第五章 细胞衰老与凋亡
细胞衰老和凋亡ppt-细胞生物学PPT课件

维持内部环境的稳定
抵御外界各种因素的干扰
2、细胞坏死是在种种不利因素影响下,由于细胞
正常代谢活动受损或中断引起的细胞损伤和死亡。
2020年10月2日
9
三、细胞的凋亡和细胞坏死
1、细胞凋亡是由基因所决定的细胞自动结束生命的过程
2020年10月2日
22
演讲完毕,谢谢观看!
Thank you for reading! In order to facilitate learning and use, the content of this document can be modified, adjusted and printed at will after downloading. Welcome to download!
生化特点 有无炎症
举例
细胞的凋亡 细胞的坏死
基因引起 与基因无关
温和的刺激或 剧烈的刺激或
药物
损伤
主动死亡
被动死亡
2020年10月2日
13
合作探究 2、细胞凋亡与细胞坏死的区别:
项目 内因 诱因
生化特点 有无炎症
举例
细胞的凋亡 细胞的坏死
基因引起 与基因无关
温和的刺激或 剧烈的刺激或
药物
损伤
主动死亡 无
诱(外)因:环境中的致癌因子的作用。
内因:原癌基因和抑癌基因发生基因突变,导 致正常的细胞生长和分裂失控而变成癌细胞。
原癌基因:负责调节细胞周期、控制细胞生 长和分裂。
抑癌基因:阻止细胞不正常的增殖。
2020年10月2日
21
细胞的衰老和凋亡高中生物教案范文

一、教学目标1. 让学生了解细胞衰老和凋亡的概念及意义。
2. 让学生掌握细胞衰老和凋亡的基本特征。
3. 让学生理解细胞衰老和凋亡在生物体生长发育中的作用。
4. 培养学生观察、分析问题的能力。
二、教学内容1. 细胞衰老和凋亡的概念2. 细胞衰老和凋亡的基本特征3. 细胞衰老和凋亡的意义4. 细胞衰老和凋亡在生物体生长发育中的作用三、教学重点与难点1. 教学重点:细胞衰老和凋亡的概念、基本特征及意义。
2. 教学难点:细胞衰老和凋亡的机制。
四、教学方法1. 采用问题驱动法,引导学生思考和探讨细胞衰老和凋亡的意义。
2. 使用多媒体教学,展示细胞衰老和凋亡的相关图像和实例。
3. 进行小组讨论,培养学生的合作能力和口头表达能力。
五、教学过程1. 引入:通过展示老年人皮肤和器官的图片,引导学生思考细胞衰老的现象。
2. 讲解:介绍细胞衰老和凋亡的概念、基本特征及意义。
3. 案例分析:分析细胞衰老和凋亡在生物体生长发育中的作用,如蝌蚪尾的消失、红细胞的寿命等。
4. 小组讨论:让学生结合实例,讨论细胞衰老和凋亡的作用和机制。
5. 总结:概括本节课的主要内容,强调细胞衰老和凋亡的重要性。
6. 作业布置:布置相关思考题,让学生进一步巩固所学知识。
六、教学策略1. 采用问题驱动法,引导学生思考和探讨细胞衰老和凋亡的意义。
2. 使用多媒体教学,展示细胞衰老和凋亡的相关图像和实例。
3. 进行小组讨论,培养学生的合作能力和口头表达能力。
4. 设计实验,让学生亲身体验细胞衰老和凋亡的过程。
5. 结合生活实例,让学生认识到细胞衰老和凋亡在生物体生长发育中的作用。
七、教学准备1. 准备相关教学课件和图片。
2. 准备实验材料和仪器。
3. 收集生活实例,用于教学。
八、教学评价1. 课堂问答:检查学生对细胞衰老和凋亡概念、特征的理解。
2. 小组讨论:评估学生在讨论中的合作能力和口头表达能力。
3. 实验报告:评价学生对实验原理和结果的理解。
高中生物《细胞的衰老与凋亡》 答辩题目及解析

高中生物《细胞的衰老与凋亡》答辩题目及解析1.请问什么是细胞凋亡?它的重要意义是什么?【参考答案】概念:细胞凋亡又称编程性细胞死亡,是在生物个体生长发育过程中由基因控制的,按照一定程序发生的细胞死亡。
意义:维持内部环境的稳定;抵御外界各种因素的干扰;多细胞生物体完成正常发育的必要过程。
2.细胞凋亡会不会引起炎症呢?【参考答案】细胞通过出芽的方式形成许多凋亡小体,凋亡小体内有结构完整的细胞器,还有凝缩的染色体,可被邻近细胞吞噬消化,因始终有膜封闭,没有内容物释放,故不会引起炎症。
细胞坏死会引起炎症。
3.细胞凋亡与细胞坏死有什么区别?【参考答案】区别细胞凋亡细胞坏死起因生理性病理变化或剧烈损伤范围单个散在细胞大片组织或成群细胞细胞体积固缩变小肿胀变大凋亡小体细胞膜完整,形成凋亡小体无,细胞自溶,细胞膜破坏蛋白质合成有无调节过程受基因调控被动进行炎症反应无,不释放细胞内容物有,释放内容物。
4. 有一天,你上课的时候,学生向你提出里一个你也不知道的怎么解答的问题,你准备怎么办?【参考答案】首先,我会赞扬学生求知好学的精神。
其次,我将以诚恳的态度向学生说明由于该问题不是我所了解的领域,因此我不能立马解答,但我会在课后查找了解后及时回复他;之后继续开展授课,保证课程正常进行。
然后,在课后,我会查阅相关资料、请教同事,通过网络查找该问题答案,并将答案进行归纳理解,以便更好地传授学生。
最后,在今后的教学工作中,我会鼓励学生积极提问,充分发挥学生好学求知能力;同时我也会利用业余时间多学习多准备,以便能更好地做好教学工作。
5.本节课中,你的教学重点是什么?如何进行突破呢?【参考答案】本节课中的教学重点是个体衰老与细胞衰老的关系。
许多学生根据已有的生活经验,会认为细胞衰老就会导致个体衰老。
因此纠正学生的认知误区,正确理解个体衰老与细胞衰老的关系尤为重要。
同时为后面学习衰老细胞的特征作铺垫,故该知识点为重点。
在教学的过程中,我将主要通过层层的问题引导,由生活实际入手,学生小组交流与教师补充的方式来总结出:对于单细胞生物,细胞衰老等于个体衰老;对于多细胞生物,二者之间不等同。
细胞的衰老和凋亡(含解析)

课时作业(二十)[学业水平层次(A)]1.关于细胞凋亡过程的叙述,错误的是()A.凋亡开始时,细胞皱缩,细胞膜破裂B.凋亡小体包含染色体片段和细胞器C.凋亡小体是细胞膜内折形成的泡状和芽状突起D.凋亡细胞最终被周围细胞吞噬消化【解析】细胞凋亡分为三个阶段。
开始时细胞皱缩,细胞连接消失,内质网和染色体都发生变化,但细胞膜依然完整。
伴随着核膜破裂,染色体断裂为大小不等的片段,并与一些细胞器聚集后被内折的细胞膜包围,在细胞膜表面形成泡状和芽状突起,即凋亡小体。
在细胞凋亡的第三阶段,脱离的凋亡小体最终被周围的细胞吞噬消化。
【答案】 A2.下列与细胞凋亡过盛有关的疾病或现象是()A.儿童早衰B.老年人行动迟缓、抗病力差C.肿瘤的发生D.再生障碍性贫血【解析】【答案】 D3.下列不属于细胞凋亡的实例的是()A.蝌蚪长到一定程度后尾巴消失B.人皮肤表皮细胞的脱落C.白细胞吞噬过多的细菌导致死亡D.脑细胞因缺氧而死亡【解析】凋亡受到严格的由遗传机制决定的程序性调控,是正常死亡。
A、B、C中提到的都是细胞正常死亡(细胞凋亡),只有D中脑细胞因缺氧而死亡属于细胞坏死。
【答案】 D4.下列有关细胞衰老和凋亡的说法中,正确的是()A.健康的成年人体内,每天都有一定数量的细胞凋亡B.细胞凋亡受环境影响大,机体难以控制C.老年人头发变白和白化病都是由酪氨酸酶活性降低引起的D.胚胎发育期,生物体的细胞衰老总与机体衰老同步进行【解析】衰老和凋亡都是细胞的正常生命现象。
在健康的成年人体内,每天都有一定数量的细胞凋亡。
细胞凋亡是一种程序性的死亡,是受机体基因组调控的细胞正常的死亡。
白化病是基因突变导致缺少合成酪氨酸酶的基因引起的。
胚胎发育期细胞的衰老并不代表机体的衰老,从整体上看,个体衰老的过程是组成个体的细胞普遍衰老的过程。
【答案】 A5.IAPs是细胞内一种控制细胞凋亡的物质,其作用原理是与细胞凋亡酶结合,从而达到抑制细胞凋亡的目的。
细胞衰老和细胞凋亡的区别

动物遗传育种与繁殖杨丰硕学号 1418302025细胞衰老和细胞凋亡的区别细胞凋亡是指由细胞基因调控的一种自主性自杀现象; 或者说细胞凋亡是细胞在一定的生理或病理条件下, 遵循自身的程序, 自我结束生命的死亡方式。
由于这种死亡方式是由基因调控引发的, 因此也被称为程序性细胞死亡.细胞凋亡会使细胞质、细胞核和细胞膜发生一系列生物化学和物理上的变化。
在细胞凋亡早期, 细胞膨胀变圆, 与邻近细胞的联系断绝并且脱离后皱缩。
在细胞质中, 内质网肿胀积液形成液泡。
在细胞核内, 染色质逐渐凝集成新月状, 附在核膜周边,嗜碱性增强。
最终细胞核裂解为由核膜包裹的碎片。
在细胞膜上, 细胞结点不再相连, 细胞膜变得更活跃进而发生内陷。
这些变化都将导致细胞裂解为由细胞膜包裹细胞内容物的凋亡小体。
在生理条件下, 细胞膜上发生特定的调节作用, 这可以使吞噬细胞识别并吞噬凋亡小体。
在细胞发生凋亡的过程中不会伴有细胞内容物渗漏和炎症反应。
外, 与细胞凋亡相比, 细胞坏死将导致细胞器的崩解、细胞膜破损, 大量细胞内容物渗漏。
但在体外培养的细胞中, 坏死的细胞能导致大量细胞凋亡, 这为具有吞噬功能的细胞发挥吞噬功能创造了条件, 这一机制被认为是对缺乏专业吞噬细胞的一种有力补充。
在体内, 正在死亡的细胞是很难被观察到的, 这是由于凋亡细胞被其邻近的细胞在无任何明显征兆的情况下吞噬和消化。
为了保持细胞内环境的稳定, 细胞群落依靠凋亡机制在增殖与消减之间保持着严格的平衡。
细胞凋亡是机体正常细胞在受到生理和病理性刺激后出现的一种自发的死亡过程,是一个主动、高度有序、基因控制及一系列酶参与的过程。
细胞凋亡在保证多细胞生物健康生存过程中扮演着关键角色,对个体的正常发育具有重要作用。
它在多细胞生物的组织分化、器官发育、机体稳态的维持中有着重要的意义。
机体在产生新生细胞的同时,衰老和突变的细胞通过凋亡机制而被清除,使器官和组织得以正常地发育和代谢。
细胞衰老与凋亡优秀课件

❖ 1963年,Orgel 提出差误理论认为,在信息转移的各个步骤(如转录和 翻译)中发生的差误,可以造成有缺陷蛋白的积累。当误差的产生超过某 一阈值时,引起机能失常,出现衰老和死亡。
限的(60~70代)。 细胞增殖次数与端粒DNA长度有关。Harley等发现人体内成纤维细
胞端粒每年约缩短14~18bp,而外周血淋巴细胞则每年缩短33bp。
2、重复基因失活学说:哺乳动物rRNA基因数随年龄而减少。
3、衰老基因学说
❖ 子女的寿命与双亲的寿命有关; ❖ 各种动物都有相当恒定的平均寿命和最高寿命;
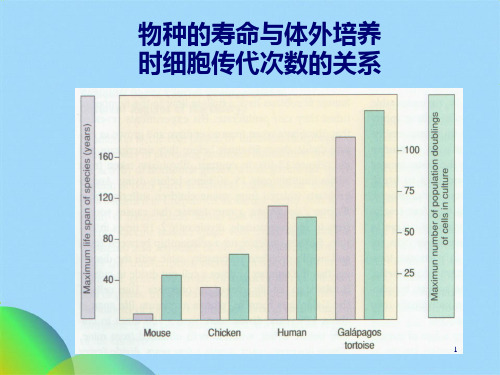
Hayflick等发现,动物体细胞在体外可传代的次数,与 物种的寿命有关。
物种 小鼠 鸡 龟
表 细胞的寿命与物种的寿命的关系
寿命(年) 培养细胞传代次数
3200
140
物种的寿命与体外培养时细胞传代次数的关系
二、细胞衰老的特征----形态和生理生化
1. 细胞水分的减少 , 导致细胞脱水收缩,体积变小。
➢ 细胞,至少是培养的细胞,不 是不死的,而是有一定的寿命 的;它们的增殖能力不是无限 的,而是有一定的界限,这就 是著名的Hayflick界限。
体外培养的年轻和老的人成纤维细胞的显微形态
✓ 左边是只分裂了几代的年轻成纤维细胞,呈 现薄层、细长的形态;
✓ 右边是分离了50次的衰老的成纤维细胞,开 始衰退,并很快死亡。
成人早衰症:平均39岁时出现衰老,47岁生命结束; 婴幼儿早衰症:1岁时出现明显的衰老,12~18岁生命结束, 早衰症患者体内解旋酶发生突变 。 Caenrhabditis elegans的平均寿命仅3.5天,该虫age-1单基因突变,
细胞衰老与凋亡教案

细胞衰老与凋亡教案(实用版)编制人:__________________审核人:__________________审批人:__________________编制单位:__________________编制时间:____年____月____日序言下载提示:该文档是本店铺精心编制而成的,希望大家下载后,能够帮助大家解决实际问题。
文档下载后可定制修改,请根据实际需要进行调整和使用,谢谢!并且,本店铺为大家提供各种类型的实用范文,如学习资料、英语资料、学生作文、教学资源、求职资料、创业资料、工作范文、条据文书、合同协议、其他范文等等,想了解不同范文格式和写法,敬请关注!Download tips: This document is carefully compiled by this editor. I hope that after you download it, it can help you solve practical problems. The document can be customized and modified after downloading, please adjust and use it according to actual needs, thank you!In addition, this shop provides various types of practical sample essays, such as learning materials, English materials, student essays, teaching resources, job search materials, entrepreneurial materials, work examples, documents, contracts, agreements, other essays, etc. Please pay attention to the different formats and writing methods of the model essay!细胞衰老与凋亡教案通过调查人体衰老的特征,讨论如何延缓衰老,延长寿命,引出个体衰老与细胞衰老的关系和细胞衰老的特征。
细胞的衰老和凋亡ppt课件

目
CONTENCT
录
• 细胞衰老 • 细胞凋亡 • 细胞衰老与凋亡的相互关系 • 抗衰老与抗凋亡的策略 • 研究展望
01
细胞衰老
定义与特征
定义
细胞衰老是指细胞在生理和形态上随年龄增长而发生退行性变化 的现象。
特征
细胞体积减小、细胞核增大、染色质固缩、细胞器数量减少等。
03
细胞衰老与凋亡的相互关系
细胞衰老与凋亡的关联
细胞衰老和凋亡是细胞生命周期中的两个重要过程 ,它们之间存在密切的关联。
细胞衰老通常是指细胞停止分裂并进入一种特殊的 状态,而凋亡则是指细胞主动地自我消亡的过程。
细胞衰老可以诱导细胞凋亡,而凋亡也可以促进细 胞衰老。
细胞衰老与凋亡的调控
02
01
03
细胞衰老和凋亡的调控涉及多个复杂的信号通路和分 子机制。
基因、环境因素和应激条件等都可以影响细胞衰老和 凋亡的过程。
调控细胞衰老和凋亡对于维持细胞稳态和组织功能具 有重要意义。
细胞衰老与凋亡在疾病中的作用
细胞衰老和凋亡在多种疾病的 发生和发展过程中发挥重要作 用。
细胞衰老可以促进肿瘤的发生 和发展,而凋亡则可以清除异 常细胞,防止肿瘤的形成。
药物治疗
药物治疗是通过药物干预来调节细胞衰老和凋亡的过程。一 些药物可以延缓细胞衰老或抑制细胞凋亡,从而延长细胞寿 命并保持细胞功能。
药物治疗的策略包括使用抗衰老药物、抗炎药物、抗氧化剂 等。这些药物可以通过不同的机制来发挥作用,如抑制自由 基的产生、调节细胞信号转导通路等。
生活方式调整
生活方式调整是通过改变日常行为和生活习惯来抗衰老和 抗凋亡。一些健康的生活方式可以延缓细胞衰老或抑制细 胞凋亡,从而延长细胞寿命并保持细胞功能。
细胞衰老与凋亡(共34张PPT)
凋亡小体的形成 ◆DNA电泳:DNA片段就呈现出梯状条带
物种的寿命与体外培养 时细胞传代次数的关系
1
早老症儿童与正常儿童的比较
2
细胞衰老的特征
◆细胞内水分减少 ◆色素生成和色素颗粒沉积 ◆细胞核 ◆内质网 ◆细胞质膜 ◆线粒体 ◆蛋白质合成的变化
3
◆ 细胞核的变化
●体外培养的二倍体细胞,细胞核随 着细胞分裂次数的增加不断增大
●细胞核的核膜内折(invagination)、 染色质固缩化
·凋亡信号通路 当细胞接受凋亡信号分子(Fas,TNF等)后,凋亡细胞表面信号分子受体相
互聚集并与细胞内的衔接蛋白(Adaptor protein)结合,这些衔接蛋白又募 集Procaspases聚集在受体部位,Procaspase相互活化并产生级联反应,使细胞
凋亡
·下游Caspases活化后,作用底物: 裂解核纤层蛋白,导致细胞核形成凋亡小体; 裂解DNase结合蛋白,使DNase释放,降解DNA形成DNA Ladder; 裂解参与细胞连接或附着的骨架和其他蛋白,使凋亡细胞皱缩、脱落, 便于细胞吞噬; 导致膜脂PS重排,便于吞噬细胞识别并吞噬。
◆彗星电泳法(comet assay) ◆流式细胞分析
根据凋亡细胞DNA断裂和丢失,采用碘化丙啶使DNA 产生激发荧光,用流式细胞仪检出凋亡的亚二倍体细
胞,同时又能观察细胞的周期状态。
高中生物第五章细胞增殖、分化、衰老和凋亡第二节细胞分化、衰老和凋亡第2讲细胞的衰老和凋亡课件苏教版必

无力
线粒体____变__大___,数目____减__少_____,呼 吸酶活性______降__低________,细胞内呼吸 速率_______减__慢_______
吸收能力下降
细胞膜通透性改变
衰老细胞的特征
水:含量减少,特别是自由水的减少, 使代谢速度降低
1.成分 酶:活性降低,代谢速度降低 染色质:凝聚、固缩,染色加深
(2)细胞核体积_______增__大_______,核膜皱折,____染__色__质____ 固缩。 (3)细胞膜_____通__透__性_______改变,物质运输功能降低。 (4)细胞内多种酶的活性_______降__低_______,呼吸速率减慢, _______色__素_______逐渐积累;细胞内_____物__质__交__流_____和信 息传递受阻。
(2)凋亡小体的形成——______细__胞__核______裂解,____细__胞__膜__ 内陷,将细胞分割成多个有膜包裹的_____凋__亡__小__体_____。 (3)细胞的解体——_____凋__亡__小__体_____逐渐与细胞分离,脱落 至细胞间质,最终被邻近的细胞或专门的_____巨__噬__细__胞_____ 吞噬并消化。 3.意义:细胞凋亡是一种正常的生理过程。对生物正常的 ____个__体__发__育______、生命活动稳定状态的维持以及抵御外界 各种因素的干扰等都具有重要作用。
易错警示 影响细胞衰老的因素
细胞的衰老主要是由细胞自身的因素决定,但良好的外部环 境和生活饮食习惯可以延缓细胞衰老,进而延缓机体衰老。
主题二 细胞的凋亡
细胞凋亡对多细胞生物体完成正常发育,维持内部环境的稳 定,以及抵抗外界因素干扰起着关键的作用。 1.下图是人手掌的发育过程图
细胞衰老与凋亡案例教案

细胞衰老与凋亡案例教案章节一:引言1.1 教案目标:使学生了解细胞衰老与凋亡的概念及其在生命过程中的重要性。
1.2 教学内容:细胞衰老与凋亡的定义,细胞衰老与凋亡的意义。
1.3 教学方法:讲授法,案例分析法。
1.4 教学步骤:1.4.1 引入话题:通过展示细胞衰老与凋亡的实例,引发学生对细胞衰老与凋亡的兴趣。
1.4.2 讲解细胞衰老与凋亡的定义:介绍细胞衰老与凋亡的基本概念。
1.4.3 分析细胞衰老与凋亡的意义:阐述细胞衰老与凋亡在生命过程中的重要作用。
章节二:细胞衰老2.1 教案目标:使学生了解细胞衰老的特点及其机制。
2.2 教学内容:细胞衰老的特点,细胞衰老的机制。
2.3 教学方法:讲授法,案例分析法。
2.4 教学步骤:2.4.1 回顾细胞衰老与凋亡的概念:巩固学生对细胞衰老与凋亡的基本理解。
2.4.2 讲解细胞衰老的特点:介绍细胞衰老的主要表现。
2.4.3 分析细胞衰老的机制:讲解细胞衰老的分子机制。
章节三:细胞凋亡3.1 教案目标:使学生了解细胞凋亡的特点及其机制。
3.2 教学内容:细胞凋亡的特点,细胞凋亡的机制。
3.3 教学方法:讲授法,案例分析法。
3.4 教学步骤:3.4.1 回顾细胞衰老与凋亡的概念:巩固学生对细胞衰老与凋亡的基本理解。
3.4.2 讲解细胞凋亡的特点:介绍细胞凋亡的主要表现。
3.4.3 分析细胞凋亡的机制:讲解细胞凋亡的分子机制。
章节四:细胞衰老与凋亡的调控4.1 教案目标:使学生了解细胞衰老与凋亡的调控机制。
4.2 教学内容:细胞衰老与凋亡的调控因子,细胞衰老与凋亡的调控途径。
4.3 教学方法:讲授法,案例分析法。
4.4 教学步骤:4.4.1 回顾细胞衰老与凋亡的概念:巩固学生对细胞衰老与凋亡的基本理解。
4.4.2 讲解细胞衰老与凋亡的调控因子:介绍调控细胞衰老与凋亡的关键因子。
4.4.3 分析细胞衰老与凋亡的调控途径:讲解细胞衰老与凋亡的调控途径。
章节五:细胞衰老与凋亡的应用5.1 教案目标:使学生了解细胞衰老与凋亡在医学和科研领域的应用。
细胞的分化、衰老和凋亡

在普通高中《生物课程标准》的内容标准中,“分子与细胞”模块的第四学习单元是“细胞的增殖”,第五学习单元是“细胞的分化、衰老和凋亡”。
同细胞代谢一样,细胞的增殖、分化、衰老和凋亡,也是细胞生命活动的具体表现。
笔者根据这两个学习单元的具体内容标准和活动建议,对其教学内容、教学目标、教学策略的构思如下:1.具体内容标准及其地位和教育意义1.1具体内容标准及地位在高中生物新课程的标准中,“细胞的增殖”学习单元的具体内容标准是:简述细胞的生长和增殖的周期性;描述细胞的无丝分裂;观察细胞的有丝分裂并概述其过程。
“细胞的分化、衰老和凋亡”学习单元的具体内容标准是:说明细胞的分化;举例说明细胞的全能性;探讨细胞的衰老和凋亡与人体健康的关系;说出癌细胞的主要特征、讨论恶性肿瘤的防治。
细胞的增殖、分化、衰老和凋亡等生命活动,对生物体的生存和发展具有重要意义。
细胞增殖是生物体生长、发育、生殖和遗传的基础;细胞分化是个体发育的一个重要阶段,胚层细胞的分化导致组织形成、器官发生和系统建成,细胞的异常分化会导致癌变;细胞的衰老和凋亡与人体健康也有着极为密切的关系。
因此,学生学习本单元知识,不仅有利于认识生命的本质,而且为继续学习生殖和发育、基因的表达和调控、基因突变和人类遗传病等知识打下必要的基础。
1.2 具体内容标准的教育意义对照“标准”中的学习目标分类说明表可知,细胞的增殖、分化、衰老和凋亡的具体内容标准涉及8项具体要求,知识性目标有5项,其中的3项是了解层次,2项是理解层次,其他3项属于情感性目标的经历水平。
在“分子与细胞”模块的所有学习单元中,只有“细胞的增殖”和“细胞的分化、衰老和凋亡”的具体内容标准中明确地提出了情感性目标要求,这是因为这部分知识与人体健康、疾病以及社会老龄化问题有关。
珍爱生命、关注健康、尊重和爱护老人是现代公民的基本素养,因此,这部分知识对培养学生情感和提高社会公德有着深刻的教育意义。
我们知道,细胞代谢是生命实现自我更新的生理基础,细胞的增殖、分化、衰老和凋亡等生命活动与其代谢水平密切联系。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
5)膜粘度增加,流动性降低
衰老细胞膜选择通透性改变,物质进出膜的速度下降,因为衰 老或有缺陷的膜常处于凝胶相或固相,产生刚性体脂肪酸尾巴 不能运动,膜流动性差;年轻健全的细胞膜具典型的液晶相, 比较柔韧使脂肪酸尾巴可以自由运动。
6)细胞连接减少
衰老时细胞的间隙连接明显减少,用3H-U标记脲嘧啶核苷酸, 通过放射自显影来研究不同年龄细胞的间隙连接交换情况,将 带3H标记和不带标记的年轻或年老细胞分别放在一起培养,发 现培养4hr后,有70%的年轻细胞从共同培养的年轻供体细胞得 到3H-U,只有30%的年老细胞从共同体培养的年老供体细胞得到 3H-U。说明衰老时细胞间的代谢协作关系显著减少。
2、细胞凋亡
细胞死亡是生物界普遍现象,人体内细胞注定要死亡,有些死 亡是生理性的,有些是病理性的。 细胞死亡分类; 坏死(necrosis),细胞突发性病理的死亡或生理环境急骤变 化(如高热,缺氧等)所致。 凋亡(apoptosis),个体发育过程中基因调控下的细胞自杀活 动,是细胞有机体为调控机体发育,维持内环境稳定,由基因 编码程序的细胞主动性死亡过程。 细胞凋亡对机体的许多正常生理功能具有重要意义。 细胞凋亡与众多疾病发生有密切关系。研究细胞凋亡生物学机 制将可能揭示许多疾病的致病机制,为新治疗药物研发提供理 论依据。
1.1、细胞动态分类
根据细胞的增殖能力,分化程度,生存时间,组织细胞 分为4类:
①更新组织细胞:执行某种功能的特化细胞,经过一定时间后衰 老死亡,由新细胞分化成熟补充。(上皮细胞、血细胞) ②稳定组织细胞,分化程度较高的组织细胞,功能专一,正常情 况下没有明显的衰老现象,细胞分裂少见,但在某些细胞受到破 坏丧失时,其余细胞也能进行分裂,以补充失去的细胞,(肝、 肾细胞) ③恒久组织细胞,属高度分化的细胞,个体一生中没有细胞更替, 破坏或丧失后不能由这类细胞分裂来补充。(神经细胞,骨骼细 胞和心肌细胞 ) ④可耗尽组织细胞,如人类的卵巢实质细胞,在一生中逐渐消耗, 而不能得到补充,最后消耗殆尽。
细胞膜起泡,细胞体积缩小 蛋白质聚集 染色质逐渐聚集成新月状附在核膜周边,嗜碱性增强, 进而断裂,但胞膜仍完整 胞膜不断出芽、脱落,细胞变成数个大小不等的由膜包 裹的凋亡小体 凋亡小体被周围有吞噬功能的细胞如巨噬细胞、上皮细 胞等吞噬、降解
细胞凋亡的形态学变化
(3)细胞凋亡的生物化学变化
第五章 细胞衰老与凋亡
1、细胞衰老
1961年Hayflick等人研究发现,培养细胞是有一定寿命的, 它们的增殖能力有一定限度,在一个范围常数之内 ──Hayflick常数,又称Hayflick界限。
动物 培养细胞传代 寿命(年) 小鼠 12 3 鸡 25 30 龟 140 200 人(胚胎成纤维细胞) 50~60 100~ 人(新生儿成纤维细胞) 30~40 人(青年成纤维细胞) 20~30 人(成年成纤维细胞) 10~30
1998年Garman等人研究认为叙利亚仓鼠胚细胞在复制分裂各阶 段始终表达端粒酶,端粒长度并没变短,经过20-30代分裂后, 细胞依照衰老。有人将去除端粒酶基因的小鼠传了5代,止今 仍未发现寿命比一般鼠要短。故此端粒钟学说还需有更多的实 验依据。
染色体外rDNA环(ERC)的积累
以一种酵母细胞为材料进行的,认为细胞衰老的步伐是 由rDNA的变化决定的。当酵母菌达到生命“中年”的某 一时刻时,经过同源重组产生的第一份环状rDNA片段就 会冲出染色体,成为染色体外的rDNA环,随着细胞不断 分裂,这种rDNA环不断复制增加,当积累到500-1000份 拷贝时,细胞就进入了衰老。双向电泳试验也证明,衰 老细胞中积累有许多密闭的环状rDNA,而年轻细胞中含 量极少。故此染色体外rDNA的积累,导致了细胞的衰老。
衰老细胞的形态变化
核 染色质 质膜 细胞质 线粒体 高尔基体 尼氏体 包含物 核膜 增大、染色深、核内有包含物 凝聚、固缩、碎裂、溶解 粘度增加、流动性降低 色素积聚、空泡形成 数目减少、体积增大、mtDNA突变或丢失 碎裂 消失 糖原减少、脂肪积聚 内陷
(二)细胞生理变化
胞内水分减少,细胞萎缩,体积变小,与正常细胞相比 衰老细胞含水量明显下降细胞皱缩。 代谢速度减慢,生活力降低,检测发现有些酶的活性下 降。 呼吸速率减慢,衰老细胞的呼吸强度下降。
daf2(编码胰岛素受体)和age1(编码磷脂酰肌醇-3-激酶P110催化亚 基):正常状态下两个基因表达,决定了正常的成虫发育。
daf16:编码一种转录活化剂:在饥饿或虫口过密的情况下,大量表达, 并朝着dauer幼虫方向发育。
在daf2和age1突变体中,daf2和age1部分表达,导致决定成虫发育和 dauer幼虫特异性防御机制同时表达,完成正常的成虫发育,又获得了 寿命延长特性(一般延长2-4倍)。
1.3、细胞衰老的机理
氧化损伤:氧自由基引发氧化性损伤积累。
端粒的变化:细胞衰老的“有丝分裂钟”
rDNA变化:染色体外rDNA 环(ERC)的积累。 沉默信息调节蛋白复合物:异染色质区阻断DNA转录。 SGS1基因、WRN基因: 编码解旋酶,sgs1突变体寿命明显短于野生型(平均9.5
2.1、简介及基本概念
(1)细胞凋亡:
细胞凋亡的概念最早是由病理学家Kerr于1972年正式提 出
又被称为“细胞程序性死亡”(programmed cell death, PCD)
细胞凋亡是一种具有形态特征和生化改变的细胞凋亡过 程,这些改变是一凋亡形态学变化
发育程序与衰老
线虫的生命周期:幼虫期为4天,然后达到性成熟并进入成虫期,延续数周后, 进入衰老与死亡。
幼虫发育过程中遇到不良环境(虫口过密、食物短缺和高温),引起性外激 素浓度变化,结果幼虫进入一种特殊的dauer幼虫阶段,可以延续6个月。 此时幼虫全身被以角质化的层、口紧闭、处于休眠状态; 如环境条件改善,又可发育成成虫,经过数周,完成其生活史。 遗传分析发现,存在着两种Dauer幼虫形成基因:
2)内质网的变化
糙面内质网总量减少,排列有序性下降。年轻动物细胞 中,糙面内质网发育良好,排列有序;年老动物中,内 质网成弥散状分散于核周围胞质中,不存在有序排列。 光镜下用碱性染料染色可以看到神经元细胞中呈颗粒状 的尼氏体。尼氏体是由糙面内质网和核糖体组成的混合 物。研究者发现小鼠和人的大脑及小脑某些神经元中尼 氏体中物质含量会随着动物年龄的增长而逐渐下降,表 明随年龄增大内质网量减少。
1.2、细胞衰老特征
(一)细胞结构的变化
1)细胞核的变化
细胞核增大,通过二倍体细胞培养发现,随着细胞分裂次数的 增加,细胞核会不断增大。 核膜内折,培养细胞和体内细胞,随着培养时间增加,传代次 数增加或年龄增长,核膜内折现象明显增多。 染色质固缩化,培养的晚代细胞核中,染色质固缩化明显,早 代细胞只有轻微的固缩,体内细胞亦有这种现象。 核仁裂解为小体,在sgs1酵母突变体衰老细胞中可看到核仁裂 解为小体。
线粒体DNA
随着年龄增长线粒体DNA的突变显著增加,并表现出异质 性 线粒体DNA的突变可能导致电子传递链不能执行正常功能。 如缺失突变引起细胞色素氧化酶的缺失,ATP水平和 NAD/NADH比率下降,促进ROS的生成。 mtDNA裸露于基质,缺乏结合蛋白的保护,最易受自 由基伤害,而催化mtDNA复制的DNA聚合酶γ不具有校 正功能,复制错误频率高。
Sir复合物
沉默信息调节蛋白复合物(Sir1、Sir2、Sir3、Sir4),一般 存在于异染色质中,该复合物的作用是阻止它们所在位点的 DNA转录。
1994年Kennedy肯尼迪等人在一种酵母突变体发现了Sir3蛋白 和Sir4蛋白由端粒处向着核仁处转移,这种转移伴随着酵母寿 命的延长。进一步研究发现,突变体中的Sir4蛋白的C端有121 个氨基酸被除掉了,除掉的这个区域本来是Sir4蛋白与端粒和 其他蛋白相互作用的部位,由于这个区域的去除,Sir复合物 能够自由定位,并从端粒处转移到了核仁,而Sir复合物到达 核仁,又抑制了rDNA的转录,使核仁rDNA处于沉默状态不能复 制,从而造成染色体外rDNA环(ERC)生成和积累量减少,ERC 量增加细胞衰老加快,减少则意味着细胞寿命延长。
3)线粒体的变化
细胞在衰老过程中,线粒体数量减少,体积增大。在人 类、大鼠、小鼠衰老的肝细胞中线粒体数量明显减少, 并发生膨大现象。
4)致密体的生成
致密体是衰老细胞中常见的一种结构。在不同的教科书 中对这种细胞成分还有一些其他的名称,比如脂褐质、 老年色素(斑)、血褐质、脂色素、黄色素、透明蜡体 及残体等等。研究表明,致密体是由溶酶体或线粒体转 化而来的。
最新研究认为,人体表皮的细胞衰老是由于葡萄糖代谢产生自 由基,它在细胞内乱撞导致细胞因子受损,细胞衰老死亡出现 斑块,自由基进入核撞击基因使基因改变,细胞发生癌变。有 人将小鼠的谷胱甘肽过氧化物酶基因及SOD1、SOD2和SOD3基因 中的一种剔除,并没发现引起细胞衰老。
端粒钟学说
随着细胞的每次分裂,端粒不断缩短,当缩短到一个阈值时, 细胞就进入了衰老。1998年Wright等人将人的端粒酶反转录酶 亚基的基因引入正常人的二倍体细胞,发现传染细胞的端粒长 度明显增加,分裂旺盛,同时作为细胞衰老标志的β半乳糖苷 酶的活性明显降低,细胞寿命较正常细胞增加了至少20代,这 一证据说明端粒长度与细胞衰老有着密切的关系。
SGSI基因 及WRN基因
酵母细胞中发现了一种突变体,该突变体因突变缺失了SGSI基 因,而且寿命短,野生型平均分裂24.5代,突变体平均分裂 9.5代,表明SGSI基因缺失影响了酵母的寿命。SGSI蛋白合成 后定位在核仁中,作用是阻止核仁裂解,如果缺少这种蛋白, 核仁就会裂解为小体。 人类发现了一种早衰疾病——Werner综合症(WS),病人30多 岁会出现一些正常老年人的衰老现象,如动脉硬化、骨质疏松、 Ⅱ型糖尿病、肿瘤等等。从WS病人细胞中分离出了一种突变基 因——WRN基因。WRN基因与酵母突变体SGSI基因同源,WRN蛋 白含有7个DNA解旋酶所特有的标志序列,WRN蛋白突变,可影 响其正常功能,使DNA不能正常解旋复制,由此造成DNA复制的 严重障碍。正常人皮肤的成纤维细胞中,WRN蛋白存在于核仁 中,WS病人的成纤维细胞中看不到典型的WEN蛋白。