新编文档-真核生物基因结构-精品文档
真核基因的结构

基底连接区 边界元件 启动子 起始密码子 内含子 终止密码子 聚合信号
增强子 基底连接区
5‘-非编码 外显子
外显子
3‘-非编码 外显子
边/部分 外显子/部分 基因/部分 选择性剪接 无内含子基因 外显子数目 外显子长度 内含子长度 基因长度 供体位点 受体位点 53435/29404 83488/21342 20791/16141 2167,10.4% 1551,7.4% 117, 5.7 1-1088,201.6 259776,2203.5 401910, 9033 58707, 98.0% 58112, 98.53%
人源真核生物基因
• • • • • •
人类基因编码的外显子比各自基因的长度要短 外显子平均长度200bp 编码和非编码外显子长度可以是几个到千个碱基 编码蛋白的外显子占整个基因的百分之几 各种重复子占人类DNA的41% 外显子占基因组序列的2-3%
计算识别的问题
• 低质量的识别会产生大量的假阳性预测 • 任何基于基因组成的识别方法通常能成 功地用于识别原核生物的基因,无法识 别真核基因的中短外显子
需要发展基于基因序列中编码功能信号的 基因预测方法
识别功能信号的方法
• • • • • • • 搜索保守序列的非随机的相似性 位点特异性识别器 内容特异性测定 基于框架特异性的蛋白编码区的识别 供体受体拼接位点的预测 启动子序列的识别 Poly(A)位点的预测
多基因预测的差异分析
• • • • 使用HMM 基于模式的多基因预测 基因识别的准确性 利用蛋白质或EST相似性信息来改进基因 预测
序列注释
• 注释可视化 • InfoGene: 已知基因和预测基因的数据库
调控区的主要特征
真核基因组结构

15.3 Composite transposons have IS modules
Figure 15.2 A composite transposon has a central region carrying markers (such as drug resistance) flanked by IS modules. The modules have short inverted terminal repeats. If the modules themselves are in inverted orientation (as drawn), the short inverted terminal repeats at the ends of the transposon are identical.
微卫星DNA:总长度小于150bp,分布在所 有染色体,重复单位为1-5bp,重复次数1060次,常见的是 (AC)n (TG)n
4.9 Satellite DNAs often lie in heterochromatin Figure 4.14 Mouse DNA is separated into a main band and a satellite by centrifugation through a density gradient of CsCl.
4.9 Satellite DNAs often lie in heterochromatin
Figure 4.15 Cytological hybridization shows that mouse satellite DNA is located at the centromeres. Photograph kindly provided by Mary Lou Pardue and Joe Gall.
真核生物基因组结构

真核生物DNA 复性曲线的模 式图
复性反应分为 三P相74,图每-相16代 表不同复杂长 度的序列类型
大部分结构基因 位于非重复的 DNA序列内
第二节 断裂基因(split gene)
不连续基因(interrupted gene)
编码某一RNA的基因中有些序列并不出现在成熟 的RNA序列中,成熟RNA的序列在基因中被其他的 序列隔开。
因家族, 一般往往以基因家族的形式存在。
Alu family(Alu 家族):
•长约300bp的片段,大多数片段含有一个限制性内切 酶AluⅠ的酶切位点(AGCT); • 均匀分散在整个基因组中的非重复序列间; • 在人类基在103mol.s/L以上,复性速度极慢, 在一个基因组中只有一个拷贝或2~3个拷贝,也 称非重复序列(单一序列、单拷贝序列)。 结构基因 (蛋白质基因)大多是单拷贝序列。
人类珠蛋白基因家族---典型的基因家族
珠蛋白 血红蛋白
血红素
α2β2 不同的亚基由各自的基因编 码
血红蛋白( Hb )
发育过程中的珠蛋白的亚基组成
类α链
α2γ2
2% 97% 1%
类β链
两种亚基的编码基因分别形成两个不同的基因簇, 并存在于不同的染色体上。
每个基因簇中的基因按其在发育过程中的表达次序从 5’→3’排列在编码链上(其中包括有功能的基因和假基 因)
1)零时复性序列:
具有反向重复结构(也称回文结构),可在 同一条链内形成双链区,变性后再复性时,在链 间复性之前就已发生链内复性,因此不遵循二级 反应动力学方程。由于这种序列的复性速度非常 快,在动力学上称为零时(或瞬时)复性序列。
DNA复性后可出现发卡形结构。这种序列常 常是DNA复制酶、转录酶以及特异蛋白质的结合 部位。
简述真核生物基因的结构特点

简述真核生物基因的结构特点真核生物基因的结构特点包括以下几个方面:1. 真核生物基因位于染色体上,是真核生物细胞中的核心结构。
染色体是由 DNA 和蛋白质组成的复合物,是在细胞分裂时传递遗传信息的基本单位。
真核生物基因组的 DNA 与蛋白质结合形成染色体,储存于细胞核内。
除配子细胞外,体细胞内的基因组是双份的 (即双倍体,diploid),即有两份同源的基因组。
2. 真核细胞基因转录产物为单顺反子 (monocistron),即一个结构基因转录、翻译成一个 mRNA 分子,一条多肽链。
真核生物的基因转录是在 DNA 模板上以 RNA 为模板进行转录,产生的 mRNA 是单链,在细胞质中由核糖体(ribosome) 进行翻译。
3. 真核生物基因组中存在大量重复序列,包括高度重复序列和中度重复序列。
高度重复序列重复频率可达 106 次,包括卫星 DNA、反向重复序列和较复杂的重复单位组成的重复序列;中度重复序列可达 103~104 次,如为数众多的Alu 家族序列,KpnI 家族,Hinf 家族序列,以及一些编码区序列如 rRNA 基因、tRNA 基因、组蛋白基因等。
4. 真核生物基因是不连续的,在真核生物结构基因的内部存在许多不编码蛋白质的间隔序列 (interveningsequences),称为内含子 (intron),编码区则称为外显子 (exon)。
内含子与外显子相间排列,转录时一起被转录下来,然后RNA 中的内含子被切掉,外显子连接在一起成为成熟的 mRNA,作为指导蛋白质合成的模板。
5. 真核生物基因组远大于原核生物的基因组,具有许多复制起点,而每个复制子的长度较小。
真核生物基因组 DNA 与蛋白质结合形成染色体,储存于细胞核内。
除配子细胞外,体细胞内的基因组是双份的 (即双倍体,diploid),即有两份同源的基因组。
真核基因的一般结构构造

真核基因的一般结构构造
真核基因的一般结构构造
真核基因是指eukaryotic细胞和细菌的基因组中的基本基因,它们是
为了创造具有特定功能的多种类型的RNA而编码的。
真核基因的一般
结构构造是由一段特定序列DNA碱基组成,它们可以被分为三个部分:起始密码子、非编码区和终止密码子。
首先,起始密码子是真核基因的一个重要部分,它定义了真核基因的
起始位置,当mRNA在这里被转录时,激活该基因的生物学功能。
这部
分的密码子被称为ATG,即开始转录的第一个反式密码子,它常常位于基因片段的开始位置,ATG中含有三个碱基:Adenine(A)、Thymine(T)和Guanine(G)。
其次,非编码区是真核基因中最长的部分,也是起码90%的真核基因序列组成。
它不参与蛋白质合成,但是起一定的调节作用,如决定基因
表达的频率和程序等。
这部分的基因序列包括各种称为基因控制元件
的元素,如原子、转录因子结合位点、启动子序列和增强子序列等。
最后,终止密码子,又被称为终止码,它是真核基因转录结束的终止点。
它通常位于编码区域的末尾,它会提前结束mRNA的翻译,然后将mRNA从细胞中分泌出去。
该部分的密码子包括三个碱基:
Cytosine(C)、Adenine(A)和Uracil(U) 。
总之,真核基因的一般结构构造由三个部分组成:起始密码子、非编
码区和终止密码子,它们是真核基因的核心结构,在这三部分的完整
性和正确性的基础上,才能确保真核基因的功能正常发挥。
《真核生物基因结构》PPT课件

保留
遗传病
3
二)Gene类别
依其功能分为: 1、结构Gene---编码蛋白和酶分子结构; 2、调控Gene---调节结构基因表达; 3、转录而不翻译的Gene:
rDNA Gene→rRNA→核仁形成区, 核糖体组成。
tRNA Gene→tRNA→转运氨基酸。
4
三)原核与真核生物基因比较
原核生物Gene:无核膜,散在细胞质。 一般只有一个染色体,即一个DNA或RNA分子,Gene 是连续的。 大多数是双链环状,少数为单链、线状; 如:大肠杆菌,双链环状DNA分子,3000~4000个 基因,4.2x106 bp,已经定位900多个基因
它是基因的调控序列,对基因的有效表达 起调控作用,包括:启动子、增强子、终止 子等。
1 0
2.1 启动子
启动子( Promoter)是一段特定的核苷酸序列,位于Gene 转录起始点上游的100bp 范围内,是RNA聚合酶的结合部位, 能促进转录过程。Promoter决定DNA中的转录链。
TATA框(TATA Box)是一段高度保守序列,7个bp,TATAA/ TAA/T,位于转录起始点上游25~30 bp(-30~50)。TATA 框与转录因子TFII结合,再与RNA 聚合酶II形成复合物,从而 准确地识别转录起始位置,对转录水平有定量效应。
真核生物基因结构
(Suitable for teaching courseware and reports)
2
一)基因的一般特性从分子水源自看,基因有以下基本特性:1、Gene自我复制------半保留复制
2、基因决定性状:
Protein
Gene→mRNA →
enzyme
淘汰
3、Gene突变
真核基因结构

真核基因结构
嘿,咱今天就来聊聊真核基因结构哈。
你说这真核基因结构啊,就像是一个超级复杂的大拼图。
这拼图里有好多好多的小零件呢!首先呢,有个叫启动子的家伙,它就像是比赛开始的发令枪,“砰”的一声,基因表达就开始啦!然后呢,还有编码区,这可是关键部分哟,就像拼图的主体图案,决定了最后会呈现出个啥样子。
再说说那些调控元件吧,它们就像一群小精灵,这儿跳跳那儿蹦蹦,指挥着基因该怎么表达,什么时候表达。
还有那些内含子,就像是拼图里的一些隐藏关卡,有时候会让你摸不着头脑,但又不能小瞧它们。
这真核基因结构啊,真的是特别神奇。
就好像是大自然这位超级厉害的设计师精心打造出来的杰作。
它决定了我们生物的各种特征和功能,从我们的长相到我们的性格,都和它有着密切的关系呢。
想象一下,如果真核基因结构是一场游戏,那这游戏可太难玩啦!那么多的规则,那么多的细节,稍不注意就会出错。
不过呢,好在大自然已经玩得很溜啦,把我们都创造得好好的。
咱平常可能不会特别去注意这个神奇的真核基因结构,但它其实一直在默默地工作着,就像一个勤劳的小蜜蜂,为我们的生命活动辛勤付出。
哎呀呀,说了这么多,其实就是想让你知道,真核基因结构虽然听起来很复杂很专业,但其实它就在我们身边,影响着我们的点点滴滴。
它就像是生命的密码,等待着我们去慢慢探索和理解。
好啦,关于真核基因结构咱就先聊到这儿咯,下次有机会再一起探讨其他有趣的生命奥秘呀!嘿嘿。
第五章真核生物基因组结构

染色质(chromatin):是指细胞周期间期细胞核内由 DNA、组蛋白、非组蛋白和少量RNA组成的复合结构, 因其易被碱性染料染色而得名。
染色体(chromosome):是指在细胞分裂期出现的 一种能被碱性染料强烈染色,并具有一定形态、结构特 征的物体。 携带很多基因的分离单位。只有在细胞分 裂中才可见的形态单位。
具体表现: 与预期的编码蛋白质的基因数量相比,基因组DN
A的含量过多,为什么? 一些物种之间的复杂性变化范围并不大,但C值却
有很大的变化范围。对于不同物种,C值差异较大。 C值的大小与物种的结构组成和功能的复杂性没有严 格的对应关系,这种现象称为C值矛盾。
08:50
12
四 基因的数量
差别很大
在原核生物中,转录的调节区都很小,大都位于启 动子上游不远处,调控蛋白结合到调节位点上可直接 促进或抑制RNA聚合酶与它的结合。
08:50
8
⑥ 真核生物的RNA在细胞核中合成,只有经转运穿过核 膜,到达细胞质后,才能被翻译成蛋白质,原核生物 中不存在这样严格的空间间隔。
⑦ 许多真核生物的基因只有经过复杂的成熟和剪接过 程,才能顺利地翻译成蛋白质。
卫星DNA 小卫星DNA:由中等大小的串联重复序列组成, 位于靠近染色体末端的区域。
微卫星DNA:由更简单的重复单位组成的小序 列,分散于基因组中。
08:50
36
第四节 真核基因组的包装
包装比(packing ratio):
用以描述DNA的浓缩程度,即DNA的原始长 度除以包装后的长度。 如人类最小的染色体(Y):DNA拉直约为1.4 cm ,在有丝分裂最致密状态下,这条染色体 长度约为2μm,因此包装比可以大到7000。
真核生物的基因结构
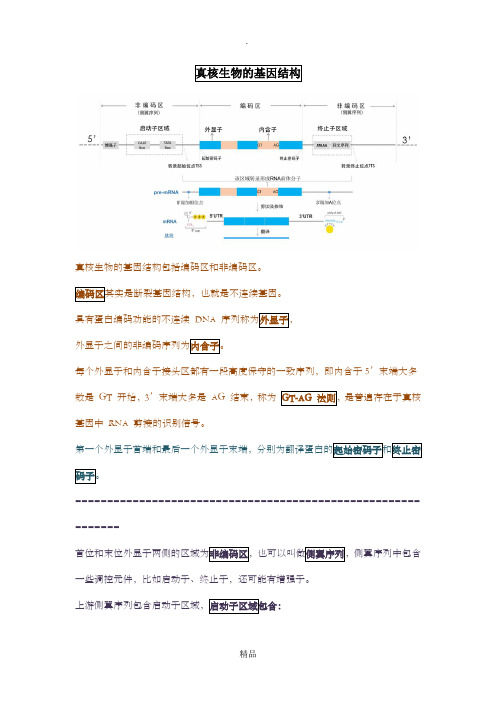
真核生物的基因结构
真核生物的基因结构包括编码区和非编码区。
编码区其实是断裂基因结构,也就是不连续基因。
具有蛋白编码功能的不连续DNA 序列称为外显子,
外显子之间的非编码序列为内含子。
每个外显子和内含子接头区都有一段高度保守的一致序列,即内含子5’末端大多数是GT 开始,3’末端大多是AG 结束,称为GT-AG 法则,是普遍存在于真核基因中RNA 剪接的识别信号。
第一个外显子首端和最后一个外显子末端,分别为翻译蛋白的起始密码子和终止密码子。
====================================================== =======
首位和末位外显子两侧的区域为非编码区,也可以叫做侧翼序列,侧翼序列中包含一些调控元件,比如启动子、终止子,还可能有增强子。
上游侧翼序列包含启动子区域,启动子区域包含:
列为TATAATAAT,是RNA聚合酶的重要的接触点,它能够使酶准确地识别转录
止密码子之间区域,不编码蛋白质。
miRNA 经常结合于3‘UTR,从而引起mRNA。
第二节真核生物基因结构及功能

ቤተ መጻሕፍቲ ባይዱ
2.4
调控序列
调控序列(Regulator Sequence)包括 启动子,增强子和终止子均属于基因的 顺式调控因子(顺式作用元件),是人 类Gene组中的一些特殊序列,起调控基 因表达的作用。 反式作用元件:TFII、CTF、SP1。
14
11
2.2
增强子
增强子(Enhancer)包括启动子上游或下游的一 段DNA序列,可以增强启动子发动转录,提高转录效率。 特点: 在任意位置都有效 无方向性 有组织特异性 例如:Beta珠蛋白Gene增强子是由串联重复的两 个72bp长的相同序列组成,位于转录起点上游-1400bp 或下游3300bp处,均可增强转录效率(活性)200倍。 增强子在转录起始点的上下游一定范围内增强转录效率。 作用可以是5’~3’,也可以是3’~5’方向。
9
2.1 启动子
启动子( Promoter)是一段特定的核苷酸序列,位于Gene 转录起始点上游的100bp 范围内,是RNA聚合酶的结合部位, 能促进转录过程。Promoter决定DNA中的转录链。 TATA框(TATA Box)是一段高度保守序列,7个bp, TATAA/ TAA/T,位于转录起始点上游25~30 bp(-30~50)。 TATA框与转录因子TFII结合,再与RNA 聚合酶II形成复合物, 从而准确地识别转录起始位置,对转录水平有定量效应。
4
二、真核生物基因结构
真核生物的结构基因的DNA序列由编码序列和非编 码序列两部分组成,编码序列是不连续的,被非编码序 列分割开来,称为断裂基因(split gene)。
1、外显子和内含子 2、侧翼序列与调控序列 2.1 启动子 2.2 增强子 2.3 终止子
真核细胞的基因结构

真核细胞的基因结构在遗传学上通常将能编码蛋白质的基因称为结构基因。
真核生物的结构基因是断裂的基因。
一个断裂基因能够含有若干段编码序列,这些可以编码的序列称为外显子。
在两个外显子之间被一段不编码的间隔序列隔开,这些间隔序列称为内含子。
每个断裂基因在第一个和最后一个外显子的外侧各有一段非编码区,有人称其为侧翼序列。
在侧翼序列上有一系列调控序列(图1)。
调控序列主要有以下几种:①在5′端转录起始点上游约20~30个核苷酸的地方,有TATA框(TATA box)。
TA TA框是一个短的核苷酸序列,其碱基顺序为TATAA TAAT。
TA TA 框是启动子中的一个顺序,它是RNA聚合酶的重要的接触点,它能够使酶准确地识别转录的起始点并开始转录。
当TA TA框中的碱基顺序有所改变时,mRNA的转录就会从不正常的位置开始。
②在5′端转录起始点上游约70~80个核苷酸的地方,有CAAT框(CAAT box)。
CAAT框是启动子中另一个短的核苷酸序列,其碱基顺序为GGCTCAATCT。
CAAT框是RNA聚合酶的另一个结合点,它的作用还不很肯定,但一般认为它控制着转录的起始频率,而不影响转录的起始点。
当这段顺序被改变后,mRNA的形成量会明显减少。
③在5′端转录起始点上游约100个核苷酸以远的位置,有些顺序可以起到增强转录活性的作用,它能使转录活性增强上百倍,因此被称为增强子。
当这些顺序不存在时,可大大降低转录水平。
研究表明,增强子通常有组织特异性,这是因为不同细胞核有不同的特异因子与增强子结合,从而对不同组织、器官的基因表达有不同的调控作用。
例如,人类胰岛素基因5′末端上游约250个核苷酸处有一组织特异性增强子,在胰岛素β细胞中有一种特异性蛋白因子,可以作用于这个区域以增强胰岛素基因的转录。
在其他组织细胞中没有这种蛋白因子,所以也就没有此作用。
这就是为什么胰岛素基因只有在胰岛素β细胞中才能很好表达的重要原因。
④在3′端终止密码的下游有一个核苷酸顺序为AATAAA,这一顺序可能对mRNA的加尾(mRNA尾部添加多聚A)有重要作用。
真核生物结构基因

真核生物结构基因真核生物是指细胞核内含有真正的染色体,其中包含着基因,这些基因是决定生物遗传性状的基本单位。
基因是由DNA序列编码的,它们携带着生命信息。
基因的结构和组成对于生命的运作和表达起着至关重要的作用。
基因的结构基因通常由三个部分组成:启动子、编码区和终止子。
启动子位于基因的起始位置,是一段DNA序列,它可以激活基因的转录,从而将基因转录成mRNA。
编码区位于启动子的下游区域,它是一个由多个外显子和内含子组成的序列,编码区的序列决定了基因所编码的蛋白质的氨基酸序列。
终止子位于编码区的下游,它是基因转录终止的信号,从而完成mRNA的合成。
基因的组成基因由DNA序列编码,DNA序列是由四种不同的核苷酸组成的碱基序列,即腺嘌呤(A)、胸腺嘧啶(T)、鸟嘌呤(G)和胞嘧啶(C)。
这四种碱基按照一定的顺序组成了DNA序列,而DNA序列又组成了基因序列。
基因序列是一段由多个碱基组成的DNA序列,这些碱基按照一定的顺序编码了蛋白质的氨基酸序列,从而决定了蛋白质的形状和功能。
基因的表达基因的表达是指基因信息从DNA转化为蛋白质的过程。
基因的表达受到多种因素的调控,其中包括启动子、转录因子和表观遗传学等。
启动子可以激活基因的转录,从而将基因转录成mRNA。
转录因子是一种特殊的蛋白质,它能够结合到启动子上,从而调控基因的转录。
表观遗传学是指与DNA序列无关的遗传学变化,如DNA 甲基化和组蛋白修饰等,这些变化会影响基因的表达。
总结基因作为生命的基本单位,决定了生物的遗传性状。
基因的结构和组成对于生命的运作和表达起着至关重要的作用。
基因由DNA序列编码,DNA序列是由四种不同的核苷酸组成的碱基序列组成的,而DNA序列又组成了基因序列。
基因的表达是指基因信息从DNA 转化为蛋白质的过程,受到多种因素的调控。
深入理解基因的结构和组成以及基因的表达,对于生命科学的研究和应用具有重要意义。
真核生物结构基因

真核生物结构基因真核生物是指细胞内有核和其他细胞器的生物,包括动物、植物、真菌和原生动植物等。
结构基因是指编码蛋白质的基因。
在真核生物中,结构基因是生命活动的重要组成部分,控制着细胞的生长和分化等重要生物过程。
真核生物的细胞结构真核生物的细胞结构主要包括细胞核、线粒体、内质网、高尔基体、溶酶体等。
其中,细胞核是真核细胞的重要特征,包含了细胞的遗传信息和控制蛋白质合成的核糖核酸。
线粒体是细胞内的能量中心,参与了细胞的呼吸作用。
内质网则参与了蛋白质的合成和运输等活动。
高尔基体则负责将蛋白质及其他分子送达其它细胞器或细胞外。
溶酶体则起到了消化细胞外物质和细胞内垃圾的作用。
真核生物的基因结构真核生物的基因结构主要包括启动子、外显子和内含子等。
启动子是基因的起始点,其中包含有调控基因转录的序列。
外显子则编码了蛋白质所需要的信息。
内含子则是指在基因转录过程中,需要被剪切掉的无用序列。
另外,真核生物的基因还包括了转录因子和RNA剪接因子等,这些因子参与了基因的表达和调控过程。
真核生物的基因表达调控真核生物的基因表达调控非常复杂,包括转录起始、RNA加工、转运和翻译等多个环节。
其中,转录因子的作用非常重要,它们可以结合到启动子上,激活或抑制基因的转录过程。
此外,RNA剪接也是基因表达的重要调控机制,它可以选择性地剪切内含子,从而调节外显子的组合方式,影响蛋白质的翻译过程。
真核生物的基因编辑技术近年来,CRISPR/Cas9等基因编辑技术的出现,使得真核生物的基因编辑变得更加方便和高效。
通过这些技术,可以对真核生物的基因进行精准的编辑和修改,从而实现对基因表达和调控的精确控制。
这些技术的出现为真核生物研究提供了新的思路和手段,也为人类疾病的治疗和基因工程的发展提供了新的可能性。
真核生物结构基因是生命活动的重要组成部分,对于真核生物的生长、分化和遗传等方面都有着重要的作用。
随着基因编辑技术的不断发展,相信我们对于真核生物结构基因的了解和应用将会更加深入和广泛。
真核基因的基本结构

真核基因的基本结构
真核基因的基本结构通常由以下几个部分组成:
1. 启动子:启动子是基因的一个序列,可以与RNA聚合酶结合,启动转录过程。
启动子通常位于基因的5'端。
2. 编码区:编码区是基因的编码序列,包括外显子和内含子。
外显子是编码蛋白质的序列,内含子则是位于编码区内的非编码序列。
3. 终止子:终止子是基因的一个序列,可以与转录因子结合,终止转录过程。
终止子通常位于基因的3'端。
4. 调控序列:调控序列是基因内的一些序列,可以与转录因子结合,调控基因的转录活性。
常见的调控序列包括增强子和反应元件等。
真核基因的编码区通常由多个外显子和内含子组成,外显子和内含子交替排列。
在转录过程中,内含子会被剪切掉,只保留外显子部分的序列,然后经过拼接形成成熟的mRNA。
这个过程被称为RNA剪接。
真核基因的结构是复杂的,其表达受到多个层次的调控。
通过对真核基因结构的分析,可以深入了解基因的表达调控机制,为研究基因功能和疾病发生提供重要信息。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
4
二、真核生物基因结构
真核生物的结构基因的DNA序列由编码序列和非编 码序列两部分组成,编码序列是不连续的,被非编码序 列分割开来,称为断裂基因(split gene)。
1
一)基因的一般特性
从分子水平看,基因有以下基本特性:
1、Gene自我复制------半保留复制
2、基因决定性状:
Protein
Gene→mRNA →
enzyme
淘汰
3、Gene突变
生物进化
保留
遗传病
2
二)Gene类别
依其功能分为: 1、结构Gene---编码蛋白和酶分子结构; 2、调控Gene---调节结构基因表达; 3、转录而不翻译的Gene:
11
2.2 增强子
增强子(Enhancer)包括启动子上游或下游的一 段DNA序列,可以增强启动子发动转录,提高转录效率。 特点: 在任意位置都有效 无方向性 有组织特异性
例如:Beta珠蛋白Gene增强子是由串联重复的两 个72bp长的相同序列组成,位于转录起点上游-1400bp 或下游3300bp处,均可增强转录效率(活性)200倍。 增强子在转录起始点的上下游一定范围内增强转录效率。 作用可以是5’~3’,也可以是3’~5’方向。
7
5`GT——AG3`法则
在每个外显子和内含子的接头区都是一段 高度保守的共有序列,内含子的5`端是GT,3 端是AG,这种接头方式称为GT-AG法则,普 遍存在于真核生物中,是RNA剪接的识别信号, 转录后的前体RAN中的内含子剪接位点。
8
2、侧翼序列与调控序列
每个结构基因的第一个和最后一个外显 子的外侧,都有一段不被转录的非编码区, 称为侧翼序列(Flanking sequence)。
1、外显子和内含子
2、侧翼序列与调控序列
2.1 启动子
2.2 增强子
2.3 终止子
5
1、外显子和内含子
在结构基因中,编码序列称为外显子(exon),表达多 肽链部分。非编码序列称为内含子(Intron),又称插入 序列。
β珠蛋白 基因(1700bp)=3个外显子+2个内含子。 DMD基因(2300kb)=79个外显子+ 78 个内含子。
rDNA Gene→rRNA→核仁形成区, 核糖体组成。
tRNA Geneห้องสมุดไป่ตู้tRNA→转运氨基酸。
3
三)原核与真核生物基因比较
原核生物Gene:无核膜,散在细胞质。 一般只有一个染色体,即一个DNA或RNA分子,Gene 是连续的。 大多数是双链环状,少数为单链、线状; 如:大肠杆菌,双链环状DNA分子,3000~4000个 基因,4.2x106 bp,已经定位900多个基因
它是基因的调控序列,对基因的有效表达 起调控作用,包括:启动子、增强子、终止 子等。
9
2.1 启动子
启动子( Promoter)是一段特定的核苷酸序列,位于Gene 转录起始点上游的100bp 范围内,是RNA聚合酶的结合部位, 能促进转录过程。Promoter决定DNA中的转录链。 TATA框(TATA Box)是一段高度保守序列,7个bp,TATAA/ TAA/T,位于转录起始点上游25~30 bp(-30~50)。TATA 框与转录因子TFII结合,再与RNA 聚合酶II形成复合物,从而 准确地识别转录起始位置,对转录水平有定量效应。
13
2.4 调控序列
调控序列(Regulator Sequence)包括 启动子,增强子和终止子均属于基因的 顺式调控因子(顺式作用元件),是人 类Gene组中的一些特殊序列,起调控基 因表达的作用。
反式作用元件:TFII、CTF、SP1。
14
12
2.3 终止子
终止子(Terminator)由一段 回文序列以及特定的序列 (PolyA)5’--AATAAA--3’组成。
回文序列为转录终止号。PolyA 为附加信号。终止子为反向重复 序列,是RNA聚合酶停止工作 的信号,反向重复序列转录后, 可以形成发夹式结构,并且形成 一串U。发夹式结构阻碍了RNA 聚合酶的移动。一串U的U与 DNA模板中的A的结合不稳定, 从模板上脱落下来,终止转录。
10
CAAT框(CAAT Box):是一段保守序列, 9bp,GGGC/TCAATAC, 位于转录起始点上游70~-80bp,转录因子CTF识别位点并与之结合, 激活转录。
GC框(GC Box):顺序为GGCGGG,有两个 拷贝,位于CAAT Box两侧,与转录因子SP1 结合。(SP1有锌指区可以与DNA结合,在N 端有激活转录的作用)GC框有激活转录的功 能。
(迄今认识的最大的基因)
6
外显子和内含子
真核生物内含子和外显子 不是完全固定不变的, 有时同一DNA 链上的某一段DNA序列,当它 作为编码某一多肽链的基因时是外显子,而作 为编码另一多肽链时,则是内含子。这样,同 一基因却可以转录两种或两种以上的mRNA。
真核生物某些结构Gene没有内含子,如组蛋 白Gene,干扰素Gene等。它们多以基因簇形 式存在,大多数的酵母结构Gene也没有内含 子。
第二节、真核生物基因结构及功能
一、基因的概念
基因的概念随着分子遗传学、分子生物学、 生物化学领域的进展而不断完善。 从遗传学角度看:
基因是生物的遗传物质,是遗传的基本单 位----突变单位、重组单位和功能单位。 从分子生物学角度看:
基因是负载着特定遗传信息的DNA分子片段, 在一定条件下能够表达这种遗传信息,执行特 定的生理功能。