公示 - 福建农林大学科学技术发展研究院
福建农林大学2013年拟推荐免试硕士研究生名单xls
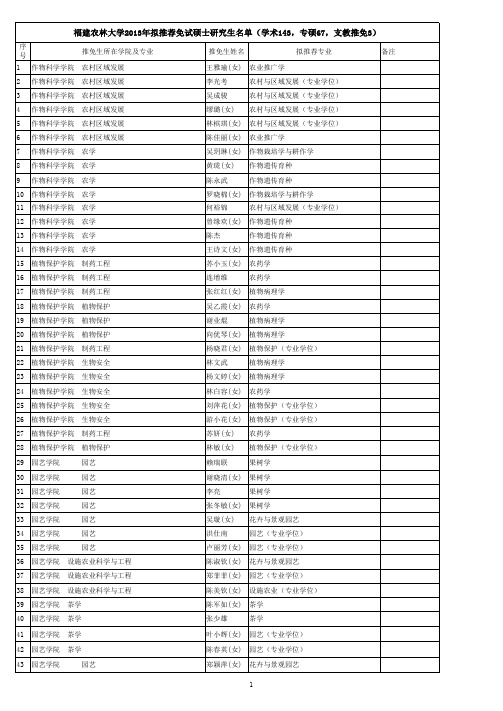
机电工程学院 机械设计制造及其自动化(现代设计) 罗龙和 机电工程学院 机械设计制造及其自动化(现代设计) 钟寿洋 机电工程学院 电气工程及其自动化 机电工程学院 电气工程及其自动化 机电工程学院 电子科学与技术 机电工程学院 电子科学与技术 机电工程学院 工业工程 机电工程学院 工业工程 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生命科学学院 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物科学(基) 生物信息学 生物工程 生物工程 生物工程 生物技术 林魁 黄诚杰 曹立
100 食品科学学院 101 食品科学学院 102 食品科学学院 103 食品科学学院 104 食品科学学院 105 食品科学学院 106 食品科学学院 107 食品科学学院 108 食品科学学院 109 食品科学学院 110 食品科学学院 111 林学院 112 林学院 113 林学院 114 林学院 115 林学院 116 林学院 林学 林学 林学 林学
福建农林大学2013年拟推荐免试硕士研究生名单(学术143,专硕67,支教推免3)
序 号 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 经济与管理学院 推免生所在学院及专业 文化产业管理 文化产业管理 工商管理 工商管理 行政管理 行政管理 旅游管理 旅游管理 会计学 会计学 金融学 金融学 国际经济与贸易 国际经济与贸易 农林经济管理 农林经济管理 农林经济管理 农林经济管理 农林经济管理 农林经济管理 工商管理 推免生姓名 谢翠玲(女) 周淑娟(女) 郑小丹(女) 陈建铃(女) 朱婷婷(女) 傅美兰(女) 苏玉卿(女) 庄惠(女) 李秀容(女) 方晓婉(女) 欧聪丽(女) 熊荣桢(女) 陈燕煌(女) 叶梅红(女) 陈翠蓉(女) 陈思莹(女) 吴骏莹(女) 叶菁菁(女) 黄朝宗 钟美玉(女) 郑小清(女) 旅游管理 农业科技组织与服务(专业学位) 企业管理 技术经济与管理 公共管理 农业科技组织与服务(专业学位) 旅游管理 农业科技组织与服务(专业学位) 会计学 会计学 应用经济学 农业科技组织与服务(专业学位) 应用经济学 农业科技组织与服务(专业学位) 农业经济管理 林业经济管理 林业经济管理 农业经济管理 人口、资源与环境经济学 农业科技组织与服务(专业学位) 企业管理 机械设计及理论 机械设计及理论 农业工程(专业学位) 农业电气化与自动化 农业工程(专业学位) 拟推荐专业 备注
福建农林大学研究生教育管理系统

福建农林大学研究生教育管理系统福建农林大学研究生教育管理系统是一种基于现代信息技术的教育管理工具。
该系统旨在提供全面的教育服务,方便研究生的学习和生活,促进教学和科研的高效运作。
本文将从系统的设计背景、功能模块、特点和应用效果等方面详细介绍福建农林大学研究生教育管理系统。
福建农林大学研究生教育管理系统的设计背景是为了满足高校研究生教育管理的需求。
随着我国高等教育的迅速发展,研究生教育在高等教育体系中占据重要地位。
研究生教育具有专业性强、知识深度高等特点,因此需要一个专门的管理系统来提供个性化、便捷的服务。
福建农林大学研究生教育管理系统的功能模块包括学籍管理、课程管理、导师管理、科研管理、学位管理、学业指导、就业服务等。
学籍管理模块主要包括学生信息的录入、修改和查询,以及学籍变更的申请和审核等功能。
课程管理模块可以帮助学生选课、查询课程信息、统计学分等。
导师管理模块提供导师信息的管理、学生指导记录的管理等功能。
科研管理模块可以帮助学生上传和下载科研文档、管理科研项目等。
学位管理模块主要包括学位申请和学位评定等功能。
学业指导模块可以提供学生论文选题、撰写和答辩等服务。
就业服务模块可以提供就业信息发布、就业指导等功能。
福建农林大学研究生教育管理系统的特点包括智能化、个性化、网络化和安全性等。
系统具有智能化特点,可以根据学生的需求提供相应的服务,如个性化选课推荐、学业指导建议等。
系统支持网络化操作,学生可以通过网络随时随地访问系统,进行相关操作。
系统具有一定的安全性,只有授权人员才能访问和修改系统数据。
福建农林大学研究生教育管理系统的应用效果主要体现在教学管理的规范化和便捷化,以及学生学习和生活的便利性。
系统通过优化教学管理流程,提高了教育管理的效率和精确度,减少了人工操作的出错率。
学生可以通过系统方便地查询课程信息、教师信息、科研文献等,提高了学习效率。
系统还可以提供个性化的学业指导和就业服务,帮助学生更好地规划未来发展。
东方蜜蜂微孢子虫nce-miR-11248及其靶基因SGT1的生物信息学和表达谱研究

第52卷 第2期2024年2月西北农林科技大学学报(自然科学版)J o u r n a l o f N o r t h w e s t A&F U n i v e r s i t y(N a t .S c i .E d .)V o l .52N o .2F e b .2024网络出版时间:2023-08-04 18:06 D O I :10.13207/j .c n k i .jn w a f u .2024.02.004网络出版地址:h t t ps ://l i n k .c n k i .n e t /u r l i d /61.1390.S .20230804.1516.004东方蜜蜂微孢子虫n c e -m i R -11248及其靶基因S G T 1的生物信息学和表达谱研究[收稿日期] 2022-11-01 [基金项目] 国家自然科学基金面上项目(32172792);国家现代农业产业技术体系建设专项(C A R S -44-K X J 7);福建农林大学硕士生导师团队项目(郭睿);福建农林大学动物科学学院(蜂学学院)科研扶持项目(郭睿);福建省大学生创新创业训练计划项目(202110389027) [联系方式] 张凯遥(1998-),女,福建福州人,在读研士,主要从事蜜蜂分子生物学研究㊂E -m a i l :k a i ya o 1223@126.c o m [通信作者] 牛庆生(1969-),男,吉林长春人,研究员,主要从事蜜蜂种质资源研究㊂E -m a i l :a pi s 1969@163.c o m 郭 睿(1987-),男,安徽六安人,副教授,主要从事蜜蜂分子生物学及组学研究㊂E -m a i l :r u i gu o @f a f u .e d u .c n 张凯遥1,赵浩东1,张艺琼1,赵 萧1,高旭泽1,邹培缘1,张奎昊1,陈大福1,2,郭 睿1,2,牛庆生3(1福建农林大学动物科学学院(蜂学学院),福建福州350002;2福建省蜂疗研究所,福建福州350002;3吉林省养蜂科学研究所,吉林吉林132000)[摘 要] ʌ目的ɔ对前期鉴定所获得的东方蜜蜂微孢子虫的n c e -m i R -11248进行表达和序列验证,预测㊁分析其靶基因,并检测n c e -m i R -11248及其靶基因在东方蜜蜂微孢子虫侵染意大利蜜蜂工蜂过程中的表达谱,为进一步开展n c e -m i R -11248调控东方蜜蜂微孢子虫侵染的功能及作用机制研究提供参考㊂ʌ方法ɔ利用S t e m -l o o p RT -P C R 和S a n g e r 测序验证n c e -m i R -11248的真实性;利用相关生物信息学软件预测n c e -m i R -11248的靶基因,并分析其编码蛋白的分子特性;使用M E M E 和T B t o o l s 软件预测S G T 1蛋白的保守基序和结构域,通过M e ga 11.0软件进行氨基酸序列多重比对,采用邻接法构建系统进化树㊂采集东方蜜蜂微孢子虫孢子侵染1,2,4,6和8d 的意蜂工蜂中肠组织,采用R T -qP C R 检测n c e -m i R -11248及其靶基因S G T 1的表达谱㊂ʌ结果ɔn c e -m i R -11248在东方蜜蜂微孢子虫孢子中真实存在,其序列长度为25b p ㊂n c e -m i R -11248的靶基因为S G T 1㊂S G T 1蛋白分子式为C 765H 1213N 195O 241S 4,分子质量约为17.10k u ,脂溶系数为82.67,等电点为5.50,亲水系数为-0.709,不含典型的信号肽和跨膜结构域,可同时定位于细胞核㊁线粒体和过氧化物酶体㊂东方蜜蜂微孢子虫㊁家蚕微孢子虫㊁泛胞虫㊁肠脑炎微孢子虫和蚱蜢脑炎微孢子虫的S G T 1中均含有4个保守基序(M o t i f 1㊁M o t i f 2㊁M o t i f 3和M o t i f 4)和1个结构域(S G T 1超家族结构域)㊂东方蜜蜂微孢子虫㊁家蚕微孢子虫㊁肠脑炎微孢子虫和蚱蜢脑炎微孢子虫的S G T 1聚为一支,且东方蜜蜂微孢子虫与家蚕微孢子虫的S G T 1进化距离最近㊂相较于侵染东方蜜蜂微孢子虫后1d ,侵染后2,4,6和8d n c e -m i R -11248的表达量均显著下调(P <0.05);侵染后2,4,6和8d S G T 1的表达量均下调,其中侵染后4,6和8d 的表达量与2d 的差异达显著水平(P <0.05)㊂ʌ结论ɔn c e -m i R -11248在东方蜜蜂微孢子虫孢子中真实存在;东方蜜蜂微孢子虫与家蚕微孢子虫的S G T 1亲缘关系最近;随着侵染时间的延长,n c e -m i R -11248及其靶基因S G T 1在东方蜜蜂微孢子虫侵染意大利蜜蜂工蜂过程中的表达量均呈持续下降;n c e -m i R -11248和S G T 1是潜在的参与调控微孢子虫侵染过程的因子㊂[关键词] 东方蜜蜂微孢子虫;意大利蜜蜂;n c e -m i R -11248;S G T 1基因;表达谱[中国分类号] S 895.2+36[文献标志码] A[文章编号] 1671-9387(2024)02-0032-08S t u d y o n b i o i n f o r m a t i c s a n d e x pr e s s i o n p a t t e r n o f n c e -m i R -11248a n d t a r g e t ge n e S G T 1i n N o s e m a c e r a n a e Z H A N G K a i y a o 1,Z H A O H a o d o n g 1,Z H A N G Y i q i o n g 1,Z H A O X i a o 1,G A O X u z e 1,Z O U P e i y u a n 1,Z H A N G K u i h a o 1,C H E N D a f u 1,2,G U O R u i 1,2,N I U Q i n g s h e n g3(1C o l l e g e o f A n i m a l S c i e n c e s (C o l l e g e o f B e e S c i e n c e ),F u j i a n A g r i c u l t u r e a n d F o r e s t r y U n i v e r s i t y ,F u ji a n ,F u z h o u 350002,C h i n a ;2A p i t h e r a p y R e s e a r c h I n s t i t u t e o f F u j i a n P r o v i n c e,F u j i a n,F u z h o u350002,C h i n a;3A p i c u l t u r e S c i e n c e I n s t i t u t e o f J i l i n P r o v i n c e,J i l i n,J i l i n132000,C h i n a)A b s t r a c t:ʌO b j e c t i v eɔI n t h i s s t u d y,p r e v i o u s l y i d e n t i f i e d n c e-m i R-11248i n N o s e m a c e r a n a e w a s e x-p r e s s e d a n d v e r i f i e d,i t s t a r g e t g e n e w a s p r o j e c t e d a n d a n a l y z e d,a n d t h e i r e x p r e s s i o n p a t t e r n s d u r i n g t h e N o s e m a c e r a n a e i n f e c t i o n t o A p i s m e l l i f e r a l i g u s t i c a w o r k e r s w e r e a s s e s s e d,a i m i n g t o p r o v i d e r e f e r e n c e s f o r f u r t h e r e x p l o r a t i o n o f t h e m e c h a n i s m u n d e r l y i n g N.c e r a n a e i n f e c t i o n r e g u l a t e d b y n c e-m i R-11248.ʌM e t h o dɔS t e m-l o o p R T-P C R a n d S a n g e r s e q u e n c i n g w e r e u s e d t o p r o v e t h e a u t h e n t i c i t y o f n c e-m i R-11248.R e l a t e d b i o i n f o r m a t i c s o f t w a r e w e r e e m p l o y e d t o p r e d i c t i t s t a r g e t g e n e a n d a n a l y z e m o l e c u l a r c h a-r a c t e r i s t i c s o f t h e e n c o d e d p r o t e i n.M E M E a n d TB t o o l s w e r e u t i l i z e d t o p r e d i c t c o n s e r v e d m o t i f s a n d s t r u-c t u r a ld o m a i n s i n t he e n c o d e p r o t e i n.M u l t i p l e a l i g n m e n t of a m i n o a c i d s e q u e n c e s w a s c o n d u c t e d b y M eg a11.0s o f t w a r e,f o l l o w e d b y c o n s t r u c t i o n o f p h y l o g e n e t i c t r e e w i t h t h e n e i g h b o r-j o i n i n g m e t h o d.T h e m i d g u t t i s s u e s o f A.m.l i g u s t i c a w o r k e r s a t1,2,4,6a n d8d p o s t N.c e r a n a e i n f e s t a t i o n w e r e c o l l e c t e d.R T-q P C R w a s u s e d t o d e t e r m i n e e x p r e s s i o n p r o f i l e s o f n c e-m i R-11248a n d t h e t a r g e t g e n e.ʌR e s u l tɔI t w a s p r o v e d t h a t n c e-m i R-11248w a s t r u l y e x i s t e d i n N.c e r a n a e s p o r e s,a n d t h e s e q u e n c e l e n g t h w a s25b p.T h e t a r g e t g e n e o f n c e-m i R-11248w a s S G T1.T h e m o l e c u l a r f o r m u l a o f S G T1w a s C765H1213N195O241S4w i t h m o l e c u l a r w e i g h t o f a p p r o x i m a t e l y17.10k u,i t s l i p o l y s i s c o e f f i c i e n t,i s o e l e c t r i c p o i n t a n d h y d r o p h i l i c c o e f f i c i e n t w e r e 82.67,5.50a n d-0.709,r e s p e c t i v e l y.T h e r e w a s n o c l a s s i c a l s i g n a l p e p t i d e a n d t r a n s m e m b r a n e d o m a i n i n S G T1,a n d i t s i m u l t a n e o u s l y l o c a t e d i n n u c l e u s,m i t o c h o n d r i o n a n d p e r o x i s o m e.A d d i t i o n a l l y,S G T1p r o-t e i n s i n N.c e r a n a e,N o s e m a b o m b y c i s,P a n c y t o s p o r a,E n c e p h a l i t o z o o n i n t e s t i n a l i s a n d E n c e p h a l i t o z o o n r o m a l e a e c o n t a i n e d f o u r c o n s e r v e d m o t i f s o f M o t i f1,M o t i f2,M o t i f3a n d M o t i f4a n d o n e s t r u c t u r a l d o-m a i n o f S G T1s u p e r f a m i l y.F u r t h e r m o r e,S G T1p r o t e i n s i n N.c e r a n a e,N.b o m b y c i s,E.i n t e s t i n a l i s a n d E.r o m a l e a e w e r e g r o u p e d i n t o o n e c l a d e,a n d N.c e r a n a e S G T1a n d N.b o m b y c i s S G T1h a d t h e c l o s e s t e v o-l u t i o n a r y d i s t a n c e.A s c o m p a r e d t o1d a y p o s t N.c e r a n a e i n o c u l a t i o n,t h e e x p r e s s i o n l e v e l o f n c e-m i R-11248w a s s i g n i f i c a n t l y d o w n-r e g u l a t e d a t2,4,6a n d8d a y s p o s t i n o c u l a t i o n(P<0.05),t h e e x p r e s s i o n l e v e l o f S G T1w a s d o w n-r e g u l a t e d a t2,4,6a n d8d a y s p o s t i n o c u l a t i o n,a n d t h e d i f f e r e n c e s i n e x p r e s s i o n l e v e l a t4,6a n d8d a y s p o s t i n o c u l a t i o n r e a c h e d s i g n i f i c a n t l e v e l s(P<0.05).ʌC o n c l u s i o nɔT h i s s t u d y s h o w e d t h a t n c e-m i R-11248w a s t r u l y e x i s t e d i n N.c e r a n a e s p o r e s.N.c e r a n a e S G T1a n d N.b o m b y c i s S G T1h a d t h e c l o s e s t g e n e t i c d i s t a n c e.B o t h n c e-m i R-11248a n d t a r g e t g e n e S G T1p r e s e n t e d c o n t i n u o u s d e-c r e a s e s d u r i n g t h e N.c e r a n a e i n f e c t i o n t o A.m.l i g u s t i c a w o r k e r s,a n d n c e-m i R-11248a n d S G T1w e r e p o-t e n t i a l f a c t o r s i n v o l v e d i n m o d u l a t i o n o f t h e m i c r o s p o r i d i a n i n f e c t i o n p r o c e s s.K e y w o r d s:N o s e m a c e r a n a e;A p i s m e l l i f e r a l i g u s t i c a;n c e-m i R-11248;S G T1g e n e;e x p r e s s i o n p a t t e r n意大利蜜蜂(A p i s m e l l i f e r a l i g u s t i c a)简称意蜂,是西方蜜蜂(A p i s m e l l i f e r a)的一个优良亚种,具有采集力强㊁不易逃群和性情温顺等优良特性,因而在养蜂生产中被广泛饲养[1]㊂东方蜜蜂微孢子虫(N o s e m a c e r a n a e)是一种专性寄生成年蜜蜂中肠上皮细胞的真菌病原,不仅能导致蜜蜂细胞结构破坏㊁能量胁迫㊁免疫抑制㊁细胞凋亡和寿命缩短[2],还能与其他生物或非生物因子一起危害蜂群健康[3]㊂m i R N A是一类较为保守的内源性小非编码R N A (n o n-c o d i n g R N A,n c R N A),可在转录后水平调节基因表达,从而在众多生物学过程中行使重要的调控功能[4]㊂动㊁植物的m i R N A研究起步较早且较为深入㊂B a i等[5]研究发现,过表达m i R-1269可以促进食管鳞癌癌细胞的增殖㊁迁移和侵袭㊂Y a n g 等[6]研究发现,在水稻(O r y z a s a t i v a L.)中过表达O s a-M I R319a或O s a-M I R319b可以改变叶片形态增加叶面积㊂在果蝇(D r o s o p h i l a)中,m i R-7通过抑制下游N o t c h受体来限制N o t c h信号转导,进而缓冲视叶发育过程中神经上皮细胞向神经母细胞的转变[7]㊂过表达m i R-14会导致家蚕(B o m b y x m o r i)幼虫发育延迟,幼虫和蛹体积变小,蜕皮激素滴度降低[8]㊂与动㊁植物相比,真菌m i R N A的研究较为滞后㊂随着深度测序技术与生物信息学的快速发展,脉孢菌(N e u r o s p o r a)[9]㊁新型隐球菌(C r y p-33第2期张凯遥,等:东方蜜蜂微孢子虫n c e-m i R-11248及其靶基因S G T1的生物信息学和表达谱研究t o c o c c u s n e o fo r m a n s )[10]和阿氏丝孢酵母(T r i c h o s -po r o n a s a h i i )[11]等真菌的m i R N A 陆续被鉴定出来㊂目前,已鉴定的东方蜜蜂微孢子虫m i R N A 非常有限,有研究人员通过二代测序和生物信息学预测到东方蜜蜂微孢子虫的9个m i R N A[12-13]㊂近期,笔者所在研究团队通过s m a l l R N A -s e q 在东方蜜蜂微孢子虫孢子中鉴定到n c e -m i R -11248㊁n c e -m i R -23928㊁n c e -m i R -12220㊁n c e -m i R -10660㊁n c e -m i R -9769㊁n c e -m i R -15338㊁n c e -m i R -34537㊁n c e -m i R -9172㊁n c e -m i R -15325㊁n c e -m i R -26675等10个m i R N A [14],且通过试验证实了n c e -m i R -23928和n c e -m i R -12220的真实性,并分析了其表达水平[15-16]㊂本研究在前期试验基础上对n c e -m i R -11248进行克隆㊁测序,预测和分析其靶基因,并检测n c e -m i R -11248及其靶基因在东方蜜蜂微孢子虫侵染意蜂工蜂过程的表达谱,以期为进一步开展n c e -m i R -11248调控东方蜜蜂微孢子虫侵染的功能及作用机制研究提供参考依据㊂1 材料与方法1.1 生物材料试验于2021年12月至2022年8月在福建农林大学动物科学院(蜂学学院)蜜蜂保护实验室进行㊂东方蜜蜂微孢子虫孢子,由福建农林大学动物科学学院(蜂学学院)蜜蜂保护实验室制备和保存[17,14]㊂未受东方蜜蜂微孢子虫感染的意蜂工蜂,取自福建农林大学动物科学学院(蜂学学院)教学蜂场的饲养蜂群㊂东方蜜蜂微孢子虫感染的意蜂工蜂,取自福州市闽侯县荆溪源安蜂场的饲养蜂群㊂1.2 n c e -m i R -11248的S t e m -l o o p RT -P C R 克隆和S a n ge r 测序根据前期预测到的n c e -m i R -11248序列(5'-U A U G G A U C A G U G G C A U U U U U A U G C C -3')[14],利用D N AMA N 软件设计特异性引物n c e -m i R -11248-L o o p 和nc e -m i R -11248,引物序列见表1,委托上海生工生物工程股份有限公司合成㊂按照本团队建立的技术流程[15-16],采用S t e m -l o o p RT -P C R 方法进行东方蜜蜂微孢子虫孢子样品总R N A 提取㊁c D N A合成(引物为n c e -m i R -11248-L o o p )及nc e -m i R -11248的P C R 扩增(引物为n c e -m i R -11248)㊂对扩增产物进行1.5%琼脂糖凝胶电泳检测㊂参照范元婵等[18]的方法回收目的片段并进行T A 克隆,采用P C R 方法筛选阳性克隆㊂委托上海生工生物工程股份有限公司对阳性克隆菌液进行S a n ge r 测序㊂表1 试验所用引物序列T a b l e 1 S e q u e n c e s o f p r i m e r s u s e d i n t h i s s t u d y引物P r i m e r序列(5'ң3')S e qu e n c e n c e -m i R -11248-L o o p C T C A A C T G G T G T C G T G G A G T C G G C A A T T C A G T T G A G G G C A T A A An c e -m i R -11248F :CG T A T G G A T C A G T G G C A T T ,R :C T C A A C T G G T G T C G T G G A 5S r R N AF :CG A G C G G T T T C C C A T C T C A G T A ,R :A A A A C A C C G G A A C T C G T C A G C T S G T 1F :G A C A C T T C T C G T A T T T C A C T ,R :T T C C A T T G C T C T T C T T G T Gβ-a c t i n F :A C A A T G G T T C A G G T A T C G T A ,R :G T G C C T C A T C T C C T A C A T A A1.3 东方蜜蜂微孢子虫侵染和工蜂中肠样品制备按照本团队建立的技术流程[19]进行东方蜜蜂微孢子虫孢子侵染与工蜂中肠样品制备:将群势较强的试验蜂群中即将出房的封盖子脾迅速提至实验室,放入(34ʃ0.5)ħ恒温恒湿箱,收集刚出房的1日龄工蜂放入干净塑料盒(35只/盒);饥饿处理2h 后单头工蜂饲喂侵染东方蜜蜂微孢子虫孢子的5μL 500g /L 蔗糖溶液(含有106个孢子);塑料盒上方插入一支内含2m L 蔗糖溶液的饲喂器,每24h 更换一支饲喂器,同时清理死蜂;分别于侵染后1,2,4,6和8d 在超净工作台中拉取工蜂中肠,每3只工蜂的中肠放入1个R N A a s e -f r e e 离心管,液氮速冻后放入-80ħ超低温冰箱保存备用㊂1.4 n c e -m i R -11248的表达谱检测以5S r R N A 为内参,采用R T -qP C R 检测n c e -m i R -11248的表达谱㊂参考G e n B a n k 中的5Sr R N A 序列(G e n B a n k I D :E F 091880.1)设计引物(表1),委托上海生工生物工程股份有限公司合成㊂利用R N A 抽提试剂盒(美国普洛麦格公司)分别提取上述5个时间点中肠样品的总R N A ,反转录得到相应的c D N A (引物为n c e -m i R -11248-L o o p )㊂以c D N A 为模板,在全自动P C R 分析系统(G e n t i e r 96R )上进行n c e -m i R -11248及内参5S r R N A 的R T -qP C R ,参照吴鹰等[16]的报道设置反应体系和程序㊂试验设3次技术重复和3次平行重复㊂以侵染后1d 的表达水平为参照,采用2-ΔΔC t法计算n c e -m i R -11248在侵染后2,4,6和8d 的相对表达量㊂1.5 n c e -m i R -11248的靶基因预测笔者团队前期已利用R N A -s e q 技术对东方蜜蜂微孢子虫的纯净孢子样品进行深度测序,获得了43西北农林科技大学学报(自然科学版)第52卷高质量的测序数据[14]㊂在此基础上,本研究利用T a r g e t F i n d e r软件(v1.6)预测n c e-m i R-11248的靶基因㊂将预测所获靶基因序列在S w i s s-P r o t(h t-t p://w w w.u n i p r o t.o r g/u n i p r o t/?q u e r y=*&f i l =r e v i e w e d%3A y e s)㊁N r(f t p://f t p.n c b i.n i h.g o v/ b l a s t/d b)㊁K E G G(h t t p s://w w w.o m i c s h a r e.c o m/ t o o l s/H o m e/S o f t/p a t h w a y g s e a)和G O(h t t p s:// w w w.o m i c s h a r e.c o m/t o o l s/H o m e/S o f t/g o g s e a)数据库中进行B L A S T比对,从而获得靶基因相应功能注释㊂1.6n c e-m i R-11248的靶蛋白生物信息学分析根据1.5节n c e-m i R-11248的靶基因预测结果,并结合东方蜜蜂微孢子虫的生物学背景[20-21]筛选到1个关键靶基因S G T1㊂利用N C B I网站(h t-t p s://w w w.n c b i.n l m.n i h.g o v/)上的O R F工具预测n c e-m i R-11248靶基因编码蛋白S G T1的氨基酸序列㊂通过E x p a s y网站(h t t p s://w w w.e x p a s y. o r g/)上的P r o t p a r a m㊁P r o t S c a l e和S W I S S-m o d e l等软件分析靶蛋白的理化性质㊁亲水性和三级结构㊂分别利用S i g n a l P4.1S e r v e r㊁N e t P h o s3.1S e r v e r㊁T M-HMM及S O P M A等软件预测靶蛋白S G T1的信号肽㊁磷酸化位点㊁跨膜结构域及二级结构㊂使用P S O R TⅡ软件预测靶蛋白S G T1的亚细胞定位㊂所有分析软件的参数均采用默认值㊂使用B L A S T工具将靶蛋白S G T1的氨基酸序列比对到N C B I G e n B a n k数据库(h t t p s://w w w.n c b i. n l m.n i h.g o v/g e n b a n k/),以搜索序列相似性较高的其他物种的相应蛋白,利用M E M E软件(h t t p s:// m e m e-s u i t e.o r g/)预测其保守基序,比对参数为:最小和最大结构域的氨基酸残基数分别为6和50,不同结构域的数量为5,其余参数为默认值㊂采用N C B I网站上的C o n s e r v e d D o m a i n s S e a r c h工具预测靶蛋白S G T1的结构域,并采用T B t o o l s软件(v.1.098761)进行分析㊂利用M e g a 11.0软件(v.1.098761)对东方蜜蜂微孢子虫和其他物种的相关蛋白进行氨基酸序列多重比对,并采用邻接法构建系统进化树,参数为默认值㊂1.7n c e-m i R-11248的靶基因表达谱测定以β-a c t i n作为内参,通过R T-q P C R检测n c e-m i R-11248靶基因S G T1在东方蜜蜂微孢子虫侵染过程的相对表达量㊂根据n c e-m i R-11248靶基因S G T1序列及G e n B a n k中β-a c t i n序列(G e n B a n k I D:36320842)设计特异性引物(表1),委托上海生工生物工程股份有限公司合成㊂参照张文德等[15]的方法进行R T-q P C R,试验设置3次技术重复和3次平行重复㊂以侵染后1d n c e-m i R-11248的靶基因S G T1表达量为参照,采用2-ΔΔC t法计算侵染后2, 4,6和8d时n c e-m i R-11248的靶基因相对表达量㊂1.8数据处理与分析采用G r a p h P a d P r i s m8软件(v.8.0.2)进行数据分析并绘图㊂通过S P S S软件(v.26)进行单因素方差分析(A N O V A),以P<0.05为显著性阈值㊂采用T u k e y检验法和字母显著标记法对试验数据进行两两比较分析㊂2结果与分析2.1n c e-m i R-11248的分子验证琼脂糖凝胶电泳结果(图1-A)显示,n c e-m i R-11248R T-P C R产物约为100b p,符合预期大小(L o o p引物长度(44b p)+n c e-m i R-11248引物长度(37b p)+n c e-m i R-11248序列长度(25b p))㊂S a n-g e r测序结果(图1-B)显示,n c e-m i R-11248片段序列长度为25b p,与前期基于转录组测序数据预测出的n c e-m i R-11248序列长度一致㊂以上结果表明, n c e-m i R-11248在东方蜜蜂微孢子虫孢子中是真实存在的㊂A.n c e-m i R-11248扩增产物电泳图;B.n c e-m i R-11248的S a n g e r测序㊂M.D N A M a r k e r;1.n c e-m i R-11248扩增产物A.E l e c t r o p h o r e s i s o f a m p l i f i c a t i o n p r o d u c t f r o m n c e-m i R-11248;B.S a n g e r s e q u e n c i n g o f p r o d u c t f r o m n c e-m i R-11248.M.D N A M a r k e r;1.A m p l i f i c a t i o n p r o d u c t o f n c e-m i R-11248图1东方蜜蜂微孢子虫n c e-m i R-11248的分子验证F i g.1 M o l e c u l a r v a l i d a t i o n o f n c e-m i R-11248i n N o s e m a c e r a n a e 53第2期张凯遥,等:东方蜜蜂微孢子虫n c e-m i R-11248及其靶基因S G T1的生物信息学和表达谱研究2.2 n c e -m i R -11248的表达谱R T -qP C R 结果(图2)显示,相较于侵染后1d ,侵染后2,4,6和8d n c e -m i R -11248的表达量均显著下调(P <0.05),分别下调为侵染后1d 时的22.80%,0.76%,0.02%和0.01%㊂图柱上标不同小写字母表示差异显著(P <0.05)㊂图7同D i f f e r e n t l o w e r c a s e l e t t e r s i n d i c a t e s i gn i f i c a n t d i f f e r e n c e (P <0.05).F i g.7a s s a m e 图2 东方蜜蜂微孢子虫侵染过程中n c e -m i R -11248的表达谱F i g .2 E x p r e s s i o n p a t t e r n o f n c e -m i R -11248d u r i n gi n f e c t i o n p r o c e s s o f N o s e m a c e r a n a e2.3 n c e -m i R -11248靶基因及其编码蛋白的分子特性根据n c e -m i R -11248的靶向预测结果筛选出1个关键靶基因 s k p1的g 2等位基因的抑制子基因S G T 1(G e n e I D :36319968),其编码蛋白S G T 1共含有441个核苷酸,编码146个氨基酸(M E F A E -T K T S V F L F L Y D N S I D T S R I S L N N P K S L F I T P Y K -S I D L Y K E V T H I Q K I K VH T K C V E I I L V K K V P S K -WY T L S G P E N I D E K K Q E Q K V I E E N D I T Q D D D I M -K V L S K I Y Q N G D E N T R R AM E K S F I E S D G T V L S -T NW E D V K N K K Y E Q )㊂S G T 1的分子式为C 765H 1213N 195O 241S 4,分子质量约为17.10k u ,脂溶系数为82.67,不稳定系数为35.84,等电点为5.50㊂S G T 1蛋白包含25个负电荷氨基酸和24个正电荷氨基酸,其中含量最高和最低的氨基酸分别是赖氨酸和半胱氨酸(表2)㊂S G T 1的平均亲水系数为-0.709,亲水氨基酸数量多于疏水氨基酸(图3-A ),说明该蛋白可能为亲水性蛋白㊂S G T 1包含9个丝氨酸磷酸化位点,3个酪氨酸磷酸化位点及8个苏氨酸磷酸化位点(图3-B )㊂表2 S G T 1蛋白的氨基酸组成T a b l e 2 A m i n o a c i d c o m po s i t i o n o f S G T 1p r o t e i n 氨基酸A m i n o a c i d数量N u m b e r频率/%F r e q u e n c y氨基酸A m i n o a c i d数量N u m b e r频率/%F r e q u e n c y丙氨酸A l a 21.40亮氨酸L e u 96.20精氨酸A r g 32.10赖氨酸L y s 1913.00天冬酰胺A s n 96.20蛋氨酸M e t 32.10天冬氨酸A s p 117.50苯丙氨酸P h e 53.40半胱氨酸C y s 10.70脯氨酸P r o 42.70谷氨酰胺G l n 64.10丝氨酸S e r 128.20谷氨酸G l u 149.60苏氨酸T h r 117.50甘氨酸G l y 32.10色氨酸T r p 21.40组氨酸H i s 21.40酪氨酸T y r 64.10异亮氨酸I l e149.60缬氨酸V a l106.80图3 S G T 1的亲水性(A )和磷酸化位点(B )F i g .3 H y d r o p h i l i c (A )a n d p h o s p h o r yl a t i o n s i t e (B )o f S G T 1 S G T 1不含典型的信号肽和跨膜结构域,说明该蛋白可能为胞内蛋白㊂S G T 1含有55个α-螺旋(占比37.67%),28个延长链(占比19.18%),58个无规则卷曲(占比39.73%),不含β-转角㊂进一步63西北农林科技大学学报(自然科学版)第52卷的三级结构分析结果显示,S G T 1与模板1r l 1.1.A 之间的同源性为20.00%㊂S G T 1可同时定位于细胞核㊁线粒体和过氧化物酶体,占比分别为73.90%,21.70%和4.30%㊂2.4 S G T 1蛋白的保守基序和结构域搜索结果显示,东方蜜蜂微孢子虫含有1个S G T 1,家蚕微孢子虫㊁肠脑炎微孢子虫㊁蚱蜢脑炎微孢子虫和蒺藜苜蓿均含有1个S G T 1,而泛胞虫含有2个S G T 1㊂保守基序分析结果(图4)显示,东方蜜蜂微孢子虫㊁家蚕微孢子虫㊁泛胞虫㊁肠脑炎微孢子虫㊁蚱蜢脑炎微孢子虫的S G T 1均含有4个保守基序,分别为M o t i f 1㊁M o t i f 2㊁M o t i f 3和M o t i f 4;而蒺藜苜蓿S G T 1仅包含M o t i f 1和M o t i f 22个保守基序㊂结构域分析结果(图5)显示,东方蜜蜂微孢子虫和泛胞虫的S G T 1包含S G T 1超家族和S G S 超家族2个结构域,而家蚕微孢子虫㊁肠脑炎微孢子虫和蚱蜢脑炎微孢子虫的S G T 1仅包含1个S G T 1超家族,蒺藜苜蓿包含1个P L N 03088结构域㊂上述结果表明,微孢子虫的S G T 1具有高度保守性㊂图4 东方蜜蜂微孢子虫和其他6个物种的S G T 1蛋白的保守基序F i g .4 C o n s e r v e d m o t i f s o f SG T 1p r o t e i n s i n N o s e m a c e r a n a e a n d o t h e r s i x s pe c i e s 图5 东方蜜蜂微孢子虫和其他6个物种的S G T 1蛋白的结构域F i g .5 S t r u c t u r a l d o m a i n s o f SG T 1p r o t e i n s i n N o s e m a c e r a n a e a n d o t h e r s i x s pe c i e s 2.5 S G T 1蛋白的系统进化分析图6显示,东方蜜蜂微孢子虫㊁家蚕微孢子虫㊁肠脑炎微孢子虫和蚱蜢脑炎微孢子虫聚为一支,2个泛胞虫聚为一支,而蒺藜苜蓿单独为一支;东方蜜蜂微孢子虫与家蚕微孢子虫的进化距离最近,与肠脑炎微孢子虫和蚱蜢脑炎微孢子虫的进化距离次之㊂图6 基于S G T 1的东方蜜蜂微孢子虫与其他6个物种的系统进化树F i g .6 P h y l o g e n e t i c t r e e o f N o s e m a c e r a n a e a n d o t h e r s i x s pe c i e s b a s e d o n S G T 1p r o t e i n s 2.6 东方蜜蜂微孢子虫侵染过程中S G T 1的表达谱图7结果显示,相较于侵染后1d ,侵染后2dS G T 1的表达量下调为侵染后1d 时的90.92%,但二者差异不显著(P >0.05);侵染4,6和8d S G T 173第2期张凯遥,等:东方蜜蜂微孢子虫n c e -m i R -11248及其靶基因S G T 1的生物信息学和表达谱研究的表达量均显著下调(P <0.05),分别下调为侵染后1d 时的30.46%,14.88%和17.61%㊂图7 东方蜜蜂微孢子虫侵染过程中S G T 1的表达谱F i g .7 E x p r e s s i o n p a t t e r n o f S G T 1d u r i n gi n f e c t i o n pr o c e s s o f N o s e m a c e r a n a e 3 讨 论本研究中,S t e m -l o o p R T -P C R 和S a n ge r 测序结果证实n c e -m i R -11248在东方蜜蜂微孢子虫孢子中是真实存在的,且其在微孢子虫孢子中可能发挥一定的作用㊂东方蜜蜂微孢子虫在西方蜜蜂工蜂中肠内的增殖周期约为4d [22]㊂本研究中n c e -m i R -11248表达谱测定结果表明,在东方蜜蜂微孢子虫第一个增殖周期刚启动时其表达水平即下调,且随着东方蜜蜂微孢子虫侵染时间的延长,其表达水平持续下调,这提示n c e -m i R -11248可能参与了调节东方蜜蜂微孢子虫的侵染过程㊂但本课题组前期研究发现,在东方蜜蜂微孢子虫侵染1~4d 的意蜂工蜂中肠内,n c e -m i R -12220表现出上升-下降-上升的表达趋势[16],n c e -m i R -34537[23]和n c e -m i R -23928[15]呈现上升-下降的表达趋势㊂上述结果表明,不同m i R N A 在东方蜜蜂微孢子虫侵染意蜂工蜂过程中的动态表达趋势不同,可能发挥的调控作用也不同㊂下一步研究拟通过饲喂模拟物和抑制物对意蜂工蜂中肠内n c e -m i R -11248进行过表达和敲减,进而明确其对东方蜜蜂微孢子虫增殖的影响㊂本研究对东方蜜蜂微孢子虫的n c e -m i R -11248靶蛋白S G T 1进行了分析,结果显示其分子式为C 765H 1213N 195O 241S 4,分子质量约为17.10k u ,脂溶系数为82.67,不稳定系数为35.84,等电点为5.50,是一种亲水性的胞内蛋白㊂本研究在东方蜜蜂微孢子虫㊁家蚕微孢子虫㊁泛胞虫㊁肠脑炎微孢子虫和蚱蜢脑炎微孢子虫S G T 1中均预测到4个相同的保守基序及1个相同的结构域(S G T 1超家族结构域),表明S G T 1在上述5种微孢子虫中较为保守㊂系统进化结果显示,东方蜜蜂微孢子虫与其他3种微孢子虫的S G T 1聚为一支,这与保守结构域和保守基序分析结果一致,表明S G T 1蛋白在上述微孢子虫中具有高度保守性㊂以上结果丰富了东方蜜蜂微孢子虫S G T 1的基本信息,为其进一步的功能研究提供了理论依据㊂S G T 1由S G T 1特异性基序S G S ㊁四肽重复结构域T P R ㊁可变区域V R 1和V R 2及C S 基序等5个结构域组成,通过与泛素连接酶复合物相互作用在伴侣蛋白H S P 90介导的蛋白稳定性和靶向蛋白降解方面起着重要作用[24]㊂动粒是细胞分裂过程中高保真染色体传递的重要组成部分,S G T 1的二聚化对动粒组装具有重要意义[25]㊂研究表明,S G T 1基因突变可导致酿酒酵母(S a c c h a r o m yc e s c e r e v i s i a e )细胞在G 1期或G 2期出现生长停滞[26]㊂虽然m i R N A 的经典调控方式为负调控基因表达,但近年来的研究表明哺乳动物和病毒m i R N A 也能通过非经典的方式正调控基因表达[27-28]㊂本研究发现,相较于侵染后1d ,侵染后2,4,6和8d S G T 1的表达量均明显下调,与n c e -m i R -11248的表达趋势一致,暗示二者之间存在潜在的相关关系,但这还需要进一步通过试验来证明㊂据本研究结果可推测,在东方蜜蜂微孢子虫感染意蜂工蜂过程中,n c e -m i R -11248的表达受到抑制,其靶基因S G T 1表达水平下调,进而调节孢子的增殖和侵染过程㊂目前,有关建立东方蜜蜂微孢子虫侵染西方蜜蜂工蜂的模型,并利用该模型开展东方蜜蜂微孢子虫基因的R N A i 研究已见诸报道[29]㊂后续将通过R N A i 对意蜂工蜂中肠内S G T 1进行干扰,进而揭示S G T 1在东方蜜蜂微孢子虫侵染过程中的功能㊂[参考文献][1] 曾志将.养蜂学[M ].北京:中国农业出版社,2017.Z e n g Z J .A p i c u l t u r e [M ].B e i j i n g :C h i n a A gr i c u l t u r e P r e s s ,2017.[2] P a r i s L ,E l A l a o u i H ,D e l b a c F ,e t a l .E f f e c t s o f t h e g u t pa r a s i t e N o s e m a c e r a n a e o n h o n e yb e e p h y s i o l o g y an d b e h a v i o r [J ].C u r r e n t O pi n i o n i n I n s e c t S c i e n c e ,2018,26:149-154.[3] 梁 勤,陈大福.蜜蜂保护学[M ].北京:中国农业出版社,2009:5-7.L i a n g Q ,C h e n D F .H o n e y b e e p r o t e c t i o n [M ].B e i j i n g:C h i n a A gr i c u l t u r e P r e s s ,2009:5-7.[4] B a r t e l D P .M i c r o R N A s :t a r g e t r e c o g n i t i o n a n d r e g u l a t o r y fu n -c t i o n s [J ].C e l l ,2009,136:215-233.[5] B a i X ,W a n g Q ,R u i X ,e t a l .U p r e gu l a t i o n o f m i R -1269c o n t r i -b u t e s t o t h e p r o g r e s s i o n o f e s o p h a g e a l s qu a m o u s c e l l c a n c e r 83西北农林科技大学学报(自然科学版)第52卷c e l l s a nd i s a s s o c i a te d w i t h p o o r p r o g n o s i s[J].T e c h n o l o g y i nC a n c e r R e s e a r c h&T r e a t m e n t,2021,20:1533033820985858.[6] Y a n g C,L i D,M a o D,e t a l.O v e r e x p r e s s i o n o f m i c r o R N A319i m p a c t s l e a f m o r p h o g e n e s i s a n d l e a d s t o e n h a n c e d c o l d t o l e-r a n c e i n r i c e(O r y z a s a t i v a L.)[J].P l a n t,C e l l&E n v i r o n-m e n t,2013,36:2207-2218.[7] C a y g i l l E E,B r a n d A H.m i R-7B u f f e r s d i f f e r e n t i a t i o n i n t h ed e v e l o p i n g D r o s o p h i l a v i s u a l s y s t e m[J].C e l l R e p o r t s,2017,20:1255-1261.[8] L i u Z,L i n g L,X u J,e t a l.M i c r o R N A-14r e g u l a t e s l a r v a l d e v e-l o p m e n t t i m e i n B o m b y x m o r i[J].I n s e c t B i o c h e m i s t r y a n d M o l e c u l a r B i o l o g y,2018,93:57-65.[9] L e e H C,L i L,G u W,e t a l.D i v e r s e p a t h w a y s g e n e r a t e m i c r o-R N A-l i k e R N A s a n d d i c e r-i n d e p e n d e n t s m a l l i n t e r f e r i n g R N A si n f u n g i[J].M o l e c u l a r C e l l,2010,38:803-814.[10]J i a n g N,Y a n g Y,J a n b o n G,e t a l.I d e n t i f i c a t i o n a n d f u n c t i o n a ld e m o n s t r a t i o n o f m i R N A s i n t h e f u n g u s C r y p t o c o c c u s n e o f o r-m a n s[J].P L o S O N E,2012,7:e52734.[11] Z h a n g M W,Z h u Z H,X i a Z K,e t a l.C o m p r e h e n s i v e c i r-c R N A-m i c r o R N A-m R N A n e t w o r k a n a l y s i s r e v e a l ed t he n o v e lr e g u l a t o r y m e c h a n i s m o f T r i c h o s p o r o n a s a h i i i n f e c t i o n[J].M i l i t a r y M e d i c a l R e s e a r c h,2021,8:19.[12] H u a n g Q,E v a n s J D.I d e n t i f i c a t i o n o f m i c r o R N A-l i k e s m a l l R N A sf r o m f u ng a l p a r a s i t e N o s e m a c e r a n a e[J].J o u r n a l o f I n v e r t e-b r a t e P a t h o l o g y,2016,133:107-109.[13]S h a o S S,Y a n W Y,H u a n g Q.I d e n t i f i c a t i o n o f n o v e l m i R N A sf r o m t h e m i c r o s p o r i d i a n p a r a s i t e N o s e m a c e r a n a e[J].I n f e c-t i o n,G e n e t i c s a n d E v o l u t i o n:J o u r n a l o f M o l e c u l a r E p i d e m i o l-o g y a n d E v o l u t i o n a r y G e n e t i c s i n I n f e c t i o u s D i s e a s e s,2021, 93:104930.[14]张文德,赵浩东,孙明会,等.东方蜜蜂微孢子虫孢子中微小R N A的鉴定与分析[J].昆虫学报,2022,65(6):708-717.Z h a n g W D,Z h a o H D,S u n M H,e t a l.I d e n t i f i c a t i o n a n da n a l y s i s o f m i c r o R N A s i n N o s e m a c e r a n a e s p o r e s[J].A c t aE n t o m o l o g i c a S i n i c a,2022,65(6):708-717.[15]张文德,胡颖,张凯遥,等.东方蜜蜂微孢子虫侵染意大利蜜蜂工蜂过程的n c e-m i R-23928及其靶基因表达谱[J].微生物学通报:2023,50(1):185-193.Z h a n g W D,H u Y,Z h a n g K Y,e t a l.E x p r e s s i o n p r o f i l e s o fn c e-m i R-23928a n d i t s t a r g e t g e n e s i n t h e i n f e c t i o n p r o c e s s o fA p i s m e l l i f e r a l i g u s t i c a w o r k e r s w i t h N o s e m a c e r a n a e[J].M i c r o b i o l o g y C h i n a:2023,50(1):185-193.[16]吴鹰,叶亚萍,张佳欣,等.东方蜜蜂微孢子虫侵染意大利蜜蜂工蜂过程中n c e-m i R-12220及其靶基因的表达谱[J].菌物学报,2022,41(10):1546-1557.W u Y,Y e Y P,Z h a n g J X,e t a l.E x p r e s s i o n p r o f i l e s o f n c e-m i R-12220a n d i t s t a r g e t g e n e s d u r i n g t h e N o s e m a c e r a n a e i n-f e c t i o n p r o c e s s o f A p i s m e l l i f e r a l ig u s t i c a w o r k e r s[J].M y c-o s y s t e m a,2022,41(10):1546-1557.[17]陈华枝,范小雪,范元婵,等.东方蜜蜂微孢子虫基因的可变剪接及可变腺苷酸化解析[J].菌物学报,2021,40(1):161-173.C h e n H Z,F a n X X,F a n Y C,e t a l.A n a l y s i s o f a l t e r n a t i v es p l i c i n g a n d a l t e r n a t i v e p o l y a d e n y l a t i o n o f N o s e m a c e r a n a eg e n e s[J].M y c o s y s t e m a,2021,40(1):161-173.[18]范元婵,王杰,孙明会,等.东方蜜蜂微孢子虫蓖麻毒素B凝集素基因的分子克隆及生物信息学分析[J].中国农业大学学报,2022,27(3):138-145.F a n Y C,W a n g J,S u n M H,e t a l.M o l e c u l a r c l o n i n g a n db i o i n f o r m a t i o n a n a l y s i s o f N o s e m ac e r a n a e r i c i n B-l e c t i n g e n e[J].J o u r n a l o f C h i n a A g r i c u l t u r a l U n i v e r s i t y,2022,27(3): 138-145.[19] C h e n D F,C h e n H Z,D u Y,e t a l.G e n o m e-w i d e i d e n t i f i c a t i o no f l o n g n o n-c o d i n g R N A s a n d t h e i r r e g u l a t o r y n e t w o r k s i n-v o l v e d i n A p i s m e l l i f e r a l i g u s t i c a r e s p o n s e t o N o s e m a c e r a-n a e i n f e c t i o n[J].I n s e c t s,2019,10:245.[20] M a r tín-H e r nán d e z R,B a r t o l o méC,C h e j a n o v s k y N,e t a l.N o s e m ac e r a n a e i n A p i s m e l l i f e r a:a12y e a r s p o s tde t e c t i o n p e r s p e c-t i v e[J].E n v i r o n m e n t a l M i c r o b i o l o g y,2018,20:1302-1329.[21] H u a n g Q,W u Z H,L i W F,e t a l.G e n o m e a n d e v o l u t i o n a r ya n a l y s i s o f N o s e m a c e r a n a e:a m i c r o s p o r i d i a n p a r a s i t e o f h o n-e y b e e s[J].F r o n t i e r s i n M i c r o b i o l o g y,2021,12:645353.[22]吴志豪,曾志将,黄强.东方蜜蜂微孢子虫传染病学分析[J].微生物学报,2021,61(9):2628-2642.W u Z H,Z e n g Z J,H u a n g Q.T h e e p i d e m i o l o g i c a l a n a l y s i s o fN o s e m a c e r a n a e[J].A c t a M i c r o b i o l o g i c a S i n i c a,2021,61(9): 2628-2642.[23]王紫馨,张文德,张佳欣,等.东方蜜蜂微孢子虫侵染过程中n c e-m i R-34537和靶基因P I P5K I的表达谱[J].微生物学报,2023,63(2):731-744.W a n g Z X,Z h a n g W D,Z h a n g J X,e t a l.I n v e s t i g a t i o n a n d e x-p r e s s i o n p r o f i l e d e t e r m i n a t i o n o f n c e-m i R-34537a n d t a r g e tg e n e P I P5K I d u r i n g t h e i n f e c t i o n p r o c e s s o f N o s e m a c e r a n a e[J].A c t a M i c r o b i o l o g i c a S i n i c a,2023,63(2):731-744.[24] M e l d a u S,B a l d w i n I T,W u J.F o r s e c u r i t y a n d s t a b i l i t y:S G T1i n p l a n t d e f e n s e a n d d e v e l o p m e n t[J].P l a n t S i g n a l i n g&B e-h a v i o r,2011,6:1479-1482.[25] B a n s a l P K,N o u r s e A,A b d u l l e R,e t a l.S G T1d i m e r i z a t i o n i sr e q u i r e d f o r y e a s t k i n e t o c h o r e a s s e m b l y[J].T h e J o u r n a l o fB i o l o g i c a lC h e m i s t r y,2009,284:3586-3592.[26] K i t a g a w a K,S k o w y r a D,E l l e d g e S J,e t a l.S G T1e n c o d e s a ne s s e n t i a l c o m p o n e n t of t h e y e a s t k i n e t o c h o r e a s s e m b l y p a t h-w a y a n d a n o v e l s u b u n i t o f t h e S C F u b i q u i t i n l i g a s e c o m p l e x[J].M o l e c u l a r C e l l,1999,4:21-33.[27] S h i m a k a m i T,Y a m a n e D,J a n g r a R K,e t a l.S t a b i l i z a t i o n o fh e p a t i t i s C v i r u s R N A b y a n A g o2-m i R-122c o m p l e x[J].P r o-c e ed i n g s o f t he N a t i o n a l A c a d e m y of S c i e n c e s o f t h e U n i t e d S t a t e so f A m e r i c a,2012,109:941-946.[28]θr o m U A,N i e l s e n F C,L u n d A H.M i c r o R N A-10a b i n d s t h e5'U T R o f r i b o s o m a l p r o t e i n m R N A s a n d e n h a n c e s t h e i r t r a n s l a t i o n[J].M o l e c u l a r C e l l,2008,30:460-471. [29] H e N,Z h a n g Y,D u a n X L,e t a l.R N A i n t e r f e r e n c e-m e d i a t e dk n o c k d o w n o f g e n e s e n c o d i n g s p o r e w a l l p r o t e i n s c o n f e r s p r o-t e c t i o n a g a i n s t N o s e m a c e r a n a e i n f e c t i o n i n t h e E u r o p e a nh o n e y b e e,A p i s m e l l i f e r a[J].M i c r o o r g a n i s m s,2021,9:505.93第2期张凯遥,等:东方蜜蜂微孢子虫n c e-m i R-11248及其靶基因S G T1的生物信息学和表达谱研究。
福建农林大学2019年硕士研究生招生专业目录

福
注:2019年我校共有90多个专业招收攻读全日制和非全日制硕士学位研究生1900名左右(含推荐免试、科、专业均面向全国招生。
004林学院
095400林业
090300农业资源与环境
福建农林大学二〇一九年硕士研究生招生专业目录
制硕士学位研究生1900名左右(含推荐免试、“退役大学生士兵”专项计划、和“少数民族高层次骨干人才计划”,具体
目录
“少数民族高层次骨干人才计划”,具体指标以教育部下达的招生计划为准),各学
137
森林培育学
3农业资源利用
运输学或物流系统规划与设计。
中国工程院院士黄璐琦来宁夏农科院调研

宁夏农林科技,Ningxia Journal of Agri.and Fores.Sci.&Tech.9月23日,中国工程院院士、国家中医药管理局副局长、中国中医科学院院长黄璐琦来宁夏农科院调研。
院党组副书记、院长刘常青,自治区卫健委纪检监察组组长赵捷等陪同调研。
在宁夏农林科学院芦花台现代农业科研基地,黄璐琦调研了生物中心承担的中央本级重大增减支项目“名贵中药资源可持续利用能力建设项目”课题“药用苦枸杞品种选育研究”的进展情况,问询了苦枸杞种质资源收集引种、遗传关系分子鉴定、扦插及组培快繁育苗、引种后表型变化分析等研究工作进展。
在听取了课题组成员汇报后,黄璐琦对课题研究进展表示了肯定,并对下一步研究工作给予了指导。
该课题针对当前枸杞品种缺失传统味“苦”药用性状的问题,与黄璐琦团队联合攻关,开展苦枸杞引种驯化、快速繁育、配套栽培、“味苦”关联组分解析、药效综合评价等研究工作,旨在建立苦枸杞“资源引选-种苗繁育-配套栽培-质量评价”技术体系,助力枸杞产业高质量发展。
此次黄璐琦调研指导,开拓了生物中心科技人员的研究思路,明确了今后重点攻关方向,为课题高水平实施奠定了良好的基础。
责任编辑:周慧中国工程院院士黄璐琦来我院调研文/院生物中心中国中医科学院和福建农林大学专家来宁夏农科院开展学术交流文/院生物中心9月11日,中国中医科学院中药资源中心副主任、国家杰出青年科学基金获得者袁媛研究员、福建农林大学张重义教授应邀到宁夏农科院开展学术交流,分别作了题为“优质药材品质形成分子机理”和“克服中药材连作障碍研究的思考”的学术报告。
生物中心全体科技人员,以及院枸杞科学所、林草所相关科技人员参加了报告会。
会上,袁媛研究员从药材基因型、表观遗传修饰等方面阐述了影响优质药材品质形成的分子机理;张重义教授以中药材地黄连作障碍为例,从根系分泌物、根际微生物组、植物免疫响应等方面探讨了中药材连作障碍形成机制以及解决连作障碍的技术方案。
2012春福建福州党群系统考录公务员和机关(单位)工作人员拟录用人员名单公示

现将福州市党群系统2012年春季考录公务员和机关(单位)⼯作⼈员拟录⽤⼈员名单予以公⽰。
公⽰时间为2012年7⽉17⽇⾄2012年7⽉23⽇。
公⽰期间,如有问题请及时向福州市委组织部党群机关⼲部管理处反映。
监督电话:0591-********。
中共福州市委组织部党群机关⼲部管理处2012年7⽉17⽇附:公⽰内容曾令锋,男,准考证号码:110140*********,拟录⽤单位:福州市纪律检查委员会、监察局,毕业院校及专业:三明学院汉语⾔⽂学专业,本科学历,⽂学学⼠学位;项振斌,男,准考证号码:110140102011057,拟录⽤单位:福州市纪律检查委员会、监察局,毕业院校及专业:厦门⼤学法学院法学专业,本科学历,法学学⼠学位;俞圣洁,⼥,准考证号码:110140103013063,拟录⽤单位:福州市纪律检查委员会、监察局,毕业院校及专业:福州⼤学法学院法学专业,本科学历,法学学⼠学位;陈⾃如,⼥,准考证号码:110143*********,拟录⽤单位:福州市效能办,毕业院校及专业:孝感学院汉语⾔⽂学专业,本科学历,⽂学学⼠学位;官⾦晶,⼥,准考证号码:110140104013629,拟录⽤单位:福州市纪律检查委员会、监察局,毕业院校及专业:福州⼤学经济学专业,本科学历,经济学学⼠学位;叶媛华,⼥,准考证号码:120140105010493,拟录⽤单位:福州市纪律检查委员会、监察局,毕业院校及专业:福建师⼤⽂学院汉语⾔⽂学专业,本科学历,⽂学学⼠学位;潘玮鸿,⼥,准考证号码:410140106011480,拟录⽤单位:福州市纪律检查委员会、监察局,毕业院校及专业:仰恩⼤学经济学专业,本科学历,经济学学⼠学位;江岚,男,准考证号码:310140*********,拟录⽤单位:中共福州市委宣传部,毕业院校及专业:南昌航空⼤学经济学专业,本科学历,经济学学⼠学位;潘志聪,男,准考证号码:310140402015417,拟录⽤单位:中共福州市委宣传部,毕业院校及专业:福州⼤学物流管理专业,本科学历,管理学学⼠学位;周颖,⼥,准考证号码:110140*********,拟录⽤单位:中共福州市委机构编制委员会办公室,毕业院校及专业:天津理⼯⼤学电⼦科学与技术专业,本科学历,⼯学学⼠学位;杨⼦健,男,准考证号码:122147001080564,拟录⽤单位:福州市红⼗字会,毕业院校及专业:解放军后勤⼯程学院应⽤化学专业,本科学历,⼯学学⼠学位;程君,⼥,准考证号码:122146001090171,拟录⽤单位:福州市⽼⼲部活动中⼼,毕业院校及专业:福州⼤学会计学专业,硕⼠研究⽣学历,管理学硕⼠学位;吴忠晗,男,准考证号码:112146002098091,拟录⽤单位:福州市⽼⼲部活动中⼼,毕业院校及专业:福建师范⼤学计算机科学与技术专业,本科学历,理学学⼠学位;王⽂娟,⼥,准考证号码:112149801095175,拟录⽤单位:福州市群众路⼲休所,毕业院校及专业:华中师范⼤学⽂学院汉语⾔⽂学专业,本科学历,⽂学学⼠学位;许菁,⼥,准考证号码:112142001083023,拟录⽤单位:福州市妇⼥联合会,毕业院校及专业:⼭东⼤学新闻学专业,本科学历,⽂学学⼠学位;张宝⽂,男,准考证号码:810019*********,拟录⽤单位:⿎楼区洪⼭镇。
福建农林大学2011-2012校内勤工岗位设置汇总表

第 2 页,共 11 页
福建农林大学2011-2012学年校内机关各部门勤工助学岗位需求汇总表(总需求人数:566人 贫困生优先) 用工 报名联系电话/ 序号 用工部门 设岗类型 岗位名称 岗位职责与要求 工作时段 报名地点 联系人 备注 人数 邮件 农学专业知识,动手能力 甘蔗综合研 临时岗位 数据整理员 录入试验数据 临时 2 徐良年 83759396 强,对甘蔗栽培、品种选育 究所 或组培感兴趣,能吃苦 农学专业知识,动手能力 甘蔗综合研 临时岗位 组培员 协助组培室组培工作 临时 2 徐良年 83759396 强,对甘蔗栽培、品种选育 究所 或组培感兴趣,能吃苦 染色体制片 甘蔗综合研 研究生,熟悉染色体原位杂 临时岗位 协助染色体制片工作 临时 1 徐良年 83759396 员 究所 交技术 有一定专利基础知识,一定 资料管理员、电脑录入、 的文字处理能力,能熟练使 知识产权管 7 固定岗位 学生助手 处务工作助理 课余时间 5 明德楼502 许海萍 83789285 用电脑,具有吃苦耐劳的 理处 精神。 专利方面信息采集,文字和数 学生助手招 固定岗位 据库处理 聘研究生 有较好的专利基础知识,有 较高的文字和数据库处理能 力,能熟练使用电脑,具有 吃苦耐劳的精神。 勤快认真,吃苦耐劳,责任 心强 无 无 无 无 要求普通话较标准、字迹工 整、熟悉电脑操作 要求家庭经济困难的研究 生,熟悉电脑操作、工作细 要求具备一定办公软件知 识,工作责任心强、细心耐 心的研究生 要求具备一定电脑办公软 件,责任心强,工作细致的
9 2 1 1 2 4 4 2
高丽榕 陈 寅 陈生农 陈生农 林和佳 林和佳 林和佳 林和佳
83789261 83789460 83719276 83719276 83776192 83776192 83776192 83776192
“股市黑嘴”与股价崩盘风险——基于中国证券监管行政处罚的实证研究

“股市黑嘴”与股价崩盘风险——基于中国证券监管行政处
罚的实证研究
李曦;陈振山;刘杰
【期刊名称】《管理现代化》
【年(卷),期】2024(44)1
【摘要】“股市黑嘴”通过在市场散播诱导性信息,影响投资者行为和股价表现,是典型的信息型市场操纵行为。
本文利用中国证监会2008年至2020年披露的148例股市黑嘴操纵案件,实证检验了股市黑嘴操纵对股价崩盘风险的影响,并考察其影响渠道。
结果表明股市黑嘴通过在市场上散播虚假信息,提高了市场信息不对称程度,加剧了操纵后的股价崩盘风险。
进一步的分析发现,对于股票流动性和信息透明度较好的公司,股市黑嘴操纵对崩盘风险的影响相对较弱。
【总页数】12页(P52-63)
【作者】李曦;陈振山;刘杰
【作者单位】福建省招标采购集团有限公司;福建农林大学经济与管理学院;福建普惠金融研究院
【正文语种】中文
【中图分类】F832.5
【相关文献】
1.公司债务期限结构与股价崩盘风险*--基于中国A股上市公司的实证证据
2.企业研发创新对股价崩盘风险的影响机制研究\r——来自中国A股市场的实证检验
3.
企业创新能力与股价崩盘风险:基于中国股市的分析4.期望绩效反馈会影响股价崩盘风险吗?——基于中国A股上市公司的实证分析5.中国环境信息规制的市场效应——基于股价崩盘风险的实证检验
因版权原因,仅展示原文概要,查看原文内容请购买。
核酸等温扩增技术在水产病原体快速检测中的应用

核酸等温扩增技术在水产病原体快速检测中的应用庞建虎;乔龙亮;朱鹏;高威芳;章礼萍【摘要】有别于费力和费时的传统病原体检测方法,基于靶基因的分子生物学检测方法以其快速和灵敏的特点逐渐成为现代水产病原体检测的首选.在众多分子生物学检测方法中,尤以核酸等温扩增技术备受关注.本文对各种核酸等温扩增技术在水产病原体检测中的应用进行了综述,阐述了各种核酸等温扩增技术的原理,对比分析了各种核酸等温扩增技术的优劣,介绍了该技术在水产病原体检测中的应用现状,展望了该技术的发展趋势,旨在促进我国水产病原体现场快速检测技术的发展.【期刊名称】《福建农林大学学报(自然科学版)》【年(卷),期】2018(047)005【总页数】11页(P530-540)【关键词】核酸等温扩增;水产病原体;快速检测【作者】庞建虎;乔龙亮;朱鹏;高威芳;章礼萍【作者单位】宁波大学海洋学院,浙江宁波315211;宁波大学海洋学院,浙江宁波315211;宁波大学海洋学院,浙江宁波315211;宁波海洋研究院,浙江宁波315832;宁波海洋研究院,浙江宁波315832;宁波海洋研究院,浙江宁波315832【正文语种】中文【中图分类】Q789近年来,我国水产养殖业蓬勃发展,养殖规模和产生的经济效益都是空前的.目前,我国水产品产量占动物性食物的1/3,处在世界首位,人均占有量高于世界平均水平[1].但随着集约化养殖方式的大规模应用,产量提高的同时也导致了病害的频繁爆发.同时,由于我国水产疫苗研究起步较晚,只有较少种类的疫苗可供使用[2].鉴于此,我国水产病害的防治主要以预防为主,快速、准确和简单的检测方法对于及时找出病原体显得尤为重要.因为只有找出真正的病原,才能对症下药,在疾病发生时尽量将损失减小到最低.然而传统的检测方法(选择性培养基和生理生化试验检测),尽管也能检测出病原体,但费力、费时和低灵敏度等缺点使得其不适合于当前水产病原体的疫情防控.如致病性嗜水气单胞菌的国标检测方法是经过一系列的生理生化试验鉴定,其检测周期长,操作十分复杂[3].分子生物学检测方法在灵敏度、特异性和检测时间等方面都优于传统病原体检测方法.核酸扩增在分子生物学检测中是常用的手段,聚合酶链式反应(polymerase chain reaction,PCR)是目前使用最频繁的核酸扩增技术.然而对精密温控仪器的依赖限制了其在条件较差的实验室及基层渔场的使用,尤其不适合现场快速检测.而核酸等温扩增技术不需要精密的温控循环设备,在一个特定的温度下通过添加不同性质的酶和特异性引物即可快速完成核酸的扩增,极大地降低了对设备的要求,缩短了反应时间,近年来正蓬勃发展.笔者通过对近十年来相关文献的整理发现:无论是从核酸等温扩增技术的开发上,还是其在病原体的检测应用上,大多数都是由国外研究者开展的,而国内研究者在这方面涉足较少.本文对目前常见核酸等温扩增技术的原理和优缺点等方面做了比较,并对它们在水产病原体的检测应用上做了系统的综述,希望能够帮助国内读者快速了解核酸等温扩增技术,促进该技术在水产病原体快速检测领域中的应用.1 常见的核酸等温扩增技术及比较目前常见的等温扩增技术有环介导等温扩增(loop-mediated isothermal amplification, LAMP)、依赖解旋酶等温扩增(helicase-dependent isothermalamplification, HDA)、依赖核酸序列等温扩增(nucleic acid sequence-based amplification, NASBA)、链替代等温扩增(strand displacement amplification, SDA)、交叉引物等温扩增(crossing priming amplification, CPA)、滚环等温扩增(rolling circle amplification, RCA)和重组酶聚合酶扩增(recombinase polymerase amplification, RPA)等[4].1.1 LAMPLAMP技术于2000年由日本Eiken公司发明[5].其原理是:根据目的基因的6个区域设计4条不同的引物(内引物和外引物),通过在反应体系中添加具有链置换特性的Bst DNA聚合酶、反应模板和基质等,在一定温度(60~65 ℃)下不断地进行DNA合成,从而达到目的基因快速扩增的目的,可在15~60 min实现109~1 010倍的扩增.LAMP技术具有对设备要求低、操作简单、灵敏度高和特异性强等优点.因其有着高度的特异性,因此只需根据扩增反应产物的有无即可对靶基因序列的存在与否作出判断.LAMP的缺点也是比较明显,如需要设计多对引物,使得引物设计较为复杂;其次是在对常规LAMP扩增产物的检测时,需要打开盖子,容易形成气溶胶,造成假阳性[6].1.2 HDAHDA是美国的研究者在2004年发明的一种等温扩增技术,其扩增原理类似于生物体内DNA的复制.主要原理如下:首先,通过解旋酶在恒等温的条件下使得DNA双链打开,随即单链DNA结合蛋白(SSB)结合到DNA单链上,抑制单链DNA重新形成双链DNA,在体系中提供了能够与引物互补结合的单链DNA模板,在DNA聚合酶催化下进行延伸(图1).刚刚合成的双链DNA随即又进入新的循环,结果使得目的基因呈指数式增长[7].图片来源于文献[6].A:解旋酶解开DNA双链;B:引物与单链DNA结合;C:DNA 聚合酶在引物基础上延伸,两条子链扩增形成;D:新的循环开始.图1 HDA 反应原理Fig.1 The principle of HDA reaction相较于传统的PCR扩增,HDA在于加入了解旋酶和单链结合蛋白,且整个扩增过程是在一个温度条件下进行的,避免了PCR扩增过程中通过温控仪来控制反应的进行.目前,研究者通过在体系中添加逆转录酶等,HDA也已经成功应用于RNA模板的扩增[8].HDA技术原理和对设备要求均简单,只需要恒温器即可,无需复杂引物.1.3 NASBANASBA技术是在PCR扩增技术的基础上发展起来的核酸扩增技术.NASBA是在等温条件下通过3种酶来控制反应,分别是来自禽流感病毒的逆转录酶(AMV)、来自大肠杆菌的核糖核酸酶(RNaseH)和来自噬菌体的T7RNA聚合酶[9].其原理是:整个扩增过程包括非循环相和循环相.在非循环相扩增过程中,反向引物在模板RNA上扫描与之互补的序列,随之反向引物与其特异性结合,在AMV的催化作用下合成互补的cDNA链,形成了RNA-DNA杂合体.随即RNAaseH特异地将RNA-DNA杂合体中的RNA链降解掉,带有启动子序列的正向引物与单链cDNA 结合,合成双链DNA.由于新合成的双链DNA带有可以被T7RNA聚合酶特异地识别的启动子序列,由此进入循环相扩增,高效、特异地合成反义RNA(图2).由于双链DNA中T7启动子序列的加入,而外来DNA不具有这样的结构,因此,NASBA不易受到污染,使得其特异性远远高于普通PCR扩增技术[10].图片来源于文献[11].A:反向引物在RNA模板扫描与之互补的序列;B:反向引物与模板特异性结合;C:RNA-DNA杂合体形成;D:RNA-DNA杂合体中的RNA链被降解;E:带有启动子序列的正向引物与单链DNA结合;F:互补链DNA延伸,带有T7启动子序列的DNA双链形成,以此为模板合成正向RNA;M、L、G、K、H、J、I:循环相步骤.图2 NASBA反应原理Fig.2 The principleof NASBA reaction1.4 SDAWalker et al[12]于1992年开发了SDA.其主要原理是:首先,通过引物与单链DNA进行特异地识别,在具有链置换活性的聚合酶催化下合成具有HincⅡ酶识别位点的目的DNA序列,由此进行SDA循环扩增.通过HincⅡ酶切割具有识别位点的目的DNA序列,通过聚合酶在切割处对3′端进行延伸扩增,互补链则被替换;新合成的替代链作为新的模板,体系中通过循环进行这些步骤,达到目的DNA序列快速扩增的目的.SDA虽然具有扩增效率高和特异强的优点,但也有缺点,如反应成本较高,且由于产物中存在单、双链DNA产物,因此电泳时拖尾现象较明显.1.5 CPACPA由杭州优思达公司完全独立研发,是目前我国首个具有完全自主知识产权的核酸等温扩增技术[13].该技术无需预变性步骤或者通过切口酶打开双链,在恒温63 ℃下引物与模板结合成混合物,反应产物中通过自发形成的泡状结构,交叉引物与5′端的结合可激发链置换反应的进行[14].CPA技术具有高灵敏度、高特异性和操作简便的优点.但该反应体系复杂、方法不稳定和假阳性现象等缺点也限制了该技术的使用[4].1.6 RCARCA技术于1998年建立的一种核酸等温扩增技术,扩增原理类似于CPA技术[15].目前主流的指数RCA的主要原理是:首先,正向引物结合到环状DNA序列上,在聚合酶作用下继续延伸,产生带有反向引物的结合位点的单链DNA,随即反向引物结合到单链DNA并进行延伸,产生的子代单链DNA充当模板,进行新的扩增(图3).扩增的结果是产生许多长短不一的双链DNA产物,扩增效率大大提高.虽然RCA技术具有高灵敏度和高效的扩增技术,但也存在着锁式探针连接效率、背景信号干扰和拓扑结构制约等问题[16].图片来源于文献[6].A:正向引物结合到环状DNA序列上并进行延伸;B:继续扩增,产生带有反向引物的结合位点的单链DNA;C:反向引物结合到单链DNA并进行延伸;D:产生的子代单链DNA充当模板,进行新的扩增.图3 指数RCA扩增原理Fig.3 The principle of hyperbranched RCA reaction1.7 RPARPA技术是被称为有望替代PCR技术的核酸扩增技术,是由英国TwistDx公司开发的核酸等温扩增产品.该技术对硬件设备的要求很低,一个简单水浴锅即可[17].它的原理是:在恒温(37~42 ℃)条件下,重组酶与引物结合形成聚合物,能够在模板链无需热变性的情况下与引物同源序列结合,通过链置换反应形成一个“D”字型环;在聚合酶催化下进行延伸,子链分离并继续延伸,对DNA序列进行指数式扩增(图4).RPA技术的关键和难点在于前期引物和探针的设计,大多数PCR引物都不适合,且目前没有专门的引物设计软件,需要研究者亲自摸索.其扩增产物可以通过琼脂糖凝胶电泳进行检测,操作步骤类似于普通PCR产物的检测;也可以通过将RPA 技术与横向流动试纸条(lateral flow dipstick, LFD)技术结合起来,通过试纸条检测反应产物;还可以在反应体系中加入探针,构建实时监测反应情况的实时荧光RPA.尽管核酸等温扩增技术一般只需要在恒定温度下就可以完成扩增,但各种核酸等温扩增所需要的温度不同,以及它们扩增需要的引物数量、酶的种类和扩增时间等方面各不相同,笔者通过对相关文献的阅读并结合本课题组的科研经历,在表1系统地列举了各种常见的核酸等温扩增技术所需引物数量、温度、时间及优缺点.图片来源于文献[18].A:重组酶与引物结合形成复合物,并与同源靶序列结合;B:通过链置换形成一个“D”字型环;C:聚合酶促使引物延伸;D:子链分离,并继续延伸;E:子链形成.图4 RPA反应原理Fig.4 The principle of RPA reaction 表1 常见核酸等温扩增技术的比较Table 1 Comparison of common isothermal nucleic acid amplification techniques技术引物/条温度/℃时间/min优点缺点LAMP460~6560特异性强,扩增效率高引物设计复杂HDA260~65120扩增效率高,引物设计简单需要复杂的反应体系优化过程NASBA237~4290~120检测速度快,有商业试剂盒可以使用不适用于DNA的扩增SDA437120效率高主要适用于短片段的扩增CPA46370操作简便反应体系复杂 RCA13760适合于单链DNA的扩增成本高RPA225~4220引物设计简单,特异性高对反应条件要求高2 在水产病原体快速检测上的应用现状通过对PubMed数据库中近十年来核酸等温扩增技术在水产病原体检测上应用文献的检索发现:各种核酸等温扩增技术在水产病原体检测上的应用频次差别较大;各种核酸等温扩增技术的检测限和检测时间也各不相同;在细菌、病毒和寄生虫等病原体的检测上均有应用.表2列举了各种常见核酸等温扩增技术近十年来的具体应用情况.2.1 在细菌快速检测上的应用LAMP技术广泛应用于细菌性水产病原体的检测上,在多种致病性弧菌和链球菌等的检测上都已成功应用(表2).王耀焕等[19]建立了创伤弧菌的LAMP快速检测方法,能在1 h内高效特异地检出创伤弧菌,通过将LAMP技术与LFD技术结合起来,建立了创伤弧菌的LFD-LAMP检测方法,能够快速、灵敏、特异地检测出创伤弧菌;Wang et al[20]建立了同时检测创伤弧菌和副溶血性弧菌的LAMP检测方法,其灵敏度是常规PCR的100倍,且可以在多种样品中同时检测出创伤弧菌和副溶血性弧菌;Suebsing et al[21]根据无乳链球菌和海豚链球菌的sodA基因设计引物,建立了罗非鱼链球菌病的LAMP检测方法,其灵敏度是巢穴PCR的10倍,结果可以直接通过观察颜色变化读取.表2 近十年等温扩增技术在水产病原体快速检测中的应用Table 2 The application of isothermal amplification in the rapid detection of aquatic pathogens in the past decade技术病原体检测限检测时间/min参考文献LAMP副溶血性弧菌和创伤弧菌250 fg和125 fg19[20]鳗弧菌0.40~6.42pg30[22]海豚链球菌和无乳链球菌123和114 CFU30[21]中华肝吸虫10-8ng35[23]日本血吸虫100 fg65[24]创伤弧菌3.7×102 CFU80[19]Laem-Singh virus100 fg60[25]第三类鲤科疱疹病毒10拷贝60[26]虾白斑综合征病毒24拷贝30[27]创伤弧菌1.5×102 CFU90[28]海豚链球菌100 fg60[29]中华鳖虹彩病毒20拷贝60[30]哈维氏弧菌17.2细胞60[31]溶藻弧菌3.7×102 CFU60[32]传染性脾肾坏死病毒10拷贝20[33]大菱鲆红体病虹彩病毒7拷贝60[34]虾白斑综合征病毒1 pg50[35]黑美人弧菌102 CFU60[36]虾白斑综合征病毒1×102拷贝60[37]鲁氏耶尔森氏菌1 pg60[38]鲤春病毒血症病毒10 TCID5060[39]创伤弧菌107 CFU60[40]虾白斑综合征病毒、传染性皮下及造血组织坏死病毒100 fg和10 pg60[41]虾白斑综合征病毒100拷贝60[42]弧菌103 CFU60[43]传染性皮下及造血组织坏死病毒100拷贝60[44]迟缓型爱德华氏菌10 CFU45[45]创伤弧菌100拷贝60[46]传染性皮下及造血组织坏死病毒6拷贝60[47]桃拉综合征病毒100 fg60[48]斑节对虾浓核病毒1 ng75[49]斑节对虾浓核病毒0.1 pg75[50]斑节对虾浓核病毒50拷贝60[51]溶藻弧菌1×102 CFU30[52]传染性脾肾坏死病毒1 fg45[53]隐性死亡病毒6.3 pg50[54]石斑鱼神经坏死病毒4.9×10-11拷贝60[55]蟾蜍米诺病毒5拷贝40[56]草鱼呼肠病毒7拷贝30[57]坏死病病毒7.5TCID5030[58]海豚链球菌12.4细胞60[59]HDA鳗弧菌30 ng130[60]副溶血性弧菌19.9 ng130[61]NASBA溶藻弧菌6.9×102 CFU90[62]副溶血性弧菌5.1×102 CFU90[63]草鱼呼肠病毒14拷贝300[64]CPA牡蛎疱疹病毒30拷贝60[65]神经坏死病毒101拷贝65[66]虾白斑综合征病毒101拷贝60[67]RPA传染性皮下及造血组织坏死病毒4拷贝7[68]虾白斑综合征病毒5拷贝6.5[69]Vibrio owensii2拷贝9[70]鲑鱼立克次氏体3×103拷贝60[71]第三类鲤科疱疹病毒10拷贝20[72]HDA技术目前仅在霍乱弧菌、副溶血性弧菌和鳗弧菌等水产病原体的检测上有应用.Tong et al[73]将HDA技术与TaqMan探针结合起来,建立了高效、快速的检测霍乱弧菌的方法.石琰璟等[61]根据副溶血性弧菌的tlh基因保守区设计引物,构建了等温条件下快速、特异地检测副溶血性弧菌的HDA检测方法,且特异性良好,检测限达到19.9 ng,其灵敏度与普通PCR相当.梁君妮等[60]根据鳗弧菌的toxR基因设计特异引物,通过对反应条件和体系的优化,成功构建了基于HDA的鳗弧菌快速检测方法.该方法能够特异地检出鱼类鳗弧菌,不会与其他鱼类病原体产生交叉反应;其检测限达到30 ng,使用构建的方法对人工污染样品的检测限为2.1×102 CFU,通过HDA对120份实际样品进行检测,与对比PCR检测技术的符合率为100%,阳性检出率优于传统的细菌培养法.秦胜利等[62]根据溶藻弧菌的hsp60基因为目的序列设计引物,构建了溶藻弧菌的NASBA快速检测体系.结果表明:该方法能特异地检测出6.9×102 CFU溶藻弧菌DNA,灵敏度比普通PCR方法高.倪鑫等[63]根据副溶血性弧菌的tlh基因为靶基因设计引物和探针,建立的NASBA检测方法对副溶血性弧菌的检测限为5.1×102 CFU,且与其他病原菌不会产生交叉反应.相比Wang et al[20]建立的LAMP检测副溶血性弧菌方法和石琰璟等[61]建立的HDA 检测副溶血性弧菌方法,LAMP检测方法的检测限和检测时间明显优于其他两种方法.2.2 在病毒快速检测上的应用LAMP技术在水产病毒病原体的检测上应用广泛,尤其在虾类病毒的检测上.如李红梅等[74]研究构建了对虾传染性皮下及造血组织坏死病毒(infectioushypodermal and hematopoietic necrosis virus, IHHNV)的LAMP检测方法.结果表明:建立的LAMP检测方法在63 ℃条件下,能够在30 min内检测出106倍稀释的IHHNV基因组DNA,与27种水生动物及对虾常见病原的DNA均无交叉反应.CPA技术在水产病原体的检测上已有报道.赵小金等[65]依据牡蛎疱疹病毒魁蚶株(OsHV-1-SB)全基因组序列保守区设计引物,建立了CPA检测体系,并对反应体系和反应条件进行了优化.结果表明:该方法能够特异地检测出30拷贝的质粒DNA,使用建立的方法在对22份实际样品的检测中显示,该方法简单、迅速、特异性强.粟子丹[66]建立的赤点石斑鱼神经坏死病毒的CPA联合切向流试纸条检测方法(CPA-LFD)简单、快速,该方法能够特异地检测出10拷贝 RNA模板,比RT-PCR的检测限还要高出10倍,与常见的鱼类病毒性病原体均不会出现交叉反应.杨昊霖[67]根据对虾白斑综合征病毒(white spot syndrome virus, WSSV)的基因组保守区设计引物构建了CPA检测方法,并将该方法与WSSV的其他核酸扩增检测方法进行了比较.结果显示,构建的CPA检测方法对WSSV的检测限为10拷贝,灵敏度高于普通PCR检测方法,与qPCR和LAMP方法有着相同的灵敏度,与其他常见的虾类病毒IHHNV、HPV和副溶血性弧菌均没有交叉反应.2.3 在寄生虫快速检测上的应用核酸等温扩增技术在寄生虫的检测上亦有少量应用.Cai et al[23]利用LAMP技术检测鱼的肝脏吸虫——中华肝吸虫,根据组织蛋白酶B3基因序列设计引物,建立的LAMP检测技术的灵敏度是传统PCR技术的100倍.余传信等[75]和Kumagai et al[24]分别建立了日本血吸虫感染性钉螺的LAMP检测方法,能够在65 min内分别检测出100 fg和1 pg血吸虫DNA.余传信等[75]将建立的LAMP检测技术在30只感染性钉螺实际样品的检测中发现,LAMP的阳性检出率为93.33%,敏感性高于传统PCR的阳性检测率(83.33%).3 存在问题及展望3.1 在水产病原体快速检测上存在的问题通过对PubMed数据库近十年来的相关文献检索发现:众多核酸等温扩增技术在水产病原体快速检测上应用最多、最成熟的是LAMP技术(41篇),在细菌、病毒和寄生虫的检测上都有应用,而其他等温扩增技术在水产病原体的检测上应用较少或至今仍没有应用.而PCR这一经典核酸扩增技术尽管需要依赖于精密仪器且存在一些其他方面的不足,但毋庸置疑仍是目前体外核酸扩增应用最为成熟和广泛的技术.在国内外有多种依据PCR技术开发的商业试剂盒可供水产病原体的检测使用,而对比核酸等温扩增技术,尽管也有相关文献报道,但大多数还是处于实验室研究阶段,仅LAMP和NASBA检测方法在致病菌检测上有相关少量的试剂盒可供使用,并建立了相应的进出口检验检疫标准(SN/T 2754和GB/T 19439—2004);但其他大多数重要的水产病原体却未能建立核酸等温检测方法,可供使用的具有商业价值的水产病原体的试剂盒较少,核酸等温扩增技术的相关检测标准也不完善,这些都限制了等温扩增技术的使用和推广.3.2 展望进入21世纪,随着生物技术的迅速发展,催生了各种核酸扩增技术,但任何病原体检测方法的研发,都要以实际应用为最终目的.核酸等温扩增技术具有对设备要求简单、操作方便、在等温条件下即可迅速完成反应等优点,受到国内外研究者的重视.然而,核酸等温扩增技术要想真正做到适应于现场快速检测,笔者认为以下几个方面将是未来研发的重点.3.2.1 开发专门的引物设计软件目前,各种核酸等温扩增技术除了LAMP技术具有专门适用的引物设计软件外,其他技术均没有专用的引物设计软件供研究者使用,研究者往往是先根据PCR引物设计软件预评估设计引物的优劣,再进行实验筛选.根据本课题组的经验得知,PCR技术软件评出的结果与实际实验结果往往有很大的区别,这样就大大增加了引物设计的工作量和成本.因此根据各种核酸等温扩增技术开发出不同的引物设计软件,对于不同核酸等温扩增技术的推广是至关重要的一步,它的完善必将推动核酸等温扩增技术的跨越式发展.3.2.2 研发重要水产病原体的商业试剂盒目前,核酸等温扩增技术虽然在水产病原体的检测上有文献报道(表2),但大多数重要的水产病原体还未有专门的商业试剂盒可供非专业的基层养殖场从业人员使用.因此,基于各种核酸等温扩增技术的扩增效率高、特异性强和灵敏度高等特点,开发出针对各种重要水产病原体的快速诊断试剂盒对于水产疾病的防控有重要的意义.3.2.3 结合不同技术各种核酸等温扩增技术尽管有诸多优点,但还存在各自的缺点,研究者需要将不同技术结合起来使用[76-78],克服技术本身的缺点,使得结果读取更为方便和快捷.如将LFD技术与RPA技术结合起来,使得反应结果通过试纸条颜色变化即可读取,摆脱了对仪器的依赖,同时也不需要操作人有专业背景,特别适合我国基层养殖场使用.本世纪以来,迅速发展的水产养殖业满足了人类对蛋白质的需求.快速、简便和灵敏的核酸等温检测技术非常适用于现场快速检测及基层养殖场普及应用.核酸等温检测技术的不断完善、新的试剂盒出现以及与其他新技术的联合应用,将极大地推动水产养殖业的发展.参考文献【相关文献】[1] 刘英杰,刘永新,方辉,等.我国水产种质资源的研究现状与展望[J].水产学杂志,2015,28(5):48-55.[2] 黄辉,齐振雄,余露军,等.我国水产疫苗的研究现状[J].湖南农业科学,2010(21):136-138.[3] 中华人民共和国国家质量监督检验检疫总局.GB/T 18652—2002 致病性嗜水气单胞菌检验方法[S].北京:中国标准出版社,2002.[4] 朱智壕,葛丽雅,涂晓波,等.等温扩增技术在食源性致病菌检测中的应用进展[J].食品安全质量检测学报,2015,6(12):4 787-4 794.[5] NOTOMI T, OKAYAMA H, MASUBUCHI H, et al. Loop-mediated isothermal amplification of DNA [J]. Nucleic Acids Research, 2000,28(12):E63.[6] YAN L, ZHOU J, ZHENG Y, et al. Isothermal amplified detection of DNA and RNA [J]. Molecular Biosystems, 2014,10(5):63-1 003.[7] 贾辉,胡兰,马晓威.依赖解旋酶DNA等温扩增技术的研究进展[J].新农业,2015(9):9-10.[8] OKUDA M, KUBOTA K, ONUKI M. Development of RT-PCR assay using a primer cocktail for eight virus species infecting melon (Cucumis melo) and cucumber (Cucumis sativus) [J]. Kyushu Plant Protection Research, 2007,53:9-13.[9] 杨海鸥,叶星晨,傅启华.依赖核酸序列扩增技术检测呼吸道合胞病毒方法的建立[J].检验医学,2016,31(3):168-172.[10] FYKSE E M, NILSEN T, NIELSEN A D, et al. Real-time PCR and NASBA for rapid and sensitive detection of Vibrio cholerae in ballast water [J]. Marine Pollution Bulletin, 2012,64(2):200-206.[11] GILL P, GHAEMI A. Nucleic acid isothermal amplification technologies: a review [J]. Nucleosides Nucleotides & Nucleic Acids, 2008,27(3):224-243.[12] WALKER G T, FRAISER M S, SCHRAM J L, et al. Strand displacement amplification-an isothermal, in vitro DNA amplification technique [J]. Nucleic Acids Research, 1992,20(7):1 691-1 69 6.[13] XU G, HU L, ZHONG H, et al. Cross priming amplification: mechanism and optimization for isothermal DNA amplification [J]. Sci Rep, 2012,2(2):57-64.[14] ZHAO Y, ZHANG X, ZHANG H, et al. Rapid and sensitive detection of Enterobacter sakazakii by cross-priming amplification combined with immuno-blotting analysis [J]. Mol Cell Probe, 2010,24(6):396-400.[15] 祁军,张霞,蒋刚强,等.交叉引物等温扩增技术检测志贺氏菌[J].食品研究与开发,2013(2):65-67.[16] 孟兆祥,张伟,檀建新,等.一种DNA扩增的新技术:利用热稳定的Bst DNA聚合酶驱动跨越式滚环等温扩增反应[J].中国生物化学与分子生物学报,2013,29(9):892-898.[17] PIEPENBURG O, WILLIAMS C H, STEMPLE D L, et al. DNA detection using recombinationp roteins [J]. PLoS Biology, 2012,4(7):1 115-1 121.[18] 高威芳,朱鹏,黄海龙.重组酶聚合酶扩增技术:一种新的核酸扩增策略[J].中国生物化学与分子生物学报,2016,32(6):627-634.[19] 王耀焕,王瑞娜,周前进,等.环介导等温扩增联合横向流动试纸条快速检测创伤弧菌检测方法的建立[J].生物技术通报,2014,6(6):81-87.[20] WANG Y, LI D, WANG Y, et al. Rapid and sensitive detection of Vibrio parahaemolyticus and Vibrio vulnificus by multiple endonuclease restriction real-time。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
项目名称:菌草生态循环产业关键技术研究与应用
申报奖种:福建省科技进步奖
成果完成人:林占熺,林兴生,刘斌,林冬梅,林辉,梁学武,杨宝学,李晶,苏德伟,刘朋虎
完成单位:福建农林大学,北京大学
项目简介:
成果所属农业科学和生态农业领域。
主要针对山地生态脆弱、菌林矛盾突出、天然饲料短缺和农业废弃物资源化高效循环利用等问题,项目组集中开展了菌草生态循环产业关键技术研究与应用,历经30多年,获得授权发明专利27项,发表论文87篇,省地方标准3项,专著1部。
菌草技术被联合国列为中国-联合国和平与发展基金重点推进项目,向全球推广,为落实联合国2030年可持续发展目标贡献“中国菌草方案”;《关于加快推进黄河菌草生态屏障建设和菌草新兴产业发展的建议》被中国工程院采纳。
李玉、王光谦、唐守正和康绍忠等11位院士评价认为:菌草生态治理投入少、见效快、效果好,有利于生态安全和环境改善,为生态治理提供了新思路、新技术和新途径。
主要创新点如下:
1.开辟了新的科学研究领域——菌草学,创立了菌草生态治理及其综合循环利用技术体系,组建了国家菌草工程技术研究中心。
先后在福建、西藏、内蒙古、青海等地的生态脆弱区,开展菌草生态治理及其产业发展研究和示范,菌草具有侵蚀控制、保水固土、改善生境和培肥地力等功能。
2.创立了菌草种质资源库(圃),选育出巨菌草、绿洲3号等适宜生态治理及产业化利用的优质菌草草种,揭示了巨菌草高光效、高水分利用率、高产、优质、耐旱机理和绿洲系列菌草抗寒机制;首次从巨菌草中发现并分离出内生固氮菌;制定了《菌草草种认定技术规程》和《菌草栽培技术规范》。
3. “以草代木”栽培食药用菌,从根本上解决了“菌林矛盾”,实现菌业可持续发展;鲜菌草栽培食药用菌,解决了菌草干燥问题,节省了成本,提高了效益。
制定了《菌草栽培灵芝技术规范》。
4.首次得到纯度97%以上的GL-PPSQ2标样,建立了《高效凝胶过滤色谱法测定灵芝多糖肽》检测方法,为菌草灵芝产品质量的检控提供科学依据。
研发出银耳多糖、冻干银耳羹、竹荪豆腐等9个营养与功能食品,带动了菌业等上下游产业的规模化开发。
建立了菌草和菌糠营养成分与抗营养因子数据库,研究出菌草和菌糠饲料化工艺,明确了菌草菌物饲料畜禽消化代谢及对生长性能的影响和
饲喂牛、羊、欧洲鳗鲡的效果。
5.创新菌草生态循环产业开发与应用模式。
运用菌草技术,建立了“菌草-生态治理-综合循环利用”模式,发展菌草生态循环产业,经济、生态和社会效益显著。
成果已在我国31个省市自治区的496个县市推广应用,建立了63个示范基地,累计种植菌草18万亩,新增产值8.7亿元。
成果技术传播到103个国家,在巴布亚新几内亚、卢旺达、斐济等13国建立了示范基地,举办了17届国际菌草产业发展研讨会,举办了174期国际菌草技术培训班,培训国外人才6964人次。
巴布亚新几内亚、圭亚那、莱索托、南非、柬埔寨、斐济6国领导人及200余个外交使团访问菌草中心,在巴布亚新几内亚坚持“一个中国”原则中起到特殊的贡献,为服务我国“一带一路”战略的实施做出了积极贡献。
经同行专家评审,一致认为“整体研究达到国际领先水平”。
主要代表性论文列表:
1. Physiological and photosynthetic responses of Giant juncao (Pennisetum giganteum) to drought stress
2. Effect of JUNCAO-cultivated Ganoderma lucidum spent mushroom substrate-hot water extract on immune function in mice.
3. Extracts of Ganoderma lucidum attenuate lipid metabolism and modulate gut microbiota in high-fat diet fed rats
4. Optimization of polysaccharides extraction from Dictyophora indusiata and determination of its antioxidant activity.
5. Ganoderma lucidum polysaccharid peptide prevents renal inschemia reperfusion injury via conunteracting oxidative stress.
6. 菌草绿洲3号快繁及遗传转化体系的初步研究.
7. 巨菌草不同生长时期的内生固氮菌群组成分析.
8. 芦竹属新材料绿洲3号的愈伤组织诱导
9. 真姬菇富硒栽培及其成分的研究
10. 菌草栽培香菇
主要代表性专利:
1. 新鲜菌草温室瓶栽紫孢平菇的方法,ZL201266145.4
2. 新鲜菌草温室瓶栽杏鲍菇的方法,ZL201262998.0
3. 一种能源草的综合循环利用方法,ZL201
4.1.0041568.8
4. 菌草栽培灰树花的方法,ZL201210181193.6
5. 一种灰树花保健口服液及其制备方法,ZL2012.1.0334780.9
6. 一种快速测定灵芝产品中多糖肽的方法,ZL201410105731.2
7. 一种方便快速测定植物物料含氮率的方法,ZL2013.1.0331746.6
8. 一种真姬菇菌糟全混合日粮及其制作方法和应用,ZL2013.1.0111722.X
9. 一种种植菌草治理崩岗的方法,ZL201210339668.X
10. 一种“巨菌草-草菇-菌肥”相互促生和转化的方法,ZL201410750383.4。