23-代谢调控
色氨酸2,3-双加氧酶调控炎症免疫反应在疾病中的作用

色氨酸2,3 双加氧酶调控炎症免疫反应在疾病中的作用李素素 常 艳△ 魏 伟△(安徽医科大学临床药理研究所,抗炎免疫药物教育部重点实验室,抗炎免疫药物安徽省协同创新中心,合肥230032)摘要 犬尿氨酸代谢通路(kynureninepathway,KP)是色氨酸代谢的主要途径,参与机体多个病理生理过程。
色氨酸2,3 双加氧酶(tryptophan2,3 dioxygenase,TDO2)是KP的初始限速酶之一,能够催化色氨酸(tryptophan,Trp)使其转化为犬尿氨酸(kynurenine,Kyn)及下游代谢产物。
多项研究表明,TDO2介导的KP代谢异常参与的炎症免疫反应调控在炎症免疫相关性疾病、肿瘤、中枢神经系统疾病中发挥重要作用。
本文综述了TDO2在炎症免疫调控中的作用及其介导的KP紊乱参与多种疾病病理机制的研究进展和药物研究现状。
关键词 色氨酸 2,3 双加氧酶;炎症;免疫;色氨酸;犬尿氨酸中图分类号 R332;R341;R345;R364;R392;R977;R59 色氨酸(tryptophan,Trp)是机体的必需氨基酸之一,主要参与能量代谢、蛋白质合成和产生活性代谢产物,介导多种生理功能。
Trp主要有两条代谢途径:犬尿氨酸(kynurenine,Kyn)和5 羟色胺(5 hydroxytryptpamine,5 HT)途径,其中95%Trp通过Kyn途径代谢,称为犬尿氨酸代谢通路(kynureninepathway,KP)[1]。
Trp在吲哚胺 2,3 双加氧酶1(indoleamine2,3 dioxygenase1,IDO1)、色氨酸 2,3 双加氧酶2(tryptophan 2,3 dioxygenase2,TDO2)和吲哚胺 2,3 双加氧酶2(indoleamine2,3 dioxyge nase2,IDO2)三个限速酶的催化下生成重要中间代谢产物Kyn和一系列下游代谢产物,包括3 羟基犬尿氨酸(3 Hydroxykynurenine,3 HK)、3 羟邻氨苯甲酸(3 hydroxyanthranilicacid,3 HAA)、犬尿喹啉酸(kynyrenicacid,KYNA)、邻氨基苯甲酸(anthra nilicacid,AA)等。
生物发酵工程中的微生物代谢调控

生物发酵工程中的微生物代谢调控生物发酵工程是一门应用广泛的学科,其涉及微生物、生物化学、传热传质等多个学科的知识,旨在利用微生物在发酵过程中生成的代谢产物来满足人们的生产和生活需求。
而在微生物代谢过程中,如何通过调控其代谢途径,实现目标产物的高效合成、减少代谢的损耗,成为了生物发酵工程中的一个重要研究方向。
微生物代谢调控的意义微生物代谢调控是指通过改变微生物生长条件和代谢途径,来调整其代谢产物合成量和比例的过程。
在工业生产中,通过微生物发酵生产大量特定代谢产物是一种非常经济、环保的加工方式,但是微生物产生的代谢产物存在很大的不确定性。
因此,通过对微生物代谢途径的调控,可以使目标产物的产量和比例得到控制,从而提高产量,减少生产成本,并且提高产物的纯度和质量。
微生物代谢途径及其调控微生物代谢途径是指微生物在生物体内通过多种化学反应途径来完成与生长、繁殖相关的代谢活动。
其中包括代谢途径的产物合成、产物分解以及代谢的能量转换等步骤。
微生物代谢调控的目的就是通过改变这些代谢途径的流量和平衡,来调整微生物产物的合成量和比例,从而达到优化目标的效果。
代表微生物代谢调控方法有以下两种:1.正向调控正向调控是指通过增加代谢途径的流量和互动,来加强目标代谢产物的合成,从而提高产量。
这种调控方法常见于工业生产中,例如酿造啤酒时,生产者为了增加酒的口感和香味,会加入辅酶、氨基酸、谷氨酸等化合物来促进酵母细胞的代谢,从而促进目标产物的生成。
2.反向调控反向调控是指通过成分的增减或运输网络的改变来降低目标代谢产物的合成,从而对微生物代谢路线进行限制。
这种方法经常用于通过发酵生成的副产物的控制。
例如,在环境中限制糖分含量、压制某些酶或基因,以限制微生物代谢途径中产生某些不必要的代谢产物的方法。
代表性工程案例在实践中,通过微生物代谢调控技术,目标代谢产物的生产得以实现。
以下列举一些代表性的工程案例:1.习得味道细节啤酒发酵后的酿造品质是由微生物代谢途径的复杂模式决定的。
生物化学 第13章 代谢调节

第十三章代谢调节一、填空题:1.生物体内的代谢调节在三种不同的水平上进行,即、和。
2.代谢途径的终产物浓度可以控制自身形成的速度,这种现象被称为。
3.酶对细胞代谢的调节是最基本的代谢调节,主要有二种方式:和。
构通糖、脂代谢的关键化合物是。
4.不同代谢途径可以通过交叉点代谢中间物进行转化,在糖、脂、蛋白质及核酸的相互转化过程中三个最关键的代谢中间物是、和。
5.1961年,法国生物学家Monod和Jacob提出了关于原核生物基因结构及表达调控的学说。
6.正调控和负调控是基因表达的两种最基本的调节形式,其中原核细胞常用调控,而真核细胞常用调控模式。
7.乳糖操纵子的天然诱导物是,实验室里常用作为乳糖操纵子的安慰诱导物诱导β-半乳糖苷酶的产生。
8.许多代谢途径的第一个酶是限速酶,终产物多是它的,对它进行,底物多为其。
9.原核细胞酶的合成速率主要在水平进行调节。
10.乳糖操纵子的诱导物是,色氨酸操纵子的辅阻遏物是。
二、选择题(只有一个最佳答案):1.下列与能量代谢有关的过程除哪个外都发生在线粒体中?()A、糖酵解B、三羧酸循环C、脂肪酸的β-氧化D、氧化磷酸化2.IPTG可以诱导乳糖操纵子(lacOperon)的表达,这是因为:()A、IPTG与乳糖操作子(lacoperator)结合,诱导转录B、IPTG与LACI基因产物结合,并抑制其活性C、抑制β-半乳糖苷酶的活性D、促进Lac阻遏物的活性E、IPTG与LACI基因产物结合,并激活其活性3.在什么情况下,乳糖操纵子的转录活性最高?()A、高乳糖,低葡萄糖B、高乳糖,高葡萄糖C、低乳糖,低葡萄糖D、低乳糖,高葡萄糖4.真核细胞参与基因表达调节的调控区比原核细胞复杂是因为()A、真核细胞的细胞核具有双层膜B、原核细胞的基因总是以操纵子的形式存在C、原核细胞调节基因表达主要是在翻译水平D、真核细胞需要控制细胞特异性的基因表达E、真核细胞基因组含有太多的重复序列5.调节物质代谢体内最基础的层次是()A、细胞水平B、激素水平C、神经调节D、整体水平E、器官水平6.磷酸果糖激酶是什么代谢途径中的别构调节酶()A、三羧酸循环B、糖异生C、葡萄糖分解D、糖原合成E、糖原分解7.三羧酸循环中的别构调节酶是()A、柠檬酸合成酶B、α-酮戊二酸脱氢酶C、琥珀酸脱氢酶D、延胡索酸酶E、苹果酸脱氢酶8.催化糖酵解与磷酸戊糖途径的酶主要分布在细胞中什么部位()A、核B、胞质C、线粒体D、微粒体E、质膜9.催化三羧酸循环与脂肪酸β-氧化的酶分布在细胞内的什么部位()A、胞质B、胞膜C、胞核D、内质网E、线粒体10.氨基酸分解代谢调节的别构酶是()A、转氨酶B、脱羧酶C、转甲基酶D、己糖激酶E、谷氨酸脱氨酶11.糖异生限速酶的别构调节激活剂是()A、A TPB、ADPC、AMPD、dA TPE、cAMP 12.各种分解途径中,放能最多的途径是:()A、糖酵解B、三羧酸循环C、 -氧化D、氧化脱氨基13.操纵子调节系统属于哪一种水平的调节?()A、复制水平的调节B、转录水平的调节C、转录后加工的调节D、翻译水平的调节14.下列关于操纵基因的论述哪个是正确的?()A、能专一性地与阻遏蛋白结合B、是RNA聚合酶识别和结合的部位C、是诱导物和辅阻遏物的结合部位D、能于结构基因一起转录但未被翻译15.以下有关阻遏蛋白的论述哪个是正确的?()A、阻遏蛋白是调节基因表达的产物B、阻遏蛋白妨碍RNA聚合酶与启动子结合C、阻遏蛋白RNA聚合酶结合而抑制转录D、阻遏蛋白与启动子结合而阻碍转录的启动16.糖酵解中,下列哪一个催化的反应不是限速反应?()A、丙酮酸激酶B、磷酸果糖激酶C、己糖激酶D、磷酸丙糖异构酶17.磷酸化酶通过接受或脱去磷酸基而调节活性,因此它属于:()A、别(变)构调节酶B、共价调节酶C、诱导酶D、同工酶18.下列与能量代谢有关的途径不在线粒体内进行的是:()A、三羧酸循环B、脂肪酸β氧化C、氧化磷酸化D、糖酵解作用19.关于共价修饰调节酶,下列哪种说法是错误的?()A、这类酶一般存在活性和无活性两种形式,B、酶的这两种形式通过酶促的共价修饰相互转变C、伴有级联放大作用D、是高等生物独有的代谢调节方式20.阻遏蛋白结合的位点是:()A、调节基因B、启动因子C、操纵基因D、结构基因21.下面哪一项代谢是在细胞质内进行的:()A、脂肪酸的β-氧化B、氧化磷酸化C、脂肪酸的合成D、TCA22.在乳糖操纵子模型中,操纵基因专门控制是否转录与翻译。
成纤维细胞生长因子23与肾脏磷代谢研究新进展

加 尿磷 的排 泄直接 影 响血 清 中磷 的水 平 ,参与钙 磷
代 谢 。它 们 包括 成 纤维 细 胞 生 长 因 子 2 3
体调节 钙磷 代谢 的主要 激素 。生理状 态 下 ,血 钙 降 低 会 刺激 甲状 旁腺 主细 胞 上钙 敏感 性 受体 活 化 , 促
进 P H生 成和 分 泌增 多 。P H主要 作 用于 肾脏 和 骨 T T
磷的排泄直接 影响血清 中磷 的水平 ,这一发现为磷代谢 的研 究提供 了新 的视 角 。成纤维细 胞生长因子 2 3 (i r b a t g o t a t r 2 , G 一 3 就是 目前研究最多的一个 。循环 F F 2 f b o l s r w h f c o 一 3 F F 2 ) G 一 3水平过高或 过低都会 引 起 机体磷平衡的改变,引发一系列病变 。血 F F 2 水平过 高患者表现 为低 磷血症、 1 2 0 )D 生成减 G一 3 , 5(a 23
的磷 稳态 [。正常 的肾功 能是保 证 P H V t 轴 有效 1 ] T - iD 作用 的基础 , 肾功 能损 害会从 多个途 径影 响机 体钙
磷代 谢 。
和 2%的氨 基酸 相似性 。FF 2 白含 有 2 1 2 G 一3蛋 5 个氨
基 酸 ,相 对 分子质 量为 3 0 ,除含 有 F F家族 同 20 0 G 源 性结 构域 N 端外 ,还包 含一 个 由 7 个氨 基酸 组成 1
(i r b a t g o t a t r 2 , F F 2 ) f b o l s r w h f c o 一 3 G 一 3 、分 泌
型 曲连 蛋 白 4 s c e e r Z l d r l t d p o (er t d f iZ e e a e r —
氨基酸感知与代谢调控的研究进展

动物营养学报2015,27(2):342⁃351ChineseJournalofAnimalNutrition㊀doi:10.3969/j.issn.1006⁃267x.2015.02.003氨基酸感知与代谢调控的研究进展许丹丹㊀何㊀艮∗(中国海洋大学水产学院,水产动物营养与饲料农业部重点实验室,海水养殖教育部重点实验室,青岛266003)摘㊀要:氨基酸不仅是蛋白质和其他含氮化合物合成的重要前体,还参与体内主要代谢途径的调控㊂当氨基酸不足时,机体内多种机制参与调节体内平衡,包括快速停止蛋白质合成㊁增加氨基酸合成和转运,以及加强自噬作用㊂越来越多的学者证明氨基酸可作为信号分子参与细胞内信号传导过程,可以调节其他营养素如脂肪和能量的代谢,最终导致机体整体代谢的改变㊂本文主要综述细胞内氨基酸的营养感知与应答机制,涉及氨基酸应答(AAR)和雷帕霉素靶蛋白(TOR)2条信号转导通路,并探讨这2条信号通路对下游营养素代谢途径的调节㊂关键词:氨基酸感知;氨基酸应答;雷帕霉素靶蛋白;代谢中图分类号:Q493;S968.1㊀㊀㊀㊀文献标识码:A㊀㊀㊀㊀文章编号:1006⁃267X(2015)02⁃0342⁃10收稿日期:2014-08-13基金项目:国家自然科学基金(31222055);国家973计划(2014CB138602)作者简介:许丹丹(1986 ),女,山东枣庄人,博士研究生,从事水产动物蛋白质代谢研究㊂E⁃mail:dandxu2008@163.com∗通信作者:何㊀艮,教授,博士生导师,E⁃mail:hegen@ouc.edu.cn㊀㊀当环境发生大范围改变,如盐度㊁酸碱度或者营养素等发生变化,多细胞生物体本身的内稳态机制能够保证其存活㊂对高等动物的研究表明,氨基酸代谢库中氨基酸的组成与含量变化引起不同的细胞应答,由相关信号传导系统介导,调控下游效应因子,实现蛋白质的合成与降解㊁基因表达与抑制以及营养素的新陈代谢,最终在宏观上表现为动物生长发育等经济性状的差别[1]㊂目前已知在细胞水平上有2条信号通路可感知氨基酸含量,1条为感知氨基酸丰度的雷帕霉素靶蛋白(tar⁃getofrapamycin,TOR)信号系统,1条为感知氨基酸平衡的氨基酸应答(aminoacidresponse,AAR)信号通路[1-2]㊂1㊀TOR信号通路的氨基酸感知与代谢调节1.1㊀TOR信号通路与氨基酸感知㊀㊀TOR是一种高度保守的丝氨酸/苏氨酸激酶,它接收并整合来自细胞内的氨基酸㊁生长因子㊁能量状态和应激等信号,调节细胞的生长和代谢[3-4]㊂TOR存在2个结构和功能上有差异的复合物,分别是对雷帕霉素和营养素敏感的TOR复合体(TORcomplex,TORC)1和对雷帕霉素和营养素不敏感的TORC2[5-6]㊂在哺乳动物细胞中,TORC1主要包含有TOR㊁TOR调节相关蛋白(regulatory⁃associatedproteinofmTOR,Raptor)和哺乳动物酵母同源致命因子Sec13蛋白8(mam⁃malianlethalwithSEC13protein8,mLST8)成分,而TORC2主要包含有TOR㊁对雷帕霉素不敏感的伴随物(rapamycin⁃insensitivecompanionofmTOR,Rictor)㊁哺乳动物应激激活蛋白激酶作用蛋白(mSIN1)㊁mLST8和富含脯氨酸蛋白(prolinerichprotein,PRR)成分[7]㊂TORC1可以综合细胞内氨基酸㊁生长因子㊁应激等信号,进而控制着多种主要的细胞生命活动,例如蛋白质合成㊁脂肪合成㊁能量代谢和自噬等过程[4,8]㊂TORC2主要由生长因子激活,进而通过磷脂酰肌醇-3-激酶(phos⁃phoinositide⁃3⁃kinase,PI3K)依赖的核糖体辅助和磷酸化蛋白激酶A㊁G㊁C相关(AGC)激酶家族,进而控制着细胞的存活㊁肌动蛋白细胞骨架的组织和其他生命过程[8-10]㊂㊀㊀对TORC1激活的机制已在哺乳动物㊁畜禽和2期许丹丹等:氨基酸感知与代谢调控的研究进展鱼类中进行了广泛研究[5,8,11]㊂其一,胰岛素可以激活TORC1的活性㊂胰岛素或类胰岛素生长因子(IGFs)首先与细胞膜表面的受体结合,使胰岛素受体底物1(insulinreceptorsubstrate1,IRS1)磷酸化,继而使PI3K在细胞膜聚集㊂PI3K通过磷酸肌醇依赖激酶1(PDK1)磷酸化蛋白激酶B(proteinkinaseB,Akt)㊂Akt可通过2种途径激活TORC1:磷酸化的结节性硬化症复合体(TSC)2使TSC1/TSC2复合物去抑制,通过脑组织中丰富表达的Ras同源类似物(Rashomologenrichedinbrain,Rheb)鸟苷三磷酸酶(GTPase)激活TORC1;Akt也可通过磷酸化富含脯氨酸Akt/PKB亚基40(PRAS40)激活TORC1[12]㊂其二,氨基酸可以激活TORC1的活性㊂氨基酸信号如何传递至TORC1目前还知之甚少,但是一些重要的信号传递蛋白已逐渐被发现[13]㊂氨基酸感知一个至关重要的步骤是由氨基酸招募TORC1至溶酶体膜上,其中Rheb对TORC1附着在溶酶体膜上起至关重要的作用[14],对所有可以激活TORC1的信号来讲都是必需的[15]㊂㊀㊀然而,TSC2敲除的小鼠胚胎成纤维细胞(MEFs)仍然可以感知氨基酸的缺乏,意味着存在其他非TSC⁃Rheb途径的复合物参与到氨基酸感知的过程中㊂TORC1在溶酶体上的激活模型已被广泛接受,其受氨基酸含量调节,涉及多个蛋白酶复合体,包括RagGTPase㊁调节子(regulator)和空泡H+⁃ATP酶(v⁃ATPase)㊂RagGTPase共有4个亚基,分别是RagA㊁RagB㊁RagC和RagD,其中通常RagA和RagB㊁RagC和RagD聚集组成没有功能的二聚体,它们通常常驻在溶酶体膜上㊂Rag复合物与鸟苷三磷酸(GTP)或鸟苷二磷酸(GDP)的结合是受氨基酸调控的㊂细胞经过氨基酸刺激之后结合核苷酸时开始翻转,RagA/B和RagC/D分别与GTP和GDP结合,这是一种Rag二聚体活化的状态㊂当氨基酸充足时,RagA/B㊃GTP⁃RagC/D㊃GDP激活复合体与TORC1复合体的Raptor相互作用而成为停靠位点,使TORC1停靠在溶酶体的表面㊂当处于氨基酸缺乏状态时,TORC1分散在整个细胞中,而当加入氨基酸刺激后,TORC1重新分布在含有溶酶体和晚期核内体标志蛋白溶酶体相关膜蛋白(LAMP2)和小GTPase酶结合蛋白7(RAB7)的小泡中㊂这意味着RagGTPase可以将氨基酸信号传导到TORC1[14-15]㊂RagGTPase定位在溶酶体表面,需要一个锚定分子,而regulator多聚复合物就是介导二者结合的骨架,其是由有丝分裂原激活蛋白激酶衔接蛋白1(MP1)㊁衔接蛋白14(p14)和衔接蛋白18(p18)组成的三聚体㊂当氨基酸改变时,溶酶体膜上的v⁃ATPase调节regulator的氨基酸应答反应㊂高度保守的v⁃ATPase由2个组分组成,包括V1结构域和V0结构域㊂V1结构域的ATP水解介导细胞质中的质子通过V0结构域的通道泵入溶酶体中㊂V1结构域与RagGTPase和regulator相互作用,其作用受氨基酸调节,当氨基酸缺乏时其作用加强,当氨基酸充足时作用减弱㊂V1结构域的ATP水解对于RagGTPase与TORC1的相互作用是必需的,从而激活TORC1㊂无论是果蝇还是哺乳动物细胞中,氨基酸均引起溶酶体膜上v⁃ATPase与RagGTPase和regulator相互作用,从而调节TORC1的活力[16]㊂㊀㊀饲料高蛋白质㊁氨基酸和胰岛素均能激活TOR信号系统,影响虹鳟的多种代谢过程[17]㊂氨基酸对动物TOR信号通路调控作用的研究发现,去除组织培养液中的氨基酸1 2h,显著抑制了TOR下游信号应答蛋白核糖体蛋白S6激酶(ribo⁃somalproteinS6kinases,S6Ks)和真核起始因子4E结合蛋白(eukaryotictranslationinitiationfactor4E⁃bindingproteins,4E⁃BPs)磷酸化作用;氨基酸恢复至基础水平,恢复了下游信号传导物的活性或磷酸化作用[18]㊂在对啮齿动物[19-20]和人类[21-22]的体内体外研究发现,氨基酸种类对TOR信号传导的调控作用存在差异,亮氨酸和精氨酸的调控作用较强,特别是亮氨酸㊂1.2㊀TOR信号通路控制蛋白质合成㊀㊀TORC1调控着整体的翻译水平,进而促进细胞生长和生命延长㊂TORC1的下游蛋白包括4E⁃BP1和S6K1,它们以磷酸化的方式调控着mRNA多方面的翻译功能[8]㊂蛋白质合成的限速步骤是翻译起始,在这个过程中小核糖体亚基聚集到mRNA5ᶄ端,识别起始密码子,随后完整的核糖体也聚集其上完成翻译[23-24]㊂小核糖体亚基与mRNA的结合需要真核细胞转录起始因子(eu⁃karyotictranslationinitiationfactor,eIF)4F复合体结合到mRNA5ᶄ帽子结构㊂eIF4F复合体包含3个起始因子,即eIF4E㊁eIF4G和eIF4A,其中eIF4E与5ᶄ帽子结构结合是组装eIF4F复合体的关键,它343㊀动㊀物㊀营㊀养㊀学㊀报27卷能够召集eIF4G和eIF4A的结合[23]㊂而4E⁃BP1与eIF4E结合抑制了eIF4F复合体形成㊂当TOR信号通路激活后,磷酸化4E⁃BP1使eIF4E与之分离,eIF4E募集eIF4G和eIF4A,起始蛋白质的合成[25]㊂㊀㊀TORC1的另一个重要应答因子是S6Ks,由TORC1磷酸化激活㊂研究表明,S6Ks在蛋白质翻译中调节翻译起始过程,且协同调节核糖体的生物合成,从而使翻译更加高效[26]㊂S6K1有多个效应因子,S6K1的磷酸化可以磷酸化下游核糖体蛋白(S6),转录起始因子eIF4B,抑制胰岛素受体底物-1(IRS⁃1)㊁凋亡蛋白(BAD)以及对真核细胞延伸因子2(eukaryoticelongationfactor2,eEF2)有抑制作用的eEF2激酶(eEF2K)等,从而调节蛋白质合成㊁糖脂代谢㊁细胞大小和分裂以及细胞存活等细胞过程[27-28]㊂S6是第1个被鉴别的S6K1底物,同样,S6的磷酸化作用也是被研究的最彻底的一个㊂S6有5个磷酸化位点,分别为苏氨酸35㊁236㊁240㊁244和247位点,其中苏氨酸236位点为最重要的磷酸化位点[27]㊂S6磷酸化促进了核糖体和5ᶄ端寡嘧啶束(5ᶄ⁃terminaloligopyrimidinetract,5ᶄ⁃TOP)mRNA,的亲和力,从而诱导5ᶄTOPmRNA进行有效翻译[29-30]㊂5ᶄTOPmRNA主要编码核糖体蛋白和其他翻译过程的必需蛋白[31]㊂㊀㊀蛋白质合成代谢主要受TOR信号通路调节,而蛋白质分解则主要是通过泛素-蛋白酶体通路调控[32-33]㊂然而,鱼类蛋白酶体的活性对营养状况敏感与否仍有待验证[34-35]㊂近年来,国外学者也针对鱼类肌肉组织中的蛋白质降解和特定氨基酸对TOR信号途径的激活状态开展了探索性研究[11]㊂研究发现,饲粮氨基酸含量是激活TOR系统的有效途径,比如通过添加谷氨酰胺来激活TOR信号通路,可提高生长性能[36]㊂而蛋白质摄入不足或氨基酸不平衡会导致IGF⁃1水平下降,而IGF结合蛋白1(IGFBP⁃1)水平升高,共同作用于TOR信号通路,引起生长抑制[37-38]㊂1.3㊀TOR信号通路与糖脂代谢调控㊀㊀尽管目前对于TOR和代谢调控的研究尚处于初级阶段,但已有不少结果显示TORC1可以在很多组织器官中,从转录㊁翻译和翻译后水平上调控多条代谢通路[3]㊂在淋巴瘤细胞中,雷帕霉素抑制TORC1活性改变了许多代谢酶基因的表达,结合代谢图谱和基因表达结果分析显示,在培养的细胞中TORC1调控着糖酵解㊁甾醇和脂质的合成,另外,磷酸戊糖途径也受TORC1的调控[39-40]㊂大量报道显示TORC1可以激活转录因子固醇调节元件结合蛋白1(sterolregulatoryelementbindingprotein⁃1,SREBP⁃1)㊂SREBP⁃1是转录因子固醇调节元件结合蛋白家族(SREBPs)的一员,调控着脂肪合成相关酶的表达,以及脂肪和胆固醇的稳态㊂完整的SREBP⁃1主要存在于内质网膜上,在各种刺激物的作用下,如降低的甾醇㊁胰岛素或者饱和脂肪酸水平,使SREBP⁃1从内质网运送到高尔基体,在高尔基体内SREBP⁃1前体被激活㊂被激活后的SREBP⁃1进而被转运到细胞核内,在细胞核内激活和促进甾醇调节原件和相关基因表达[41]㊂TORC1通过促进SREBP⁃1转录㊁翻译和翻译后的加工,使其活性提高,从而诱导甾醇和脂肪的合成,以及磷酸戊糖途径相关基因的转录[39]㊂由TORC1上调的SREBP⁃1活性对于由Akt磷酸化引起的脂肪合成是必需的[42]㊂㊀㊀TORC1也可以促进缺氧诱导因子(hypoxiainduciblefactor,HIF)基因表达,这是通过4E⁃BP1依赖的方式调控其翻译[43-44]㊂HIF可激活100200个涉及到细胞内代谢和细胞适应低氧环境的基因的转录,且TORC1依赖性的HIF的激活足以上调这些基因的表达[39]㊂HIF⁃1对糖酵解途径的激活被认为是低氧代谢适应的关键,通过增加葡萄糖转化为丙酮酸盐进而分解为乳酸来实现㊂HIF⁃1也可以间接激活丙酮酸脱氢酶激酶(PDK1),通过三羧酸循环(TCA)来有效抑制代谢过程[45]㊂当营养素变化后,TORC1通过调节HIF和SREBP⁃1的活性,从而使机体能量代谢和脂肪合成代谢发生适应性改变㊂TORC1另一重要功能是,可以调控线粒体数量和功能㊂在小鼠骨骼肌中,缺乏Raptor导致调控线粒体生物合成酶的表达量降低,且氧化能力损失,Cunningham等[46]发现TORC1促进了转录因子PPARγ联合激活因子1a(PGC1a)的转录活性,其在调节线粒体合成和氧化代谢方面起重要作用㊂另外,目前对于TORC1信号如何影响和调节转录网络的了解仍然很少,仍需要大量的工作㊂㊀㊀与TORC1受抑制时对相关基因表达的调节一致,S6K1或4E⁃BPs突变的小鼠表现出极大的代谢改变㊂S6K1缺失的小鼠由于降低了β细胞的数量显现出低胰岛素症状,但是同时也升高了4432期许丹丹等:氨基酸感知与代谢调控的研究进展对胰岛素信号的敏感性[47]㊂另外,S6K1缺失的小鼠能够抵抗由饮食和年龄引起的肥胖[48],尽管摄食量未受影响,S6K1的缺失通过增加甘油三酯的分解阻止了脂肪在脂肪组织的富集,并增加了脂肪酸在脂肪组织和肌肉中的氧化㊂而4E⁃BP1和4E⁃BP2缺失的小鼠表现出相反的表观症状,尽管这些小鼠都正常存活,但与正常野生型小鼠相比在16周的研究时间内,它们明显肥胖且体重增加30%,这些小鼠脂肪组织的重量显著升高,且血浆中的胰岛素和胆固醇的水平升高,进而建立起研究肥胖的模型㊂㊀㊀大量研究证明,TOR信号系统与氨基酸应答通路在从酵母到人的不同生物中功能保守,TOR信号通路上的大部分信号蛋白,如TOR㊁S6K1㊁S6和4E⁃BP1,都已经在虹鳟上证明是保守性存在的[11,17]㊂体内和体外试验表明胰岛素[11]㊁氨基酸[49]㊁胰岛素和氨基酸[50]㊁植物蛋白质源替代[51]以及碳水化合物和蛋白质比例不同的饲粮[17,52]均显著影响TOR信号通路和肝脏中的次级代谢相关的基因表达㊂简要来讲,TOR信号通路和其他相关因子的响应状况与畜禽和其他高等动物中的研究结果一致㊂在斑马鱼中的研究结果表明,餐后TOR通路被激活,饲喂显著上调了餐后糖酵解基因[葡萄糖激酶(GK)㊁己糖激酶(HK1)]和脂肪合成基因[脂肪酸合成酶(FAS)㊁葡萄糖-6-磷酸脱氢酶(G6PD)㊁乙酰辅酶A羧化酶α(ACCα)]的表达,显著抑制了斑马鱼肝胰脏糖异生基因[磷酸烯醇式丙酮酸羧激酶(cPEPCK)]和脂肪分解途径关键酶基因[肉碱棕榈酰基转移酶1b(CPT1b)]的表达[53]㊂而腹腔注射雷帕霉素后,显著下调虹鳟脂肪合成和糖酵解途径关键酶基因的表达[54]㊂这表明氨基酸和胰岛素依赖的TOR通路的激活或抑制调控鱼体糖脂代谢过程㊂2㊀AAR信号通路的氨基酸感知与代谢调节2.1㊀AAR信号通路与氨基酸应答㊀㊀蛋白质缺乏或必需氨基酸缺乏将激活氨基酸应答通路[55]㊂一般来说,酵母转录激活因子(GCN)2作为氨基酸感知传感器,通过与非负载tRNA结合从而感知细胞内氨基酸含量,当GCN2与任一非负载转运RNA(tRNA)结合均能激活GCN2,从而使AAR信号通路激活㊂当氨基酸缺乏时,细胞内空载tRNA增多,使GCN2激酶去磷酸化被激活,进而引起eIF2α丝氨酸51位点的磷酸化㊂eIF2α的磷酸化使体内大部分蛋白质的合成减少,但也会通过转录水平调控另一些基因的表达㊂转录激活因子(activatingtranscriptionfac⁃tor,ATF)4就是其中之一,当氨基酸缺乏时上调其表达量㊂ATF4与CCAAT增强子结合蛋白(CCAAT/enhancer⁃bindingprotein,C/EBP)形成二聚体,与C/EBP激活转录因子应答元件(C/EBPactivatingtranscriptionfactorresponseelements,CARE)结合,激活大量下游基因转录,包括氨基酸转运载体㊁氨基酸代谢酶㊁氧化状态调节因子㊁能量调节因子等,从而调控着细胞内的氨基酸缺乏应答[56]㊂研究发现,在人肝癌细胞系(HepG2)细胞培养液中任何一种必需氨基酸缺乏都可以激活AAR信号通路[57]㊂另外,eIF2α的磷酸化也是会提高其他转录调节因子水平,包括转录因子ATF5以及生长停滞与DNA损害可诱导基因34(GADD34)㊂㊀㊀最重要的是当必需氨基酸缺乏时,ATF4可以诱导氨基酸转运载体和氨基酸合成酶基因的表达,从而促进氨基酸的合成与吸收,保证正常的生理机能㊂ATF4还可上调其他基因转录,如C/EBPβ㊁ATF3和C/EBP同源蛋白(CHOP),这些可以作为ATF4的负调节因子[56,58]㊂在动物中,饲喂低蛋白质的饲粮会增加C/EBPmRNA表达量和活性[59]㊂细胞感知多种胁迫信号均可上调ATF3的表达[60]㊂Pan等[61]和Jiang等[62]发现ATF3基因的表达量受氨基酸应答和UPR通路的上调,这些调节分别需要eIF2α激酶GCN2和蛋白激酶R样内质网激酶(PERK)㊂在GCN2敲除[62]和ATF4敲除[63]的纤维母细胞中发现,ATF3mRNA表达量显著降低㊂在营养缺乏情况下,AAR信号通路的激活对癌细胞存活和增殖有至关重要的作用[64]㊂关于ATF4调控的下游目的基因及其具体功能在Kilberg等[56]的综述中有详尽的描述㊂另外,受AAR调节的下游基因4E⁃BP1和发育和DNA损伤应答调节基因1(regulatedindevelop⁃mentandDNAdamageresponse1,REDD1)均为TOR信号应答通路的负调节因子,且2条信号通路均可通过调节ATF4的表达来而调控合成代谢,说明在营养状态改变时,2条信号通路共同作出应答使机体处于最优生长状态[65-67]㊂543㊀动㊀物㊀营㊀养㊀学㊀报27卷2.2㊀AAR信号通路与代谢调控㊀㊀氨基酸的不平衡不仅可以调控氨基酸代谢,而且会对糖脂代谢造成显著影响㊂当必需氨基酸缺乏时,GCN2是蛋白质和脂肪代谢过程中关键的代谢调节因子[68-69]㊂氨基酸缺乏会激活AAR信号通路,导致细胞内大多数蛋白质的翻译起始受抑制,从而抑制体内大部分蛋白质的翻译[63]㊂GCN2作为氨基酸缺乏时的感受器能够感知任何一种必需氨基酸的缺乏,eIF2α磷酸化抑制细胞内大多数蛋白质的翻译起始,从而降低整体蛋白质合成㊂因此,GCN2能够确保维持生长和细胞功能的最低氨基酸量;同时,eIF2α可以调节特异基因的翻译㊂在酵母中研究发现,转录因子GCN4的翻译可促进一系列基因的表达,包括所有的氨基酸合成酶㊁氨基酸转运载体,以使酵母抵抗氨基酸缺乏应激[70]㊂AAR信号通路可以保证足够的氨基酸前体的供应,从而使机体在必需氨基酸缺乏时维持关键蛋白的合成㊂同酵母一致,哺乳动物对氨基酸缺乏也有一套应答机制[71],包括提高氨基酸合成酶和氨基酸转运载体的表达量,同时降低整体蛋白质的合成㊂敲除GCN2基因的小鼠当饲喂氨基酸平衡的饲粮时可以正常存活,但当饲喂氨基酸不平衡的饲粮时发育受损㊂敲除GCN2的细胞,在氨基酸缺乏状态下,不能使eIF2α磷酸化[67,72]㊂㊀㊀除调节蛋白质的翻译外,AAR信号通路还可调节机体脂肪代谢㊂Guo等[68]发现饲粮缺乏亮氨酸后影响了小鼠脂质代谢过程:当亮氨酸缺乏时,小鼠肝脏和脂肪组织中甘油三酯的合成受抑制;但敲除GCN2基因的小鼠摄食亮氨酸缺乏的饲粮后肝脏脂肪合成升高,且出现脂肪肝,同时,小鼠脂肪组织中的脂肪氧化降解降低;然而,继续敲除转录因子固醇调节元件结合蛋白1c(sterolregula⁃toryelementbindingprotein⁃1c,SREBP⁃1c)后肝脏甘油三酯的积累降低,与同窝野生型小鼠差异不显著㊂在HepG2细胞中的研究结果与之一致,当细胞培养液中缺乏亮氨酸时抑制了脂肪的合成[57]㊂另外,敲除ATF4基因的小鼠能够抵抗高脂饮食诱导肥胖与脂肪肝[73]㊂给小鼠饲喂赖氨酸和苏氨酸缺乏的小麦面筋会引起胆固醇合成的降低;给小鼠饲喂完全不含蛋白质的饲粮时也会导致这些基因表达量的降低[74];同时,饲喂这2种饲粮的组中小鼠血浆胆固醇含量也均降低㊂以上研究均证明饲粮必需氨基酸缺乏显著影响脂肪合成,即饲粮中氨基酸的含量会影响到体内非蛋白质类营养物质的动态平衡㊂2.3㊀AAR信号通路与摄食调控㊀㊀当以某一种必需氨基酸缺乏的饲粮饲喂动物时,这种不平衡会很快被动物体所识别并降低摄食量㊂事实上,血浆中的氨基酸缺乏是和摄食量的降低直接联系起来的,研究还发现大脑的前梨状皮质区域可以监测到蛋白质的质量或者氨基酸的平衡性[75-76]㊂氨基酸应答通路对摄食量的调控方式为:氨基酸不平衡时缺乏的氨基酸相应tRNA出现去乙酰化,激活GCN2激酶,进而磷酸化转录启动因子eIF2α,引起摄食量下调㊁抑制蛋白质合成[77]㊂GCN2有非常保守的㊁调控氨基酸平衡的功能,在酵母中它通过调控氨基酸合成而获得氨基酸平衡;在哺乳动物和畜禽中是通过调控摄食行为进行㊂在饥饿的动物中,GCN2通过抑制蛋白质合成从而减少肌肉重量[78]㊂对GCN2敲除小鼠的研究发现,大脑对氨基酸不平衡的识别需要GCN2感知细胞内空载tRNA的量[79]㊂ATF4敲除的成纤维细胞和肝癌细胞中的微阵列分析显示ATF4调控着大量的和氨基酸转运㊁代谢㊁氧化状态和能量调控相关的基因[63,80]㊂体内研究发现,给GCN2敲除小鼠饲喂亮氨酸缺乏的饲粮会导致其肌肉生长受损,但是并未引起肝脏损伤,而在野生小鼠中,亮氨酸的缺乏同时降低了肌肉和肝脏的重量[67]㊂3㊀小㊀结㊀㊀氨基酸可以作为信号传导通路的调节因子引起信号通路应答,从而调节相关蛋白的翻译与糖脂代谢过程,尽管其在哺乳动物中有较深入的研究,但是在水产营养领域中仍是一个较新的概念㊂尤其对于肉食性鱼类,氨基酸不平衡问题是限制鱼粉蛋白质源替代研究的一个主要原因,因此深入探究其机制将对鱼类饲料中非鱼粉蛋白质源的大量应用有一定的指导作用㊂另外,尽管TOR信号通路与代谢调节在鱼类中有部分研究,而AAR信号通路在鱼类营养研究中未见相关报道㊂因此氨基酸与胰岛素㊁氨基酸本身,尤其是必需氨基酸对鱼类AAR和TOR信号通路的调节作用与响应阈值,以及对糖脂代谢的调控机制有待深入研究㊂6432期许丹丹等:氨基酸感知与代谢调控的研究进展参考文献:[1]㊀GALLINETTIJ,HARPUTLUGILE,MITCHELLJR.Aminoacidsensingindietary⁃restriction⁃mediatedlongevity:rolesofsignal⁃transducingkinasesGCN2andTOR[J].BiochemicalJournal,2013,449(1):1-10.[2]㊀吕子全,郭非凡.氨基酸感应与糖脂代谢调控的研究进展[J].生命科学,2013,25(2):152-157.[3]㊀POLAKP,HALLMN.mTORandthecontrolofwholebodymetabolism[J].CurrentOpinioninCellBiology,2009,21(2):209-218.[4]㊀WULLSCHLEGGERS,LOEWITHR,HALLMN.TORsignalingingrowthandmetabolism[J].Cell,2006,124(3):471-484.[5]㊀KIMJ,GUANKL.AminoacidsignalinginTORac⁃tivation[J].AnnualReviewofBiochemistry,2011,80:1001-1032.[6]㊀LOEWITHR,JACINTOE,WULLSCHLEGERS,etal.TwoTORcomplexes,onlyoneofwhichisrapamy⁃cinsensitive,havedistinctrolesincellgrowthcontrol[J].MolecularCell,2002,10(3):457-468.[7]㊀ZINZALLAV,STRACKAD,OPPLIGERW,etal.ActivationofmTORC2byassociationwiththeribo⁃some[J].Cell,2011,144(5):757-768.[8]㊀LAPLANTEM,SABATINIDM.mTORsignalingingrowthcontrolanddisease[J].Cell,2012,149(2):274-293.[9]㊀CYBULSKIN,HALLMN.TORcomplex2:asigna⁃lingpathwayofitsown[J].TrendsinBiochemicalSciences,2009,34(12):620-627.[10]㊀HAGIWARAA,CORNUM,CYBULSKIN,etal.HepaticmTORC2activatesglycolysisandlipogenesisthroughAkt,glucokinase,andSREBP1c[J].CellMe⁃tabolism,2012,15(5):725-738.[11]㊀SEILIEZI,GABILLARDJC,SKIBA⁃CASSYS,etal.AninvivoandinvitroassessmentofTORsignalingcascadeinrainbowtrout(Oncorhynchusmykiss)[J].AmericanJournalofPhysiology:Regulatory,Integra⁃tiveandComparativePhysiology,2008,295(1):R329-R335.[12]㊀BHASKARPT,HAYN.ThetwoTORCsandAKT[J].DevelopmentalCell,2007,12(4):487-502.[13]㊀KIME,GORAKSHA⁃HICKSP,LIL,etal.Regula⁃tionofTORC1byRagGTPasesinnutrientresponse[J].NatureCellBiology,2008,10(8):935-945.[14]㊀SANCAKY,BAR⁃PELEDL,ZONCUR,etal.Ragu⁃lator⁃RagcomplextargetsmTORC1tothelysosomalsurfaceandisnecessaryforitsactivationbyaminoacids[J].Cell,2010,141(2):290-303.[15]㊀SANCAKY,PETERSONTR,SHAULYD,etal.TheRagGTPasesbindraptorandmediateaminoacidsignalingtomTORC1[J].Science,2008,320(5882):1496-1501.[16]㊀JEWELLJL,RUSSELLRC,GUANKL.AminoacidsignallingupstreamofmTOR[J].NatureRe⁃viewsMolecularCellBiology,2013,14(3):133-139.[17]㊀SEILIEZI,PANSERATS,LANSARDM,etal.Dieta⁃rycarbohydrate⁃to⁃proteinratioaffectsTORsignalingandmetabolism⁃relatedgeneexpressionintheliverandmuscleofrainbowtroutafterasinglemeal[J].A⁃mericanJournalofPhysiology:Regulatory,IntegrativeandComparativePhysiology,2011,300(3):733-743.[18]㊀HARAK,YONEZAWAK,WENGQP,etal.AminoacidsufficiencyandmTORregulatep70S6kinaseandeIF⁃4EBP1throughacommoneffectormecha⁃nism[J].JournalofBiologicalChemistry,1998,273(23):14484-14494.[19]㊀KIMBALLSR,JEFFERSONLS.Signalingpathwaysandmolecularmechanismsthroughwhichbranched⁃chainaminoacidsmediatetranslationalcontrolofpro⁃teinsynthesis[J].TheJournalofNutrition,2006,136(1):227S-231S.[20]㊀STIPANUKMH.Leucineandproteinsynthesis:mTORandbeyond[J].NutritionReviews,2007,65(3):122-129.[21]㊀邓会玲,刘国华,刘宁.氨基酸介导的TOR信号传导通路研究进展[J].动物营养学报,2011,23(4):529-535.[22]㊀DRUMMONDMJ,RASMUSSENBB.Leucine⁃en⁃richednutrientsandtheregulationofmammaliantar⁃getofrapamycinsignallingandhumanskeletalmuscleproteinsynthesis[J].CurrentOpinioninClinicalNu⁃trition&MetabolicCare,2008,11(3):222-226.[23]㊀PESTOVATV,KOLUPAEVAVG,LOMAKINIB,etal.Molecularmechanismsoftranslationinitiationineukaryotes[J].ProceedingsoftheNationalAcade⁃myofSciencesoftheUnitedStatesofAmerica,2001,98(13):7029-7036.[24]㊀GEBAUERF,HENTZEMW.Molecularmechanismsoftranslationalcontrol[J].NatureReviewsMolecularCellBiology,2004,5(10):827-835.743㊀动㊀物㊀营㊀养㊀学㊀报27卷[25]㊀GINGRASAC,KENNEDYSG,O LEARYMA,etal.4E⁃BP1,arepressorofmRNAtranslation,isphos⁃phorylatedandinactivatedbytheAkt(PKB)signa⁃lingpathway[J].GenesDevelopment,1998,12(4):502-513.[26]㊀JASTRZEBSKIK,HANNANKM,TCHOUBRIEVAEB,etal.Coordinateregulationofribosomebiogene⁃sisandfunctionbytheribosomalproteinS6kinase,akeymediatorofmTORfunction[J].GrowthFactors,2007,25(4):209-226.[27]㊀RUVINSKYI,SHARONN,LERERT,etal.Riboso⁃malproteinS6phosphorylationisadeterminantofcellsizeandglucosehomeostasis[J].GenesDevelop⁃ment,2005,19(18):2199-2211.[28]㊀RUVINSKYI,MEYUHASO.RibosomalproteinS6phosphorylation:fromproteinsynthesistocellsize[J].TrendsinBiochemicalSciences,2006,31(6):342-348.[29]㊀JEFFERIESHB,FUMAGALLIS,DENNISPB,etal.Rapamycinsuppresses5ᶄTOPmRNAtranslationthroughinhibitionofp70S6K[J].TheEMBOJour⁃nal,1997,16(12):3693-3704.[30]㊀HORNSTERNE,TANGH,MEYUHASO.MitogenicandnutritionalsignalsaretransducedintotranslationalefficiencyofTOPmRNAs[C]//Coldspringharborsymposiaonquantitativebiology.ColdSpringHarbor,NY:ColdSpringHarborLaboratoryPress,2001:477-484.[31]㊀MEYUHASO,AVNID,SHAMAS.TranslationalcontrolofribosomalproteinmRNAsineukaryotes[M].ColdSpringHarbor,NY:ColdSpringHarborLaboratoryPress,1996:363-364.[32]㊀LECKERSH,JAGOERT,GILBERTA,etal.Multi⁃pletypesofskeletalmuscleatrophyinvolveacommonprogramofchangesingeneexpression[J].TheFASEBJournal,2004,18(1):39-51.[33]㊀HOCHSTRASSERM.Ubiquitin⁃dependentproteindegradation[J].AnnualReviewofGenetics,1996,30(1):405-439.[34]㊀MARTINSAM,BLANEYS,BOWMANAB,etal.Ubiquitin⁃proteasome⁃dependentproteolysisinrain⁃bowtrout(Oncorhynchusmykiss):effectoffooddep⁃rivation[J].PflügersArchiv,2002,445(2):257-266.[35]㊀SALEMM,KENNEYPB,REXROADCE,etal.Mi⁃croarraygeneexpressionanalysisinatrophyingrain⁃bowtroutmuscle:auniquenonmammalianmuscledegradationmodel[J].PhysiologicalGenomics,2007,28(1):33-45.[36]㊀WANGY,LIK,HANH,etal.Potentialofusingablendofrenderedanimalproteiningredientstoreplacefishmealinpracticaldietsformalabargrouper(pi⁃nephelusmalabricus)[J].Aquaculture,2008,281(1):113-117.[37]㊀STRAUSDS.Nutritionalregulationofhormonesandgrowthfactorsthatcontrolmammaliangrowth[J].TheFASEBJournal,1994,8(1):6-12.[38]㊀DUANCM,RENHX,GAOS.Insulin⁃likegrowthfactors(IGFs),IGFreceptors,andIGF⁃bindingpro⁃teins:rolesinskeletalmusclegrowthanddifferentia⁃tion[J].GeneralandComparativeEndocrinology,2010,167(3):344-351.[39]㊀DUVELK,YECIESJL,MENONS,etal.ActivationofametabolicgeneregulatorynetworkdownstreamofmTORcomplex1[J].MolecularCell,2010,39(2):171-183.[40]㊀YECIESJL,MANNINGBD.TranscriptionalcontrolofcellularmetabolismbymTORsignaling[J].CancerResearch,2011,71(8):2815-2820.[41]㊀ESPENSHADEPJ,HUGHESAL.Regulationofste⁃rolsynthesisineukaryotes[J].Genetics,2007,41:401-427.[42]㊀PORSTMANNT,SANTOSCR,GRIFFITHSB,etal.SREBPactivityisregulatedbymTORC1andcon⁃tributestoAkt⁃dependentcellgrowth[J].CellMetab⁃olism,2008,8(3):224-236.[43]㊀THOMASGV,TRANC,MELLINGHOFFIK,etal.Hypoxia⁃induciblefactordeterminessensitivitytoin⁃hibitorsofmTORinkidneycancer[J].NatureMedi⁃cine,2005,12(1):122-127.[44]㊀MAJMUNDARAJ,WONGWJ,SIMONMC.Hy⁃poxia⁃induciblefactorsandtheresponsetohypoxicstress[J].MolecularCell,2010,40(2):294-309.[45]㊀KIMJW,TCHERNYSHYOVI,SEMENZAGL,etal.HIF⁃1⁃mediatedexpressionofpyruvatedehydrogen⁃asekinase:ametabolicswitchrequiredforcellularad⁃aptationtohypoxia[J].CellMetabolism,2006,3(3):177-185.[46]㊀CUNNINGHAMJT,RODGERSJT,ARLOWDH,etal.mTORcontrolsmitochondrialoxidativefunctionthroughaYY1⁃PGC⁃1[agr]transcriptionalcomplex[J].Nature,2007,450(7170):736-740.[47]㊀PENDEM,KOZMASC,JAQUETM,etal.Hypoin⁃sulinaemia,glucoseintoleranceanddiminishedβ⁃cellsizeinS6K1⁃deficientmice[J].Nature,2000,4088432期许丹丹等:氨基酸感知与代谢调控的研究进展(6815):994-997.[48]㊀UMSH,FRIGERIOF,WATANABEM,etal.Ab⁃senceofS6K1protectsagainstage⁃anddiet⁃inducedobesitywhileenhancinginsulinsensitivity[J].Nature,2004,431(7005):200-205.[49]㊀LANSARDM,PANSERATS,PLAGNES⁃JUANE,etal.L⁃leucine,L⁃methionine,andL⁃lysinearein⁃volvedintheregulationofintermediarymetabolism⁃relatedgeneexpressioninrainbowtrouthepatocytes[J].TheJournalofNutrition,2011,141(1):75-80.[50]㊀LANSARDM,PANSERATS,PLAGNES⁃JUANE,etal.Integrationofinsulinandaminoacidsignalsthatregulatehepaticmetabolism⁃relatedgeneexpressioninrainbowtrout:roleofTOR[J].AminoAcids,2010,39(3):801-810.[51]㊀LANSARDM,PANSERATS,SEILIEZI,etal.He⁃paticproteinkinaseB(Akt)⁃targetofrapamycin(TOR)⁃signallingpathwaysandintermediarymetabo⁃lisminrainbowtrout(Oncorhynchusmykiss)arenotsignificantlyaffectedbyfeedingplant⁃baseddiets[J].BritishJournalofNutrition,2009,102(11):1564-1573.[52]㊀SKIBA⁃CASSYS,PANSERATS,LARQUIERM,etal.Apparentlowabilityofliverandmuscletoadapttovariationofdietarycarbohydrate:proteinratioinrain⁃bowtrout(Oncorhynchusmykiss)[J].BritishJournalofNutrition,2013,109(8):1359-1372.[53]㊀SEILIEZI,MÉDALEF,AGUIRREP,etal.Postpran⁃dialregulationofgrowth⁃andmetabolism⁃relatedfac⁃torsinZebrafish[J].Zebrafish,2013,10(2):237-248.[54]㊀DAIWW,PANSERATS,MENNIGENJA,etal.Post⁃prandialregulationofhepaticglucokinaseandli⁃pogenesisrequirestheactivationofTORC1signallinginrainbowtrout(Oncorhynchusmykiss)[J].JournalofExperimentalBiology,2013,216(23):4483-4492.[55]㊀PADDON⁃JONESD,SHEFFIELD⁃MOOREM,ZHANGXJ,etal.Aminoacidingestionimprovesmuscleproteinsynthesisintheyoungandelderly[J].AmericanJournalofPhysiology:EndocrinologyandMetabolism,2004,286(3):E321-E328.[56]㊀KILBERGMS,SHANJX,SUN.ATF4⁃dependenttranscriptionmediatessignalingofaminoacidlimita⁃tion[J].TrendsinEndocrinologyandMetabolism,2009,20(9):436-443.[57]㊀DUDEKSM,SEMENKOVICHCF.EssentialaminoacidsregulatefattyacidsynthaseexpressionthroughanunchargedtransferRNA⁃dependentmechanism[J].JournalofBiologicalChemistry,1995,270(49):29323-29329.[58]㊀CHAVEROUXC,JOUSSEC,CHERASSEY,etal.I⁃dentificationofanovelaminoacidresponsepathwaytriggeringATF2phosphorylationinmammals[J].Mo⁃lecularandCellularBiology,2009,29(24):6515-6526.[59]㊀MARTENN,SLADEKF,STRAUSD.Effectofdiet⁃aryproteinrestrictiononlivertranscriptionfactors[J].BiochemicalJournal,1996,317(2):361-370.[60]㊀HAIT,WOLFGANGCD,MARSEEDK,etal.ATF3andstressresponses[J].GeneExpression,1999,7(4/5/6):321-335.[61]㊀PANYX,CHENH,SIUF,etal.Aminoaciddepriva⁃tionandendoplasmicreticulumstressinduceexpres⁃sionofmultipleactivatingtranscriptionfactor⁃3mR⁃NAspeciesthat,whenoverexpressedinHepG2cells,modulatetranscriptionbythehumanasparaginesyn⁃thetasepromoter[J].TheJournalofBiologicalChem⁃istry,2003,278(40):38402-38412.[62]㊀JIANGHY,WEKSA,MCGRATHBC,etal.Acti⁃vatingtranscriptionfactor3isintegraltotheeukaryot⁃icinitiationfactor2kinasestressresponse[J].Molec⁃ularandCellularBiology,2004,24(3):1365-1377.[63]㊀HARDINGHP,ZHANGY,ZENGH,etal.Aninte⁃gratedstressresponseregulatesaminoacidmetabolismandresistancetooxidativestress[J].MolecularCell,2003,11(3):619-633.[64]㊀YEJB,KUMANOVAM,HARTLS,etal.TheGCN2⁃ATF4pathwayiscriticalfortumourcellsur⁃vivalandproliferationinresponsetonutrientdepriva⁃tion[J].TheEMBOJournal,2010,29(12):2082-2096.[65]㊀KIMBALLSR,DOAND,KUTZLERL,etal.RapidturnoverofthemTORcomplex1(mTORC1)re⁃pressorREDD1andactivationofmTORC1signalingfollowinginhibitionofproteinsynthesis[J].TheJour⁃nalofBiologicalChemistry,2008,283(6):3465-3475.[66]㊀JINHO,SEOSK,WOOSH,etal.SP600125nega⁃tivelyregulatesthemammaliantargetofrapamycinviaATF4⁃inducedRedd1expression[J].FEBSLetters,2009,583(1):123-127.[67]㊀ANTHONYTG,MCDANIELBJ,BYERLEYRL,etal.Preservationofliverproteinsynthesisduringdiet⁃aryleucinedeprivationoccursattheexpenseofskele⁃943㊀动㊀物㊀营㊀养㊀学㊀报27卷talmusclemassinmicedeletedforeIF2kinaseGCN2[J].TheJournalofBiologicalChemistry,2004,279(35):36553-36561.[68]㊀GUOFF,CAVENERDR.TheGCN2eIF2αkinaseregulatesfatty⁃acidhomeostasisintheliverduringdeprivationofanessentialaminoacid[J].CellMetab⁃olism,2007,5(2):103-114.[69]㊀DEVALC,CHAVEROUXC,MAURINAC,etal.A⁃minoacidlimitationregulatestheexpressionofgenesinvolvedinseveralspecificbiologicalprocessesthroughGCN2⁃dependentandGCN2⁃independentpathways[J].TheFEBSJournal,2009,276(3):707-718.[70]㊀HINNEBUSCHAG.TranslationalregulationofGCN4andthegeneralaminoAcidcontrolofyeast[J].AnnualReviewofMicrobiology,2005,59:407-450.[71]㊀KILBERGMS,PANYX,CHENH,etal.Nutritionalcontrolofgeneexpression:howmammaliancellsre⁃spondtoaminoacidlimitation[J].AnnualReviewofNutrition,2005,25:59-85.[72]㊀ZHANGPC,MCGRATHBC,REINERTJ,etal.TheGCN2eIF2αkinaseisrequiredforadaptationtoami⁃noaciddeprivationinmice[J].MolecularandCellularBiology,2002,22(19):6681-6688.[73]㊀DESOUSA⁃COELHOAL,MARREROPF,HAROD.Activatingtranscriptionfactor4⁃dependentinduc⁃tionofFGF21duringaminoaciddeprivation[J].Bio⁃chemicalJournal,2012,443(1):165-171.[74]㊀ENDOY,FUZW,ABEK,etal.Dietaryproteinquantityandqualityaffectrathepaticgeneexpression[J].TheJournalofNutrition,2002,132(12):3632-3637.[75]㊀KOEHNLETJ,RUSSELLMC,MORINAS,etal.Dietsdeficientinindispensableaminoacidsrapidlydecreasetheconcentrationofthelimitingaminoacidintheanteriorpiriformcortexofrats[J].TheJournalofNutrition,2004,134(9):2365-2371.[76]㊀郑溜丰,彭健.中枢神经系统整合外周信号调节采食量的分子机制[J].动物营养学报,2013,25(10):2212-2221.[77]㊀MAURINAC,JOUSSEC,AVEROUSJ,etal.TheGCN2kinasebiasesfeedingbehaviortomaintainami⁃noacidhomeostasisinomnivores[J].CellMetabo⁃lism,2005,1(4):273-277.[78]㊀DEVERTE,HINNEBUSCHAG.GCN2whetstheappetiteforaminoacids[J].MolecularCell,2005,18(2):141-142.[79]㊀HAOSZ,SHARPJW,ROSS⁃INTACM,etal.Un⁃chargedtRNAandsensingofaminoaciddeficiencyinmammalianpiriformcortex[J].Science,2005,307(5716):1776-1778.[80]㊀LEEJI,DOMINYJE,Jr,SIKALIDISAK,etal.HepG2/C3Acellsrespondtocysteinedeprivationbyinductionoftheaminoaciddeprivation/integratedstressresponsepathway[J].PhysiologicalGenomics,2008,33(2):218-229.0532期许丹丹等:氨基酸感知与代谢调控的研究进展∗Correspondingauthor,professor,E⁃mail:hegen@ouc.edu.cn(责任编辑㊀菅景颖)AdvancesinAminoAcidSensingandMetabolicRegulationXUDandan㊀HEGen∗(KeyLaboratoryofAquacultureNutritionandFeedsofMinistryofAgriculture,KeyLaboratoryofMaricultureofMinistryofEducation,OceanUniversityofChina,Qingdao266003,China)Abstract:Aminoacids,consideredastheimportantbuildingblocksforthesynthesisofproteinandothernitro⁃gencompounds,haverecentlyshowntoactasmodulatorsofintracellularsignaltransductionpathways.Toa⁃dapttointracellularaminoaciddeprivation,multipleadaptivemechanismshaveevolved,includingaquickces⁃sationofnewproteinsynthesis,anincreaseinaminoacidtransportandbiosynthesis,andautophagy.Evidencehasaccumulatedthataminoacidsalsofunctionassignalmoleculewhichprofoundeffectsonregulationofcellsignalingandgeneexpression.Metabolicadaptationincludinglipidandenergymetabolismisrequiredtocopewithepisodesofproteinoraminoacidsavailableleadingtooverallmetabolicchangesinmulticellutarorgan⁃isms.Here,thecurrentknowledgeaboutthemechanismsofintracellularaminoacidsensingandresponse,in⁃volvingintheaminoacidresponse(AAR)pathwayandthetargetofrapamycin(TOR)signalingpathway,andhowthesetwopathwaysregulatedownstreamnutrientsmetabolismwerereviewed.[ChineseJournalofAnimalNutrition,2015,27(2):342⁃351]Keywords:aminoacidsensing;TOR;AAR;metabolism153。
生化试题——精选推荐

⽣化试题试卷5: ⽣物氧化、糖代谢、脂代谢姓名()学号()⼀、是⾮题(每题1分,共20题,答对给1分,答错倒扣0.5分,不答不给分,请⽤"+"和"-"分别表⽰"对"和 "错")9、在消耗ATP的情况下,电⼦可从复合体Ⅳ流动到复合体Ⅰ。
10、ATP是磷酸果糖激酶的底物,因此它的浓度越⾼,则相关的反应速度就越快。
11、由于癌细胞的⽣长和分裂⽐正常细胞要旺盛,因⽽更依赖于ATP的产⽣,这表现在它们的糖酵解速率明显减弱。
12、HMGCoA合成酶是胆固醇合成途径中的限速酶。
13、脂肪酸β氧化产⽣的⼄酰CoA可通过TCA循环⽽形成OAA,OAA可通过糖异⽣合成葡萄糖,因⽽动物细胞可通过这样的⽅式⽽将脂肪酸净转变为糖。
14、磷酸化的⼄酰CoA羧化酶才有活性。
15、可以使⽤化学渗透学说解释F1/F0-ATPase合成ATP的机制。
16、植烷酸是在体内通过α-氧化分解产⽣ATP的。
17、砷酸和亚砷酸均可以解除巴斯德效应。
18、⼄醛酸循环和三羧酸循环中都有琥珀酸的净⽣成。
19、天然的酮体包括丙酮、⼄酰⼄酸和L-β羟丁酸。
20、过氧化物酶体也能进⾏脂肪酸的β-氧化。
⼆、选择题(每题1分,共15题,每道题只有1个答案,答错不倒扣)6、苍术苷酸能够抑制氧化磷酸化是因为它直接作⽤于A、复合体ⅠB、复合体ⅡC、复合体ⅢD、复合体ⅣE、ADP/ATP交换体7、F1/F0ATPase的活性中⼼位于A、α亚基B、β亚基C、γ亚基D、δ亚基E、ε亚基8、下列哪⼀种物质最不可能通过线粒体内膜?A、PiB、苹果酸C、柠檬酸D、丙酮酸E、NADH9、将离体的线粒体放在⽆氧的环境中,经过⼀段时间以后,其内膜上的呼吸链的成分将会完全以还原形式存在,这时如果忽然通⼊氧⽓,试问最先被氧化的将是内膜上的哪⼀种复合体?A、A、复合体ⅠB、复合体ⅡC、复合体ⅢD、复合体ⅣE、复合体Ⅴ10、在离体的完整的线粒体中,在有可氧化的底物的存在下,加⼊哪⼀种物质可提⾼电⼦传递和氧⽓摄⼊量?A、更多的TCA循环的酶B、ADPC、FADH2D、NADHE、氰化物11、下列反应中哪⼀步伴随着底物⽔平的磷酸化反应?A、葡萄糖→葡萄糖-6-磷酸B、⽢油酸-1,3-⼆磷酸→⽢油酸-3-磷酸C、柠檬酸→α-酮戊⼆酸D、琥珀酸→延胡索酸E、苹果酸→草酰⼄酸12、帮助FFA进⼊线粒体的有机分⼦是(A)CoA (B)⾁毒碱 (C)硫⾟酸 (D) 磷脂 (E)α-磷酸⽢油⼆、填充题(每空1分,共35个空)9、细胞内代谢物上脱下来的氢如果直接与氧⽓结合则形成()。
第十章:发酵过程优化与放大概论

• 3. 串级反馈控制
• 由两个以上控制器对一种变量进行联合控 制的方法
• 4. 前馈/反馈控制
• 如废水处理系统,分析悬浮固体含量前馈到排 放控制器
• 根据排出悬浮固体含量对排放率进行反馈调节
• 1.2.2 发酵控制系统的硬件结构
• 1. 传感器 • 2. 变送器与过程接口 • 3. 执行机构和转换器 • 4. 监控计算机
• 2. 溶氧
• 复膜溶氧探头
• 银阴极和铅阳极组成的原电池
• 管状银阳极、铂丝阴极、氯化银电解液和极 化电源组成的极谱型
• 产生的电流正比于通过膜扩散入探头的氧量
• 复膜溶氧探头实际测量的是氧分压,与溶氧 浓度并不直接相关,结果用溶氧压(DOT) 表示
• 一般以空气中氧饱和的百分度表示
• 3. 氧化还原电位
不同水平问题的相关
Rpm ↓ DO ↓
OUR
当DO低于临界
氧浓度(CLc)时,
VHB
OUR下降,细胞代
谢由好氧向厌氧途径 迁移。
CLC
DO
分子水平 基因工程血红蛋白 hemoglobin (VHB) 对氧的亲和力提高(由图所示),临界氧下降。
2.3 发酵工程控制中二个基本问题——优化与放大
代谢调控研究 代谢工程研究
• 缺乏以细胞代谢流分析与控制为核心的
研究内容
不同尺度的网络状 态关系
•生命所特有的信息流、 物质流、能量流 •具有“变化着的结构” 不仅是线性或动力学 因素
• 跨尺度测量与控制 往往以各自研究的技 术背景从单一尺度去 理解和分析研究生物 过程的特点
呈网络多输入多输出关系
•不是简单的统计热力学关系 •网络结构表现在不同尺度的网络状态的互动关系
微生物代谢及调控

目录1.绪论 (2)1.1.微生物的代谢 (2)1.2.微生物代谢的调节 (2)2.微生物的初级代谢 (2)2.1.能量代谢 (2)2.1.1.能量代谢的载体ATP (2)2.1.2.ATP的代谢方式 (2)2.2.分解代谢 (3)2.2.1.糖的代谢 (3)2.2.2.脂肪和脂肪酸的代谢 (3)2.2.3.氨基酸的代谢 (4)2.2.4.核酸的代谢 (4)2.3.合成代谢 (4)2.3.1.糖类的合成 (4)2.3.2.脂类的合成 (5)2.3.3.氨基酸的合成 (5)2.3.4.蛋白质的合成 (6)2.3.5.核苷酸与核酸的合成 (7)3.微生物的次级代谢 (7)3.1.次级代谢概述 (7)3.2.次级代谢的意义 (8)3.3.次级代谢的生物合成 (8)3.4.次级代谢的特点 (8)4.微生物代谢的调节 (9)4.1.代谢调节的部位 (9)4.1.1.原核微生物细胞的代谢调节部位 (9)4.1.2.真核微生物细胞的代谢调节部位 (9)4.2.代谢调节的方式 (9)4.3.酶活性的调节 (9)4.3.1.酶活性调节的调节机制 (9)4.3.2.前馈与反馈 (10)4.3.3.反馈抑制 (10)4.4.酶合成的调节 (10)4.4.1.酶合成的诱导 (10)4.4.2.酶合成的阻遏 (10)4.4.3.酶合成诱导和阻遏的机制 (10)5.总结 (11)微生物代谢及其调节1.绪论1.1.微生物的代谢微生物代谢包括在微生物细胞中进行的所有生物化学反应的总和。
在代谢过程中,凡是能释放能量的物质分解过程称为分解代谢;吸收能量的物质合成过程称为合成代谢,因其导致新物质的生化合成也称为生物合成。
通过代谢,细胞吸收营养物质,并把它们转化为细胞成分,同时将废物排泄到体外。
无论是分解代谢还是合成代谢,代谢途径都是由一系列连续的酶促反应构成,其前一步反应的产物是后续反应的底物。
细胞通过各种方式有效的调节相关的酶促反应来保证整个代谢途径的协调性与完整性,从而使微生物细胞的生命活动的以正常进行。
微生物遗传育种名词解释(二)

1、自然选育:从自然界直接分离和筛选菌种或在生产中利用自发突变选育优良菌株。
2、诱变育种:对出发菌株进行诱变,然后运用合理的程序与方法筛选符合要求的优良菌株。
3、代谢调控育种:利用现有的代谢调控知识,筛选特定突变型,改变代谢流量或流向,从而提高目的产物产量的一种育种技术。
4、重组育种;利用微生物间的遗传重组来改变其遗传物质组成及结构的工业微生物育种技术。
5、原生质体融合育种;通过人为方法,使遗传性状不同的两细胞的原生质体发生融合,从而实现遗传重组的工业微生物育种技术。
6、基因工程育种技术:在体外构建重组DNA分子并导入宿主内高效表达,从而获得重组微生物的育种技术。
7、突变:遗传物质核酸中的核苷酸序列发生了稳定的可遗传的变化。
8、突变体:带有突变基因的细胞或个体9、突变型:突变体的基因型或表型称为突变型,和其相对的原存在状态称为野生型。
10、自发突变(spontaneous mutagenesis):未经任何人为处理而自然发生的突变;11、诱发突变(induced mutagenesis):由人们有意识地利用物理或化学手段对生物体进行处理而引起的突变。
12、整倍体:含有完整的染色体组。
13、非整倍体:含有不完整状态的染色体组,一般是指二倍体中成对染色体成员的增加或减少。
14、部分二倍体:原核生物中由一整条染色体和外来染色体片段所构成的不完整二倍体。
增变基因(mutator gene):其基因突变会导致整个基因组的突变频率明显上升的一些基因。
15、前突变:诱变剂所造成的DNA分子某一位置的损伤16、光复活:指细菌在紫外线照射后立即用可见光照射,可以显著地增加细菌的存活率,降低突变率。
17、表型延迟phenotype lag:突变体表型改变落后于其基因型改变的现象。
18、分离性延迟segregational lag :突变基因由杂合状态到纯合状态所造成的表型迟延19、生理性延迟physiological lag :由于基因产物的“稀释”过程所造成的表型迟延野生型(wild type):从自然界分离到的任何微生物在其发生营养缺陷突变前的原始菌株;基因重组:由于不同DNA链的断裂和连接而产生DNA片段的交换和重新组合,形成新的DNA分子,进而形成新遗传个体的方式称为基因重组。
各论II 23、代谢综合征

一、内脏脂肪积聚
(一)脂肪组织的生理功能
– 储存能量的器官 – 体内最大的内分泌器官
• 瘦素、脂联素、抵抗素、肿瘤坏死因子、内 脏脂肪因子、补体D、白介素6、纤溶酶原激 活物抑制物-1、血管紧张素Ⅱ、自由脂肪酸、 胰岛素样生长因子、血管内皮生长因子等
• 调节胰岛素敏感性、血压水平、内皮功能、 纤溶活动及炎症反应等
– 人类肥胖相关易感基因:90多个
β-肾上腺素受体基因家族的ADRB1、
肥 胖
影响能量摄入
ADRB2、ADRB3基因 解偶联蛋白家族UCP2、UCP3基因
相
关
影响能量消耗
易
过氧化物酶体增殖物激活受
感
体-γ基因( PPARγ )
基影响脂肪细胞贮存脂肪因19– 遗传学缺陷
• 单基因肥胖:人类的一些单基因的突变可以不依赖环境 而导致个体严重肥胖,其特点为早发性极度肥胖
14
1、瘦素---脂肪细胞分泌的饱感信号
– 抑制食欲,增加能量消耗 – 抑制胰岛素分泌,促进内脏脂肪分解,减少非
脂肪细胞甘油三酯的堆积 – 具有调节免疫和炎症的作用
15
2、脂联素
– 调脂作用 – 降糖作用:可促进糖吸收利用,抑制肝糖异生 – 抗炎作用:抑制TNF-α的生成与释放 – 胰岛素增敏作用 – 抗动脉粥样硬化作用
缺氧
进入脂肪
大量增加
脂肪细胞释放促炎因子
25
二、胰岛素抵抗(Insulin resistance,IR)
(一)血糖的调控--胰岛素和胰高血糖素
26
• 胰岛素的作用机制
– 磷酸酰肌醇-3激酶(PI-3K)途径 – Ras-MAPK途径
– 机制 • 存在瘦素抗体或瘦素拮抗剂 • 血脑屏障对瘦素的通透性降低 • 瘦素与受体结合障碍:瘦素受体基因( LEPR )突变 • 受体后信号转导缺陷:JAK-STAT途径 • 衰老
成纤维细胞生长因子23对磷代谢的调控及其临床意义

( K .T i rv w s m r e c n i i t it terg l i f G 2 np op a o e s s n i u ss h C D) hs e i u ma zs e e t n g s n e u t no F 3 o h sh t h m ot i a dds s e e i r sh oh ao F e as c et
C E ax ,Z A u I N H om n ,Y ii H N Y —i H NG K n ,TA a — ig U X -e j
Lbrtr o n or o g n tblm,W s C iaH si l i u nU wm  ̄,C eg u6 04 , hn a oao E dc nl ya dMe oi yf i o a s et hn o t ,Sc a n e i pa h hn d 10 1 C i a
D I139.n6 —92230 O:09/i .72 10 ..1 .6 js145.100 sFra bibliotek・ 专
论 ・
成 纤维 细胞 生 长 因子 2 3对磷 代 谢 的调控 及 其 临床 意义
陈亚希 ,张 坤 ,田浩 明 ,余希 杰
[ 要] 成纤维细胞生长 因子 2 ( G 2 0 摘 3 F F 3 )是最近发现 的一种新 型 内分泌激 素 ,主要 在骨细胞 和成骨 细胞 中表达 。F F 3需要 与 F F 一一 l h G2 G R 0 Ko o复合物结合 发挥其 生理功能。F F 3通过下调肾脏近曲小管上皮细 【 t G2 胞 内 N T a和 N T c P2 P 2 表达降低磷从原尿的重吸收 ,下调 1羟基酶 ( y 2 b )在 肾脏 近 曲小管 的表达 降低 活 一 Cp7 1 性12 ( H)D ,5 O , 的合成 ,进而减少小肠对磷 的吸收 。磷代谢 紊乱 常见于 先天性 代谢疾 病和慢 性 肾病 ( K 。 C D) 明确 F F 3在其中的病理作用将为上述疾病的诊 断和治疗 提供新思路和 目标。 G2 [ 关键词 ] 磷代谢 ;成纤维细胞生长因子一3 2 ;成纤维 细胞生 长因子受体 ;0 Ko o 【 lh 一 t
分子诊断学概论

分子诊断学概论一、分子诊断的基本概念与历史发展二、分子诊断的现状三、分子诊断的主要技术四、分子诊断的标准化与质量控制五、分子诊断的未来趋势分子诊断基本概念◆1953年,Watson和Crick提出了DNA双螺旋结构模型,为揭开人类生命现象的本质奠定了基础,标志着分子生物学的开端,也使得对疾病发病机制的认识从整体、细胞水平逐渐深入到分子水平◆分子诊断学(Molecular diagnostics),是以分子生物学理论为基础,利用分子生物学的技术和方法,研究人体内源性或外源性生物大分子和大分子体系的存在、结构或表达调控的变化,为疾病的预防、诊断、治疗和转归提供信息和依据的一门学科◆通常所称的基因诊断,指针对DNA或RNA的分子诊断技术临床检验诊断体外诊断(IVD )报告,影响约70%临床决策影像学诊断临床诊断疾病的检验诊断核磁共振辅助检验B 超CT体格检查病史临床检验诊断(实验室检验诊断)临床体液、血液检验临床化学检验临床免疫、血清学检验临床微生物学检验(细菌室)临床细胞分子遗传学检验CT (computed tomography ,电子计算机断层扫描)临床检验诊断发展阶段发展阶段历史时期技术类型典型特征简单划分第一代早期细胞形态学检验诊断•以疾病的表型改变为依据•非特异、滞后•难以早期诊断传统的临床检验诊断学学科第二代1950年代生物化学检验诊断第三代1960年代免疫学检验诊断第四代1970年代末基因检验诊断 (分子生物学检验诊断)•以疾病基因为探测对象•特异、敏感•早期诊断、预测新型的临床检验诊断学学科分子诊断(临床分子生物学检验诊断)分子生物学医学检验(临床检验诊断)分子生物学(molecular biology)1953年Watson&Crick发现DNA双螺旋结构模型70年代以来,成为生命科学最具活力的学科前沿分子医学(molecular medicine)、基因诊断(genetic diagnosis)分子生物学理论和技术方法被应用于临床分子生物学与医学的交叉和渗透国际首例基因诊断1970年代末美籍华裔简悦威(Yuet Wai Kan)分子杂交技术,α地中海贫血、镰状红细胞贫血我国基因诊断里程碑1984年,上海市儿童医院曾溢滔点杂交技术,α地中海贫血,发表在《Lancet》•以基因突变位点 (导致单基因遗传病) 为靶标第一代•核心技术:DNA或RNA分子杂交技术•以基因组特异性核酸序列 (DNA、RNA) 为靶标第二代•核心技术:Sanger测序技术、PCR技术•以基因组特异性核酸序列、蛋白质分子为靶标第三代•核心技术:生物芯片技术(高通量)•以基因组特异性核酸序列、蛋白质分子、代谢物为靶标第四代•核心技术:新一代测序技术、质谱技术分子诊断生物标志物◆核酸序列信息•个体差异基因:微卫星、SNP、mtDNA等•病原体基因组:病毒、细菌、真菌等•基因转录水平:mRNA、microRNA、lncRNA、circRNA、cfRNA等◆核酸序列变化•染色体变异:T21、T18、T13、CNV等•基因突变:点突变、插入/缺失突变、倒位突变、重复突变等◆核酸修饰•DNA甲基化•RNA甲基化◆蛋白质表达水平、修饰◆代谢产物、多糖链和脂质分子分子诊断学任务、特点、辨别◆任务•利用基础医学和生命科学的理论和方法,研究疾病发生和发展的分子机制•确定在疾病过程中特异的分子标志物•建立分子标志物的临床检验方法和评价体系•建立分子生物学检验的质量控制◆特点•主要是直接以疾病基因为探查对象,属于病因学诊断•对基因的检测结果不仅具有描述性,更具有准确性•可准确诊断疾病的基因型变异、基因表型异常以及由外源性基因侵入引起的疾病◆辨别•临床分子生物学检验技术=临床分子诊断技术•分子诊断VS基因诊断•分子诊断学包括:核酸诊断(DNA/RNA)、蛋白质检测诊断等分子诊断学概论一、分子诊断的基本概念与历史发展二、分子诊断的现状三、分子诊断的主要技术四、分子诊断的标准化与质量控制五、分子诊断的未来趋势医疗机构临床检验项目(2013版)临床体液、血液专业临床化学检验专业临床免疫、血清学专业临床微生物学专业临床细胞分子遗传学专业哪些专业含有基因诊断项目?临床免疫、血清学专业(摘录)序号项目名称1甲型肝炎病毒(HAV)RNA检测2乙型肝炎病毒(HBV)DNA测定3乙型肝炎病毒(HBV) YMDD变异检测4乙型肝炎病毒(HBV)前核心变异检测5乙型肝炎病毒(HBV)核心变异检测6乙型肝炎病毒(HBV)基因分型测定7丙型肝炎病毒(HCV)RNA测定8丙型肝炎病毒(HCV)分型9丁型肝炎病毒(HDV)RNA测定10庚型肝炎病毒核糖核酸定性(HGV-RNA)测定11戊型肝炎病毒(HEV)RNA测定12弓形体核酸测定13风疹病毒RNA测定14巨细胞病毒(CMV)DNA测定15水痘—带状疱疹病毒核酸测定16人乳头瘤病毒(HPV)基因检测17呼吸道合胞病毒核酸测定18流行性出血热病毒核酸测定19EB病毒核酸测定20副流感病毒核酸测定21人轮状病毒核酸测定22狂犬病毒核酸测定23乙型脑炎病毒核酸测定序号项目名称26柯萨奇病毒核酸测定27森林脑炎病毒(TBE)核酸测定28甲型流感病毒核酸测定29乙型流感病毒核酸测定30SARS冠状病毒核酸测定31BK病毒核酸测定32禽流感病毒核酸测定33埃可病毒核酸测定34西尼罗河病毒核酸测定35斑疹伤寒杆菌核酸测定36布氏杆菌核酸测定37结核分枝杆菌核酸测定38脑膜炎奈瑟菌核酸测定39幽门螺杆菌核酸测定40淋球菌核酸测定41嗜肺军团菌核酸测定42肺炎支原体核酸测定43生殖道支原体核酸测定44解脲脲原体核酸测定45肺炎衣原体核酸测定46鹦鹉热衣原体核酸测定47沙眼衣原体核酸测定48立克次体核酸测定临床细胞分子遗传学专业(摘录)序号项目名称备注1利用Southern blot分子杂交技术的白血病融合基因检查包括血友病A、血友病B、血菅性血友病、其它凝血因子缺陷症基因分析2利用Southern blot分子杂交技术的白血病融合基因检查1、 Ph染色体的分子杂交检查2、 RARA基因的分子杂交检查3、 AML1基因的分子杂交检查4、 E2A基因的分子杂交检查5、 MLL基因的分子杂交检查3利用RT-PCR或real time PCR技术的白血病融合基因检查1、Bcr-abl融合基因检查2、 AML1-EVI1融合基因检查3、 PML-RARA融合基因检查4、 DEK-CAN融合基因检查5、 AML1-MTG8融合基因检查6、 E2A-PBX1融合基因检查4单基因遗传病基因突变检查包括:1、进行性肌营养不良基因突变检查2、遗传性舞蹈病的基因突变检查3、其它5遗传性凝血因子缺陷症基因突变包括:1、血友病A的基因突变检查2、血友病B的基因突变检查3、混合型血友病的基因突变检查6α地中海贫血的基因突变检查7β地中海贫血的基因突变检查8苯丙酮尿症的基因突变检查9HLA低分辨基因分型检查10HLA高分辨基因分型检查序号项目名称备注12SRY的基因检查13P53基因的基因突变检查14K-Ras基因的基因突变检查15视网膜母细胞瘤RB1基因的基因突变检查16家族性乳腺癌基因的基因突变检查包括:1、BRCA1基因的基因突变检查2、BRCA2基因的基因突变检查3、其它17多发性内分泌腺瘤RET基因的基因突变的检查18遗传性非息肉性大肠癌的基因突变检查1、hMLH1基因的基因突变检查2、hMSH2基因的基因突变检查3、PMS1基因的基因突变检查4、PMS2基因的基因突变检查19遗传性大肠癌微卫星不稳定性(MSI)的基因检测20大肠癌易感基因的基因检测1、APC基因的基因检测2、DCC基因的基因检测21用于病毒、细菌用药指导的基因检测1、拉米夫定用药指导的基因检测2、结核病用药指导的基因检测3、肠球菌耐万古霉素用药指导的基因检测22用于化学药物用药指导的基因检测1、硝酸甘油用药指导的基因检测2、5-氟尿嘧啶用药指导的基因检测P450家族代谢酶基因的基包括CYP2C9、CYP2C19、CYP2D6、全国医疗服务项目技术规范(2023年版)◆检验+病理诊断项目合计1818项,增加了近60%,成为了11个大类中新增比例最高的板块实验室自建检测项目 (LDT)2022年12月《国家药监局综合司国家卫生健康委办公厅关于开展医疗机构自行研制使用体外诊断试剂试点工作的通知》,试点医疗机构包括:北京协和医院、北京医院、中日友好医院、中肿、阜外医院、北大一院等6家医院LDT(Laboratory developed test,实验室自建检测项目)感染领域:临床病原体检测方法微生物学检测:病原体培养/涂片病原体颗粒检测免疫学检测:检测血清学标志Ag、Ab分子诊断:检测DNA/RNA•耗时长•阳性率低•难培养•简便、快速•适于大规模筛查•可定性/定量检测•存在“窗口期”问题•不能早期诊断•灵敏度较低•快速、高通量•灵敏、特异•早期(缩短窗口期)•可分型•检测病原体突变•检测耐药基因•治疗监测病原体分子诊断检测病原体是否存在病原体分型(包括亚型)耐药基因检测相关的人类基因多态性检测标本类型外周血有核细胞血清血浆组织器官体液分泌物排泄物适宜分子诊断病原体类型难培养的如CT 、MG 、病毒培养较慢的如TB镜检容易弄错的如NG 、阴道毛滴虫免疫交叉反应较多的如CT 需要分型的如HPV 、HSV胞内病原体如衣原体、支原体、病毒CT (Chlamydia trachomatis ,沙眼衣原体)MG (Mycoplasma genitalium ,生殖支原体)TB (Mycobacterium tuberculosis ,结核分枝杆菌)NG (Neisseria Gonorrhoeae ,淋病奈瑟菌)HPV (human papillomavirus ,人乳头瘤病毒)遗传领域:镰状红细胞贫血症◆红血球不正常带来严重后果,问题在于血红蛋白ß链一个谷氨酸残基变成了缬氨酸残基◆常染色体隐性遗传病•基因点突变•Mst II 限制性内切酶位点改变•RFLP技术:酶切+电泳胚胎着床前分子诊断◆取1-2个囊胚期细胞进行基因诊断,从而将人类的遗传缺陷控制在最早期阶段无创产前诊断(NIPT )19972008卢煜明发现母体外周血中存在胎儿游离DNA高通量测序分析胎儿游离DNA 用于唐氏综合征筛查2009中国开始NIPT 临床试验2011中国、美国开始NIPT 临床服务2012美国妇产科协会推荐高危人群进行NIPT 201520172016中国无创单病开始临床应用卫计委推出NIPT 临床应用指南美国多种单基因疾病NIPT 临床服务2022美国妇产科协会推荐全人群进行NIPT国家药监局发布NIPT 注册指南◆胎儿游离DNA ◆高通量测序肿瘤领域:肿瘤靶向治疗◆高通量测序为主循环肿瘤DNA(ctDNA)年份事件1948血中游离DNA的发现1965肿瘤与血中游离DNA的相关性1966-1973系统性红斑狼疮等疾病患者血中游离DNA水平增高1977血中游离DNA水平与肿瘤病程及疗效相关1989发现血中游离DNA与原发肿瘤突变相似1994-1999更多证据表明血中游离DNA与原发肿瘤基因突变的一致性1997孕妇血中胎儿DNA的发现1998移植器官核酸可称为游离核酸成分的发现2000-2010游离DNA与多种疾病的诊断和预后相关2010游离DNA致癌性的确定ctDNADNA文库构建捕获扩增DNA&质控富集效率高通量测序和数据分析个体化用药领域:药物基因组药物作用靶点相关基因药物代谢相关基因药物副作用相关基因药物相关基因◆P53:50%以上人类肿瘤会发生p53基因突变◆BRCA1和BRCA2:乳腺癌易感基因1和2◆EGFR:表皮生长因子受体,细胞增殖和信号传导功能◆细胞色素P450超家族:人体内最大的药物代谢系统分子诊断学概论一、分子诊断的基本概念与历史发展二、分子诊断的现状三、分子诊断的主要技术四、分子诊断的标准化与质量控制五、分子诊断的未来趋势DNA->RNA->蛋白质->代谢产物◆基因(产物) 修饰•甲基化•乙酰化•磷酸化◆代谢及代谢调控分子诊断主要技术1. 分子杂交技术•遗传性疾病的基因诊断2. PCR技术•感染性疾病的基因诊断3. 生物芯片技术•复杂性疾病的基因诊断4. 基因测序技术•复杂性疾病的基因诊断5. 质谱技术•核酸质谱、蛋白质组学6. 人工智能辅助•AI辅助的分子诊断(AI+)1. 分子杂交技术杂交类型检测目的及范围Southern印迹杂交经凝胶电泳分离且转移至膜上,DNA分子Northern印迹杂交经凝胶电泳分离且转移至膜上,RNA分子菌落杂交固定在膜上,经裂解从细菌释放,DNA分子斑点杂交固定在膜上,DNA或RNA分子原位杂交(FISH)细胞或组织中,DNA或RNA分子液相分子杂交在溶液中,DNA或RNA分子,引入磁珠2. PCR技术◆痕量核酸模板体外扩增,提高了检测灵敏度和反应特异性•1971年,Korana提出核酸体外扩增的设想•1985年,Mullis发明聚合酶链反应,Klenow片段•1988年,Keohanog,T4DNA聚合酶•1988年,Saiki,TaqDNA聚合酶•1993年,Mullis因聚合酶链反应技术获得诺贝尔奖荧光定量PCR 技术◆也称为real-time PCR ,实现了核酸的实时定量检测◆Log 浓度与循环数呈线性关系,根据达到阈值的循环数计算样品所含模板量•荧光染料:SYBR green•荧光探针:Taqman 、molecular beacon 、复合探针•举例:新冠病毒检测荧光强度---循环数曲线初始模板量对数---Ct 循环数标准曲线10410310610510210数字PCR技术◆dPCR,又称为单分子PCR,近年来迅速发展起来的绝对定量PCR技术◆不依赖于扩增曲线的循环阈值进行定量,不受扩增效率的影响,也不必采用看家基因和标准曲线,具有很好的准确度和重现性,可以实现绝对定量分析3. 生物芯片技术◆广义指在微小空间中能够高通量处理或分析生物相关物质的集成式技术◆狭义指微阵列芯片技术,将大量基因探针/基因片段/蛋白/多肽,按特定的排列方式固定在支持物表面上,实现高通量处理或分析功能•固相芯片(玻片、硅片、塑料等)、液相芯片(微珠)•特点:高通量、微型化、自动化微流控芯片技术◆Microfluidics 技术,指的是使用微管道(尺寸为数十到数百微米)处理或操纵微小流体(体积为微升到纳升)的系统所涉及的科学和技术,是一门涉及化学、流体物理、微电子、新材料、生物学和生物医学工程的新兴交叉学科◆也被称为芯片实验室(lab on a chip )和微全分析系统(micro-total analytical system ),具有微型化、集成化等特征优势集成小型化与自动化样本量需求少试剂消耗少高通量污染少不足缺规范与标准技术难度不低生产成本较高开发周期较长4. 基因测序技术◆核酸测序技术,是分子诊断中基因序列确定的金标准ABI Prism310 1986年Roche 4542005年Illumina GA2006年ABI SOLiD2007年Helicos HeliScope2008年PacBio RS2010年ONT MinION2013年第一代(Sanger)第二代(NGS)第三代第四代或合称第三代(TGS)Sanger测序和NGS测序双脱氧末端终止法可逆终止、边合成边测序法单分子测序技术◆SMRT单分子实时合成测序技术,零模波导孔,荧光◆纳米孔单分子测序技术,纳米孔,电信号5. 质谱技术质量分析器离子源检测器多肽离子化 真空环境获得质谱图进样系统引入样品根据荷质比分离离子 检测记录离子信号计算机数据处理系统◆离子源•电子电离•快原子轰击离子化(FAB)•电喷雾离子化(ESI )•基质辅助激光解析离子化(MALDI)◆质量分析器•四极杆质谱(直流电极+射频电极,共4组)•飞行时间质谱(TOF)•离子阱质谱◆离子源与质量分析器组合•MAIDL-TOF-MS (基质辅助激光解析电离飞行时间质谱)•ESI-四极杆MS •ESI-串联MS6. AI辅助分子诊断◆AI+自动化流水线(包含分子诊断)•打通从标本到检验到临床的数据通路•及时准确地将“标本信息”转化为“检验数据”•再将“检验数据”转化为“临床诊疗信息”•大幅提高实验室咨询服务能力•医学检验工作向着更精准、高效的方向发展分子诊断学概论一、分子诊断的基本概念与历史发展二、分子诊断的现状三、分子诊断的主要技术四、分子诊断的标准化与质量控制五、分子诊断的未来趋势临床分子诊断方法性能评价◆定量检测方法和程序的分析性能验证内容,至少应包括准确度、精密度、可报告范围等◆定性检测项目验证内容,至少应包括检出限及符合率等,验证结果应经过授权人审核分子诊断存在的问题及原因◆假阳性问题◆假阴性问题◆重复性问题•同一实验室不同批次间重复测定,结果存在差异•不同实验室对同一标本检测,结果存在差异◆检测对象的多态性◆标本采集◆诊断试剂方法•准确性•特异性•检测限•检测范围•重复性•稳定性◆微量反应体系◆测定操作 (人员素质)◆仪器设备的维护校准 (定期)◆数据处理及结果报告个体差异样本量差异检测平台差异样本采集差异样本保存、运输差异分子诊断技术监管◆申请获批医疗器械证,有严格的管理•项目报批:卫健委批准•实验室:通过验收,定期校验仪器与器材•试剂:国家食品药品监督管理局(NMPA)批准•工作人员:经过培训,持证上岗•质量控制:室内质量控制(IQC),室间质量评价( EQA)◆LDT?国内正在摸索监管➢推荐“微专业-体外诊断与大数据分析”,《体外诊断产品注册与监管》,由项光新、李伟、连国军等老师授课国家如何监管医疗器械NMPA产品上市许可制度企业医疗器械生产企业许可国家机构法规生产质量管理规范规范性文件法律规章法规不良事件检测和报告医疗器械召回稽查局、法规司省和县级药监器械司、注册司质量监督机构技术审评机构分子诊断学概论一、分子诊断的基本概念与历史发展二、分子诊断的现状三、分子诊断的主要技术四、分子诊断的标准化与质量控制五、分子诊断的未来趋势将成为本世纪检验医学的主导技术◆应用面更广:扩展到复杂性疾病,检测未知病原体◆使用更便捷:自动化、智能化、普及化◆诊断更准确:致病根源、致病机制,定性->定量◆诊断更早期:早发现、早治疗,诊已病->诊未病•病原体的确认和定量、分型、耐药性检测1. 感染性疾病分子诊断•对遗传病进行确诊、分型和早期诊断2. 遗传病分子诊断•肿瘤的早期诊断、分型和伴随诊断3. 肿瘤分子诊断•药物基因组学、用药指导4. 个体化用药指导•公共卫生、器官移植、个体识别、基因治疗5. 其他领域美国《2030年全球趋势》未来分子诊断学的准确性将促使医疗体系变革基因检测方法将加速疾病诊断,同时帮助医师确定个性化最佳治疗方案感染领域:病原体检测⚫国内总体:年均非新冠的标本量约为1亿例⚫常规感染样本量:约为9000万例/年⚫危重感染样本量:约为1000万例/年,多数病原不明WHO 公布2019年全球十大健康威胁,与感染密切相关有6个:流感、耐药、埃博拉、登革热、艾滋病、疫苗犹豫临床宏基因组测序遗传领域:人类基因组临床应用Collins, FS & McKusick VA. Implications of the Human Genome Project for medical science. JAMA, 2001, 285: 540-554.单基因病无创产前筛查◆利用母体外周血中的胎儿游离DNA 的进行分子生物学检验,开展无创性性产前诊断,取代羊膜穿刺或采集绒毛进行无创性产前诊断方法8000病种多1%发病率高20%致死率高治疗方式少1%努南综合征1:2500 -1:1000Rett综合征(女性)1:23000 -1:10000Kabuki 综合征1:32000致死性骨发育不良1:10000-1:5000CHARGE 综合征1:15000 -1:8500软骨发育不全1:10000结节性硬化1:5,800马凡综合征1:10000 -1:5000单基因病占总出生缺陷的22.2%(染色体10%)复杂性疾病诊断。
上海交大医学院宁光团队发现肥胖治疗新靶标

上海交大医学院宁光团队发现肥胖治疗新靶标肥胖的基因治疗研究上海交大医学院宁光团队发现肥胖治疗新靶标—LGR4基因研究显示,LGR4基因的缺失可促进白色脂肪转化为棕色脂肪,增加机体能量消耗,减轻体重。
上海交通大学医学院附属瑞金医院宁光教授领衔的研究团队发现,通过调控LGR4基因可以影响肥胖的发生;这一基因好比神奇的开关,可调节白色脂肪和棕色脂肪之间的转化,因此,阻断LGR4 信号极可能成为肥胖干预的新靶点。
人体有两种脂肪组织,白色脂肪和棕色脂肪,白色脂肪将能量以甘油三酯形式储存起来,主要分布于皮下和内脏器官周围,白色脂肪过多就是肥胖;棕色脂肪将能量以热的形式散失掉,成年人体内只有很少量的棕色脂肪。
通俗来讲,白色脂肪是导致肥胖的“坏脂肪”,而棕色脂肪各项代谢指标,包括血糖、血脂、血压等均明显好转。
欧洲一个著名人类遗传学研究小组近期发表了一项研究成果,发现极少数欧洲后裔携带LGR4 失活突变,这一人群的体重明显低于正常对照人群。
这一成果进一步支持上海瑞金医院肥胖研究团队的研究结果,LGR4 极有可能是一个全新的肥胖致病基因。
上海瑞金医院肥胖研究团队的相关研究已经在《自然-细胞生物学》发表;接下来,研究人员将努力发现LGR4 的配体和阻断剂,以推动LGR4 作为肥胖靶点的干预研究,造福肥胖患者。
上海交大科研团队发现黄连素或可治疗肥胖上海交通大学医学院附属瑞金医院副院长宁光教授领衔的团队新近研究发现,中药黄连素可激活白色脂肪和棕色脂肪组织的生热作用,能够调控机体的能量平衡,在肥胖治疗中可能具有潜在的应用前景。
该研究论文日前发表在《自然·通讯》杂志上。
专家介绍,脂肪组织主要分为两种类型:白色脂肪组织和棕色脂肪组织。
白色脂肪广泛分布于皮下组织和内脏周围,主要以甘油三酯的形式储存能量并分泌一些影响能量平衡的激素和细胞因子。
棕色脂肪则主要存在于人类新生儿和幼小的哺乳动物体内,它们作为机体的“加热器”通过线粒体的脂质氧化解耦联产热从而消耗能量,同时也可分泌少量的脂肪因子。
原核生物基因表达调控

20
同位素示踪实验
把大肠杆菌细胞放在加有放射性35S标记的氨基酸,但没 有半乳糖诱导物的培养基中繁殖几代然后再将这些带有 放射活性的细菌转移到不含35S、无放射性的培养基中 随着培养基中诱导物的加入, β-半乳糖苷酶便开始合成。 分离β-半乳糖苷酶, 发现这种酶无35S标记说明酶的合 成不是由前体转化而来的, 而是加入诱导物后新合成的。
• Jacob和Monod认为诱导酶(他们当时称为适应酶)
现象是个基因调控问题, 可以用实验方法进行研究, 因此
选为突破口, 终于通过大量实验及分析, 于1961年建立
了该操纵子的控制模型。
-
21
酶的诱导
-
22
• 酶的诱导现象是生物进化过程中出现的一种合理、 经济地利用有限资源的本能。
• 酶诱导已证明是低等生物的普遍现象。
倒位片段
鼠伤寒沙门菌鞭毛素基- 因的调节
H1鞭毛素
10
鼠伤寒沙门氏菌(S.typhimrium)的相转变(phase variation)
-
11
2.σ 因子对原核生物转录起始的调控
σ因子:原核生物RNA聚合酶的一个亚基,是转录起 始所必需的因子,主要影响RNA聚合酶对转录起始 位点的正确识别,这种σ因子称σ70,此外还有分子量 不同,功能不同的其他σ因子 。
PO
操纵子可视为原核生物的转录单位,它可以逐个
地从原核生物基因组中分离出来,对其结构功
能加以研究。
-
15
3.乳糖操纵子
1) 乳糖操纵子的结构
启动子 操纵基因
调节蛋白
(阻遏蛋白)
-
结构基因
16
3个编码的结构基因
• Z编码β-半乳糖苷酶: 将乳糖水解成葡萄糖和半乳糖,还能 将乳糖转变为异构乳糖
代谢生物化学第二十三章 三羧酸循环
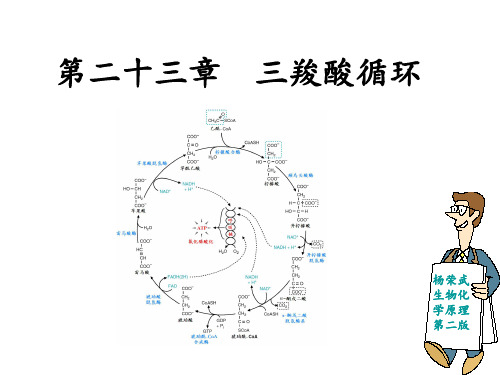
α-酮戊二酸的氧化脱羧
反应5:底物水平的磷酸化
TCA循环唯一的一步底物水平磷酸化反应
« 琥珀酰-CoA合成酶 « ATP或GTP被合成 « 它的催化过程牵涉到一系列高能分子的形成,
因此能量的损失微乎其微 « 反应机制涉及一个磷酸组氨酸
底物水平的磷酸化
反应6:琥珀酸的脱氢
« 由苹果酸脱氢酶催化 «ΔGo‘ = +30 kJ/mol,意味着在热力学上极不
利于正反应的进行,但在体内,反应产物草 酰乙酸可以迅速被下一步不可逆反应消耗, NADH则进入呼吸链被彻底氧化,因此,整 个反应被“强行拉向”正反应。 tca_cycle_v9.swf
草酰乙酸的再生
TCA 循环总结
ó 脂肪酸的β氧化 ó 氨基酸的氧化分解 ó 丙酮酸的氧化脱羧——由丙酮酸脱氢酶系催化
反应1:柠檬酸的合成
« 这是一步不可逆反应 « 由柠檬酸合酶催化 « 柠檬酸合酶由两个相同的亚基组成,它被视为酶“诱导
契合”学说又一代表性的例子
« 在无底物结合时,酶两个亚基的构象呈开放型;当结合 底物以后,则被诱导为紧密型。在反应中,OAA首先与 酶活性中心结合,这种结合迅速诱导活性中心的构象发 生变化,从而创造出乙酰-CoA的结合位点。随后,乙酰CoA结合到酶活性中心,并与OAA形成柠檬酰-CoA。这 时,酶的构象再次发生变化,远离活性中心的一个关键 的Asp残基被拉入到柠檬酰-CoA上的硫酯键,很快硫酯键 被切开,终产物辅酶A和柠檬酸被依次释放。
« 乙酰-CoA是TCA循环第一步反应的底物,因此,机体 还可以通过控制它的形成来控制TCA循环,而这必然 牵涉到细胞内参与乙酰-CoA合成的酶,例如丙酮酸脱 氢酶系和参与脂肪酸β-氧化的酶。
生物化学第13章代谢调节

第十三章代谢调节一、填空题:1.生物体内的代谢调节在三种不同的水平上进行,即、和。
2.代谢途径的终产物浓度可以控制自身形成的速度,这种现象被称为。
3.酶对细胞代谢的调节是最基本的代谢调节,主要有二种方式:和。
构通糖、脂代谢的关键化合物是。
4.不同代谢途径可以通过交叉点代谢中间物进行转化,在糖、脂、蛋白质及核酸的相互转化过程中三个最关键的代谢中间物是、和。
5.1961年,法国生物学家Monod和Jacob提出了关于原核生物基因结构及表达调控的学说。
6.正调控和负调控是基因表达的两种最基本的调节形式,其中原核细胞常用调控,而真核细胞常用调控模式。
7.乳糖操纵子的天然诱导物是,实验室里常用作为乳糖操纵子的安慰诱导物诱导β-半乳糖苷酶的产生。
8.许多代谢途径的第一个酶是限速酶,终产物多是它的,对它进行,底物多为其。
9.原核细胞酶的合成速率主要在水平进行调节。
10.乳糖操纵子的诱导物是,色氨酸操纵子的辅阻遏物是。
二、选择题(只有一个最佳答案):1.下列与能量代谢有关的过程除哪个外都发生在线粒体中?()A、糖酵解B、三羧酸循环C、脂肪酸的β-氧化D、氧化磷酸化2.IPTG可以诱导乳糖操纵子(lacOperon)的表达,这是因为:()A、IPTG与乳糖操作子(lacoperator)结合,诱导转录B、IPTG与LACI基因产物结合,并抑制其活性C、抑制β-半乳糖苷酶的活性D、促进Lac阻遏物的活性E、IPTG与LACI基因产物结合,并激活其活性3.在什么情况下,乳糖操纵子的转录活性最高?()A、高乳糖,低葡萄糖B、高乳糖,高葡萄糖C、低乳糖,低葡萄糖D、低乳糖,高葡萄糖4.真核细胞参与基因表达调节的调控区比原核细胞复杂是因为()A、真核细胞的细胞核具有双层膜B、原核细胞的基因总是以操纵子的形式存在C、原核细胞调节基因表达主要是在翻译水平D、真核细胞需要控制细胞特异性的基因表达E、真核细胞基因组含有太多的重复序列5.调节物质代谢体内最基础的层次是()A、细胞水平B、激素水平C、神经调节D、整体水平E、器官水平6.磷酸果糖激酶是什么代谢途径中的别构调节酶()A、三羧酸循环B、糖异生C、葡萄糖分解D、糖原合成E、糖原分解7.三羧酸循环中的别构调节酶是()A、柠檬酸合成酶B、α-酮戊二酸脱氢酶C、琥珀酸脱氢酶D、延胡索酸酶E、苹果酸脱氢酶8.催化糖酵解与磷酸戊糖途径的酶主要分布在细胞中什么部位()A、核B、胞质C、线粒体D、微粒体E、质膜9.催化三羧酸循环与脂肪酸β-氧化的酶分布在细胞内的什么部位()A、胞质B、胞膜C、胞核D、内质网E、线粒体10.氨基酸分解代谢调节的别构酶是()A、转氨酶B、脱羧酶C、转甲基酶D、己糖激酶E、谷氨酸脱氨酶11.糖异生限速酶的别构调节激活剂是()A、ATPB、ADPC、AMPD、dATPE、cAMP12.各种分解途径中,放能最多的途径是:()A、糖酵解B、三羧酸循环C、(-氧化D、氧化脱氨基13.操纵子调节系统属于哪一种水平的调节?()A、复制水平的调节B、转录水平的调节C、转录后加工的调节D、翻译水平的调节14.下列关于操纵基因的论述哪个是正确的?()A、能专一性地与阻遏蛋白结合B、是RNA聚合酶识别和结合的部位C、是诱导物和辅阻遏物的结合部位D、能于结构基因一起转录但未被翻译15.以下有关阻遏蛋白的论述哪个是正确的?()A、阻遏蛋白是调节基因表达的产物B、阻遏蛋白妨碍RNA聚合酶与启动子结合C、阻遏蛋白RNA聚合酶结合而抑制转录D、阻遏蛋白与启动子结合而阻碍转录的启动16.糖酵解中,下列哪一个催化的反应不是限速反应?()A、丙酮酸激酶B、磷酸果糖激酶C、己糖激酶D、磷酸丙糖异构酶17.磷酸化酶通过接受或脱去磷酸基而调节活性,因此它属于:()A、别(变)构调节酶B、共价调节酶C、诱导酶D、同工酶18.下列与能量代谢有关的途径不在线粒体内进行的是:()A、三羧酸循环B、脂肪酸β氧化C、氧化磷酸化D、糖酵解作用19.关于共价修饰调节酶,下列哪种说法是错误的?()A、这类酶一般存在活性和无活性两种形式,B、酶的这两种形式通过酶促的共价修饰相互转变C、伴有级联放大作用D、是高等生物独有的代谢调节方式20.阻遏蛋白结合的位点是:()A、调节基因B、启动因子C、操纵基因D、结构基因21.下面哪一项代谢是在细胞质内进行的:()A、脂肪酸的β-氧化B、氧化磷酸化C、脂肪酸的合成D、TCA22.在乳糖操纵子模型中,操纵基因专门控制是否转录与翻译。
成纤维细胞生长因子23信号通路在慢性肾脏病及相关并发症中的作用

㊃综述㊃基金项目:河北省自然科学基金资助项目氨基化纤维素治疗慢性肾衰大鼠高磷血症效果及减少主动脉钙化机制(H 2018201289)通信作者:邹丹丹,E m a i l :z d d 1992512@163.c o m成纤维细胞生长因子23信号通路在慢性肾脏病及相关并发症中的作用邹丹丹,张海松,汪 煜(河北大学附属医院肾内科,河北保定071000) 摘 要:慢性肾脏病(C K D )是由多种因素共同作用的慢性肾功能减退性疾病,其发生㊁发展及转归机制尚未完全阐明㊂C K D 发病隐匿,早期无明显体征,而随着病情进展,肾功能不断减退,由钙磷代谢紊乱引起的心血管并发症是C K D 患者的主要致死原因㊂成纤维细胞生长因子23(F G F 23)对钙磷代谢具有调节作用,可反映早期肾功能变化情况,因此C K D 患者血清中F G F 23及其相关信号通路的变化有助于及早评估肾功能㊂本文就F G F 23及其相关信号通路的研究进展作一综述㊂关键词:肾疾病;成纤维细胞生长因子23;信号通路;研究进展中图分类号:R 692 文献标志码:A 文章编号:1004-583X (2019)12-1143-05d o i :10.3969/j.i s s n .1004-583X.2019.12.019 慢性肾脏病(C K D )作为世界范围内的一个主要公共卫生问题日益受到重视㊂美国肾脏数据系统(U S R D S )2019年度数据报告显示,美国C K D 的患病率仍逐年稳步上升,从2016至2017年,C K D 确诊比例从13.8%上升到14.5%,而终末期肾病(E S R D )的患病率增长了2.6%㊂在这些新增的E S R D 患者中,血液透析(H D )仍是他们首选的肾脏替代治疗模式㊂中国肾脏疾病监测网络(C K -N E T )2015年度数据报告显示,在1850万住院患者中C K D 患者占4.8%,合并其他非传染性慢性病患者中C K D 比例增加,尤其是糖尿病和高血压患者[1]㊂C K D 患者院内病死率为2.6%,伴有心力衰竭的患者院内病死率显著增加㊂大量的临床研究表明,年龄㊁性别㊁高血压㊁糖尿病㊁心血管疾病史及高尿酸血症都与C K D的发生发展密切相关[2]㊂C K D 患者因肾功能进行性衰退导致的E S R D 病例数逐年上升,而H D 和腹膜透析(P D )作为维持E S R D 患者正常病理生理水平的重要手段,其治疗成本较高㊂另外,透析并发症及相关感染,也使得E S R D 患者的生存质量存在着不可预知的风险㊂因此,对C K D 患者进行早期诊断㊁早期进行肾脏保护治疗㊁延缓肾功能的衰退㊁减少C K D 患者年透析次数不仅可以提高C K D 及E S R D 患者的生存质量,而且还可以减少相关治疗的医疗费用㊂1 F G F -23及K l o t h o 蛋白简介1.1 F G F 23 F G F 家族包括22个具有不同功能的成员,其中F G F 23是一类主要由骨细胞和成骨细胞产生的多肽,其相对分子质量为32000(251个氨基酸),定位在染色体12p 13上㊂在分泌过程中,血清全段成纤维细胞生长因子23(i F G F 23)可被降解成为N 末端F G F 23(n F G F 23)和C 末端F G F 23(c F G F 23),n F G F 23具有F G F 23受体(F G F R )结合域,可与F G F R 结合,c F G F 23则可与K l o t h o 蛋白结合,但只有i F G F 23才具有生物活性㊂F G F 23不同于F G F 家族其他成员的功能,其主要通过抑制近端肾小管对钠依赖性磷酸盐重吸收以及1α-羟化酶活性来调节血清钙磷水平㊂在C K D 早期,血清F G F 23水平随肾小球滤过率(e G F R )的降低逐渐升高,提示F G F 23可能作为C K D 的早期生物学标志[3]㊂F G F 23在肾脏㊁甲状旁腺等组织中起调节磷因子的作用,主要参与磷和维生素D 的代谢调控㊂F G F 23信号通路的异常与多种代谢性疾病尤其是C K D 的发生密切相关㊂F G F 23水平的不断升高是导致C KD 患者疾病进程加快㊁诱发并发症甚至最终死亡的主要因素㊂1.2 K l o t h o 蛋白 K l o t h o 蛋白作为F G F 23的结合蛋白,是F G F R 激活所必需的㊂事实上,K l o t h o 可以与多个F G F R 结合,并增强它们与F G F 23的亲和力㊂广泛表达的F G F R 与有限表达的K l o t h o 的共同表达提示可能是F G F 23作用的靶点㊂K l o t h o 蛋白主要分为3种亚型:α-K l o t h o ㊁β-K l o t h o 和γ-K l o t h o ㊂其中α-K l o t h o 主要以膜型(m K l o t h o)和分泌型(s K l o t h o )2种形式存在,m K l o t h o 主要在肾脏㊁甲状旁腺和脑组织中表达,参与F G F 23相关的钙㊁磷㊁骨代谢过程,s K l o t h o 则通过K l o t h o 基因选㊃3411㊃‘临床荟萃“ 2019年12月20日第34卷第12期 C l i n i c a l F o c u s ,D e c e m b e r 20,2019,V o l 34,N o .12Copyright ©博看网. All Rights Reserved.择性剪切㊁翻译表达后直接进入循环,在抗氧化㊁调控离子传递和W n t信号方面起着重要作用㊂2调控F G F23生成因素2.1局部因素 F G F23的翻译后修饰过程和受体结合特性一直受到广泛关注㊂然而,F G F23的调控机制尚不清楚,现有研究主要有以下几方面:①骨细胞分泌的牙本质基质蛋白1(D M P1)㊁磷酸盐调节蛋白(P H E X)㊁细胞外基质磷酸化糖蛋白(M E P E)及硬骨素(S O S T)能够调节F G F23水平㊂D M P1参与黏着斑激酶(F A K)介导的丝裂原活化蛋白激酶(MA P K)信号通路,下调F G F23表达[4]㊂而D M P1和P H E X基因突变可激活骨中典型的F G F/F G F R 通路导致F G F23产量增加[5],M E P E的C端富含丝氨酸-天门冬氨酸基序(A S A R M基序)能够与P H E X 结合,并阻止M E P E被组织蛋白酶B降解为游离的A S A R M基序,从而促进骨的矿化过程㊂S O S T能够和W n t蛋白竞争低密度脂蛋白受体相关蛋白(L R P5/6)在胞外的结合位点,从而抑制W n t信号通路,干扰成骨细胞介导的骨形成,研究发现敲除S O S T基因的小鼠F G F23水平降低[6]㊂因此, D M P1㊁P H E X㊁M E P E和S O S T均对F G F23的表达具有调控作用㊂②目前研究发现,人类F G F2基因能够编码22k D a-34k D a的高分子量(HMW)核亚型和18k D a的低分子量(L MW)核亚型,HMW亚型通常存在于细胞内,能够协助细胞内成纤维细胞生长因子受体1(F G F R1)/环磷酸腺苷反应元件结合蛋白(C R E B)的结合蛋白(C B P)/C R E B信号通路的核定位和激活,与F G F23启动子中的环磷酸腺苷反应元件(C r e)结合启动转录,L MW亚型则通过膜F G F R介导的磷酯酶Cγ(P L Cγ)和MA P K分别激活活化T细胞核因子(N F A T)和E26转录因子1(E T S1)以刺激成骨细胞中F G F23启动子活性[7]㊂③N o t c h受体胞内段(N I C D)是N o t c h信号通路的活性片段,在N o t c h受体与配体结合时,由γ-分泌酶作用使之从细胞膜水解出来,并转移到细胞核内,与N o t c h的下游效应蛋白C S L结合,从而激活N o t c h 靶基因的转录㊂N I C D转基因小鼠实验表明,F G F23在骨细胞中的表达受N o t c h信号途径的正向调控,并与H e s1和H e y1基因的上调有关[8]㊂2.2全身因素1,25(O H)2V i t D增加了E T S1水平,并且E T S1-V D R E/N u r r1复合元件在维生素D 激素调控F G F23m R N A的过程中起重要作用[9]㊂E T S1通过协助维生素D受体(V D R)使之与维甲酸X受体(R X R)结合形成V D R-R X R异二聚体,该二聚体与维生素D反应元件(V D R E)结合,激活下游核受体相关因子1(N u r r1),上调F G F23的表达㊂甲状旁腺激素(P T H)可与骨表面甲状旁腺激素1受体(P T H1R)结合,从而激活蛋白激酶A(P K A)通路,通过N u r r1结合于F G F23启动子区域,从而使得F G F23表达增加(图1)㊂C K D患者长骨中局部表达的K l o t h o可使F G F23水平升高[10],而M a等[11]指出s K l o t h o可通过抑制W n t/β-连环蛋白(β-c a t e n i n)信号通路抑制成骨样细胞UM R-106合成F G F23㊂这些数据表明,m K l o t h o和s K l o t h o对合成F G F23反应具有拮抗作用㊂其他因素如促红细胞生成素(E P O)㊁磷酸盐㊁钙㊁醛固酮㊁缺铁㊁代谢性酸中毒㊁雌激素㊁瘦素等也可增加血清中F G F23水平㊂3F G F23对C K D患者相关靶器官的调控作用3.1 F G F23对肾脏的调控α-K l o t h o与F G F R1c㊁F G F R3c和F G F R4结合形成的二元复合物允许F G F23与之结合,而硫酸乙酰肝素(H S)使形成的三元复合物进一步二聚化,从而激活下游的信号通路㊂F G F23与α-K l o t h o结合后,可使成纤维生长因子受体底物2(F R S2)和MA P K磷酸化,进而下调近端小管钠依赖性磷酸盐转运蛋白2a(N a P i-2a)和钠依赖性磷酸盐转运蛋白2c(N a P i-2c)在细胞膜上的表达,减少磷酸盐的重吸收㊂同时,在近端小管中,F G F23能够在K l o t h o存在时激活上皮细胞中细胞外调节蛋白激酶1/2(E R K1/2),下调1α-羟化酶(C Y P27B1)的表达,并增加24-羟化酶(C Y P24A1)的表达,从而降低1,25(O H)2V i t D的生成㊂S a k a n 等[12]指出,随着G F R的升高,K l o t h o在肾脏中的表达下降,因此,在C K D晚期F G F23对肾脏的作用减弱,高F G F23水平不仅不能有效调节钙磷平衡,反而出现一系列毒性作用,如继发性甲状旁腺功能亢进(S H P T)㊁佝偻病㊁左心室肥厚(L V H)㊁血管钙化等㊂3.2 F G F23与肾性贫血临床研究表明,血清高F G F23水平㊁低血红蛋白水平与贫血总体风险发生率之间存在相关性[13]㊂为了明确F G F23是否在红细胞生成中发挥作用,C o e等[14]利用F G F23完全缺失的小鼠和流式细胞术分析等工具,发现随着F G F23基因的失活,骨髓和外周血中造血干细胞(H S C)和红系祖细胞明显增加,并且E P O水平明显高于野生型对照组㊂更重要的是,将外源性F G F23注射到野生型小鼠腹腔后可产生相反的结果㊂另外,A g o r o等[15]利用5/6肾切除(N x)小鼠术后8周使用F G F23阻断肽进行贫血治疗,在该N x模型中, F G F23阻断肽单次腹腔注射可挽救肾性贫血㊂提示血清F G F23升高可负调控红细胞生成,降低肾脏㊃4411㊃‘临床荟萃“2019年12月20日第34卷第12期 C l i n i c a l F o c u s,D e c e m b e r20,2019,V o l34,N o.12Copyright©博看网. All Rights Reserved.E P O的释放,是H S C向红系分化的负中介;FG F23还可直接降低细胞周期G2/M期红细胞的比例,促进红细胞凋亡㊂此外,F G F23也可间接导致肾性贫血㊂S i n g h 等[16]指出,通过使用F G F23过表达的动物模型显示,F G F23可直接作用于肝脏,增强炎症环境㊂在血清F G F23水平不受控制的情况下,F G F23与肝细胞表面的F G F R4结合,激活P L Cγ/钙调神经磷酸酶(C a N)/N F A T信号通路,导致白细胞介素6(I L-6)和C反应蛋白(C R P)的合成和分泌增加,并且在F G F R4被药物抑制后,这些炎性标记物及其肝脏产物得到改善[17]㊂急性和慢性炎症可促进F G F23的生成,而血清F G F23水平过高又可直接刺激多种促炎性细胞因子的合成和分泌,从而形成一种恶性循环,导致广泛的组织损伤和C K D的加速进展㊂由于炎症环境的增强,促炎性细胞因子上调了肝内铁调素的产生[18]㊂而铁调素通过抑制胃肠道铁吸收并阻止循环铁从网状内皮系统释放到血浆中,导致铁代谢紊乱,使红细胞生成减少,引起肾性贫血[19]㊂3.3 F G F23对甲状旁腺的调控 S H P T是C K D患者的常见并发症,F G F23调节P T H的合成与分泌,它们之间存在着负反馈调节作用㊂在正常生理条件下,甲状旁腺细胞膜上可同时表达m K l o t h o及F G F R1,F G F23与m K l o t h o-F G F R1复合体结合可激活细胞内MA P K信号传导途径,使E R K1/2磷酸化,提高细胞内早期生长反应因子1(E g r-1)的转录水平,抑制P T H转录㊂同时,O l a u s o n等[20]发现,在K l o t h o基因缺失的小鼠中,F G F23激活N F A T C2途径,实现了其抑制P T H水平的作用,并且当该途径被阻断后,F G F23对P T H的调节作用也随之消失,可见F G F23可独立于K l o t h o信号通路抑制P T H的产生㊂然而,G a l i t z e r等[21]对C K D大鼠模型研究发现,与对照组相比,E S R D大鼠甲状旁腺组织中K l o t h o和F G F R1的转录及翻译水平均降低㊂在以尿毒症患者为研究对象的临床观察中,也得到了类似的结论[22]㊂因此,由于甲状旁腺对高水平F G F23的抵抗,血清P T H和F G F23水平均升高㊂引起S H P T的可能原因如下:血钙升高可激活甲状旁腺内的钙敏感受体(C a S R),导致MA P K级联反应,而E S R D的C a S R表达下降抑制该反应致使P T H水平升高,同时双特异性磷酸酶(D u s p)可加速MA P K失活;D u s p是F G F23-MA P K-E R K1/2信号传导途径的特异性负反馈调节因子,D u s p的过表达导致E R K失活;F G F23对1,25(O H)2V i t D存在负反馈调节,而1,25(O H)2V i t D可与V D R结合上调K l o t h o基因的转录,在C K D伴S H P T患者中,甲状旁腺V D R明显减少,F G F23可能通过下调1,25 (O H)2V i t D水平继而减少V D R表达,降低K l o t h o 水平㊂3.4 F G F23对心血管的调控3.4.1心肌肥厚动物实验表明,K l o t h o可能通过减少细胞表面存在的瞬时受体电位阳离子通道6 (T R P C6),在一定的应激条件下导致内向钙电流的减少,从而抑制心肌肥厚和炎症因子的产生㊂研究表明,L V H的C K D患者心肌细胞中F G F23表达增加[23]㊂此外,有证据表明,α-K l o t h o在心肌中的表达水平在生理上并不显著,但血清高浓度的F G F23可通过F G F R4介导的非依赖K l o t h o的信号转导途径直接作用于心肌细胞,F G F23与F G F R4结合后激活P L Cγ/C a N/N F A T信号通路,导致心室重构[24]㊂F G F23的局部分泌增加也可刺激F G F R4介导的信号传导,因此F G F23也可以旁分泌的方式参与L V H 的发生发展㊂3.4.2血管钙化血管钙化是一种异位性软组织矿化过程,可显著增加心血管疾病的病死率㊂既往对F G F23和血管钙化之间的相关性进行了研究,但结果并不一致㊂部分研究发现循环中F G F23水平与冠状动脉钙化之间呈正相关[25]㊂另有研究发现K l o t h o表达于体外培养的人类主动脉血管平滑肌细胞(H A S M C s)表面,F G F23通过K l o t h o-F G F R1途径上调S M C s中骨保护素(O P G)表达,O P G通过抑制N o t c h1-重组信号序列结合蛋白(R B P-J K)信号途径抑制成骨细胞特异性基因M S X2表达,进而抑制S M C s向成骨细胞分化及骨矿化过程[26]㊂但也有研究者观察到在人类S M C s中并无K l o t h o受体,即使在s K l o t h o存在的情况下,血浆F G F23与冠状动脉钙化的患病率或严重程度无关[27]㊂总之,F G F23与血管钙化之间的相关性尚无确切证据,需要进一步研究㊂此外,目前还不清楚F G F23是否在不同血管床上有不同的作用㊂3.5 F G F23对骨骼的调控骨骼是一个不断重塑的动态器官,骨骼在形成过程中受到W n t/β-c a t e n i n 通路㊁O P G/核因子κB受体活化因子配体(R A N K L)/核因子κB受体活化因子(R A N K)通路㊁转化生长因子β(T G F-β)/骨形成蛋白(B M P)/S m a d 通路㊁N o t c h通路㊁磷脂酰肌醇-3-激酶(P I3K)/蛋白激酶B(A K t)通路等的调节,这些通路中许多成分的作用相互重叠,而F G F23对这些通路的作用机制尚未完全阐明㊂骨细胞局部分泌的F G F23可能以自分泌/旁分泌的方式调节骨的矿化过程㊂F G F23过㊃5411㊃‘临床荟萃“2019年12月20日第34卷第12期 C l i n i c a l F o c u s,D e c e m b e r20,2019,V o l34,N o.12Copyright©博看网. All Rights Reserved.表达可引起佝偻病或骨软化㊂研究发现,F G F23通过F G F R3介导的非K l o t h o依赖途径,从转录水平上抑制成骨细胞中骨矿化调节剂 非特异性碱性磷酸酶(T N A P)的表达,抑制焦磷酸盐(P P i)裂解为磷酸盐(P i),进而抑制骨的矿化过程[28]㊂此外,F G F23和s K l o t h o的联合作用通过上调D K K1来增加β-c a t e n i n的非活性形式,从而抑制间充质干细胞分化和成骨细胞矿化[29]㊂F G F23的心血管调控见图1㊂图1F G F23的多效性作用近年来,C K D发病率逐年上升,早期难以发现,严重威胁着人类健康㊂随着病情进展,发展至E S R D 患者需要肾脏替代治疗来维持正常生活,但由此引发的并发症及相关感染会加重患者病情,给患者生存带来极大危险㊂F G F23作为参与钙磷代谢紊乱的调节因子,在C K D早期即可出现升高,早于血磷及P T H水平的改变,并随病情进展动态演变,有助于C K D的早期筛查和随访跟踪㊂同时,肾脏㊁甲状旁腺㊁心血管和骨骼等器官均可受到F G F23的调控,为C K D-M B D(慢性肾脏病-矿物质与骨异常)的进展提供早期评估指标㊂因此,通过对F G F23及其相关信号通路的研究,将有助于了解C K D的发生发展过程,为早期筛查疾病㊁延缓病情进展㊁改善患者生活质量提供新的治疗途径及依据㊂参考文献:[1] W a n g F,Y a n g C,L o n g J,e t a l.E x e c u t i v e s u mm a r y f o r t h e2015A n n u a l D a t a R e p o r t o f t h e C h i n a K i d n e y D i s e a s eN e t w o r k(C K-N E T)[J].K i d n e y I n t,2019,95(3):501-505.[2]康英丽,李英.2018年肾脏疾病研究进展[J].临床荟萃,2019,34(1):56-63.[3]I s a k o v aT,W a h lP,V a r g a s G S,e ta l.F i b r o b l a s t g r o w t hf a c t o r23i s e l e v a t e d b e f o r e p a r a t h y r o i d h o r m o n e a n d p h o s p h a t ei nc h r o n i ck i d n e y d i s e a s e[J].K i d n e y I n t,2011,79(12):1370-1378.[4] L e e J W,Y a m a g u c h i A,I i m u r aT.F u n c t i o n a l h e t e r o g e n e i t y o fo s t e o c y t e s i nF G F23p r o d u c t i o n:t h e p o s s i b l e i n v o l v e m e n to fD M P1a s ad i r e c t n e g a t i v e r e g u l a t o r[J].B o n e k e y R e p,2014,3:543.[5] M a r t i nA,L i uS,D a v i d V,e ta l.B o n e p r o t e i n sP H E Xa n dD M P1r e g u l a t e f i b r o b l a s t i c g r o w t hf a c t o rF g f23e x p r e s s i o n i no s t e o c y t e s t h r o u g h a c o mm o n p a t h w a y i n v o l v i n g F G F r e c e p t o r(F G F R)s i g n a l i n g[J].F A S E BJ,2011,25(8):2551-2562.[6] K u m a rR,V a l l o n V.R e d u c e dr e n a l c a l c i u m e x c r e t i o ni nt h ea b s e n c e o f s c l e r o s t i ne x p r e s s i o n:e v i d e n c e f o r an o v e l c a l c i u m-r e g u l a t i n g b o n ek i d n e y a x i s[J].JA m S o cN e p h r o l,2014,25(10):2159-2168.[7] H a n X,X i a o Z,Q u a r l e s L D.M e m b r a n e a n di n t e g r a t i v en u c l e a r f i b r o b l a s t i c g r o w t h f a c t o r r e c e p t o r(F G F R)r e g u l a t i o no fF G F-23[J].JB i o l C h e m,2015,290(16):10447-10459.[8] T a m a m u r a Y,S a k a m o t o K,K a t s u b e K I,e t a l.N o t c hs i g n a l i n g i s i n v o l v e d i nF G F23u p r e g u l a t i o n i no s t e o c y t e s[J].B i o c h e m B i o p h y sR e sC o mm u n,2019,518(2):233-238.[9] K a n e k o I,S a i n i R K,G r i f f i nK P,e t a l.F G F23g e n e r e g u l a t i o nb y1,25-d i h y d r o x y v i t a m i n D:o p p o s i n g e f f ec t s i nad i p o c y te sa n do s t e o c y t e s[J].JE n d o c r i n o l,2015,226(3):155-166.[10] K a l u d j e r o v i c J,K o m a b aH,S a t oT,e t a l.K l o t h oe x p r e s s i o ni n l o n g b o n e s r e g u l a t e sF G F23p r o d u c t i o nd u r i n g r e n a l f a i l u r e[J].F A S E BJ,2017,31(5):2050-2064.[11] M aL,G a oM,W uL,e t a l.T h e s u p p r e s s i v e e f f e c t o f s o l u b l eK l o t h oo n f i b r o b l a s t i c g r o w t h f a c t o r23s y n t h e s i s i nUM R-106o s t e o b l a s t-l i k e c e l l s[J].C e l lB i o lI n t,2018,42(9):1270-1274.㊃6411㊃‘临床荟萃“2019年12月20日第34卷第12期 C l i n i c a l F o c u s,D e c e m b e r20,2019,V o l34,N o.12Copyright©博看网. All Rights Reserved.[12]S a k a nH,N a k a t a n iK,A s a iO,e t a l.R e d u c e d r e n a lα-K l o t h oe x p r e s s i o n i nC K D p a t i e n t sa n di t sef f e c to nr e n a l p h o s p h a t eh a n d l i n g a n dv i t a m i nD m e t a b o l i s m[J].P L o S O n e,2014,9(1):e86301.[13] T s a iMH,L e uJ G,F a n g YW,e t a l.H i g hf i b r o b l a s t g r o w t hf a c t o r23l e v e l s a s s o c i a t e d w i t h l o w h e m og l o b i n l e v e l si np a t i e n t s w i t h c h r o n i c k i d n e y d i s e a s e s t a g e s3a n d4[J].M e d i c i n e(B a l t i m o r e),2016,95(11):e3049. [14] C o eL M,M a d a t h i lS V,C a s uC,e t a l.F G F-23i san e g a t i v er e g u l a t o r o f p r e n a t a la n d p o s t n a t a le r y t h r o p o i e s i s[J].JB i o lC h e m,2014,289(14):9795-9810.[15] A g o r oR,M o n t a g n aA,G o e t zR,e t a l.I n h i b i t i o n o f f i b r o b l a s tg r o w t h f a c t o r23(F G F23)s i g n a l i n g r e s c u e s r e n a l a n e m i a[J].F A S E BJ,2018,32(7):3752-3764.[16]S i n g hS,G r a b n e r A,Y a n u c i l C,e ta l.F i b r o b l a s t g r o w t hf a c t o r23d i r e c t l y t a rg e t sh e p a t o c y t e s t o p r o m o t ei n f l a mm a t i o ni n c h r o n i c k i d n e y d i s e a s e[J].K i d n e y I n t,2016,90(5):985-996.[17] C z a y aB,F a u lC.T h er o l eo fF i b r o b l a s t g r o w t hf a c t o r23i ni n f l a mm a t i o na n d a n e m i a[J].I n tJ M o lS c i,2019,20(17):E4195.[18] C h u n g B,V e r d i e r F,M a t a kP,e t a l.O n c o s t a t i nMi s a p o t e n ti n d u c e r o f h e p c i d i n,t h e i r o nr e g u l a t o r y h o r m o n e[J].F A S E BJ,2010,24(6):2093-2103.[19] A k c h u r i nO,S u r e s h b a b uA,D o t y S B,e t a l.L a c ko f h e p c i d i na m e l i o r a t e s a n e m i a a n d i m p r o v e s g r o w t h i n a n a d e n i n e-i n d u c e dm o u s e m o d e lo fc h r o n i ck i d n e y d i s e a s e[J].A m J P h y s i o lR e n a l P h y s i o l,2016,311(5):F877-F889.[20] O l a u s o nH,L i n d b e r g K,A m i nR,e t a l.P a r a t h y r o i d-s p e c i f i cd e l e t i o n o f K l o t h o u n r a v e l s a n o v e l c a l c i n e u r i n-d e p e n d e n tF G F23s i g n a l i n gp a t h w a y t h a tr e g u l a t e sP T H s e c r e t i o n[J].P L o SG e n e t,2013,9(12):e1003975.[21] G a l i t z e r H,B e n-D o vI Z,S i l v e rJ,e ta l.P a r a t h y r o i d c e l lr e s i s t a n c e t o f i b r o b l a s t g r o w t h f a c t o r23i n s e c o n d a r yh y p e r p a r a t h y r o i d i s mo f c h r o n i c k i d n e y d i s e a s e[J].K i d n e y I n t,2010,77(3):211-218.[22] Y a nJ,J i n g b o C,W a n g D,e ta l.A c o r r e l a t i o n b e t w e e nd e c r e a s e d p a r a t h y r o i dα-K l o t h oa n df i b r o b l a s t g r o w t hf a c t o rr e c e p t o r1e x p r e s s i o n w i t h p a t h o l o g i c a l c a t e g o r y a n dp a r a t h y r o i d g l a n d v o l u m ei n d i a l y s i s p a t i e n t s[J].I n t U r o lN e p h r o l,2015,47(4):701-706.[23] L e i f h e i t-N e s t l e r M,G r oβe S i e m e r R,F l a s b a r t K,e t a l.I n d u c t i o no fc a r d i a cF G F23/F G F R4e x p r e s s i o ni sa s s o c i a t e dw i t hl e f t v e n t r i c u l a r h y p e r t r o p h y i n p a t i e n t s w i t h c h r o n i ck i d n e y d i s e a s e[J].N e p h r o lD i a lT r a n s p l a n t,2016,31(7): 1088-1099.[24] G r a b n e rA,A m a r a lA P,S c h r a mm K,e ta l.A c t i v a t i o no fc a rd i a c f i b r o b l a s t g r o w t h f a c t o r re c e p t o r4c a u s e s l ef tv e n t r i c u l a r h y p e r t r o p h y[J].C e l lM e t a b,2015,22(6):1020-1032.[25] H uX,M aX,P a n X,e ta l.F i b r o b l a s t g r o w t hf a c t o r23i sa s s o c i a t e dw i t h t h e p r e s e n c e o f c o r o n a r y a r t e r y d i s e a s e a n d t h en u m b e r o f s t e n o t i cv e s s e l s[J].C l i nE x p P h a r m a c o lP h y s i o l, 2015,42(11):1152-1157.[26] N a k a h a r aT,K a w a i-K o w a s eK,M a t s u iH,e t a l.F i b r o b l a s tg r o w t hf a c t o r23i n h i b i t s o s t e o b l a s t i c g e n e e x p r e s s i o n a n di n d u c e so s t e o p r o t e g e r i ni nv a s c u l a rs m o o t h m u s c l ec e l l s[J].A t h e r o s c l e r o s i s,2016,253:102-110.[27]S c i a l l aJ J,L a u W L,R e i l l y M P,e ta l.F i b r o b l a s t g r o w t hf a c t o r23i sn o ta s s o c i a t e d w i t ha n dd o e sn o t i n d u c ea r t e r i a lc a l c i f i c a t i o n[J].K id ne y I n t,2013,83(6):1159-1168.[28] M u r a l i S K,R o s c h g e r P,Z e i t zU,e t a l.F G F23r e g u l a t e s b o n em i n e r a l i z a t i o n i na1,25(O H)2D3a n dk l o t h o-i n d e p e n d e n tm a n n e r[J].JB o n eM i n e rR e s,2016,31(1):129-142.[29] C a r r i l l o-Lóp e zN,P a n i z oS,A l o n s o-M o n t e sC,e ta l.D i r e c ti n h i b i t i o no fo s t e o b l a s t i c W n t p a t h w a y b y f i b r o b l a s t g r o w t hf a c t o r23c o n t r i b u t e s t o b o n e l o s s i n c h r o n i c k i d n e y d i s e a s e[J].K i d n e y I n t,2016,90(1):77-89.收稿日期:2019-11-23编辑:王秋红㊃7411㊃‘临床荟萃“2019年12月20日第34卷第12期 C l i n i c a l F o c u s,D e c e m b e r20,2019,V o l34,N o.12Copyright©博看网. All Rights Reserved.。
fgf23 基因

fgf23 基因FGF23基因是人体中的一种重要基因,它在调控骨骼系统和矿物质代谢方面起着重要的作用。
本文将从FGF23基因的结构与功能、与疾病的关联以及治疗方法等方面进行探讨。
FGF23基因编码一种成纤维细胞生长因子23(fibroblast growth factor 23),它主要由骨骼组织和肾小管上皮细胞合成。
FGF23基因编码的蛋白质具有促进肾小管对磷酸盐的排泄和抑制肠道对磷酸盐的吸收的作用,从而维持体内磷酸盐的平衡。
此外,FGF23基因编码的蛋白质还能抑制1,25-二羟维生素D的合成,降低肠道对钙的吸收,从而调节体内钙磷代谢。
近年来的研究发现,FGF23基因与一些疾病的发生和发展密切相关。
例如,肾性佝偻病是由于FGF23基因突变导致的一种常见的骨骼疾病。
在这种疾病中,FGF23基因突变导致FGF23蛋白质的功能异常,使其失去对磷酸盐的调控作用,从而导致体内磷酸盐的积累和骨骼的异常发育。
此外,FGF23基因还与慢性肾脏病相关,慢性肾脏病患者常常伴随着FGF23基因的异常表达,导致磷酸盐代谢紊乱和骨质疏松。
针对FGF23基因与疾病的关联,科学家们开展了大量的研究工作,并取得了一些重要的进展。
一方面,研究人员通过基因编辑技术,成功地纠正了FGF23基因的突变,恢复了FGF23蛋白质的正常功能,为肾性佝偻病等疾病的治疗提供了新的思路和方法。
另一方面,科学家们还发现,通过调节FGF23基因的表达水平,可以有效地调节磷酸盐和钙的代谢,为慢性肾脏病等疾病的治疗提供了新的途径。
除了疾病治疗方面,FGF23基因的研究还对骨骼系统和矿物质代谢的生理机制有着重要的启示。
通过深入研究FGF23基因的调控机制和信号转导途径,可以更好地理解骨骼系统和矿物质代谢的调节过程,为未来的疾病治疗和药物研发提供理论依据。
FGF23基因作为调控骨骼系统和矿物质代谢的重要基因,在疾病治疗和生理机制研究方面具有重要的意义。
通过对FGF23基因的深入研究,我们可以更好地理解疾病的发生和发展机制,并为疾病的预防和治疗提供新的思路和方法。
成纤维细胞生长因子23在骨矿物质代谢中的作用及其调控机理

成纤维细胞生长因子23在骨矿物质代谢中的作用及其调控机理董中亮;任晓曼;卜舒扬;闪爱婷;王玉婷;杨建成【摘要】骨源性激素成纤维细胞生长因子23(FGF23)介导由甲状旁腺、肾脏、骨骼和维生素D组成的负反馈回路,建立"骨骼-肾脏-甲状旁腺"内分泌轴,参与骨矿物质代谢并发挥重要作用.钙、磷、铁、维生素D、甲状旁腺素(PTH)、成纤维细胞生长因子受体(FGFR)/FGF以及蛋白质翻译后修饰调控FGF23的分泌、活性和胞内过程.随着深入的研究,探索出了一些以FGF23为靶点治疗骨矿物质代谢障碍疾病的新疗法.本文综述了FGF23在骨矿物质代谢中的作用及其调控机理的研究进展,以期为相关研究提供参考依据.%fibroblast growth factor?23 ( FGF23 ) as a bone?derived hormone plays an important role in bone mineral metabolism by mediating the negative feedback loops formed by parathyroid gland, kidney, bone and vitamin D to constructing abone?kidney?parathyroid gland endocrine axis. The secretion, activity and intracel?lular processes of FGF23 are regulated by calcium, phosphate, iron, vitamin D, parathyroid hormone ( PTH) , fibroblast growth factor receptor ( FGFR)/FGF and posttranslational modifications. With the increase of study on FGF23, the researchers have explored new therapeutic interventions on bone mineral metabolism disorder. In order to provide reference for relevant research, functions and regulation mechanism of FGF23 in bone min?eral metabolism were reviewed in this paper.【期刊名称】《动物营养学报》【年(卷),期】2017(029)010【总页数】5页(P3467-3471)【关键词】成纤维细胞生长因子23;骨矿物质代谢;钙;磷;铁【作者】董中亮;任晓曼;卜舒扬;闪爱婷;王玉婷;杨建成【作者单位】沈阳农业大学畜牧兽医学院,沈阳 110866;沈阳农业大学畜牧兽医学院,沈阳 110866;沈阳农业大学畜牧兽医学院,沈阳 110866;沈阳农业大学畜牧兽医学院,沈阳 110866;沈阳204医院,沈阳 110043;沈阳农业大学畜牧兽医学院,沈阳110866【正文语种】中文【中图分类】S852.2钙、磷是动物生长、骨骼发育和维持机能所必需的矿物质元素,是骨骼的基本组成成分,它们结合生成的羟基磷灰石构成骨盐。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
(生糖氨基酸) 生糖氨基酸)
氨基酸
α-酮酸 酮酸
糖
脂类代谢与蛋白质代谢的相互联系
甘油 脂肪 脂肪酸 乙酰CoA 乙酰 氨基酸碳架 氨基酸 蛋白质 磷酸二羟丙酮
蛋白质
氨基酸 酮酸或乙酰CoA 酮酸或乙酰 生酮氨基酸) (生酮氨基酸)
脂肪酸
脂肪
核酸与糖、脂类、蛋白质代谢的联系
核酸是细胞内重要的遗传物质,控制着蛋白质的合成, 核酸是细胞内重要的遗传物质,控制着蛋白质的合成,影响细 胞的成分和代谢类型 核酸生物合成需要糖和蛋白质的代谢中间产物参加, 核酸生物合成需要糖和蛋白质的代谢中间产物参加,而且需要 酶和多种蛋白质因子。 酶和多种蛋白质因子。 各类物质代谢都离不开具备高能磷酸键的各种核苷酸, 各类物质代谢都离不开具备高能磷酸键的各种核苷酸,如ATP 是能量的“通货” 此外 参与多糖的合成, 是能量的“通货”,此外UTP参与多糖的合成,CTP参与磷脂合 参与多糖的合成 参与磷脂合 参与蛋白质合成与糖异生作用。 成,GTP参与蛋白质合成与糖异生作用。 参与蛋白质合成与糖异生作用 核苷酸的一些衍生物具重要生理功能( 核苷酸的一些衍生物具重要生理功能(如CoA、NAD+,NADP+, 、 , , cAMP,cGMP)。 , )
第39章、细胞代谢调控 章
第一节 物质代谢的相互联系 第二节 代谢调节 第三节 基因表达调控
思考
?
第一节 物质代谢的相互联系
一、糖代谢与脂类代谢的相互关系 二、糖代谢与蛋白质代谢的相互联系 三、脂类代谢与蛋白质代谢的相互联系 核酸与糖、脂类、 四、核酸与糖、脂类、蛋白质代谢的联系
糖代谢与脂类代谢的相互联系
b、酶合成的诱导和阻遏 、酶合成的诱导和阻遏 实例:诱导型操纵子 实例:诱导型操纵子乳糖操纵子 色氨酸操纵子(自学) 阻遏型操纵子 色氨酸操纵子(自学)
A.有活性阻遏蛋白 有活性阻遏蛋白
调节基因 启动基因 操纵基因 结构基因 阻遏蛋白阻挡操纵基因 结构基因不表达
酶 的 诱 导 和 阻 遏 操 纵 子 模 型
CAP
cAMP -CAP
葡萄糖降解物与cAMP的关系 的关系 葡萄糖降解物与 ATP 腺苷酸 环化酶 cAMP 磷酸二 酯酶
抑制
葡萄糖 分解代 谢产物
激活
降低cAMP浓度 呈失活状态 cAMP 使CAP呈失活状态
CAP:降解物基因活化蛋白(catabolic :降解物基因活化蛋白( gene activation protein) )
(1)限速步骤和标兵酶 (2)前馈和反馈激活 (3)反馈抑制
(4)前馈和反馈调节中酶活性调节的机制
反馈调节中酶活性调节的机制
代谢物
别 构 中 心
活性 中心
共价修饰
酶分子中的某些基团,在其它酶的催化下, 酶分子中的某些基团,在其它酶的催化下, 可以共价结合或脱去,引起酶分子构象的改变, 可以共价结合或脱去,引起酶分子构象的改变, 使其活性得到调节, 使其活性得到调节,这种方式称为酶的共价修饰 )。目前已知有六种修饰方 (Covalent moldification )。目前已知有六种修饰方 磷酸化/去磷酸化 乙酰化/去乙酰化 去磷酸化, 去乙酰化, 式:磷酸化 去磷酸化,乙酰化 去乙酰化,腺苷酰 去腺苷酰化, 去尿苷酰化, 化/去腺苷酰化,尿苷酰化 去尿苷酰化,甲基化 去腺苷酰化 尿苷酰化/去尿苷酰化 甲基化/ 去甲基化, 去甲基化,氧化(S-S)/还原(2SH)。 ) 还原 。
反馈激活和前馈激活示意图
+ + E A→B → C → D → F → C E A→B → → G → D
+
例1:糖代谢途径中丙酮酸积累激活丙酮酸羧化酶, 1:糖代谢途径中丙酮酸积累激活丙酮酸羧化酶, 糖代谢途径中丙酮酸积累激活丙酮酸羧化酶 例2:乙酰CoA的积累激活PEP羧化酶 乙酰CoA的积累激活PEP羧化酶 CoA的积累激活PEP
第二节 代 谢 调 节
一、代谢调节的概念 二、细胞区域化调节 三、酶水平的调节 四、激素调节
代谢调节
生命是靠代谢的正常运转维持的。 生命是靠代谢的正常运转维持的。生命有限的空间内同 时有那麽多复杂的代谢途径在运转, 时有那麽多复杂的代谢途径在运转,必须有灵巧而严密的调 节机制, 节机制,才能使代谢适应外界环境的变化与生物自身生长发 育的需要。调节失灵便会导致代谢障碍, 育的需要。调节失灵便会导致代谢障碍,出现病态甚至危及 生命。在漫长的生物进化历程中,机体的结构、代谢和生理 生命。在漫长的生物进化历程中,机体的结构、 功能越来越复杂,代谢调节机制也随之更为复杂。 功能越来越复杂,代谢调节机制也随之更为复杂。 代谢调节的四级水平: 代谢调节的四级水平: 酶水平调节 细胞水平调节 激素水平调节 神经水平调节 多细胞整体水平调节
第三节 基因表达的调控
一、原核和真核基因组 二、原核生物酶合成调节的遗传机制 ——操纵子学说 操纵子学说 三、真核生物基因表达的调控
原核生物酶合成调节的遗传机制
操纵子学说
操纵子——基因表达的协同单位 a、操纵子 基因表达的协同单位
结构基因(编码蛋白质, ) 结构基因(编码蛋白质,S) 操纵子 控制部位 启动子( 启动子(premotor, P) ) 操纵基因( 操纵基因(operator, O) )
mRNAZ 乳糖 阻遏蛋白 有活性) (有活性) 阻遏蛋白 (无活性) 无活性)
mRNAY
mRNAa
基 因 表 达
乳糖操纵子的正调控
CAP 基因 R T CAP结 结 合部位 P RNA 聚合酶 mRNAZ mRNAY mRNAa O LacZ
结构基因
LacY Laca T 基 因 表 达
mRNA
内质网:蛋白质合成 内质网 蛋白质合成 ;磷脂合成 磷脂合成
酶活性的前馈和反馈调节
前馈( 和反馈( 前馈(feedforward )和反馈(feedback )是来自 电子工程学的术语,前者的意思是“输入对输出的影响” 电子工程学的术语,前者的意思是“输入对输出的影响”, 后者的意思是“输出对输入的影响” 后者的意思是“输出对输入的影响”,这里分别借用来说 明底物和代谢产物对代谢过程的调节作用。 明底物和代谢产物对代谢过程的调节作用。这种调节作用 是通过酶的变构效应 变构效应来实现的 是通过酶的变构效应来实现的。
A、乳糖操纵子的结构 、
乳 糖 操 纵 子 的 负 调 控
调节 基因 R mRNA
Байду номын сангаас
启 动 操纵 子 基因 P O LacZ
乳糖结构基因 LacY Laca
基 因 关 闭
阻遏蛋白 有活性) (有活性)
B、乳糖酶的诱导 、
调节 基因 R mRNA
启 动 操纵 子 基因 P O
乳糖结构基因 LacZ LacY Laca
5
磷酸化酶激酶(活性) 磷酸化酶激酶(活性)
ATP ADP
5、磷酸化酶 b
(无活性) 无活性) 磷酸化酶 a(活性) (活性)
×106
6
×108
葡萄糖
6、糖原
1-磷酸葡萄糖 磷酸葡萄糖
血液
葡萄糖
6-磷酸葡萄糖 磷酸葡萄糖
反馈抑制
由代谢终产物作为变构剂来抑制在此产 由代谢终产物作为变构剂来抑制在此产 变构剂 物合成过程中某一酶(通常为限速酶) 物合成过程中某一酶(通常为限速酶)活 性的作用,称为反馈抑制。 性的作用,称为反馈抑制。这是一种负反 馈机制,多数情况下控制合成代谢。 馈机制,多数情况下控制合成代谢。 类型: 类型:顺序反馈抑制 协同反馈抑制 累积反馈抑制 同工酶反馈抑制
脂肪代谢和糖代谢的关系
3-磷酸甘油 磷酸甘油 三酰甘油 三酰甘油 脂肪酸 甘油
糖原(或淀粉) 糖原(或淀粉) 1,6-二磷酸果糖 , 二磷酸果糖 磷酸二羟丙酮
β 氧 化
合 成
丙酮 酸
磷酸烯醇丙酮酸
草酰乙酸
乙酰 CoA 植物或微 生物
三羧酸 循环
苹果酸
乙醛酸 循环
延胡索酸
琥珀酸
糖的分解代谢和 糖异生的关系
酶水平的调节
1、酶原激活 2、酶的别构效应 酶活性的前馈和反馈调节 3、酶的共价修饰与级联放大机制 4、辅因子对已有酶活性的调节
酵解;磷 细胞质:酵解 磷 戊糖途径;糖原 戊糖途径 糖原 合成;脂肪酸合 合成 脂肪酸合 成;
酶 定 位 的 区 域 化
细胞核:核酸合成 细胞核 核酸合成
线粒体:丙酮酸氧化 三羧 线粒体 丙酮酸氧化;三羧 丙酮酸氧化 酸循环;β 氧化 氧化;呼吸链电 酸循环 β-氧化 呼吸链电 子传递;氧化磷酸化 子传递 氧化磷酸化
葡萄糖
1,6-二磷酸果糖 , 二磷酸果糖
酶 的 反 馈 激 活
磷酸烯醇式丙酮酸 羧化酶 活化 草酰乙酸
丙酮酸
乙酰CoA 乙酰 α-酮戊二酸 酮戊二酸 拧檬酸
谷氨酰氨合成酶的累积反馈抑制
辅因子对已有酶活性的调节
能荷对代谢的调节 [NADH]/[NAD+]对代谢的调节 金属离子浓度对代谢的调节 金属离子浓度对代谢的调节
(PEP) ) 丙氨酸 (胞液) 胞液) (线粒体) 线粒体) 天冬氨酸
(转氨基作用) 转氨基作用)
谷氨酸
蛋白质
核酸
核苷酸
淀粉、 淀粉、糖原
1-磷酸葡萄糖 磷酸葡萄糖 6-磷酸葡萄糖 磷酸葡萄糖
脂肪
糖 类 脂 类 氨 基 酸 和 核 苷 酸 之 间 的 代 谢 联 系
氨基酸
生糖氨基酸
甘氨酸 天冬氨酸 谷氨酰氨 丙氨酸 甘氨酸 丝氨酰 苏氨酸 半胱氨酸 天冬氨酸 天冬酰氨 酪氨酸 天冬氨酸 苯丙酰氨 异亮氨酸 甲硫酰氨 苏氨酸 缬氨酸 谷氨酸 谷氨酰氨 组氨酸 脯氨酸 精氨酸
例:糖原磷酸化酶的共价修饰
ATP
激酶 磷酸酯酶