褐飞虱体内类酵母共生菌
褐飞虱体内类酵母共生菌共29页

•
7、心急吃不了热汤圆。
•
8、你可以很有个性,但某些时候请收 敛。
•
9、只为成功找方法,不为失败找借口 (蹩脚 的工人 总是说 工具不 好)。
•
10、只要下定决心克服恐惧,便几乎 能克服 任何恐 惧。因 为,请 记住, 除了在 脑海中 ,恐惧 无处藏 身。-- 戴尔. 卡耐基 。
END
16、业余生活要有意义,不要越轨。——华盛顿 17、一个人即使已登上顶峰,也仍要自强不息。——罗素·贝克 18、最大的挑战和突破在于用人,而用人最大的突破在于信任人。——马云 19、自己活着,就是为了使别人过得更美好。——雷锋 20、要掌握书,莫被书掌握;要为生而读,莫为读而生。——布尔沃
不同虫源及致害性褐飞虱体内类酵母共生菌的形态特性、数量及其种类鉴定[博士后学位论文]
![不同虫源及致害性褐飞虱体内类酵母共生菌的形态特性、数量及其种类鉴定[博士后学位论文]](https://img.taocdn.com/s3/m/06d22bb665ce0508763213a9.png)
浙江大学博士后学位论文不同虫源及致害性褐飞虱体内类酵母共生菌的形态特性、数量及其种类鉴定姓名:陈法军申请学位级别:博士后专业:生态学指导教师:俞晓平;叶恭银20060622浙江大学(浙江省农科院)博b后研究工作报告第一幸国内外研究进展2共生菌的分类地位2.1形态学研究酵母类真菌的常规分类主要依赖其形态和生理生化指标。
在“昆虫.共生菌”这一共生关系研究中,共生菌的离体培养至关重要(Baumanneta1.,1995;苗雪霞和丁德诚,2003)。
至今,国内外尚无有关褐飞虱体内的类酵母共生菌的离体培养报道。
这严重阻碍了该类共生菌的生理生化特性实验的开展。
而作为单细胞个体,其常规的形态分类又受到很大的限制。
从而使得该类共生菌的分类研究至今止步不前。
根据菌体离体后的显微结构观察和常规形态(见图1.1A,B,C和D个体形态;图1.2A,B,C和D出芽繁殖;图1.3A,B,C和D离体培养后观察到的假菌丝)分类,褐飞虱体内共生菌是类酵母共生菌(Yeast.1ikesymbiotes,YLS),属于子囊菌亚门的核菌纲(Fyrenomyeetes)、果囊菌亚纲(Pyrenomycete)的假丝酵母属(Candidae)(Nasu&Suenaga,1958;Nasu,1963;Nasuelal.,1981;Noda'1977;Chenetal.,198la'1981b;Eyaetal.,1989)。
图l—l示褐飞虱体内的类酵母共生菌的个体形态(陈法军和张珏锋,未发表资料)(A一杆状;B一梭形;C一球形;D一卵形)(分离子卵内;光学显微镜F观察)Fig.I-IShapesofyeast-likeendosymbiote(YLES)inbrownplanthopper(BPH),Nilaparvatalugens(ChenFa-Jun&ZhangJue-Feng,unpublisheddata)(A·poleshape;B—fusiformshape;C·globoseshape;D—ovalshape)(YLESisolatedfromBPHeggs;lightmicroscope;×100oillensand10lambar)塑坐艘i!坚型望垡黧熬!!型竺登曼!避墼!§!堂蹩塾21芝竺!!h型嬲l-2示褐飞虱熔内的类酵母共生蓥的出芽繁臻方式(陈法翠釉张珏锋,未发表资棼努份一杼获:B一梭形;C一璩彤;D一卵形)(分离于弗内;兜学显徽镜F观察)Fig.1-2Buddingproliferationofyeast*likeendosymbiote(YLES)inBPH,Ⅳlugens(ChenFa-Jun&ZhangJue-Feng,unpublisheddata)缓一poleshape;B—fusiformshape;C*globoseshape;D·ovalshape)(YLESisolatedfromBPHeggs;lightmicroscope;X100oillensand10pmbar)蚕14示离体培养勰褥飞蠢僖肉豹类酵母共生藏的德藿丝《陈法攀期张珏锋,来发表资睾葺)(光学显微镜F观察;陈法军等,未发表资料)Fig.1-3Pseudomyeeliumofthecultivatedyeast-likeendosymbiotes(YLES)加vitroisolatedfromtheeggsofBPH糙lugens(ChertFa-Jun&ZhangJue-Feng,unpublisheddata)(1ightmicroscope;。
褐飞虱体内细菌群落的 PCR-DGGE 分析

褐飞虱体内细菌群落的 PCR-DGGE 分析徐红星;郑许松;杨亚军;王新;傅强;叶恭银;吕仲贤【期刊名称】《中国水稻科学》【年(卷),期】2014(000)002【摘要】应用变性梯度凝胶电泳(denatured gradient gel electrophoresis,DGGE)分析了褐飞虱体内细菌的群落结构。
结果表明,褐飞虱1龄~5龄若虫体内细菌的菌群结构随其生长发育而异,而羽化后的成虫与1龄若虫的菌群结构具有较高的相似性。
根据 DGGE 聚类分析结果,9个地理种群褐飞虱体内细菌可分为3类,菲律宾单独一组,泰国与我国的海南、云南和浙江种群为一组,越南与我国的广西、湖南和江西种群为一组。
取食感虫品种的 TN1种群褐飞虱体内细菌的群落结构相似,与取食抗虫品种的 ASD7种群和 Mudgo 种群之间具有较明显的差异,但取食不同抗虫水稻品种的褐飞虱还未能得到有效区分。
%The bacterial community structure in different virulent populations of brown planthopper (BP H ), Nilaparvata lugens (Stål),was analyzed by denatured gradient gel electrophoresis (DGGE).The results showed that the bacterial community in BPH nymph from the 1 st to the 5th instars varied with nymphal growth and development, and the bacterial community in the 1 st instar BPH nymph was similar to that in adults.Nine geographic BPH populations were obviously divided into three groups according to the cluster analysis based on DGGE fingerprint.The first group came from the Philippines,the second from Thailand and the provinces of Hainan,Yunnan and Zhejiang of China,and the third from Vietnam and the provinces ofGuangxi,Hunan,and Jiangxi of China.BPH populations on Mudgo (with resistant gene Bph1 )and ASD7 (with resistant gene bph2 )have similar bacterial community,while differed to that fed on susceptible rice variety,TN1 .【总页数】6页(P217-222)【作者】徐红星;郑许松;杨亚军;王新;傅强;叶恭银;吕仲贤【作者单位】浙江大学昆虫科学研究所,杭州 310029; 浙江省农业科学院植物保护与微生物研究所,杭州 310021;浙江省农业科学院植物保护与微生物研究所,杭州 310021;浙江省农业科学院植物保护与微生物研究所,杭州 310021;浙江省农业科学院植物保护与微生物研究所,杭州 310021;中国水稻研究所,杭州310006;浙江大学昆虫科学研究所,杭州 310029;浙江省农业科学院植物保护与微生物研究所,杭州 310021【正文语种】中文【中图分类】S435.112+.3;Q18【相关文献】1.褐飞虱体内细菌型共生菌的分布 [J], 唐明;徐小蓉;洪鲲;乙引2.基于16S rRNA高通量测序的灰飞虱体内细菌群落结构及多样性分析 [J], 杨晓晴;王正亮;王天召;俞晓平3.钉螺体内细菌群落结构PCR-DGGE分析方法的建立 [J], 倪加加;余育和;胡合华4.经除草剂处理的水稻对褐飞虱体内几种酶及水稻受褐飞虱为害程度的影响 [J], 刘井兰;吴进才;袁树忠;徐建祥;姜永厚5.南方水稻黑条矮缩病对白背飞虱和褐飞虱体内3种水解酶活性的影响 [J], 杨洪;蒋德春;金道超;周操;杨熙彬因版权原因,仅展示原文概要,查看原文内容请购买。
褐飞虱唾液分泌物中细菌多样性的初步分析

褐飞虱唾液分泌物中细菌多样性的初步分析作者:唐明等来源:《江苏农业科学》2014年第04期摘要:褐飞虱是我国水稻生产中的重要害虫,其唾液在取食过程中起着重要的作用。
在笔者的前期研究中发现,褐飞虱在取食时会分泌凝胶状唾液和水溶性唾液。
笔者收集了褐飞虱的水溶性唾液分泌物,通过建立16S ribosomal DNA(rDNA)文库检测了唾液分泌物中的微生物组成,共发现6种细菌,它们都属于变形菌门,其中属于γ变形菌亚门、β变形菌亚门的分别有4种、2种。
此外研究还对褐飞虱取食后残留在水稻叶鞘中的口针鞘内部进行了透射电镜观察,发现在口针鞘内部和周围都有微生物的存在。
研究结果可为进一步认识褐飞虱的取食行为以及褐飞虱体内的细菌型共生菌的生物学特性提供参考和依据。
关键词:褐飞虱;唾液分泌物;细菌多样性;细菌型共生菌中图分类号: S435.112+.3 文献标志码: A 文章编号:1002-1302(2014)04-0090-03收稿日期:2013-08-17基金项目:国家自然科学基金(编号:31201514);贵州省科技基金(编号:黔科合J字[2012]2277号);贵州省高层次人才科研条件特助经费项目(编号:TZJF-2011年-39号);贵阳市社会发展与民生科技项目(编号:筑科合同[2012103]85号)。
作者简介:唐明(1979—),男,贵州遵义人,博士,讲师,主要从事植物保护研究。
Tel:(0851)6702541;E-mail:tangming2006@。
通信作者:乙引,男,重庆合川人,博士,教授,主要植物生理生化研究。
Tel:(0851)6702541;E-mail:yiyin@。
褐飞虱又名稻褐虱[Nilaparvata lugens (Stl)],属同翅目(Homoptera)蝉亚目(Cicademorpha)飞虱科(Delphacidae)。
褐飞虱是典型的刺吸式口器昆虫,通过口针取食水稻韧皮部维管束中的汁液,因此褐飞虱是水稻生产中的重要害虫,具有季节性、迁飞性、暴发性和猖獗性等特点,给我国水稻生产造成了严重损失[1]。
不同地理种群褐飞虱体内共生酵母菌的个体大小及数量差异

不同地理种群褐飞虱体内共生酵母菌的个体大小及数量差异陈法军;张珏峰;周彦铨;叶恭银;吕仲贤;俞晓平【期刊名称】《昆虫知识》【年(卷),期】2006(43)4【摘要】褐飞虱腹部脂肪体内普遍存在共生酵母菌,该类共生菌在褐飞虱的生理代谢和营养利用等方面起着重要作用。
采用冷冻切片技术结合显微摄像系统观察法测定不同地理种群褐飞虱体内共生酵母菌的个体大小和数量。
试验结果表明,不同地理种群褐飞虱雌(雄)成虫体内共生菌的个体大小和数量差异显著,依次为:对照种群>广西种群>浙江种群>福建种群。
结合虫体内脂肪和糖元含量的分析得出,褐飞虱体内共生酵母菌的长、宽度和数量与脂肪和糖元含量显著正相关。
文章从共生酵母菌的角度解释了不同地理种群褐飞虱致害性变异的内在原因,并推测,迁飞过程所导致的褐飞虱体内脂肪和糖元的消耗影响了该类共生菌的数量和质量,并最终导致褐飞虱对抗性品种水稻的致害性减弱。
【总页数】6页(P460-465)【关键词】褐飞虱;共生酵母菌;个体大小;数量;地理种群;致害性【作者】陈法军;张珏峰;周彦铨;叶恭银;吕仲贤;俞晓平【作者单位】浙江省农业科学研究院植物保护与微生物研究所,杭州310021;浙江大学应用昆虫学研究所,杭州310029;浙江省农业科学研究院卫生所,杭州310021【正文语种】中文【中图分类】S812;S433【相关文献】1.水稻与褐飞虱互作过程中虫体内类酵母共生菌的个体大小及数量变化 [J], 陈法军;张珏锋;陈建明;郑许松;陈列忠;俞晓平2.褐飞虱不同致害性种群体内共生菌 18S rDNA部分序列比较 [J], 张珏锋;吕仲贤;陈法军;陈建明;郑许松;徐红星;陈列忠;俞晓平3.不同虫源和致害性的褐飞虱体内共生菌的种群动态 [J], 吕仲贤;俞晓平;陈建明;郑许松;徐红星4.不同褐飞虱寄主种群类酵母共生菌形态和数量的比较 [J], 孙佳音;傅强;赖凤香;王渭霞5.高温处理后褐飞虱体内共生酵母菌和氨基酸需求的变化 [J], 傅强;张志涛;胡萃;赖凤香因版权原因,仅展示原文概要,查看原文内容请购买。
褐飞虱体内酵母类共生菌的形态观察

褐飞虱体内酵母类共生菌的形态观察陈法军;张珏锋;夏湛恩;吕仲贤;俞晓平【期刊名称】《动物分类学报》【年(卷),期】2006(031)001【摘要】试验采用冷冻切片技术结合显微摄像系统观察研究了褐飞虱雌成虫体内酵母类共生菌的形态特性和在寄主虫体内的存在状态.光学显微镜观察证实,褐飞虱雌成虫的头部和胸部均未观察到共生菌,在其腹部脂肪体中却发现大量酵母类共生菌的存在,且有大量的菌胞出现.菌胞已经成为褐飞虱特有的细胞器,其发育可划分为发育初期、共生菌增殖适应期、对数增殖期(高峰期)、释放期和衰竭期5个阶段.按照形态划分,褐飞虱体内的酵母类共生菌有4大类型,即长梭形、杆状、卵形和球状.其中,长梭形和杆状个体最多、占绝大多数,卵形和球状个体很少见.此外,还发现具假膈梭形和不规则共生菌的存在.同时观察到该类共生菌通过多边芽殖进行无性繁殖,且以顶端芽殖(包括两端芽殖)为主.【总页数】8页(P55-62)【作者】陈法军;张珏锋;夏湛恩;吕仲贤;俞晓平【作者单位】浙江省农业科学院植物保护与微生物研究所杭州 310021;浙江大学农业与生物技术学院杭州 310029;浙江省农业科学院植物保护与微生物研究所杭州 310021;浙江省农业科学院植物保护与微生物研究所杭州 310021;浙江省农业科学院植物保护与微生物研究所杭州 310021;浙江省农业科学院植物保护与微生物研究所杭州 310021【正文语种】中文【中图分类】Q964【相关文献】1.水稻与褐飞虱互作过程中虫体内类酵母共生菌的个体大小及数量变化 [J], 陈法军;张珏锋;陈建明;郑许松;陈列忠;俞晓平2.褐飞虱类酵母共生菌菌体蛋白质提取方法的比较 [J], 张海强;陈建明;张珏锋3.褐飞虱类酵母共生菌菌体蛋白质提取方法的比较 [J], 张海强;陈建明;张珏锋;4.褐飞虱体内类酵母共生菌与氨基酸营养的关系 [J], 王国超;傅强;赖凤香;陈铭学;牟仁祥;张志涛5.类酵母共生菌中两个组氨酸合成基因在褐飞虱发育中的作用 [J], 唐耀华;万品俊;郝培应;傅强;俞晓平因版权原因,仅展示原文概要,查看原文内容请购买。
一株褐飞虱内共生菌的分离及分子鉴定

Ke r s y wo d :br wn p a t o e ; c l u e i v t o;y a t l e y b ot s;Ya r w i i l c o l n h pp r u t r n i r e s — i s m i e k r o a l po yi a;s r i io at n;m o e u a tan s l i o lc lr
维普资讯
中 国水 稻 科 学 ( h ns c c) O 7 1 5 :5 ~ 5 4 C iee RieS i ,2 O ,2 ( ) 5 1 5 J
h t f www. ie c. n tp f www. i s i c . r tห้องสมุดไป่ตู้ f r sic lht : f c rc ce e o g e n
( Z e i n o etU ie st L n n3 1 0 h ja g F rs n v ri y, i ' 1 3 0,C ia I si t f P a t oe t na d Mir boo y, h ja gAc d my o a h n ; n tt e l n t i n co ilg Z e i n u o Pr c o a e fAg iu t r l rc l a u S in e ,Ha g h u3 0 2 ce cs n z o j 0 1,C ia C r ep n i ga t o , - i : x @c l . d . n hn; o r s o d n u h r E mal y p j u e u c )
c) n
A t a n I o a e r m o a t p e n t o e ul rI n iia i n S r i s l t d f o Br wn Pl n ho p ra d Is M l c a de tfc to
稻飞虱体内的类酵母共生菌及其营养功能3

与该类昆虫的营养需求有密切关系≈ ∗ ∀ 稻
普遍存在共生菌 ∀ 由于此类昆虫所吸食的植物 韧皮部汁液的营养很不均衡 一般认为共生菌
半翅目昆虫以吸食植物的汁液为食 体内
关键词 稻飞虱 类酵母共生菌 营养
系提供了有利条件 可以从基因水平证明二者的相互依赖性 ∀
蛋白质和固醇类等营养物质 ∀ 现代分子生物学技术的发展为深入研究稻飞虱和类酵母共生菌的相互关
∏2≤
ı 3
王国超 傅 33 强 张志涛
中国水稻研究所 水稻生物学国家重点实验室 杭州
稻飞虱体内的类酵母共生菌及其营养功能 3
专论与综述
昆虫知识 ≤
∏
∞
#
#
以及在传递过程中包被物的变化有关 其详细
向最小方向进化≈ ∀ 而飞虱体内的共生酵母
虫的免疫系统破坏 可能与 ≠ ≥ 自身的膜结构
说明 Βυχηνερα 为适应共生生活方式 其基因组
年≈ ∀
胞包被的合胞体中的 ≠ ≥ 通过胞吐作用释放
≠ ≥ 在飞虱体内的生存史应该不到
万
雌虫体内 ≠ ≥ 的传递过程为 存在于脂肪体细
病原物分化出来后进一步分化出 ≠ ≥ 因此
果 在稻飞虱的精囊和受精囊中均未发现 ≠ ≥
测 Χορδψχεπσ 可能是在 万年前从昆虫真菌
Βυχηνερα 的 传 递 方 式 相 同≈ ∀ 显 微 观 察 结
顺利通过昆虫的血腔成功传递给后代而不被昆
一般的细菌中基因组最小的为
胞≈ ∀ 在 ≠ ≥ 的垂直传播过程中 ≠ ≥ 何以
向 种蚜虫体内共生菌的基因组最小的仅有
每个 卵 的 后 端 会 清 晰 的 看 到 一 个 圆 形 的 菌
Βυχηνερα 的基因组研究结果即符合这一进化方
一株褐飞虱内共生菌的分离鉴定

一株褐飞虱内共生菌的分离鉴定陈鹏宇;梁靖皎;裴亚贺;王小兰【期刊名称】《广东农业科学》【年(卷),期】2014(041)009【摘要】为了解水稻褐飞虱中肠内共生菌的种类,用高氏1号和马铃薯葡萄糖琼脂两种培养基对褐飞虱肠道进行分离培养,对分离纯化后的菌株用菌类16S rDNA的通用引物进行PCR扩增鉴定,测序结果经BLAST比对分析,发现其与粉虱科的杀雄菌属内共生菌(Arsenophonus sp.)相似度达99%,说明所分离得到的为杀雄菌属的菌株.将所得这条序列与7条来自粉虱科不同种及来自褐飞虱的杀雄菌属内共生菌的16S rDNA序列用邻接法构建进化关系树,发现其与来自褐飞虱杀雄菌属内共生菌的16S rDNA (GU124505)距离较近,初步推断为同一种.【总页数】6页(P159-164)【作者】陈鹏宇;梁靖皎;裴亚贺;王小兰【作者单位】广州大学生命科学学院,广东广州510006;广州大学生命科学学院,广东广州510006;广州大学生命科学学院,广东广州510006;广州大学生命科学学院,广东广州510006【正文语种】中文【中图分类】Q933【相关文献】1.褐飞虱类酵母共生菌菌体蛋白质提取方法的比较 [J], 张海强;陈建明;张珏锋2.共生菌Arsenophonus、水稻品种和温度对褐飞虱黄绿绿僵菌发病率的影响 [J], 朱欢欢;陈洋;万品俊;王渭霞;赖凤香;傅强3.类酵母共生菌中两个组氨酸合成基因在褐飞虱发育中的作用 [J], 唐耀华;万品俊;郝培应;傅强;俞晓平4.一株褐飞虱内共生菌的分离及分子鉴定 [J], 张珏锋;吴鸿;陈建明;郑许松;陈列忠;俞晓平5.褐飞虱体内共生菌多样性研究进展 [J], 申屠旭萍;史嘉腾;宋阳;俞晓平因版权原因,仅展示原文概要,查看原文内容请购买。
【浙江省自然科学基金】_褐飞虱_期刊发文热词逐年推荐_20140811
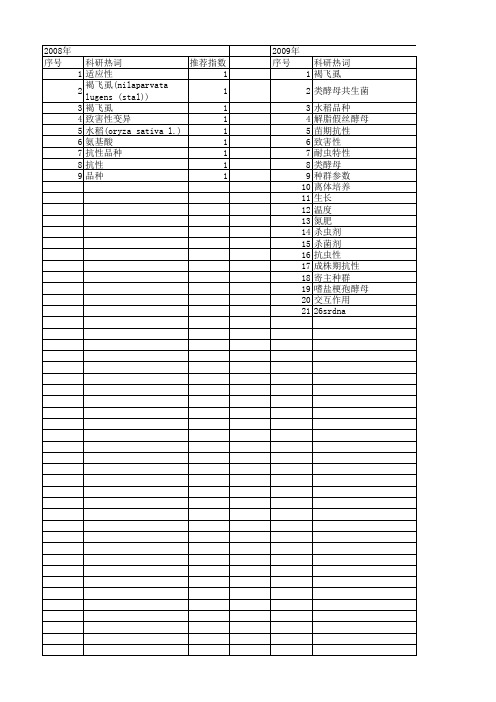
科研热词 褐飞虱 类酵母共生菌 水稻品种 解脂假丝酵母 苗期抗性 致害性 耐虫特性 类酵母 种群参数 离体培养 生长 温度 氮肥 杀虫剂 杀菌剂 抗虫性 成株期抗性 寄主种群 嗜盐梗孢酵母 交互作用 26srdna
推荐指数 6 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9
科研热词 推荐指数 适应性 1 褐飞虱(nilaparvata lugens (stal)) 1 褐飞虱 1 致害性变异 1 水稻(oryza sativa l.) 1 氨基酸 1 抗性品种 1 抗性 1 品种 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
2011年 科研热词 褐飞虱 取食行为 解脂假丝菌母 褐飞虱共生菌 苦杏仁酸 离体培养 水稻品种 敏感菌株 揭飞虱 抗药性菌株驯化 抗生性 抗氧化酶活性 抗感吡虫啉褐飞虱 抗性表现 抗性机制 抗吡虫啉菌株 吡虫啉 刺探电位图谱 刺探电位图 不同抗虫品种 推荐指数 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2013年 序号 1 2 3 4 5 6
科研热词 褐飞虱 表达谱 表皮蛋白 荧光定量pcr 基因克隆 rna干扰
推荐指数 2 2 2 2 2 2
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14
科研热词 褐飞虱 酵母类共生菌 系统发育分析 稻飞虱 白背飞虱 灰飞虱 抑制消减杂交 干旱 基因克隆 吡虫啉 取食行为 产卵选择性 its-5.8s rdna 18srdna
褐飞虱体内类酵母共生菌

血细胞计数器上分离的褐飞虱的YLS
Part
3
褐飞虱与YLS的关系
褐飞虱与YLS的营养关系
由于此类昆虫以吸食植物的汁液为食,而 植物汁液的营养很不均衡,因此在长期进化 过程中,昆虫与其体内共生菌形成互惠互利, 相互依存的关系。 昆虫为共生菌提供其生长环境和生长所需 的营养,而共生菌为昆虫提供一些生长发育 所必须的物质,如氨基酸,甾醇类物质 人为地减少虫体内共生菌的数量,会导致 宿主昆虫生长发育延迟、存活率和羽化率降 低、羽化后成虫体重减轻、产卵量下降等不 良反应
褐飞虱体内类酵母共生菌 的垂直传播机制
汇报人:肖尹
2018.5.29
目录
1 2 3
褐飞虱 类酵母共生菌(YLS) 褐飞虱与YLS的营养关系
4
YLS在褐飞虱体内的传播
Part 1 褐飞虱
褐飞虱
中文名 英文名 拉丁学名 褐飞虱 Brown rice planthopper Nilaparvata lugens(Stal)
(e)在卵巢管柄附近的卵母细胞后端形成共生球
YLS在褐飞虱体内的传播
卵内发育:
在产卵后,在宿主雌性后代的胚胎发育过程
中,共生球与每个卵中的胚胎移动,最后在
胚胎发育的后期终止于胚胎的后部。此时, YLS 被限制在腹部脂肪体内,完成垂直传播 的过程。
THANK YOU FOR YOUR ATTENTION!
卵巢侵染 (即细胞内吞作用 ; Endocytosis):
褐飞虱卵巢基本结构 褐飞虱端滋式卵巢
YLS在褐飞虱体内的传播
卵巢侵染 (即细胞内吞作用 ; Endocytosis):
这些共生菌在血腔中是自由的,游 离的YLSs感染位于卵巢管末端的 上皮塞(epithelial plug) ,通过上皮 细胞内吞进入初级卵母细胞(末端 卵母细胞)( ocyte )。
类酵母共生菌中两个组氨酸合成基因在褐飞虱发育中的作用

n a; ∗C o r r e s o n d i n u t h o r, E G m a i l: u i a n a a sc . n; x l ue . d uc . n) p ga f q g@c y p@c j
,WANP ,HAO P , T AN GY a o h u a i n u n e i i n e ta l . R o l e so f t w og e n e s i n v o l v e di nh i s t i d i n eb i o s n t h e t i cp a t h w a n j y g y yi ) , ( ) : e a s t G l i k es m b i o n t i nd e v e l o m e n to fN i l a a r v a t al u e n s( S t å l . C h i nJR i c eS c i 2 0 1 6, 3 0 4 4 0 6 G 4 1 6. y y p p g : A b s t r a c t N i l a a r v a t al u e n si sas e r i o u sp h l o e m G f e e d i n e s to fr i c ei nC h i n a . B a s e do ng e n o m ea n dt r a n s c r i t o m e gp p p g , d a t ao fN . l u e n sa n dy e a s t G l i k es m b i o n t( Y L S, a l s on a m e dE n t o m o m c e sd e l h a c i d i c o l as t r . N L U) t h em a o r y j g y p , , b i o s n t h e s i sp a t h w a s f o r a m i n oa c i d s i nN l . u e n sw e r e c o n s t r u c t e d . I n t h i s s t u d w e c l o n e d t w og e n e s E d e H i s 2a n d y y y g , E d e H i s 6,w h i c hc a t a l z ec r i t i c a l s t e s i nh i s t i d i n eb i o s n t h e s i sp a t h w a a n dr e v e a l e dt h en e a t i v ee f f e c t so fd o u b l e G y p y y g ,d s t r a n d e dR NA ( d s R NA)o nt h eg r o w t h e v e l o m e n ta n ds u r v i v a lr a t eo fN. l u e n s.H o m o l o e a r c h e sa n d p g ys g , h l o e n e t i ca n a l s i s s h o w e d t h a tE d e H i s 2a n dE d e H i s 6o r i i n f r o mY L Sg e n o m e s h a r eh i hs i m i l a r i t i e sw i t h t h a t o f p y g y g g ,w M e t a r h i z i u ma c r i d u ma n df o r m e dac l a d i nt h ep h l o e n e t i ct r e e h e r e a sn oH i s 2 Go rH i s 6 G l i k eg e n e sw a sf o u n di n y g Nl . u e n sg e n o m e . T e m o r a l e x r e s s i o np r o f i l e so fE d e H i s 2a n dE d e H i s 6s h o w e dt h a tb o t hg e n e sw e r eu b i u i t o u s l p p q y g , b u tu n e v e n l x r e s s e da m o n h ed i f f e r e n tl i f es t a e s a n dt h es a t i a le x r e s s i o np a t t e r ns h o w e dt h e a v eh i h e r ye p gt g p p yh g , , , e x r e s s i o n l e v e l s i nt h e f a tb o d a t h e r t h a nh e a d l e i n t e u m e n t a n dm i d u t . F u r t h e r m o r e n o t a r e tp r o d u c t sw e r e p yr g g g g , a m l i f i e d i nh e a da n dw i n e n o m i cD NAo fN . l u e n s, r a t h e r t h a nt h a t i na b d o m e ng e n o m i cD NA. A t t w o f o u ra n d p gg g , s i xd a sa f t e rd s E d e H i s 2o rd s E d e H i s 6i n e c t i o n t h emR NAa b u n d a n c eo f t a r e t e n e sw a sd e c r e a s e db 5%-6 0% y j g g y4 ( , E d e H i s 2) o r2 7% -5 5% ( E d e H i s 6) c o m a r i n i t hb l a n kc o n t r o l . D o w n G r e u l a t i o no fE d e H i s 2o rd s E d e H i s 6 p gw g s l i h t l i n c r e a s e dt h em o r t a l i t 3% o r9. 2% , a n dd e l a e dt h en m h a ld u r a t i o no fm a l ea n df e m a l eb 4 3a n d g y yb y8. y y p y0. , , 0 ������ 6 5d a n d0. 3 3a n d0. 3 6d a r e s e c t i v e l . M o r e o v e r b o t hm a l e( 1 1% ) a n df e m a l ea d u l t( 1 3% ) s h o w e dw i n ya y p y g , d e f o r m a t i o na f t e r i n e c t i o no f d s E d e H i s 6, h i h e r t h a nt h a t i nt h eb l a n kc o n t r o l . I nc o n c l u s i o n E d e H i s 2a n dE d e H i s 6 j g , , t h a to r i i nf r o mY L Sw e r ei n v o l v e di nh i s t i d i n eb i o s n t h e s i sp a t h w a c o n t r i b u t e dt ot h es u r v i v o r d e v e l o m e n ta n d g y y p w i n o r m a t i o no fN . l u e n s. gf g : ; ;H K e o r d s N i l a a r v a t al u e n s; e a s t G l i k es m b i o n t h i s t i d i n e i s 2;H i s 6; R NA i y y p g yw
褐飞虱不同致害性种群体内共生菌 18S rDNA部分序列比较

褐飞虱不同致害性种群体内共生菌 18S rDNA部分序列比较张珏锋;吕仲贤;陈法军;陈建明;郑许松;徐红星;陈列忠;俞晓平【期刊名称】《昆虫学报》【年(卷),期】2006(49)3【摘要】分离纯化了褐飞虱3种不同致害性种群体内类酵母共生菌(yeast-like symbionts,YLS),并对其18S rDNA基因序列进行了比较.结果表明,褐飞虱3种不同致害性种群体内类酵母共生菌18S rDNA均扩增出600 bp左右的片断.依据获得的18S rDNA特异性序列,结合已知真菌的18S rDNA部分序列,构建了不同宿主的YLS分子系统树.结果显示,褐飞虱3种不同致害性种群体内的YLS同属子囊菌亚门(Ascomycotina)的核菌纲(Pyrenomycetes),并与此纲中的Hypomyces chrysospermus亲缘关系相对最近.【总页数】5页(P528-532)【作者】张珏锋;吕仲贤;陈法军;陈建明;郑许松;徐红星;陈列忠;俞晓平【作者单位】浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021;浙江省农业科学院植物保护与微生物研究所,杭州,310021【正文语种】中文【中图分类】Q969【相关文献】1.褐飞虱内共生菌的分离及其26S rDNA部分序列分析 [J], 张珏锋;陈建明;陈法军;郑许松;陈列忠;俞晓平2.不同致害性褐飞虱种群的DNA多态性研究 [J], 关秀杰;傅强;王桂荣;赖凤香;张志涛3.不同虫源和致害性的褐飞虱体内共生菌的种群动态 [J], 吕仲贤;俞晓平;陈建明;郑许松;徐红星4.不同致害性褐飞虱种群刺吸电位图(EPG)的比较 [J], 潘建红;陈峰;何佳春;赖风香;傅强5.不同褐飞虱寄主种群类酵母共生菌形态和数量的比较 [J], 孙佳音;傅强;赖凤香;王渭霞因版权原因,仅展示原文概要,查看原文内容请购买。
高温处理后褐飞虱体内共生酵母菌和氨基酸需求的变化

高温处理后褐飞虱体内共生酵母菌和氨基酸需求的变化
傅强;张志涛;胡萃;赖凤香
【期刊名称】《昆虫学报》
【年(卷),期】2001(044)004
【摘要】对卵期高温(35℃,72 h)处理后褐飞虱Nilaparvata lugens体内共生酵母菌及该虫氨基酸需求的变化进行了研究,结果表明:1)卵期高温处理后,若虫期体内的共生酵母菌数量明显低于正常对照试虫;2)将若虫饲养于缺少单种氨基酸的22种全纯饲料上,以若虫羽化率为指标,发现高温处理试虫对组氨酸、赖氨酸、亮氨酸、异亮氨酸、苏氨酸、缬氨酸、苯丙氨酸和色氨酸等8种必需氨基酸的需求明显增加.由此推论,褐飞虱对上述8种必需氨基酸的需求与共生酵母菌有关.
【总页数】7页(P534-540)
【作者】傅强;张志涛;胡萃;赖凤香
【作者单位】中国水稻研究所;浙江大学;中国水稻研究所,;浙江大学,;中国水稻研究所
【正文语种】中文
【中图分类】Q968
【相关文献】
1.褐飞虱体内细菌型共生菌的分布 [J], 唐明;徐小蓉;洪鲲;乙引
2.水稻与褐飞虱互作过程中虫体内类酵母共生菌的个体大小及数量变化 [J], 陈法军;张珏锋;陈建明;郑许松;陈列忠;俞晓平
3.褐飞虱体内类酵母共生菌与氨基酸营养的关系 [J], 王国超;傅强;赖凤香;陈铭学;牟仁祥;张志涛
4.褐飞虱体内共生菌多样性研究进展 [J], 申屠旭萍;史嘉腾;宋阳;俞晓平
5.不同地理种群褐飞虱体内共生酵母菌的个体大小及数量差异 [J], 陈法军;张珏峰;周彦铨;叶恭银;吕仲贤;俞晓平
因版权原因,仅展示原文概要,查看原文内容请购买。
昆虫体内共生菌初探

昆虫体内共生菌初探概要:昆虫共生菌已经成为当前昆虫学领域研究的热点之一。
随着分子生物学、基因组学、转录组学和蛋白质组学的快速发展,越来越多有关共生菌文献的发表,科研工作者可以利用各种手段不断探索共生菌的种类、功能和作用,进而扩展到与寄主昆虫的相互作用模型和分子機制,及共生菌之间的相互作用等。
昆虫在食用(饲用、食用、药用)、环境治理、工业资源、观赏、土壤改良等方面发挥着极其重要的作用。
研究表明,昆虫在实现以上功能时都有体内的微生物的参与。
昆虫体内含有各种共生微生物,这些微生物在一般存在于昆虫肠道中、特殊的器官中或者细胞内。
这些微生物几乎参与了宿主所有的生命活动,一旦昆虫体内感染共生菌,会产生新的生物学性状,这些新的性状在昆虫的适应、进化和多样性方面发挥了巨大作用。
一般来说,共生菌与昆虫生长发育繁殖息息相关,经过长时间的进化,与昆虫形成了互利共生的关系。
共生菌可为宿主提供必要的营养,使其对环境耐受影响其交配、繁殖、代谢、免疫等功能,还可以介导解毒昆虫取食,增强宿主自身防御病原微生物、寄生物的能力,保护宿主逃避天敌,避免被寄生或捕食,甚至可以赋予宿主对杀虫剂产生抗性。
共生菌与宿主的互利共生为昆虫的生存带来便利,同时昆虫共生菌在开发害虫新型生物防治、废弃物生物降解和虫媒传染病的阻断控制手段中具有重要的应用前景,还可以利用昆虫与共生菌的关系来进行病虫害防控。
近年來,昆虫与共生菌的关系受到越来越多的关注,随着高通量测序工具和分析软件的发展,使得越来越多的共生菌功能被明确,这些都在为共生菌——宿主——植物的互作研究奠定基础。
本文简要概述了昆虫共生菌的研究进展,重点围绕昆虫共生菌的多样性、生物学功能、与宿主互作机制,以及共生菌——昆虫——植物互作研究进行综述,并对未来研究提出展望。
1昆虫共生菌1.1 昆虫共生菌的起源、分布和种类内共生菌是由一个自由的有机体演变而来,其结构及功能与线粒体、叶绿体相似,与昆虫的共生关系可追溯到2~2.5亿年前,在宿主昆虫体内逐渐进化成一个类细胞器的结构,含有与宿主完全不同的核酸、蛋白质合成系统,且不受宿主免疫系统的抵御,正是因为这种长期的协同进化过程,才最终形成了稳定的、可遗传的共生系统。
稻飞虱体内的类酵母共生菌及其营养功能

稻飞虱体内的类酵母共生菌及其营养功能王国超;傅强;张志涛【期刊名称】《昆虫知识》【年(卷),期】2005(42)4【摘要】类酵母共生菌(yeastlikesymbiotes,YLS)是稻飞虱体内的一种共生菌,存在于稻飞虱的脂肪体细胞中,出芽生殖,通过卵直接传给后代,其数量随飞虱的生长发育而增加,但对于单位鲜虫重而言,基本保持恒定;系统进化研究结果显示,YLS在分类上属于子囊菌亚门(Ascomycotina)、核菌纲(Pyrenomycetes),其基因组大小与自由生活的酵母菌类似;共生体系中YLS可能为宿主提供必需氨基酸、蛋白质和固醇类等营养物质。
现代分子生物学技术的发展为深入研究稻飞虱和类酵母共生菌的相互关系提供了有利条件,可以从基因水平证明二者的相互依赖性。
【总页数】5页(P353-357)【关键词】稻飞虱;类酵母共生菌;营养;共生菌;酵母菌;营养功能;体内;现代分子生物学技术;基因组大小;子囊菌亚门【作者】王国超;傅强;张志涛【作者单位】中国水稻研究所水稻生物学国家重点实验室【正文语种】中文【中图分类】Q949.326.1;S435.112.3【相关文献】1.灰飞虱体内类酵母共生菌异常毕赤酵母的分离培养及其对杀菌剂的敏感性 [J], 曹伟;马正;俞晓平2.三种稻飞虱体内类酵母共生菌18S rDNA和ITS-5.8S rDNA序列克隆及进化分析 [J], 周岩岩;董胜张;白旭;俞晓平3.褐飞虱体内类酵母共生菌与氨基酸营养的关系 [J], 王国超;傅强;赖凤香;陈铭学;牟仁祥;张志涛4.三种稻飞虱成虫体内酵母类共生菌的形态差异 [J], 陈法军;曾敏;张珏锋;陈列忠;吕仲贤;叶恭银;俞晓平5.稻飞虱酵母类胞内共生菌的组织学研究进展 [J], 陈法军;张珏锋;俞晓平因版权原因,仅展示原文概要,查看原文内容请购买。
共生菌在褐飞虱防治中的应用

Ke r s s mb o e ; l p r a a l g n t l ma a e n ; p l ai n y wo d : y it s Ni a v t u e s S  ̄ ; n g me t a p i to a c
共生菌( m it ) s boe 是昆虫体内与昆虫在进化过程中长 存在于 褐飞虱体 内腹部脂 肪体 中I ,并在 寄主脂肪体 y s 5 】
期 共 生 的微 生 物 ,它 在 寄 主 的生 长 、繁殖 过 程 中起 着 十 细 胞 中 形成 其特 有 的器 官 一 菌 胞 ( y eo ye]6 M c tc t) .。褐 】 分 重要 的作 用 。褐 飞虱 ( i p raalgn t1 内 N l av t u e sSh) 飞 虱 共 生 菌 长 约 1 m ,表 面 有 一 层 厚 的细 胞 壁 , 其 a 体 5
在 的问题 等 , 旨在 为深 入开 展 “ 抑菌 防 虫 ”研 究 提供 参 考 。
关 键 词 :共 生 菌 ;褐 飞 虱 ;防 治 ;应 用 中图分类号 :T 5 . Q4 0 1 文献标识码 :A
文章编号 :1 0 - 4 ( 0 6 1 - 7 60 6 0 1 2 0 )10 2 -4 0 3
存在类酵母共生菌( es l esm i e ,简称 Y S[ 。 中以长 梭形的个体 最多 ,杆状 类型其 次 ,而 卵形 共生 Y ati y bo s —k t L ). I
近年 来 ,对褐 飞 虱 共生 菌 的 研究 越 来 越 引起 国 内外 的重 菌 最 少 i。共 生菌 营芽 孢生 殖 ,并 以 卵母 细 胞垂 直 传递 引 视 。有 研 究 表明 ,共 生菌 是 褐 飞虱 正 常 生 长和 繁 殖所 必 的方 式 直接 传给 子 代 。褐 飞 虱体 内共 生 菌 的数 量 并 不 需 的 ,并 且共 生 菌 在 褐 飞虱 适 应环 境 、抵抗 杀 虫 剂 产生 稳定 ,正常条件下 ,褐飞虱体 内共生菌的数量随虫体的
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
褐飞虱与YLS的营养关系
(一)提供必需氨基酸 补偿机制
(二)合成胆固醇 麦角甾-5,7,24(28)-三烯甘油酯(ergosta-5,7,24(28)-trienol)
(三)参与飞虱体内的氮素循环 高活性的尿酸酶
(四)卵黄蛋白的合成 胚胎发育
Part
4
YLS在褐飞虱体内的传播
YLS在褐飞虱体内的传播
雄虫阳基侧突似蟹钳状,顶 部呈尖角状向内前方突出
褐飞虱
若虫分5龄
褐飞虱为不完全变态昆虫,生活史分3期: 卵—幼期—成虫。
不完全变态昆虫的幼虫被称为若虫(nymph)
褐飞虱
卵多产叶鞘中央肥厚部分和叶片组织内,排 成一条,称为“卵条”。 每头雌虫一般产卵300—700粒。 卵粒香蕉型,长约1mm,宽0.22mm。 卵帽顶端圆弧,清晰可数。 初产时乳白色,渐变淡黄至锈褐色,并出现 红色眼点。
褐飞虱
具有远距离迁飞习性,是我国和许多亚洲 国家当前水稻上的首要害虫。
作为单食性害虫,只能在水稻和普通野生 稻上取食和繁殖后代
①直接吸食为害 ②产卵为害 ③传播或诱发水稻病害
Part
2 类酵母共生菌 Yeast-like Symbiote
类酵母共生菌
褐飞虱体内普遍存在类酵母共生菌(Yeast-like Symbiotes, YLS) ,与酵母菌极其相似,属 于真菌类。类酵母共生菌属于子囊菌亚门核菌纲(Pyrenomycetes),假丝酵母属 (Candida)。 YLS 主要存在于飞虱腹部脂肪体内或者卵内,通过卵垂直传递给下一代,出芽生殖。YLS 的长度一般在10-15μm之间.宽度3-6μm。不同的飞虱种类中,YLS的形状和大小有所不 同。
血细胞计数器上分离的褐飞虱的YLS
Part
3
褐飞虱与YLS的关系
褐飞虱与YLS的营养关系
由于此类昆虫以吸食植物的汁液为食,而 植物汁液的营养很不均衡,因此在长期进化 过程中,昆虫与其体内共生菌形成互惠互利, 相互依存的关系。 昆虫为共生菌提供其生长环境和生长所需 的营养,而共生菌为昆虫提供一些生长发育 所必须的物质,如氨基酸,甾醇类物质 人为地减少虫体内共生菌的数量,会导致 宿主昆虫生长发育延迟、存活率和羽化率降 低、羽化后成虫体重减轻、产卵量下降等不 良反应
界 纲 目 亚目 科
动物界 昆虫纲 半翅目 同翅亚目 飞虱科 (Homoptera:Delphacidae)
褐飞虱
成虫:长翅型和短翅型
触角 头 胸 足 腹 前翅
长翅型 体长3.6~4.8毫米
短翅型 体长2.5~4.0毫米 前翅伸达腹部第5~6节, 后翅均退化
雌虫产卵器基部两侧,第1载瓣片 的内缘基部突起呈半圆形。
YLS在褐飞虱体内的传播
卵巢侵染 (即细胞内吞作用 ; Endocytosis):
(B)终末卵母细胞(terminal oocyte)和 倒数第二卵母细胞(penultimate oocyte ) 之间的上皮组织中的YLS。末端卵母细胞含 有卵黄并形成卵壳
YLS在褐飞虱体内的传播
一旦进入卵母细胞,YLS可能 在其后端聚集,在卵母细胞的 后部形成共生球。YLS松散地 分布在共生球中,并且它们被 膜状结构包围
卵巢侵染 (即细胞内吞作用 ; Endocytosis):
褐飞虱卵巢基本结构 褐飞虱端滋式卵巢
YLS在褐飞虱体内的传播
卵巢侵染 (即细胞内吞作用 ; Endocytosis):
这些共生菌在血腔中是自由的,游 离的YLSs感染位于卵巢管末端的 上皮塞(epithelial plug) ,通过上皮 细胞内吞进入初级卵母细胞(末端 卵母细胞)( ocyte )。
YLS在褐飞虱体内的传播
脂肪体内增殖:YLSs位于腹部脂肪体中的菌胞(mycetocyte)。成虫出 现48h后,YLS的数量大大增加,并且填满了脂肪体的内部。成虫出现96h后, 完整的含胞体(syncytia)形成时,更多的YLSs分布在含胞体中,并被卵巢管 附近的一层脂肪体细胞所包围
(D)雌性成虫中的菌胞和脂肪体细胞
褐飞虱体内类酵母共生菌 的垂直传播机制
汇报人:肖尹
2018.5.29
目录
1 2 3
褐飞虱 类酵母共生菌(YLS) 褐飞虱与YLS的营养关系
பைடு நூலகம்
4
YLS在褐飞虱体内的传播
Part 1 褐飞虱
褐飞虱
中文名 英文名 拉丁学名 褐飞虱 Brown rice planthopper Nilaparvata lugens(Stal)
类酵母共生菌在寄主褐飞虱腹部脂肪体细胞
中形成其特有的器官 ———菌胞 ( Mycetocyte) 羽化后两三天,菌胞似乎形成含胞体 ( syncytium )(含有由一层细胞膜包绕的多
个核的一团细胞质,这通常是由于发生了细胞
融合或一系列不完全细胞分裂周期所致),并 被认为是可分解的。
褐飞虱雌成虫腹部脂肪体内的菌胞
(e)在卵巢管柄附近的卵母细胞后端形成共生球
YLS在褐飞虱体内的传播
卵内发育:
在产卵后,在宿主雌性后代的胚胎发育过程
中,共生球与每个卵中的胚胎移动,最后在
胚胎发育的后期终止于胚胎的后部。此时, YLS 被限制在腹部脂肪体内,完成垂直传播 的过程。
THANK YOU FOR YOUR ATTENTION!
褐飞虱体内YLS由亲代在虫体内通过卵巢垂直 传递给正在发育的卵,没有在体外停留的机会。雌虫
体内YLS的侵染过程 :脂肪体内增殖、 菌胞释放和
含胞体胞外分泌 ( Exocytosis) 、 卵巢侵染 (即细胞 内吞作用 ; Endocytosis) 和卵内发育 4 个阶段。
YLS在褐飞虱体内的传播
脂肪体内增殖
(d)羽化后96小时,形成良好的含胞体
YLS在褐飞虱体内的传播
菌胞释放和含胞体胞外分泌 ( Exocytosis)
一些共生菌从含胞体的边界离开,通过胞吐作用 ( Exocytosis )释放到血腔( hemocoel )内。
(b)通过胞吐作用离开含胞体边界的共生菌(箭头)
YLS在褐飞虱体内的传播
褐飞虱与YLS的营养关系
宿主昆虫对其共生菌在生理和营养上存在依赖性:
1、共生菌和宿主昆虫氮素营养的关系主要表现在三个方面:一 是合成和提供必需氨基酸供宿主昆虫利用;二是合成蛋白质;三 是利用氮素代谢废物(NH3和尿酸等) 2、参与脂类物质的合成:1)固醇类物质的合成;2)脂肪酸的 合成 3、维生素的合成 4、提供能源