiCMK基因的功能分析(一)转基因植物的构建与筛选我们构建了LiCMK基因的过表达和敲除的转基因植物,并通过PCR和RT-PCR等方法对转基因植物进行了筛选和验证。
(二)功能分析通过对转基因植物的生长、发育和生理特性的观察和测定,我们发现LiCMK基因在植物的生长过程中起到了重要作用。
过表达LiCMK基因的植物表现出更强的生长势和抗逆性,而敲除LiCMK基因的植物则表现出生长缓慢和抗逆性减弱的现象。
这表明LiCMK基因在西伯利亚百合的生长发育及抗逆过程中发挥了重要的调控作用。
五、讨论与结论本实验成功克隆了西伯利亚百合的LiCMK基因,并对其进行了序列分析和功能分析。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
CCTTGCTGAGTGCGGGACG竹C
CCGGATCCTCTAGAGCGGCCGC(T)17
CCGGATCCTCTAGAGCGGCCGC ATGGAACTGGAATGGllAAG ATAGCAACp汀‘AC声汀AGCAGG
MDHAR—F
MDHAR.R
TmGGCGTTCAAGGAGCAG
CACCCACAACAACA A,l’GTCAGC
嚣瓣麓隧蠹瓣霪l蕊灞爹}剖j蒌l萋鐾i}麟凄
黼蠡攀篓蕊夔lll麟霆瓣澍鏊建l瓣霪|||
滋|鬟|I童瀚;i瓣蓍瓣}l整鎏器薰}蠢鹚i
激I舞 ㈣矮潮鞣露瓣撼霞
:岛紫戮纛?“A麓t:鬻。O“s:祟警“M瓣c;篇麓
u雨舟;Br:^自菜, 拟m并; 水稻 冰¨}J巾花
FIE 5 Homology
c01¨parIson
圈3MDIIAR箍嗣5’片段巢式PCR扩增『U诛田卅 M:DNA分f量DL2000t l扩增产糟
ElHmphorKIsⅡnes{PCRⅡMDHAR gene 5’end M:MarkerDL2000;h PCR pred.ct
Fig.3
M87”DNA鬈滞D。L20蚺001“1譬荔努潮”
t
分,置
t扩增产物
…Fig.4 E。:::::黜蕊:裟:黜≯
2
2百合MDHAR序列分析 将克隆得到的保守区片段,3’端核廿醴序列和5’端核心酸序列拼接.扶得了百台单脱氢
抗坏血酸还帐酶摹州MDHARl502bp的全K eDNA.5+UTR和yUTR分别为26bp和17lbp,
包青一个1305b口的开放瞄读框(ORF)(嘲4),编码434个氰基酸。从氮基酸序列同源性
Key words:Lilium longiforum;monodehydroascorbate
reduetase gene(MDHAR);Cloning;Expression
抗坏血酸(ascorbic acid,AsA)作为植物细胞内重要的自由基清除剂,是通过参与一系
列的氧化还原反应而发挥抗氧化剂的功能。单脱氢抗坏血酸还原酶(monodehyroascorbate
the root,bulb and leaf,and it axprcsscxt highly in the
Wflq increased with the raising of
and leaf.Under
H202 stress,the expression
ofLIMDHAR gene
H202
concentration.
“
+物技术
比较图谱可以看…(图5),百合与水稻和冰叫日中花的l_源性最高,分别达到83%和82%:
与人白菜和拟南芥的I可源性也分刖让到76%,f11 74%。系统进化树分析也表明谈基Ⅲ与水稻
_l冰111日中花的亲缘笑系较近(图6),我HJ将儿命名为LIMDHAR。
粼蠹嚣蠢蠹纛髓霭嚣霪藿勰飘
i
other plants
!塾垒!!!
引物
PRl PF2 PF3 PR2 PR3 PR4 AP i AP2 Actin.F Actin—R Primer
!坠!E:i巴!!:骂竺:巴!£
5'-3’
5'-3’核酸序列Seqence
GAGTAGAARTATGGNAGRTARTC CCTGAACCTTGGTGCATGCC GCTGTAGGGGATGTCGCCAC .CCAAGCTCAAGACCAATGTA"rCC CCTTGCTGAGTGCGGGACGTTC
二i::鲨::I::i::I:.I::=|
圈6 n台、lDIlAR々n种枷斯省自缸*融m弹”别的蒙娄分析
Flz 6Cluslering nfthe ded uced
LI:^台,Br:人口柴:^|:拟南井,Os:水月.31c:冰【2 rj坤托 a洲,lo acid s2quP|Ice s ofM[)lIAR’、ith h¨molo“u¨s a111]110
Cloning and expression analysis of MDHAR from Lilium
Chen Li
long洳lff,lm
Xin Hai-bo
Li Xiao—yah
Li Xiao-xin
and
Yi Ming-fang・
(Department ofOrnamental Horticulture and Landscape Architecture,China Agricultural University,
reductase,MDHAR)在抗坏血酸.谷胱甘肽循环中,可以将氧化型抗坏血酸(MDHA)还原 为还原型抗坏血酸(ASC),在维持植物体内抗坏血酸的正常代谢和植物体氧化还原平衡方 面起着重要作用(Chert 2003;Chen 2005)。 百合作为主要的切花和盆花材料,在国内外花卉市场上占有重要地位。百合¨1eI plants
ac…Iequences
LI:Lillumn’w批ⅧⅢ.B“日…Ⅱ’叩口sllbsp pckme叫s(”__{q7R^)I、I J、lc:№删^删^㈨ⅢH
0s:Or_l=a㈣v口I—oporticaGroup(D85164
2.3百台iIDIL4R
of BmillO acid sequeflce5
align…z ofl.iIvMDHAR”lfh
LhLiliumlongmotum;Br:Brassi*uMp4 subsp pekinensi s{A、039786);At:4rabldop、isl|№tmna(NM・12044); Os:Oo_:aⅫr枷Japonica Gruu p(085764);3,1“I,exeBtbOabghetltuttl crjjlalllmtm(AJ301553l
1.4百合MDHAR基因3’端和5’端的克隆
根据已克隆的百合MDHAR保守区片段,设计两条3’RACE上游引物PF2和PF3,以 APl引物进行逆转录,以AP2为下游引物,进行巢式PCR,对3’端进行扩增。反应程序为:
94℃5 min,94℃45s,52℃45s,72℃lmin,35cycle,72℃延伸lOmin。
根据已克隆的百合MDHAR保守fx.片段,设计3条5’RACE下游引物PR2、PR3和PR4, 以PR2特异引物进行逆转录,对cDNA模板进行加尾反应,再以其为模板进行PCR反应。 PCR反应程序为:94℃5min,94℃45s,50℃45s,72℃lmin,35cycle,72℃延伸10min。
表1引物序列
1.5序列测定 将所有PCR产物凝胶电泳后同收目的片段,与PMDI 8-T载体连接夕i:转化DH5a,重组 质粒鉴定后送北京华人基因公司进行测序。
1.6
RT-PCR的表达分析 根据百合MDHAR eDNA序列设计一对基闪特异引物MDHAR.F和MDHAR-R,以Actin
基凶作为内参,采用半定量RT-PCR方法研究MDHAR在根、鳞苇、叶不同组织器官的表达
百合MDHAR基因的克隆与表达分析
陈莉,辛海波,李晓艳,李晓昕,义鸣放・
(中国农业大学观赏冈艺与园林系,北京100193) 摘要:采用RACE技术,从铁炮百合‘白天堂’(‘white heaven’)组培苗叶片中克隆得到一个单脱氢 抗坏m酸还原酶摹因(MDHAR)的cDNA序列,伞长1502bp,推断其编码434个氨纂酸。序列分析表明 该基因与其他植物的MDHAR幕因具有较高的同源性。RT-PCR表达分析表明,该基因在百合的根、鳞茎、 叶中均有表达,在根和叶中表达量较高。在H202氧化胁迫下,该幕因的表达量随着H202浓度的提高而增 加。 关键词:铁炮百合;MDHAR;克隆:表达
模式,以及trl’片在0、O.5、5、50、100mMH202溶液中处理lh后的表达情况。
2009年中l目球擞花卉年会空“【论文柴
生物技术
2结果与分析
2.1百合单脱氢抗坏血酸还原酶基因MDHAR全长cDNA序列的克隆 以百合叫片攫取的RNA逆转录的cDNA为模扳,以PFI和PRI为引物.进行保守隧的 PCR扩增(圈I).经剽序』£核廿酸序列人小为920吣.剧源序列比对结粜显示,与水稻『目
润气候,越夏的百合经常出现生长停滞、植株低矮、花朵败育等现象,严重影响了切花质量,
并造成百合种球退化。夏季高温已成为限制国内百合周年生产的重要因素。 本研究采用RACE技术从铁炮百合‘白天堂’叶片中克隆得到MDHAR的eDNA全长, 并对其不同组织器官及氧化胁迫下时空表达特征进行了研究,为通过基因工程的方法提高植 物活性氧清除酶活性,从而提高植物的抗氧化胁迫能力,增强抗氧化代谢的水平,提高植物 的抗逆性提供理论依据。
源性逃77%,与白菜lHi!I_性业74%,与拟南芥剧源性选72%,
2000bp
2000bp
000bp 750bp 500bp
1000bp
150抽
500bD
250bp 100bp
250bp
lOObp
崮1MDHAR基闻中问J}段电泳目谢 M:DNA分r皿DL2000;1:扩增产物
Fi91
图2MDItAR差固3’片段巢式PCR扩增电诛围谌 M:DNA分f量DL20∞;1:扩增产物
序结粜表啊该3’PACE产物人小为597bp(I目2)。结合保守K片段设计』条下游引物,用基
闲特异引物PR2进行逆转录.PR3和pR4分别进行5’端第一轮和第二轮PcR扩增,经测序 该5’端核序酸序列人小为J90bp(图3)。
2000bp
zooobp
∞瓣∞
250bp 1 OObp
750bp 500bp 250bp 100bp
胶电泳,然后进行逆转录。
1.3保守片段的克隆
根据GenBank核酸数据库中报道的拟南芥、大白菜、芥菜、冰叶日中花MDHAR的mRNA 序列,通过DNAMAN设计保守Ix.兼Jt:弓l物PFl和PRl。PCR反应程序为:94℃5 rain,94℃