浙江大学生物化学与分子生物学笔记生物膜和运输
生物化学[第六章生物膜与物质运输]课程复习
![生物化学[第六章生物膜与物质运输]课程复习](https://img.taocdn.com/s3/m/3fe2d5f9f90f76c661371a44.png)
第六章生物膜与物质运输许多生命现象都直接或间接地依赖于生物膜,如运动、生长、繁殖、代谢等。
在流动镶嵌模型中,生物膜是一个脂质双分子层,蛋白质就在脂质双分子层中流动,膜蛋白在很大程度上决定了膜的生物功能。
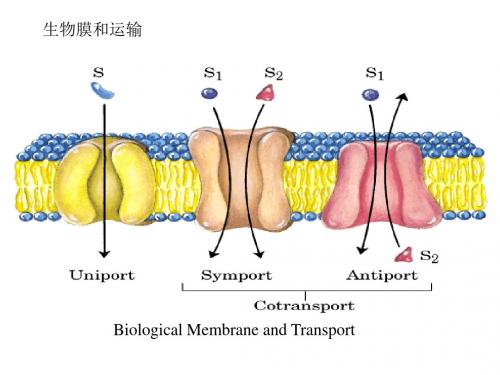
生物膜的一个重要功能就是物质的跨膜运输,根据物质的不同,分成主动运输、被动运输、协助运输和基团运输等方式。
6.3.1膜结构6.3.1.1 膜脂膜脂主要是磷脂,此外还有胆固醇(细菌不含胆固醇)和糖脂。
膜的性质和结构与膜脂的两性性质有关。
通常膜脂具有一个亲水的极性基团(头部)和两条疏水的碳氢链(尾部)。
在水环境中,极性头部与水接触,疏水的碳氢链被水排斥而挤在一起,使膜脂自发形成平行排列的双分子层,并自我封闭成微囊。
除此之外,脂类分子还能聚集在一起形成微团结构。
生物膜中脂类分子通常总是以脂质双层结构存在,或者组成平面膜,或者形成微囊,这是膜结构的基础。
膜脂还与膜的下列性质有关。
(1)膜的流动性膜的流动性包括侧面扩散、自旋转和翻转。
不饱和脂肪酸含量越高,流动性越强,胆固醇能增加膜的稳定性而不显著影响流动性,因为它有一个刚性结构(环)和一个弹性结构(碳氢链尾巴)。
(2)选择透过性由于高度疏水性,膜双分子层对于离子和生物性分子几乎是不可透过的,必须借助于膜蛋白。
要穿过膜,极性物质必须部分或全部释放出它的水化层(hydratuen spaere),结合到载体蛋白上跨膜转运或直接通过水性的蛋白通道。
跨膜的水分运动是与离子运输相结合的,非极性物质直接沿浓梯度扩散又穿过脂双分子层。
(3)自缝合能力当脂双分子层被破坏时,它们能立即自动缝合起来。
(4)不对称性生物膜是不对称的,也就是说双分子层的两上半层的脂的组成是不同的。
例如,人的细胞膜外层含有较多的磷脂酰胆碱和鞘磷脂。
6.3.1.2 膜蛋白生物膜的大部分功能需要蛋白质分子。
膜蛋白根据在膜上的位置可以分成膜周边蛋白和膜内在蛋白质。
膜蛋白以其功能大致可分为五类:酶类、运输蛋白、运动蛋白、信息接受与传递蛋白、支持与保护蛋白。
浙江大学生物化学与分子生物学笔记生物膜和运输

pH改变、螯 合剂、尿素、 碳酸盐可除 去外周蛋白
外周蛋白
嵌入(膜内) 蛋白
外周蛋 白和膜 内 (嵌入) 蛋白
糖蛋白
去污剂
外周蛋白与膜的连接是可逆的
• 许多外周蛋白通过与嵌入蛋白的亲水区 域或膜脂的极性头部以静电作用或氢键 结合到膜上,通过温度的改变或破坏静 电或破坏氢键作用(如加入螯合剂、尿 素、碳酸盐或改变pH)可被释放出来。 • 这些外周蛋白可作为膜结合酶的调节因 子、或作为连接膜内蛋白与胞间结构的 中介物、或一些膜蛋白的流动性。
Байду номын сангаас
病毒进入宿主细胞的膜融合
跨膜运输
所有生物细胞都要从环境获得原材料为其生物 合成和能量消耗,同时还需释放其代谢物到环境 中去。质膜可以识别并允许细胞所需物如糖、氨 基酸、无机离子等进入细胞,有时这些成分进入 细胞是逆浓度梯度的,即它们是被“泵”入细胞 的,同样一些分子是被“泵”出细胞的。很少有 例外小分子物质的跨膜是直接通过蛋白的,而是 通过跨膜的通道(channels)、载体(carriers)或泵 (pumps)。
膜内(嵌入)蛋白与脂 通过疏水作用维系在膜中
嵌入蛋白通常富 含 疏 水 氨 基 酸 区 域 (可在中间段,也可在氨基端或羧基 端),有些可有多个疏水序列,如 - 螺 旋,可横贯整个膜脂双分子层。
膜内(嵌 入)蛋白
(Integral Membrane Proteins)
有些外周膜蛋白 共价泊锚在膜脂上
生物膜中兼性的磷脂和固醇形成一个脂质双分 子层,非极性部分相对构成双分子层的核心,极 性的头部朝外;脂质双分子层结构中,球状蛋白 以非正规间隔埋于其中;另一些蛋白则伸出(突 出)膜的一面或另一面;还有一些蛋白跨越整个 膜。蛋白质在脂双分子层中的方向是不对称的, 表现为膜蛋白功能的不对称。脂质与蛋白质之间 构成一个流动的镶嵌结构。
生物化学下-第21章 生物膜与物质运输-精品文档

一、被动运输与主动运输
第21章 生物膜与物质运输
红细胞葡萄糖转运蛋白介导被动运输(Passive Transport)
葡萄糖进入红细胞是通过葡萄糖特异性转运蛋白的协助扩散来实现的。
红细胞的葡萄糖转运蛋白(glucose transporter, GluT1)结构模型
外周蛋白
固醇
脂双层
膜内在蛋白 (单跨膜螺旋)
与脂质共价连 接的外周蛋白
膜内在蛋白 (多跨膜螺旋)
第21章 生物膜与物质运输
第21章 生物膜与物质运输
生物膜功能:
1. 能量转换 —— 氧化磷酸化(线粒体)、光合磷酸化(叶绿体) 2. 信息识别与传递 —— 生物信号 (G-蛋白偶联受体和第二信使、视
觉、 嗅觉、味觉的信号传导、致癌基因、肿瘤抑制基因等)
一个溶质的耗能运输与另一溶质的释放流
反应偶联。
动相偶联,释放的能量由初级主动运输提
ATP水解释放能量驱动溶质X逆 电化学梯度运动。
供的。 初级主动转运建立X离子的浓度梯度,X顺
电化学梯度移动为第二溶质S逆电化学梯度
主动转运是一种能量消耗途径。
运动提供能量。
如太阳光的吸收、氧化反应、ATP分解等。
一、被动运输与主动运输
③当胰岛素水平下降, GluT4通过胞吞作用重新从 质膜上移回胞内,形成囊泡。
心肌 骨骼肌
肌糖原
脂肪细胞 三酰甘油
这些细胞膜含有葡萄糖转运蛋白GluT4
GluT4
① GluT4贮存在细 胞内的囊泡膜上。
Ⅰ型糖尿病人不能释放胰岛素,导致 肌肉和脂肪组织对葡萄糖的吸收率很 低,后果血糖增高。
最新办公文档浙江大年夜师长教师物化学与分子生物学笔记生物膜和运输-药学医学精品资料
膜内(嵌入)蛋白与脂 通过疏水作用维系在膜中
• 嵌入蛋白通常富含疏水氨基酸区 域(可在中间段,也可在氨基端或羧基 端),有些可有多个疏水序列,如 -螺 旋,可横贯整个膜脂双分子层。
膜内(嵌 入)蛋白 (Integra l Membran e Proteins)
有些外周膜蛋白 共价泊锚在膜脂上
• 有些膜外周蛋白与膜脂有一个或 多个共价结合位点,如长链脂肪酸、或 磷脂酰肌醇糖基化衍生物。连接的脂提 供了一个疏水的锚以插入脂双分子层。
膜结构的流动镶嵌模型 (Fluid Mosaic Model)
脂双分子层是基本的结构
• 脂类与水相共存时会迅速形成一种脂双 分子层结构而避开水的作用,生物膜的 厚度(电镜测定为5-8 nm)是由3 nm的 脂双分子层和蛋白的厚度决定的,所有 证据都支持生物膜由脂双分子层构成。 • 膜脂对于脂双分子层两面是不对称的, 但尽管不对称,也不象蛋白质,脂的不 对称不是绝对的。
每种膜都有一个特征的脂质组成
• 各种来源的膜的化学分析显示了一 个共同的特征,即膜脂组成因不同的界、 不同的种、不同的组织、特定细胞中不同 的细胞器而不同。细胞有一种清楚的机制, 可以精确控制膜脂合成的种类和数量,以 及定位到特定的细胞器上。
不同组织质膜的主要成分
鼠肝细 胞膜及 细胞器 膜的脂 质类型
温度引起侧 链热运动
脂双层平面 的扩散 跨膜扩散 :“翻跟 头”
膜脂的 运动
膜蛋白透过或跨过脂双分子层
• 生物膜的冰冻蚀刻电镜观察到的 一个蛋白或多蛋白复合体分布情况显示, 一些蛋白仅存在于膜的一面,另一些则 横跨整个膜双分子层,有些穿过膜的另 一表面。 • 膜蛋白在脂双分子层上可侧向运 动。
N-连接 O-连接四 糖: 2Neu5Ac ,Gal, GalNAc -螺旋
生物化学必看知识点总结优秀

引言概述:生物化学是研究生物体内化学成分的组成、结构、功能以及各种生物化学过程的机理的学科。
掌握生物化学的基本知识是理解生物体内各种生命现象的基础,也是进一步研究生物医学、生物工程等领域的必备知识。
本文将从分子生物学、酶学、代谢、蛋白质和核酸等五个方面,总结生物化学中必看的知识点。
正文内容:1.分子生物学1.1DNA的结构和功能1.1.1DNA的碱基组成1.1.2DNA的双螺旋结构1.1.3DNA的复制和转录过程1.2RNA的结构和功能1.2.1RNA的种类和功能区别1.2.2RNA的结构和特点1.2.3RNA的转录和翻译过程1.3蛋白质的结构和功能1.3.1氨基酸的结构和分类1.3.2蛋白质的三级结构和四级结构1.3.3蛋白质的功能和种类1.4基因调控1.4.1转录调控和翻译调控1.4.2基因的启动子和转录因子1.4.3RNA的剪接和编辑1.5遗传密码1.5.1遗传密码的组成和特点1.5.2密码子的解读和起始密码子1.5.3用户密码监测2.酶学2.1酶的分类和特点2.1.1酶的命名规则和酶的活性2.1.2酶的结构和功能2.1.3酶的催化机制2.2酶促反应动力学2.2.1酶反应速率和反应速率常数2.2.2酶的最适温度和最适pH值2.2.3酶的抑制和激活调节2.3酶的应用2.3.1酶工程和酶的改造2.3.2酶在医学和工业上的应用2.3.3酶和药物相互作用3.代谢3.1糖代谢3.1.1糖的分类和代谢路径3.1.2糖酵解和糖异生3.1.3糖的调节和糖尿病3.2脂代谢3.2.1脂的分类和代谢途径3.2.2脂肪酸的合成和分解3.2.3脂的调节和脂代谢疾病3.3氮代谢3.3.1氨基酸的合成和降解3.3.2尿素循环和氨的排出3.3.3蛋白质的降解和合成3.4核酸代谢3.4.1核酸的合成和降解途径3.4.2核酸的功能和结构特点3.4.3DNA修复和基因突变3.5能量代谢调节3.5.1ATP的合成和利用3.5.2代谢途径的调节和平衡3.5.3能量代谢和细胞呼吸4.蛋白质4.1蛋白质的结构和维持4.1.1蛋白质结构的层次和稳定性4.1.2蛋白质质量控制和折叠4.2蛋白质表达和合成4.2.1蛋白质的翻译和翻译后修饰4.2.2蛋白质的定位和运输4.2.3蛋白质合成的调节和失调4.3蛋白质与疾病4.3.1蛋白质异常与疾病的关系4.3.2蛋白质药物和治疗策略4.3.3蛋白质组学在疾病研究中的应用5.核酸5.1DNA的复制和修复5.1.1DNA复制的机制和控制5.1.2DNA损伤修复和维持稳定性5.1.3DNA重组和基因转座5.2RNA的合成和调控5.2.1RNA转录的调节和翻译5.2.2RNA剪接和编辑5.2.3RNA和疾病的关系5.3RNA干扰和基因沉默5.3.1RNA干扰机制和调控5.3.2RNA干扰在基因治疗中的应用5.3.3RNA沉默和抗病毒防御总结:生物化学是研究生物体内化学成分和生物化学过程的重要学科,掌握其中的关键知识点对于理解生命的本质和生物体的正常功能至关重要。
高等生物化学第七章生物膜与跨膜转运

7.4 溶质的跨膜转运
▪ 离子选择通道对特定离子具有高选择性; ▪ 离子通道有开放和关闭两种状态,在两种状态中相
互转变;并且可调控。 ▪ 离子通道的分类:
–配体门控离子通道; –电压门控离子通道。
▪ 离子通道与离子转运体的差别:
–离子通道的流速比转运体高若干个数量级,且无限接近 扩散的理论最大值;
双分子层平面里,有极大的流动性。
–翻转运动
▪ 概率小,需能,翻转酶可促进
–旋转运动 –交换 –脂肪链的运动
7.3 膜的超分子结构
▪ 生物膜的流体镶嵌模型 [Singer & Nicholson (1972), Science 1972 Feb. 18 175:23 720-731]
– 描述了蛋白质与脂双层的关系; – 膜蛋白好像液体脂分子海洋中的圆形冰山; –整体蛋白跨越脂双层,通过疏水作用结合在膜上; –外周蛋白与膜表面松散地结合(如:通过静电作
7.4 溶质的跨膜转运
▪ 细胞膜的半透性:
–亲脂性的小分子,不带电荷的水分子可以通过; –带电荷的,极性的分子不能通过。
▪ 膜蛋白可以辅助特定分子的跨膜转运,转运 方式包括:
–被动运输;如:离子通道。 –主动运输;如:离子泵。 –胞吞胞吐作用。
7.4 溶质的跨膜转运
图7-18 溶质的浓度梯度
•浓度梯度是指膜两 侧的浓度差 (C2-C1)。
▪ 将一个红细胞放在低渗的溶液中(如:纯水),红 细胞便会因水的内流而破裂(溶血)。
▪ 植物细胞与细菌细胞由于有坚固的细胞壁,可避免 在低渗溶液中破裂。这会使细胞内产生膨压。当膨 压与渗透压相等时,反渗透便会停止。
7.4 溶质的跨膜转运
图7-22 葡萄糖转运体1(Glu T1)的建议结构。(a) 12个跨膜螺旋中有9个含有3个以上 的极性或带电的氨基酸残基,通常被几个疏水残疾隔离;
浙江大学生物化学与分子生物学笔记a2_核酸化学_rna

绿色标识表 示平均频率 请对照各氨基酸在二级结 构中出现的频率
RNA的加工类型 1. Cleavage 2. Trimming 3. Splicing 4. Editing 5. Modification
RNA的功能
1. 遗传密码的中间担体(mRNA) 2. mRNA剪接等加工的活性成份(snRNA) 3. 核糖体的骨架结构及与mRNA的识别 (rRNA) 4. 活化氨基酸的担体(tRNA) *5. 酶的活性成份 *6. 病毒基因组的担体
Crick的中心法则
DNA
1957年 转录
DNA
反转录 1970年
4. RNA分子可以通过由蛋白质酶催化的反应转化 为DNA分子 (Ribonucleoside diphosphate reductase)。 5. RNA分子在细胞中同时扮演着信息的媒介作用 和执行者的作用,mRNA为前者,rRNA、 tRNA和snRNA为后者。因此很自然的认为生 命的早期可以称为RNA world。 6. RNA分子比DNA分子更容易受到修饰,均有其 用途(作用), 如mRNA的5’帽子能启动蛋白质 的翻译,tRNA上的修饰可以增加tRNA的寿命。 7. 真核细胞的基因拥有令人吃惊的特点,顺序中 插入了很多不编码氨基酸的废物(Introvening sequence, 或叫内含子),这些序列得到转录后 在mRNA成熟之前被除去。Fra bibliotekRNA
翻译
RNA
Protein
Protein
细胞中的RNA
原核生物的基因结构
转录起始 DNA
转录终止
启动子
结构基因1
结构基因1
转录终止子
结构基因1
RNA转录自DNA
真核生物的基因结构
动物生物化学6生物膜与物质运输

STEP2
STEP1
生物膜的基本结构是脂质双层,蛋白质或镶嵌在膜上或结合在膜的表面,膜上的寡糖链总是指向膜的胞外一侧.
膜上的成分是运动的,随温度变化,脂质双层呈液晶态或凝胶态.膜的相变温度(Tm)与膜上脂肪酸烃链的长度和饱和程度有关。
脂质双层的组成成分呈不对称分布.
要点:
3.1 物质运输的功能
微团
双分子层
脂质体
膜的基本结构——脂质双层(Lipid bilayer)
water
water
膜蛋白
分布于双层脂膜的外表层。 与膜的结合比较疏松,容易从膜上分离出来。 外周蛋白比较亲水,能溶解于水。
外周蛋白(peripheral protein)
蛋白的部分或全部嵌在双层脂膜的疏水层中。 溶于水,且不容易从膜中分离出来。 主要以-螺旋形式存在。 膜蛋白具有重要的生物功能,是生物膜实施功能的场所。可以分为外周蛋白和内在蛋白。它们是受体,酶,抗原,通道和骨架蛋白等。
内在蛋白(integral protein)
跨膜蛋白(transmembrane protein)
1.3 膜糖
生物膜中的寡糖链信息传递和细胞的相互识别方面具有重要作用。 糖蛋白上的寡糖链总是指向细胞的外面
生物膜上各种化学组成之间的关系
2. 生物膜的性质与结构特点
2.1 生物膜的运动
膜脂分子:分子摆动、旋转运动、侧向运动等。 膜蛋白:扩散运动 、旋转运动。
维持细胞的容积、形态、渗透压、电解质的浓度,为细胞的生理活动提供适宜的环境
从环境摄取营养物质,向环境排出代谢废物
3.物质的过膜转运
3.2 小分子物质和离子的过膜转运
简单扩散(simple diffusion)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
温度引起侧 链热运动
脂双层平面 的扩散 跨膜扩散: “翻跟头”
膜 脂 的 运 动
膜蛋白透过或跨过脂双分子层
生物膜的冰冻蚀刻电镜观察到的一个 蛋白或多蛋白复合体分布情况显示,一 些蛋白仅存在于膜的一面,另一些则横 跨整个膜双分子层,有些穿过膜的另一 表面。 膜蛋白在脂双分子层上可侧向运动。
N-连接
溶质通过透过性膜的移动
不带电
带电
被动运输是由膜蛋白促进的 顺浓度梯度的扩散
• 生物体内的简单扩散因膜把胞内和胞外环境所 阻止,膜是一种选择性通透屏障,要通过脂双 分子层,极性分子或带电溶质必需解除水化膜 的水的作用,然后透过约3nm 的介质(膜)。 • 水是一种例外,可很快透过生物膜,机制尚不 清楚,膜两侧溶质浓度差异大时,渗透压的不 平衡引起膜两侧水的流动,直至两侧的渗透压 相等。 • 极性溶质或离子的过膜运输由膜上的蛋白降低 活化能而对特异的物质提供过膜路径而过膜的 双分子层,引起促进扩散。
生物膜和运输
Biological Membrane and Transport
生物膜和运输(Biological Membrane and Transport)
生物膜的形成对于生物能量的贮存及 细胞间的通讯起着中心作用。膜的生物 活性来自于膜自身显著的特性:膜连接 紧密但有弹性;膜自我封闭,对极性分 子有选择性通透;膜的弹性允许膜在细 胞生长和运动中改变形状;暂时破裂且 可自封闭的能力可保证两个细胞或两个 膜状包裹物的融合。
三类主要运输系统
两种类型的主动运输
磷酸和钒酸
有三种形式的运输ATPases
Na+K+-ATPase是一种运输蛋白的典型 形式(Prototype)被称为P-type ATPase (可以可逆磷酸化的);另一类为Vtype ATPase—Proton pumps(V-vacuole); 第三种,ATP-splitting proton pumps,对 细菌、线粒体和叶绿体中的能量消耗反 应起中心作用,即F-type ATPases(F: energy-coupling factors),催化质子可逆 地跨膜运输,由ATP水解供能。
细胞体
纤毛
线粒体
膜 的 电 镜 横 切 面 照 片
消化泡 内质网 分泌泡
膜的生化特性
膜不仅仅是被动的屏障,膜上含有一 系列的特化蛋白质启动或催化一定的分 子事件;膜上的泵可以逆跨膜梯度移动 (运送)特定的有机物和无机离子;能 量转化器可以把一种形式的能量转化为 另一种形式的能量;质膜上的受体能够 感受胞外信号,并转化为细胞内的分子 事件。
膜的超分子结构
所有生物膜拥有共同的基本特征:对 多数极性分子或带电分子不通透,允许 非极性分子通透;约5-8 nm厚,横切电 镜照片近似三层结构。
流动镶嵌模型 (Fluid Mosaic Model)
生物膜中兼性的磷脂和固醇形成一个脂质 双分子层,非极性部分相对构成双分子层的核 心,极性的头部朝外;脂质双分子层结构中, 球状蛋白以非正规间隔埋于其中;另一些蛋白 则伸出(突出)膜的一面或另一面;还有一些 蛋白跨越整个膜。蛋白质在脂双分子层中的方 向是不对称的,表现为膜蛋白功能的不对称。 脂质与蛋白质之间构成一个流动的镶嵌结构。
膜的分子组成
生物膜几乎所有的质量都由蛋白质和 极性脂质组成,少量的碳水化合物也是 糖蛋白或糖脂的一部分。蛋白质和脂类 的相对比例因不同的膜而不同,反映着 膜生物作用的广泛性。神经元的髓鞘主 要由脂类构成,表现为一种被动的电子 绝缘体;但细菌、线粒体、叶绿体的膜 上由许多酶催化的代谢过程发生,含有 的蛋白比脂类要多。
1.D-Glc与T1特异 结合降低构象改 变的活化能 2.T1转变为T2影响 Glc跨膜通道
3.Glc由T2释放 到胞质
4.T2构象变回T1
入葡 红萄 细糖 胞运 模输 型进
氯化物和碳酸氢盐跨红细胞 膜的运输为协同运输(Cotransport)
• 红细胞存在另一种促进扩散系统—阴离子交换 体,这对于肌肉及肝脏中CO2 回到肺中的运输 是必需的。呼吸组织产生的废气—CO2 由血浆 进入红细胞,在红细胞中转化为HCO3-,HCO3重新回到血浆中被运输到肺组织。因HCO3- 比 CO2 的溶解度大,这种变化增加了由组织到肺 的CO2 运输的血容量,在肺中HCO3- 重回红细 胞被转化为CO2,被缓慢呼出。 • 氯化物-碳酸氢盐交换体也被称为阴离子交换蛋 白,可增加HCO3-对红细胞膜的透过,这一系 统也被称为协同运输系统。
膜内(嵌入)蛋白与脂 通过疏水作用维系在膜中
嵌入蛋白通常富 含 疏 水 氨 基 酸 区 域 (可在中间段,也可在氨基端或羧基 端),有些可有多个疏水序列,如-螺 旋,可横贯整个膜脂双分子层。
( Integral Membrane Proteins)
膜 内 ( 嵌 入 ) 蛋 白
有些外周膜蛋白 共价泊锚在膜脂上
( Membrrane Fusion)
膜 பைடு நூலகம் 融 合
膜融合事件
• 两个膜融合需要:相互识别;相互表面靠近并相对 (排除水分子);双层结构部分破坏;两个双分子层 融合为一个连续的脂双分子层。受体调节的内吞或控 制的分泌还需要融合发生在合适的时间或者是对特异 信号的反应。 • 融合蛋白(嵌入蛋白)(fusion protein)参与以上融合 事件,引起特异识别和短暂、局部脂双层结构变形促 使膜融合。融合蛋白可搭起两个膜融合的桥,并带来 融合区域脂双分子层的暂时恢复。 • 膜联蛋白(annexin)(一种Ca2+活化后可与膜磷脂结 合的蛋白)是一类紧挨质膜的蛋白质,需要Ca2+,与 脂双分子层的磷脂结合,可通过交叉连接两个不同膜 的脂质分子。
每种膜都有一个特征的脂质组成
各种来源的膜的化学分析显示了一个 共同的特征,即膜脂组成因不同的界、 不同的种、不同的组织、特定细胞中不 同的细胞器而不同。细胞有一种清楚的 机制,可以精确控制膜脂合成的种类和 数量,以及定位到特定的细胞器上。
不同组织质膜的主要成分
鼠 肝 膜细 的胞 脂膜 质及 类细 型胞 器
红细胞质膜内外单层膜 磷脂的不对称分布
不同功能的膜含有不同的蛋白质
• 不同来源膜的蛋白质组成比其脂质组成的变化 更大,反映在膜功能的特化上。脊椎动物视网 膜杆状细胞对于接受光为高度特化,90%以上 的膜蛋白是光吸收蛋白—视紫红质;特化较低 的红细胞质膜约含20种显著的蛋白及十几种较 少的蛋白,其中多数的蛋白为运输载体,每一 种蛋白运输一种跨膜的溶质等。 • 有些膜蛋白还有一个或多个脂共价结合,后者 可能形成一种疏水的稳定体系以保证蛋白质存 在于膜中。
膜结构的流动镶嵌模型 (Fluid Mosaic Model)
脂双分子层是基本的结构
• 脂类与水相共存时会迅速形成一种脂双 分子层结构而避开水的作用,生物膜的 厚度(电镜测定为5-8 nm)是由3 nm的 脂双分子层和蛋白的厚度决定的,所有 证据都支持生物膜由脂双分子层构成。 • 膜脂对于脂双分子层两面是不对称的, 但尽管不对称,也不象蛋白质,脂的不 对称不是绝对的。
红细胞膜膜上氯-碳酸氢盐交换体
主动运输(Active Transport) 引起物质的逆浓度梯度运输
被动运输总是顺浓度梯度运输,不会引起 物质的积累,相反,主动运输总是逆浓度梯度 运输,引起物质的积累。主动运输直接或间接 地依赖于一些放能过程,非热力学自动发生, 往往伴随有光的吸收、氧化作用、ATP水解或 其他顺浓度梯度的运输。在初级主动运输中, 物质的积累直接与放能反应(如ATPADP+Pi) 相连接;次级主动运输发生于由初级主动运输 引起的逆浓度积累的顺浓度梯度运输。
病毒进入宿主细胞的膜融合
跨膜运输
所有生物细胞都要从环境获得原材料为其 生物合成和能量消耗,同时还需释放其代谢物 到环境中去。质膜可以识别并允许细胞所需物 如糖、氨基酸、无机离子等进入细胞,有时这 些成分进入细胞是逆浓度梯度的,即它们是被 “泵”入细胞的,同样一些分子是被“泵”出 细胞的。很少有例外小分子物质的跨膜是直接 通过蛋白的,而是通过跨膜的通道 (channels)、载体(carriers)或泵(pumps)。
三种类型离子运输 ATPase亚基结构
四种类型的运输ATPases
F类型ATPases
+K+ Na
ATPase
主动协同Na+,K+的运输 由ATP所供能
每个动物细胞与环境相比维持较低的 [Na+]和较高的[K+],这种不平衡由质膜 上的主动运输所建立和维持,涉及 Na+K+-ATPases,偶联ATP水解,引起 Na+,K+的逆浓度梯度运输。每水解1ATP, 偶联运输2个K+进质膜内,3个Na+出质膜 外。Na+K+-ATPases是一种膜蛋白,由2 个亚基组成,都是跨膜蛋白。
Na+ K+ ATPase
pH改变、螯 合剂、尿素、 碳酸盐可除 去外周蛋白
外周蛋白
嵌入(膜内) 蛋白
外 (周 嵌蛋 入白 )和 蛋膜 白内
糖蛋白
去污剂
外周蛋白与膜的连接是可逆的
• 许多外周蛋白通过与嵌入蛋白的亲水区 域或膜脂的极性头部以静电作用或氢键 结合到膜上,通过温度的改变或破坏静 电或破坏氢键作用(如加入螯合剂、尿 素、碳酸盐或改变pH)可被释放出来。 • 这些外周蛋白可作为膜结合酶的调节因 子、或作为连接膜内蛋白与胞间结构的 中介物、或一些膜蛋白的流动性。
O-连接四糖: 2Neu5Ac, Gal,GalNAc
-螺旋
糖红 蛋细 白胞 的上 跨血 膜型
冰冻蚀刻(Freeze-fracture Technique)撕开膜双层
膜内(嵌入)蛋白不溶于水
膜蛋白可分为两类,外周蛋白 ( peripheral ) 和 膜 内 ( 嵌 入 ) 蛋 白 (integral),前者与膜结合松散,可逆, 很容易释放,是水溶性的;后者与膜结 合紧密,由膜上释放时要用特别的试剂 (去污剂、有机溶剂、变性剂等),及 时嵌入蛋白由膜上释放出来,一旦去除 变性剂或去污剂会立即引起蛋白沉淀 (不溶聚积物)析出。