昆虫飞行肌蛋白质3
昆虫的飞行能力实验

昆虫的飞行能力实验昆虫的飞行能力一直是科学家们探索和研究的领域之一。
尽管昆虫的身体相对微小,但它们却能够以令人难以置信的速度和灵活度飞行。
为了更好地理解昆虫的飞行机制以及为人类创造更高效的飞行器,科学家们开展了一系列的实验。
为了研究昆虫飞行的动力学,科学家们首先需要分析昆虫翅膀的结构。
研究发现,昆虫的翅膀由许多薄而坚韧的膜片构成,这些膜片与昆虫的肌肉相互配合,通过频繁而协调的振动产生飞行。
为了揭示昆虫翅膀振动的详细情况,科学家们设计了一种高速摄像技术来捕捉昆虫飞行过程中的细节。
在实验中,科学家们首先选择了一种常见的飞行昆虫,如苍蝇或蜜蜂。
他们使用准确的测量工具来获取昆虫的尺寸、重量和翅膀形状等数据。
然后,科学家们将昆虫放置在一个透明的容器内,容器内可以控制温度、湿度和氧气浓度等环境因素。
为了捕捉昆虫的飞行过程,科学家们将高速摄像机设置在适当的角度和位置上。
他们将摄像机调整至合适的帧速率,并使用适当的照明设备来确保拍摄的图像清晰可见。
这样,科学家们就可以记录下昆虫在飞行过程中翅膀的振动方式和速度等相关信息。
实验过程中,科学家们发现昆虫的翅膀振动呈现出复杂而精确的模式。
不同种类的昆虫振动频率和幅度各异,这也解释了为什么昆虫可以够以不同的速度和灵活度飞行。
通过对昆虫飞行过程中翅膀振动的分析,科学家们可以更好地理解昆虫飞行的机制并将其应用到人类飞行器的设计中。
此外,科学家们还研究了昆虫的飞行稳定性和控制能力。
他们使用精密的测量仪器来记录昆虫在不同姿态下飞行时的姿态角度和灵活度。
通过观察昆虫的飞行行为,科学家们发现昆虫具备精确的调节能力,可以根据环境条件的变化来保持稳定的飞行状态。
总体来说,昆虫的飞行能力实验为科学家们揭示了昆虫飞行的机制以及飞行稳定性等关键参数提供了重要的数据和信息。
这些实验不仅有助于更好地理解昆虫的生物特性,还为人类创造更高效的飞行器提供了重要的参考和启示。
随着技术的不断进步,相信在未来的实验中,我们还将揭开更多关于昆虫飞行的奥秘。
昆虫生理生化

昆虫生理生化《昆虫生理生化》第一章体壁引言昆虫体壁(integument)又称外骨骼,它具有高等动物皮肤和骨骼的双重功能,使虫体具有坚强的支撑,并为肌肉提供了着生点,但在体壁的某些区域,仍然有柔软的性质,关键部位有许多关节,保证虫体能行动自如。
体壁也是一个复杂的代谢库,这些过程都受到激素的调节和控制。
体壁又是虫体与环境之间的界面,是抵御外界异物和阻止杀虫剂渗透的屏障,特别对保持水分有很大的作用。
第一节体壁的组成昆虫体壁包括皮细胞、表皮和基底膜三部分,表皮是皮细胞分泌的产物,但基底膜则是由血细胞分泌的。
一皮细胞1.昆虫的皮细胞主要分布在虫体外围,局部区域已在胚胎发育时随着外胚层的内陷成为前肠、后肠、气管或生殖道的管壁细胞;另一部分特化成腺细胞、毛原细胞和膜原细胞。
皮细胞的生理特点是具有周期性吸收、合成和分泌能力。
2.在极大多数昆虫中,皮细胞都是以单层形式排列的,细胞的纵切面大多呈柱状,也有不规则的。
3.皮细胞的大小与密度因虫种和发育阶段不同而异,并且与生长期间是否进行有丝分裂有关。
(一) 形态结构1.昆虫皮细胞的形态结构是随变态和脱皮周期而不断变化的,皮细胞在脱皮期间分泌作用较弱,顶膜与底膜平直,侧膜不明显,细胞核不清晰。
2.沉积新表皮开始时期顶膜弯曲,常有原生质丝突入表皮层内,侧膜出现,胞核明显。
3.在溶离旧表皮与沉积新表皮期间,细胞的合成、分泌和吸收都很旺盛,顶膜微绒毛发达,基膜褶深陷,底膜也常随基膜一起内陷,并有血淋巴进入到内褶中,增加了基膜与血淋巴接触机会和吸收面积。
(二) 胞间联系1.皮细胞能够识别自身所处的位置,并与周围细胞进行联系和协调,按程序分泌和沉积特定的表皮。
2.皮细胞之间复杂的横向通讯联络,主要通过侧膜联接来实现。
大量平行排列的皮细胞间,在侧膜的上半部形成隔壁联结与间隙联结,皮细胞之间也借此进行信息交流,甚至结成合胞体。
3.皮细胞之间有离子偶联和由浓度梯度产生的信息交流,还能调节细胞生长和影响形态发生。
第二十五章昆虫的肌肉及其生理

缩机理。他认为肌肉收缩是由于粗肌丝和细肌丝相对滑动造成的。
移动,另一端伸向A带的H区,呈游离状态。肌肉收缩时细肌丝 的游离端沿粗肌丝的长轴向H区滑行,两组细肌丝在滑行中互相 靠拢,重叠,使肌节变短变粗,而肌丝本身的长度不发生改变。 该学说获得广泛公认
四、兴奋—收缩偶联
•
粗肌丝的肌球蛋白分子的头部具有ATP酶活性, 在细肌丝的原肌球蛋白上结合有ATP,当肌球蛋白的 横桥与肌动蛋白结合形成肌动球蛋白时,便放出ATP 酶,水解ATP,放出能量。
• 体壁肌着生在体壁下或体壁的内突上,按体节着生。 • 功能:
• 司体节、附肢和翅的运动。
• 肌肉的基本单位是肌纤维 • 肌纤维的基本单位是肌原纤维——收缩单元。 • 按肌原纤维在肌纤维中的排列状况,将体壁肌分为三种类型:
束状肌
•
•
管状肌
纤维状肌
(一) 束状肌
• 肌纤维外包着1层不含肌原纤维的肌浆,根据
• 肌原纤维由数百条粗纤丝和细纤丝所组成。
ቤተ መጻሕፍቲ ባይዱ肉的组织结构
• (一)肌纤维muscle fiber
是一类长形的多核细胞, 肌纤维包有一层肌膜,
肌膜内含有肌原纤维, 含糖元和脂肪的肌质充塞于 肌原纤维之间 肌膜垂直内陷分化成许多横向小管, 构成肌肉 的横管系统(T系统),能通过电扩散传导神经脉冲。
在肌原纤维之间, 有许多纵行的小管网络, 构成
三、肌细胞核
•
•
• 肌细胞核位于肌膜内,1个肌纤维通常有多个细胞核。以控 制早期肌细胞的分裂、分化和肌细胞的更新和代谢。
四、肌原纤维
肌原纤维是肌纤维中特有的功能性细胞器,是肌细胞的基本 功能单位。肌原纤维在肌细胞内呈细丝纤维状,在光学显微镜下
昆虫生理学第十章昆虫的运动系统

肌 球 蛋 白 和 肌 动 蛋 白
3)原肌球蛋白
一种纤维状蛋白,以双螺旋镶嵌在肌动蛋白缠 绕的凹槽内。
功能:当它覆盖肌球蛋白头部的活性部位 时,能阻止肌动球蛋白横桥的形成。
4)肌钙蛋白
一种钙结合蛋白,是肌肉收缩的一种重要的蛋白。
5)副肌球蛋白
一种氨基酸含量很高的蛋白,与肌肉长时间 紧张性收缩有关,在粗肌丝中形成一个核。功能 不详。
第六节 杀虫剂对肌肉系统的影响
肌肉毒剂
邻苯二甲酰胺类 邻甲酰胺基苯甲酰胺类
美洲中部和南部的植被群譬如尼亚那灌木,尤其是一种 广泛生长于该地区的灌木Ryania speciosa Vahl (大风子科灌 木),在上世纪中叶曾经受到默克公司科学家的关注。
1948年Rogers等从ryania中找到一种活性组分yanodine。 但ryania的杀虫作用效果并不是很有效,而且在今天看来也 不是很安全。实际上,它对哺乳动物就象以前的一支毒箭一 样具有致命效果。兰尼碱的结构复杂,非常难以进行化学合 成。
三、肌纤维上的神经分布
昆虫肌肉上的神经多数呈多点式分布,即每条肌纤维都 与运动神经末梢形成多个突触联结,运动神经末梢与肌肉的 连接点又称运动终板(terminal lamella)。但也有其他分布 形式。
昆虫的运动神经元分为兴奋性和抑制性两类,兴奋性神 经又分为快神经、慢神经及一些中间类型。
肌肉的神经分布
横桥末端产生摆动, 拉动细肌丝沿粗肌丝移动,
在高浓度Ca2+ 继续收缩
收缩结束 Ca2+主动吸回
肌质网
肌钙蛋白钙结合
其信息通过肌动蛋白
释放出
亚基与Ca2+结合,
结合亚基传递给原肌
肌
Ca2+
昆虫学3
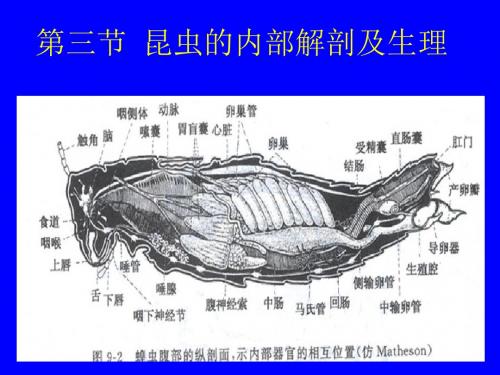
郊蟋蟀 美国家蠊
血腔(haemocoele) 背血窦(dorsal sinus) 腹血窦(ventral sinus) 围脏窦(perivisceral sinus) 背膈(dorsal diaphragm) 腹膈(ventral diaphragm)
消化系统及营养 循环系统 排泄系统及废物排除 气管系统和气体交换 神经系统及协调 内分泌系统及激素的功能 生殖系统
单极神经元
双极神经元
多极神经元
感觉神经元
运动神经元 联络神经元
神经节(ganglion)
神经髓(neuropile)
神经索
腹神经索
中枢神经系统(central nervous system) 脑 腹神经索 神经索(connective) 神经连锁(commissure) 咽下神经节(suboesophageal ganglion)
六、内分泌系统及激素的功能
• 激素(hormone):是指由 内分泌器官分泌的、具有 高度活性的微量化学物质 • 激素的发现 切头试验、结扎试验、接 躯试验、腺体移植试验、 腺体摘除试验
1. 昆虫的内分泌细胞和器官
类型:神经内分泌细胞 腺体内分泌器:心侧体,咽侧体 前胸腺
A 神经分泌细胞 (neurosecretory cell, NSC)
微气管(tracheole)
气囊(air sac)
气门(spiracle)
气管系统的呼吸机制
通风作用
扩散作用
昆虫的其它适应性呼吸 方式
体壁呼吸 气管鳃呼吸 气管鳃(tracheal gill)
直肠鳃(rectal gill)
气泡和气膜呼吸
五.神经系统及协调
神经系统的基本构造
昆虫的飞行力学昆虫的飞行机制和飞行能力的研究

昆虫的飞行力学昆虫的飞行机制和飞行能力的研究昆虫的飞行力学昆虫是地球上最成功的生物之一,它们在飞行方面展现了令人惊叹的能力。
昆虫的飞行机制和飞行能力一直是科学家们的研究重点。
本文将介绍昆虫飞行力学的研究,探讨昆虫的飞行机制和其在工程设计中的应用。
一、昆虫的飞行机制1. 翅膀结构昆虫的翅膀是其飞行的主要器官。
翅膀由薄而坚硬的外壳覆盖,内部则由与鸟类的羽毛不同的膜翅组成。
膜翅上布满了细小的静脉,这些静脉起到了加强翅膀的结构并保持其形状的作用。
2. 翅肌的调节昆虫的胸部上有一对强有力的肌肉,称为翅肌。
翅肌通过不同的收缩和放松来控制翅膀的运动。
这种精确而高效的肌肉控制使得昆虫可以根据需要调整翅膀的频率、幅度和相位。
3. 空气动力学昆虫的飞行受到空气动力学原理的支配。
研究表明,昆虫的翅膀在下颤翅飞行时产生的上扬和向前方推力是产生升力的主要原因。
昆虫通过改变翅膀的运动方式和翅膀形状来实现对升力和阻力的控制。
二、昆虫的飞行能力1. 短途飞行昆虫中有许多种类适应于短途飞行,如蜜蜂、蝴蝶等。
它们通常以花粉或花蜜为食物,并在花朵间短距离飞行。
这些昆虫的飞行速度相对较慢,飞行姿态相对较稳定。
2. 长途迁徙另一些昆虫则展示了惊人的长途迁徙能力,如蚂蚁、螽斯等。
它们能够持续飞行数小时或数天,覆盖数十甚至数百公里的距离。
这些昆虫通常以觅食或繁殖为目的进行长途迁徙。
三、昆虫飞行力学在工程设计中的应用昆虫的飞行机制和能力对工程设计具有重要的启示。
以下是一些应用领域:1. 空飞器设计受到昆虫的飞行机制启发,科学家们尝试设计出更加高效和稳定的空飞器。
例如,研究人员通过模仿蜻蜓的飞行方式,设计出了具有优秀悬停和机动性能的微型无人机。
2. 生物医学工程昆虫的飞行机制对生物医学工程领域也有潜在的应用。
例如,通过研究蚊子的飞行机制,可以改进昆虫仿生机器人在狭小环境中的操控能力,从而应用于微创手术等领域。
3. 轻巧材料设计昆虫的翅膀结构和飞行力学为轻巧材料的设计提供了灵感。
昆虫活性蛋白

• 二、免疫清道夫 昆虫活性蛋白富 含几丁质。而且昆虫几丁质的纯度要 比虾蟹类几丁质纯好多倍,几丁质是 一切生物生命力的重要支柱之一,被 誉为继蛋白质、糖、脂肪、维生素、 矿物质之后的“第六生命营养要素”。 它就像辛勤的园丁,在人体免疫系统 内起着三调(双向免疫调节、调节pH 值、调节荷尔蒙)、三排(排细胞和人体的内 部环境。
• 五、免疫激活 除抗菌肽、防御素外,昆 虫活性蛋白还含外源性凝集素,它可以促进 细胞相互粘接并抑制其增殖,不仅能使正常 细胞更富活性,并能杀灭变异细胞,抵御病 毒蔓延,激活免疫力,有效防治胃肠道炎症 及各种感染性疾病。
• 谢谢大家 •09105050 马超婕
既然这么有用,那就来具体的介绍 一下它的生理功能
• 一、免疫特警 昆虫活性蛋白独含 的抗菌肽成分,当肌体受损或病原微 生物入侵时,抗菌肽快速跟踪、追杀 入侵者,抗菌肽就像免疫特警一样, 用锐利的尖刀迎击“敌人”,在细菌、 病毒的胞膜结构上凿出离子通道,使 细菌胞膜结构破坏,引起细胞内水溶 性物质外流,从而彻底杀死细菌。人 类常食昆虫蛋白,就象给身体安上了 一道道屏障,百病难浸!
大家知道蛋白质的种类吗?
• 第一代蛋白质是植物蛋白,属于营养型蛋 白。主要从豆类、五谷、坚果、花生等食物中 提取。植物蛋白的缺点是营养素不全面——未 全部包含人体必需的8种氨基酸。 • 第二代蛋白质是动物蛋白,属于调理型蛋 白。主要从乳、蛋、肉、鱼、禽等食物中提取。 第二代蛋白的营养比第一代全面,但缺点是胆 固醇过高。 • 第三代蛋白质是昆虫活性蛋白,作为换代 产品,昆虫活性蛋白属于全效型蛋白,集预防、 保健、修复、营养四效于一体,功效明显优于 第一、第二代蛋白,并科学地解决了第一、第 二代蛋白的缺陷
• 我们现在2 1 世纪的主导食品是功能 性食品,而昆虫食品正是典型的功能 性食品。鉴于昆虫是一大类发展潜力 巨大的生物资源和营养宝库,能食能 药,可以作为运动员、老年人、少年 儿童的高级营养品和有效药品,所以今 天由我来为大家介绍一下昆虫蛋白
昆虫生理学基础—昆虫的循环系统概述

2,血糖:
海藻糖 Trehalose 优点:它具有保护生物细胞和生物活性物质
在脱水、干旱、高温、冷冻、高渗透压及有毒试剂 等不良环境条件下活性免遭破坏的功能。
由于海藻糖具有广泛而独特的生物学功能, 广泛受到各个国家的密切关注,以致引发了世界性 的海藻糖研究和开发热潮。
海藻糖,海藻糖作为血糖对昆虫的开放式循环非常有利。
血液的组成和物理性状
昆虫的血液包括血细胞和血浆两部分,除少数昆虫(如 摇蚊幼虫)因含血红素而呈红色外,大多数呈黄色、橙色 或蓝绿色。昆虫的血液一般占虫体容积的15%一75%。 一、血 细 胞
血细胞(Hemocytes)指悬浮在血浆中的游离细胞,约占 血液的2.5%,昆虫血细胞种类常因观察方法的不同而有较 大的差异,但最基本的血细胞可分为6类:原血细胞、浆 血细胞、粒血细胞、珠血细胞、类绛色细胞和凝血细胞。
通常位于触角、翅和附肢的基部或内部,一 般不与背血管相连,他们由含肌纤丝的薄隔所组 成,有膜状、瓣状、管状或囊状等多种形状。
三、造血器官 造血器官是昆虫体内不断分化并释放
血细胞的囊状构造,周围有膜包被,膜囊 内有相互交织的类胶原纤维和网状细胞。
造血器官只在幼虫时期存在,到成虫时期就会 消失。
在不同的昆虫中,造血器官的位置也不同: 1,膜翅目幼虫的造血器官位于胸部的脂肪体附 近。 2,鳞翅目幼虫的造血器官位于翅芽周围。 3,双翅目幼虫的造血器官位于大动脉上
5,其他: 非肽类的激素、色素、有机酸、多元醇、神
经调节物质以及消化道吸收和共生菌合成的其他 物质。
心脏的搏动与血液循环
昆虫的血液循环主要靠心脏和辅搏器的搏动以及膈 膜和肌肉的运动来完成。
一、心 搏 昆虫的心脏由单细胞层的心肌所组成,里面为一 层很薄的基膜,外周是结缔组织构成的围膜。昆 虫的心脏是肌原性的,它不受神经的支配,可自 发产生动作电位引起收缩,随后由心脏壁的弹性 产生舒张,从而进行有节律性的搏动。
虫类蛋白质与免疫抗体的结构与功能分析

虫类蛋白质与免疫抗体的结构与功能分析在生物学中,蛋白质是最重要的有机分子之一,扮演着许多关键生物学过程中不可或缺的角色,其中包括激素、酶、抗体、结构蛋白等。
虫类蛋白质和免疫抗体作为其中的两种,具有重要的生理学意义。
本文将从虫类蛋白质和免疫抗体的结构与功能等方面进行分析和探讨。
一、虫类蛋白质的结构与功能虫类蛋白质结构主要由α螺旋和β结构组成。
虫类蛋白质分为丝蛋白、角蛋白、胶原蛋白和肌凝蛋白。
丝蛋白存在于丝绸等昆虫的丝绸中,主要有光泽、光滑、均匀等特点。
角蛋白则存在于昆虫的外骨骼(表皮),能够形成类似甲壳的硬结构,扮演着保护昆虫体内器官的作用,另外角蛋白还具有与丝蛋白不同的柔韧、透气等性质。
胶原蛋白存在于许多节肢动物的组织中,如肌肉、骨骼、软骨、皮肤、血管、腱等,是维持这些组织的主要成分。
肌凝蛋白存在于昆虫肌肉中,是构成肌肉纤维的主要成分。
虫类蛋白质的功能主要表现在两个方面,第一个是生物学作用,例如:肌凝蛋白能够通过肌肉收缩机制调节昆虫的运动。
此外,角蛋白可增加外骨骼的硬度和强度,防御来自外部环境的机械冲击、攻击或捕食。
第二个作用是工业技术应用,例如丝蛋白可制作衣物、绸被等,角蛋白可制作人造可分离式手术缝线等。
二、免疫抗体的结构与功能免疫抗体是一种对抗外部入侵的物质,其结构由多种蛋白质组成,形成具有高度变异性的特定结构和功能。
抗体由基础分子(IgM,IgA,IgG,IgE和IgD)组成,基础分子在表现出了在保持某些结构的同时,将某些域暴露在抗原上,从而实现免疫保护效果。
免疫抗体是一种特殊的蛋白质,主要的功能是与外部的抗原结合,防御入侵。
其结构形成了一个Y形,其中最上面的“Y”形部分为抗原结合部位。
抗原在抗体结构中与氨基酸侧链相互作用,使得抗体与抗原结合。
抗体作为免疫系统的主要组成部分,处于抵御感染的前沿。
实验和临床研究表明,通过对免疫系统的信息学探究对抗感染是必要的,而免疫抗体在防范疾病传播或治疗是否有效等方面都有很好的应用性。
昆虫飞行原理

昆虫飞行原理
昆虫的飞行原理是一项复杂但高效的生物学奇迹。
它们利用了气流动力学和肌肉的协同作用来实现飞行。
首先,昆虫的翅膀是其飞行的关键。
翅膀构造轻巧且柔软,由透明而坚韧的蛋白质组成。
这种构造使得翅膀能够快速振动,并产生足够的升力来支持昆虫的身体。
昆虫的飞行过程中,翅膀的振动方式也很关键。
它们采用了交替挥动的方式,即一对翅膀在上下运动时是反向的。
这种挥动方式可以产生稳定的升力,并使昆虫保持平衡。
另外,昆虫的翅膀还可以调整其形状和角度。
通过调整翅膀的形状,昆虫可以改变所受到的气流阻力,以便更好地适应不同的飞行条件。
此外,昆虫还可以调节翅膀的角度来控制飞行方向和速度。
肌肉是昆虫飞行的另一个重要组成部分。
昆虫的胸部有一组强大的飞行肌肉,这些肌肉通过快速的收缩和放松来驱动翅膀的振动。
此外,昆虫的胸部还有一些辅助肌肉,用于控制翅膀的角度和形状。
在昆虫飞行中产生升力的基本原理与飞机类似。
当昆虫向下挥动翅膀时,翅膀和空气之间会形成一个较大的气流区域。
此时,翅膀上的压力较低,而下方的气流速度较高。
根据伯努利定律,气流速度较高的地方压力较低,从而在翅膀上产生了一个向上的升力。
总结起来,昆虫飞行的原理主要包括翅膀的振动方式、翅膀的形状和角度调整、胸部肌肉的驱动以及气流动力学原理。
通过这些生物学的适应和机制,昆虫能够实现高效、灵活的飞行。
昆虫的肌肉系统

第四节 肌肉的收缩机制
4.1 肌纤丝
——细肌丝 ——细肌丝
由肌动蛋白、原肌球蛋白和肌钙蛋白3种蛋白组成。 肌动蛋白、原肌球蛋白和肌钙蛋白3种蛋白组成。 肌动蛋白(actin)单体呈球状,串联成两条肌动蛋白链, 肌动蛋白(actin)单体呈球状,串联成两条肌动蛋白链,形成细纤 丝的主体。肌肉收缩时,能与肌球蛋白结合成肌动球蛋白横桥。 丝的主体。肌肉收缩时,能与肌球蛋白结合成肌动球蛋白横桥。 原肌球蛋白(tropomyosin)纤维状,镶嵌在肌动蛋白双链的凹槽内。 原肌球蛋白(tropomyosin)纤维状,镶嵌在肌动蛋白双链的凹槽内。 个分子可覆盖7个肌动蛋白单体,并可与肌钙蛋白分子结合。 1 个分子可覆盖 7 个肌动蛋白单体, 并可与肌钙蛋白分子结合。 肌肉收 缩时,可阻止肌动球蛋白横桥的形成。 缩时,可阻止肌动球蛋白横桥的形成。 肌钙蛋白( troponin) 肌钙蛋白 ( troponin ) 是 一种调节蛋白, 一种调节蛋白 , 在细肌丝上 每隔7个肌动蛋白单体就有1 每隔 7 个肌动蛋白单体就有 1 个肌钙蛋白分子。 个肌钙蛋白分子 。 含有 3 个 亚基,即肌动蛋白结合亚基、 亚基 , 即肌动蛋白结合亚基 、 钙结合亚基和原肌球蛋白结 合亚基。 合亚基。
第一节 肌肉的类型
1.2 体壁肌
由于所有节肢动物属于外骨骼动物, 由于所有节肢动物属于外骨骼动物, 所以昆虫的体壁 肌(skeletal muscle)相当于高等动物的骨骼肌。 muscle)相当于高等动物的骨骼肌。 昆虫的骨骼肌由长形的平行肌纤维组成, 昆虫的骨骼肌由长形的平行肌纤维组成,着生在体壁 下或由体壁内陷形成的内突上, 下或由体壁内陷形成的内突上, 常按体躯的自然分节而 分节,专司体节、附肢和翅的运动。如背纵肌、腹纵肌、 分节, 专司体节、附肢和翅的运动。如背纵肌、腹纵肌 、 背腹肌等。 背腹肌等。 组成肌肉的基本单位是肌纤维, 组成肌肉的基本单位是肌纤维, 肌纤维中含有大量的 肌原纤维(肌肉收缩的基本单元) 肌原纤维( 肌肉收缩的基本单元)。根据肌原纤维在肌 纤维中的排列情况,可把体壁肌分为束状肌 管状肌、 束状肌、 纤维中的排列情况, 可把体壁肌分为束状肌、 管状肌、 纤维状肌3种类型。 纤维状肌3种类型。
昆虫是如何飞行的

昆虫是如何飞行的昆虫是地球上最为成功的生物之一,它们在进化过程中发展出了独特的飞行能力。
昆虫的飞行方式多种多样,包括蝴蝶的翅膀振动、蜜蜂的翅膀摆动、蚊子的翅膀扇动等。
本文将介绍昆虫飞行的原理和机制。
一、昆虫翅膀的结构昆虫的翅膀是它们飞行的关键器官,翅膀的结构对于昆虫的飞行能力起着至关重要的作用。
昆虫的翅膀由薄而坚韧的外骨骼和透明的膜翅组成。
膜翅上有许多细小的脉络,这些脉络使得翅膀具有足够的强度和刚度,同时又能保持足够的柔韧性。
二、昆虫飞行的原理昆虫的飞行原理主要包括空气动力学和肌肉运动两个方面。
1. 空气动力学昆虫的翅膀在飞行过程中产生了空气动力学效应。
当昆虫振动翅膀时,翅膀上的脉络会产生气流,形成一个高压区和低压区。
高压区使得昆虫向上升起,低压区则使得昆虫向前推进。
这种空气动力学效应使得昆虫能够在空中飞行。
2. 肌肉运动昆虫的飞行依赖于肌肉的运动。
昆虫的胸部有强大的飞行肌肉,这些肌肉通过收缩和放松来驱动翅膀的运动。
昆虫的翅膀在飞行过程中以高频率振动,每秒钟可以振动数百次甚至上千次。
这种高频率的振动使得昆虫能够产生足够的升力和推力,从而实现飞行。
三、昆虫飞行的机制昆虫的飞行机制主要有两种,分别是直翅目和膜翅目。
1. 直翅目直翅目昆虫的翅膀与身体相连,不能自由摆动。
它们通过改变翅膀的角度和形状来调节飞行姿态和飞行速度。
直翅目昆虫的飞行速度较快,飞行稳定性较高。
2. 膜翅目膜翅目昆虫的翅膀与身体之间有一条薄膜相连,可以自由摆动。
它们通过振动翅膀来产生升力和推力,实现飞行。
膜翅目昆虫的飞行速度较慢,飞行稳定性较差。
四、昆虫飞行的适应性昆虫的飞行适应性非常强,它们可以在各种环境条件下飞行。
昆虫的翅膀结构和飞行机制使得它们能够在狭小的空间中飞行,如蜜蜂在花丛中采蜜。
昆虫的飞行速度和灵活性使得它们能够逃避天敌的追捕,如蝴蝶在花园中飞舞。
昆虫的飞行能力还使得它们能够迁徙和寻找食物,如蚊子在夜晚中寻找血液。
总结起来,昆虫的飞行是通过翅膀的振动和空气动力学效应实现的。
蜜蜂翅膀肌肉的生理生态学

蜜蜂翅膀肌肉的生理生态学在微小而神奇的昆虫世界里,蜜蜂代表着勤奋与完美。
它们为人类提供了蜜蜡,蜜蜂蜜以及传粉等重要服务。
蜜蜂的翅膀是它们能够飞行、采花採蜜的关键器官,而这一切的源动力就是蜜蜂翅膀的肌肉。
本文将着重阐述蜜蜂翅膀肌肉的生理生态学特征。
一、蜜蜂翅膀肌肉的结构蜜蜂翅膀肌肉是由高度肌纤维化的横纹肌构成,以适应机体需要的高强度变幅振动运动。
由于蜜蜂需要长时间在空中飞行,因此翅膀肌肉的代谢需求十分强烈。
在翅膀肌肉中,有丰富的线粒体和肌纤维,能够快速地氧化葡萄糖,释放出大量的能量。
此外,蜜蜂翅膀肌肉的肌纤维要比其他昆虫的翅膀肌肉更丰富。
因此,蜜蜂在飞行过程中,能够更加灵活地控制翅膀的变幅振动,从而保持飞行的平稳度。
蜜蜂的翅膀还有一个独特的设计是翅膀根部有一个特殊的连接器,使得蜜蜂可以控制翅膀的相位和波形,能够在高速飞行中维持测向性控制。
二、蜜蜂翅膀肌肉的生物力学特性蜜蜂的飞行速度可以达到每小时40km/h,翅膀频率可以高达200次/秒,而每次翅膀运动的幅度也可以达到90度。
以上的高速和高频振动需要蜜蜂的翅膀肌肉具有较高的生物力学特性。
在蜜蜂的翅膀肌肉中,运动头节点是肌肉的重要结构特征,这些节点的分布和连通是决定翅膀运动的关键因素。
同时,整个肌群的条状结构、路径和血管分布等特点也使得蜜蜂的翅膀肌肉拥有较高的稳定性和灵活度。
三、蜜蜂翅膀肌肉的代谢特征由于长时间的飞行需要消耗较大的能量,蜜蜂翅膀肌肉的代谢特征十分显著。
翅膀肌肉的功能主要通过三种代谢途径实现,即三磷酸腺苷(Adenosine Triphosphate, ATP)酶、肌酸激酶和磷酸化代谢。
其中,ATP酶是肌肉瞬间分解葡萄糖释放出来的能量,主要在短时间高强度运动中发挥作用。
肌酸激酶可以将磷酸转移至肌酸上,部分进入ATP产生途径。
而磷酸化代谢则是通过蛋白质的磷酸化,以ATP合成为能量的方式来进行长时间的运动。
四、蜜蜂翅膀肌肉的环境适应蜜蜂的生存环境广泛,不仅在不同的季节通过对群体进化适应了不同的生存环境。
第十五章昆虫的肌肉系统

2. 肌肉的组织结构
肌细胞(肌纤维) 肌细胞(肌纤维) 肌原纤维
2.1 肌细胞
肌肉是肌细胞的总称, 肌肉是肌细胞的总称 , 肌肉运动就是肌细胞活动的表 现。因肌细胞呈细长的纤维状,故又称肌纤维 fiber) (muscle fiber)。 在肌原纤维之间,有许多纵行的小管网络, 在肌原纤维之间 , 有许多纵行的小管网络 , 构成纵管 系统,它们由内质网分化而来,因此又称肌质网 肌质网。 系统,它们由内质网分化而来,因此又称肌质网。 肌原纤维间还有排列整齐的大型线粒体即肌粒 肌原纤维间还有排列整齐的大型线粒体即 肌粒 , 是肌 肌粒, 原纤维收缩时ATP的直接供应者 的直接供应者。 原纤维收缩时ATP的直接供应者。 线粒体附近还有穿过肌膜的微气管, 线粒体附近还有穿过肌膜的微气管 , 为线粒体氧化代 谢提供充足的氧气。 谢提供充足的氧气 。 昆虫内脏肌的肌纤维通常是单核 而体壁肌大多是多核的( 15-ZH) 的,而体壁肌大多是多核的(图15-ZH)。
3.3 肌肉收缩及滑行学说
1954年 Huxley&Hanxon根据甘油抽提肌原纤维的 1954年 , Huxley&Hanxon根据甘油抽提肌原纤维的 相差显微镜观察,以及Huxley Niedergerke用活肌肉纤 相差显微镜观察,以及Huxley & Niedergerke用活肌肉纤 维的干涉显微镜观察, 分别独立地提出了肌丝滑行学说 维的干涉显微镜观察 , 分别独立地提出了 肌丝滑行学说 : 肌丝滑行学说:
引起肌丝滑行的动力是肌动球蛋白横桥键角的改变 引起肌丝滑行的动力是 肌动球蛋白横桥键角的改变 , 肌动球蛋白横桥键角的改变, 当肌膜的兴奋由横管系统传人肌质网时, 当肌膜的兴奋由横管系统传人肌质网时 , 肌质网便释 放出大量的Ca2+ ,Ca2+浓度取决于肌纤维的兴奋程度。 浓度取决于肌纤维的兴奋程度。
昆虫体表成分

昆虫体表成分
昆虫的体表(包括外皮)是由几种主要成分构成的:
1.几丁质(Chitin):这是昆虫外骨骼的主要成分,通常存在于几丁质甲壳素的形式。
几丁质是一种高分子聚合物,与蛋白质和其他碳水化合物相结合,形成一种坚硬而轻便的结构,可保护昆虫免受外部环境的伤害。
2.蛋白质(Protein):在昆虫的体表中也存在蛋白质,虽然不如几丁质那么显著。
蛋白质在组成外骨骼和肌肉等方面起到重要作用。
3.脂肪(Fat):脂肪是一种广泛存在的脂质,它们主要是用来存储能量和作为隔热材料。
脂肪通过蜡膜形式的蜡酯和内生脂质表现在昆虫体表。
4.氨基糖(Amino Sugar):氨基糖是几丁质的组成部分之一,其主要的两种类型是N-乙酰葡萄糖胺和N-乙酰半乳糖胺。
5.内生脂质(Endogenous Lipid):这些脂质存在于昆虫表皮的蜡层中,使其变得不透水,对植食性昆虫而言尤其重要,因为这种防水特性可以减少水的蒸发。
6.蜡质(Wax):这是一种外源性成分,昆虫通常通过蜡酯合成并将其应用于外表皮肤以保护皮肤。
此外,它还可用于昆虫之间的通信。
这些组分一起构成了昆虫体表的结构,并在保护、渗透调节、保湿、生理代谢、感知等方面发挥作用。
它们是昆虫体内外环境的重要桥梁。
储藏物昆虫的生理学—肌肉系统及其生理

四、昆虫肌肉收缩的机制和能量来源 昆虫肌肉收缩的能量来源一般分为两种情况: 足肌和一般体壁肌在肌肉收缩时,细胞中的中间代谢物α-磷酸甘油酯、丙酮酸 和乳酸被释放出米,参加线粒体中的三羧酸循环与呼吸链进行的“氧化磷酸化耦联” 等反应合成ATP,开供应昆虫活动所需要的能量。 在昆虫的飞翔肌中,主要以“磷酸二羟丙酮”参加α-磷酸甘油酯循环和呼吸链 进行有氧代谢,供应快速飞翔需要的大量能量。昆虫在飞翔中不产生乳酸或产生很 少量的乳酸,产生的丙酮酸也较少。此外,昆虫翅肌中还含有占体内全磷量50%的 磷酸化合物及75%的磷酸腺苷(ATP、ADP、AMP)和磷酸精氨酸等能量转移化合物。
第八节 肌肉系统及其生理
三、肌肉的结构 昆虫肌肉上的神经分布形式属于多支神经分布型,即每一条肌纤维上分布有几 个神经元发出的神经末梢。这些神经末梢在肌纤维上相互间隔的距离很小,分布非 常均匀。在成对分布的神经纤维中,有一条控制肌肉快速收缩的神经纤维,称为 “快收缩神经”;另一条控制慢收缩和松弛的神经纤维,称为“慢收缩神经”。有 些种类昆虫的神经末梢分成很多细枝,分布到肌纤维上或环绕着肌纤维;有些种类 的神经末梢不伸入肌纤维而终止于肌纤维之间;还有些种类的神经末梢形成一扁形 或锥形的“端板”(叫杜氏锥)终止于肌肉的表面。昆虫的神经与肌肉的接点也形 成突触性接触。但许多资料报道,昆虫体躯肌肉的神经传导递质不是乙酰胆碱,而 是氨基丁酸和谷氨酸。
第八节 肌肉系统及其生理
三、肌肉的结构 昆虫的肌肉由许多平行排列的肌纤维组成。肌纤维是一种细长的多核细胞,外 面包有一薄层肌膜,其内由许多细而平行的和充塞于其间的肌质所组成。各肌原纤 维的明暗带相并列,使整条肌肉纤维呈现出明暗相间的多段现象,这种肌肉被称为 横纹肌。 每根肌原纤维由若干个较宽的A带和较窄的I带交替组成。A带由含肌球蛋白的 粗肌丝和含肌动蛋白的细肌丝所构成。I带由只含肌动蛋白的细肌丝所构成。在光学 显微镜下,密度大、折光系数高的A带发暗,称为暗带;密度小、折光系数低的I带 发亮,称为明带。在暗带中央有一小段发亮的小区,称为中带,中带只含有由肌球 蛋白构成的粗肌丝。在明带中央有一条致密而狭窄的Z线,叫做端膜,两端膜之间的 一段称为一个肌节。
蛋白质的三级结构和特点是什么

蛋白质的三级结构和特点是什么
蛋白质三级结构指一条多肽链在二级结构或者超二级结构甚至结构域的基础上,进一步盘绕,折叠,依靠次级键的维系固定所形成的特定空间结构称为蛋白质的三级结构。
蛋白质的三级结构
蛋白质分子处于它的天然折叠状态的三维构象。
三级结构是在二级结构的基础上进一步盘绕,折叠形成的。
三级结构主要是靠氨基酸侧链之间的疏水相互作用,氢键,范德华力和静电作用维持的。
蛋白质三级结构的特点
1.含多种二级结构单元;
2.有明显的折叠层次;
3.为紧密的球状或椭球状实体;
4.分子表面有一空穴(活性部位);
5.疏水侧链埋藏在分子内部,亲水侧链暴露在分子表面。
蛋白质的结构
蛋白质是以氨基酸为基本单位构成的生物高分子。
蛋白质分子上氨基酸的序列和由此形成的立体结构构成了蛋白质结构的多样性。
蛋白质具有一级、二级、三级、四级结构,蛋白质分子的结构决定了它的功能。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
昆虫飞行肌蛋白质3杨 璞 余海忠 程家安 祝增荣33浙江大学应用昆虫学研究所 杭州Τηεπροτεινσινφλιγητµυσχλεσοφινσεχτσ ≠ °∏ ≠ 2 ≤ ∞ 2 2 33ΙνστιτυτεοφΑππλιεδΕντοµολογψ ΖηεϕιανγΥνιϖερσιτψ ∏ ≤Αβστραχτ ∏ ∏ ∏ × ∏ ∏ ∏ √ ∏ Κεψωορδσ∏ 摘 要 昆虫飞行肌的肌原纤维不仅含有粗肌丝!细肌丝!纤肌丝 还含有很多其它蛋白质参与肌原纤维的组装和调节文章介绍了 余种蛋白质的结构!功能及其在肌原纤维中的位置和功能 对于了解昆虫飞行肌的发育和探索昆虫飞行能力差异的原因具有重要意义∀关键词 飞行肌 肌原纤维 肌丝 蛋白质3国家科技部重点基础研究计划 项目 ≤ 2 ∀33通讯作者 ∞2 ∏ ∏ ∏收稿日期 2 2 修回日期 2 2昆虫是无脊椎动物中惟一有翅的一类 飞行肌是昆虫特有的肌肉类型尽管昆虫的飞行能力各不相同 然而其飞行肌却有相似的微观结构肌细胞呈细长的纤维状 故又称肌纤维 其中有特化的功能细胞器)))肌原纤维 在偏光显微镜下呈现出明暗相间的带状构造 在明带 带 中部有薄膜 盘 贯穿其间 相临薄膜之间的部分构成肌节是肌细胞收缩的基本单位∀电镜下可见肌原纤维由粗!细 种肌丝构成 它们沿肌纤维的长轴并按规则的空间布局互相穿插平行排列∀粗肌丝的成分是肌球蛋白 细肌丝的主要成分是肌动蛋白 辅以原肌球蛋白和肌钙蛋白 图 ∀∏¬ 和¬ 根据观察结果提出了著名的肌丝滑动模型 肌肉收缩是粗肌丝和细肌丝的相对滑动引起的 而粗细肌丝本身长短不发生变化肌丝滑动的动力是肌动球蛋白横桥键角的改变∀该模型说明了肌节如何有效地把肌球蛋白分子的运动转化成肉眼可见的肌肉纤维的伸缩≈ 并未从分子水平说明各种蛋白质如何协调并组装成高度有序的肌原纤维∀随着实验技术的发展和研究的深入 不断又有新的蛋白质发现∀图1 果蝇飞行肌肌节结构及其组成蛋白质[4]1 粗丝结合蛋白在无脊椎动物横纹肌中有些蛋白只和粗肌丝结合 比如果蝇的副肌球蛋白!小副肌球蛋白!肌球杆蛋白以及飞行蛋白≈ ∀在果蝇的某些肌肉中这几种蛋白质都出现在 带≈ ∀下面介绍一下这几种粗丝结合蛋白∀111肌球杆蛋白肌球杆蛋白 ° 分子量 ⁄ 它是飞行肌肌原纤维的主要组分之一 是一种新发现的收缩性蛋白∀果蝇肌球蛋白重链 √ ≤ 由一个单基因产生∀该基因通过 的可变剪接 可产生 种不同的 ≤ 它还含有第 个转录单元 始于第 个内含子 编码 °∀ °包括 ≤杆状区域和 端含有 个独特的氨基酸残基的区域 这 个氨基酸取代了重链的运动区域和头部 ≥ 的轻链结合部位∀ °还存在于心脏和成虫的内脏肌肉以及幼虫胚胎中∀在这些部位 °沿着 ≤聚合成粗肌丝∀免疫印迹分析表明 在许多成虫的直接飞行肌中 °与 ≤的比例为 Β ∀ °影响肌节的结构∀ °存在于快速收缩肌肉 间接飞行肌 !慢速收缩肌肉 肠道 以及收缩性很强的肌肉 幼虫体壁 中 这说明 °的形成并不只限于某些特定的肌肉中 它在果蝇肌肉中普遍具有重要作用≈ ∀112副肌球蛋白副肌球蛋白 和肌球蛋白是粗肌丝中含量最高的蛋白 所有研究过的无脊椎动物肌肉中都含有副肌球蛋白∀它是一个杆状分子 其活动中心有 个Α 螺旋∀果蝇副肌球蛋白分子量大约是 ⁄ 由 个氨基酸残基组成 中间的 个氨基酸残基形成该Α 螺旋∀ 个副肌球蛋白能够聚合成卷曲螺旋型的二聚体∀对肌球蛋白和副肌球蛋白氨基酸序列分析发现一个明显的模式 个氨基酸残基重复序列的带电中心 这些电荷中心带有相反的电荷 交替排列 它们之间的相互作用对于蛋白质组装成粗肌丝非常重要≈ ∀一般认为副肌球蛋白帮助粗肌丝组装 对秀丽新小杆线虫Χαενορηαβδιτισελεγανσ突变体的分析表明 副肌球蛋白的含量影响粗肌丝的长度和直径 为了解副肌球蛋白在肌原纤维组装过程以及肌肉收缩过程中的作用 和 通过果蝇副肌球蛋白编码基因启动子区域的°因子 破坏果蝇副肌球蛋白编码基因 纯合的突变体果蝇在胚胎后期死亡 突变体在成肌细胞融合和肌原纤维组装中都有缺陷 肌节组装不当 肌肉没有伸缩性 说明副肌球蛋白在成肌细胞融合!肌原纤维组装以及肌肉收缩等方面都有重要作用≈ ∀113飞行蛋白飞行蛋白 是果蝇间接飞行肌中一种多磷酸化的肌原纤维蛋白≈ ∀∂ ∏¬等测定了果蝇飞行蛋白的氨基酸序列 其分子量为 ∗ ⁄ ∀这种蛋白质只在非同步飞行肌中出现 它对肌肉伸展活性具有重要作用≈ ∀飞行蛋白是一种位于间接飞行肌粗肌丝的特异性蛋白 它使肌球蛋白疏水性氨基酸和带电氨基酸重复排列 并指导粗肌丝的正确组装≈ 有的研究已表明 飞行蛋白对正常肌节的结构和肌肉收缩具有重要作用 它与肌球蛋白杆部相互作用 对粗肌丝形成正确的长度以及肌肉收缩后粗肌丝的稳定是必需的≈ 昆虫飞行肌中缺少飞行蛋白会出现粗肌丝长度增加 表明它具有调节粗肌丝组装的作用≈ ∀2肌联蛋白家族迄今为止 普遍认为肌原纤维是由粗肌丝和细肌丝有规律排列所形成的∀然而 在经过拉长的肌肉细胞内又观察到一种新的超细肌丝 它将粗肌丝与 线联接起来 不同学者给予它不同的名字 如第三肌丝!纤肌丝!≤ 肌丝!≥ 肌丝!× 肌丝!端肌丝!缝隙肌丝!弹性肌丝≈ ∀211肌联蛋白肌联蛋白 也称粗丝联接蛋白∀ 年代中期 由 ∏ 在肌动蛋白和肌球蛋白溶解残余物中 经≥⁄≥2° ∞发现 条蛋白染色带 一条是肌动蛋白 另一条是分子量极大的蛋白质 称为肌联蛋白∀与此同时 王世宽在进行骨骼肌肌动蛋白结合蛋白质的研究中 也意外地发现了这种分子量巨大的蛋白质 定名为 ≈ ∀免疫细胞化学观察到抗肌联蛋白抗体结合于纤肌丝 这清楚地证明了肌联蛋白是纤肌丝的构成成分≈ ∀肌联蛋白是横纹肌中已知分子量最大的蛋白质∀其含量在肌肉中占第 位∀脊椎动物肌联蛋白分子量是 ⁄ 其中 是由 个氨基酸残基组成的免疫球蛋白 ≤ 结构区域和纤连蛋白 ƒ 样结构域的重复单位串联而成 是独特的 序列 包括°∞∂ 富含脯氨酸°!谷氨酸∞!缬氨酸∂!赖氨酸 结构域和一个 激酶催化结构域≈ ∀在成熟的肌肉中 肌联蛋白纤丝长度大于 Λ 占肌节的一半 它的 端位于 盘 ≤ 端位于 带 一般认为肌联蛋白与粗肌丝和细肌丝缠绕在一起 通过 重复结构区域和特殊的°∞∂ 区域使肌肉具有弹性≈ ∀已表明肌联蛋白与肌球蛋白 包括肌球蛋白重链 ≤ !肌动蛋白结合蛋白! 带相关蛋白之间有多种相互作用≈ ∀在肌细胞生成时 肌联蛋白是出现最早的分子之一 早于肌动蛋白!肌球蛋白和其它收缩性成分≈ ∀ ∏ 等发现肌节蛋白质的形成顺序为 肌联蛋白 盘部分!肌球蛋白重链!Α 辅肌动蛋白!肌钙蛋白× ! 蛋白≈ ∀学者们对于肌联蛋白的作用产生以下 种假设 在肌肉发育过程中作为肌节组装的模板或骨架 作为成熟肌肉中的弹性成分∀大量的数据支持第 种假设≈ ∀研究表明 肌联蛋白的串珠结构的间距为 1 与肌球蛋白分子杆部Α螺旋的 个氨基酸残基的重复间距一致 而且在肌肉发育过程中 肌联蛋白是先于其它肌原纤维蛋白发生表达的≈ ∀在肌丝形成时 肌联蛋白可与Α 辅肌动蛋白! 等细胞骨架蛋白形成分子支架 肌球蛋白在此分子支架上组装 同时与肌动蛋白的组装相协调≈ ∀由此推测肌联蛋白有 种生理功能 首先 将粗肌丝与 线联接以维持肌原纤维的完整性和稳定性 其次 可能作为粗肌丝组装的模板蛋白质 第三 肌联蛋白≤端具有肌球蛋白轻链激酶的催化功能域 可使肌联蛋白自身或者肌球蛋白发生磷酸化而调节肌球蛋白的活性和控制粗肌丝的组装≈ ∀肌联蛋白基因家族的成员在脊椎动物和无脊椎动物中都存在 在无脊椎动物肌肉中也发现类肌联蛋白 称为颤缩蛋白 !凸出蛋白 也叫弹射蛋白 或者小肌联蛋白 2 ≈ ∀在果蝇中已经鉴定出了肌联蛋白家族中的 个成员 凸出蛋白≈ !连锁蛋白 ≈ !⁄ 肌联蛋白 ⁄2 ≈ ∀212凸出蛋白凸出蛋白 是果蝇中鉴定出的第 个肌联蛋白家族成员 它与颤缩蛋白 线虫的一种肌肉蛋白 的序列十分接近≈ 大约有 ⁄ 这种蛋白质首先在果蝇成虫间接飞行肌中被鉴定出来≈ 它从 盘伸出 结合在粗肌丝表面 另一端连在 盘 凸出蛋白分子链太短 只覆盖了不到半个肌节的长度≈ ∀凸出蛋白和脊椎动物中的肌联蛋白!秀丽新小杆线虫和海蛞蝓Στιχηοπυσϕαπανιχυσ中的颤缩蛋白以及几种无脊椎动物中的小肌联蛋白同属于一个家族≈ ∀免疫学研究表明不同无脊椎动物中 凸出蛋白处于肌节的不同位置≈ ∀在果蝇间接飞行肌中 抗体标记表明凸出蛋白在 带 然而在果蝇的其它肌肉中 抗体标记在肌节附近 不超出 带位置 与其在间接飞行肌中的位置正好相临≈ ∀对于凸出蛋白的进一步研究发现 在几种肌肉中它表现出不同形式 长度也不同 间接飞行肌中的凸出蛋白最小 远小于同步飞行肌中的 带≈ ∀和脊椎动物的肌联蛋白一样 凸出蛋白也有一个激酶区域 而且有很多 重复序列和ƒ 区域 但是凸出蛋白中没有°∞∂ 区域≈ ∀各种凸出蛋白部分水解消化以后基本相同 各种凸出蛋白可能只有部分区域相同 这可能是由于转录后加工所致 但最有可能是通过 的不同剪接造成的≈ ∀一种昆虫的肌联蛋白抗体能与其它昆虫中相应的蛋白质产生交叉反应 说明凸出蛋白和小肌联蛋白是不同肌肉的不同表达形式 果蝇凸出蛋白抗体和线虫小肌联蛋白交叉反应说明了这一点∀但凸出蛋白或小肌联蛋白和颤缩蛋白也不完全对应 它们的功能可能并不相同≈ ∀213连锁蛋白连锁蛋白 是一种很大的 盘蛋白 最早在桂花蝉Λετηοχερυσινδιχυσ中被鉴定≈ ∀在小龙虾和家蚕Βοµβψξµορι中也发现了与果蝇的 性质相似的肌肉蛋白≈ ∀其它分子量大的肌肉蛋白也有与 相似的 重复区域 比如在脊椎动物横纹肌中的肌联蛋白 所以 可能属于肌联蛋白家族≈ ∀为了进一步研究 的结构和功能 等测定了果蝇 的全部核苷酸序列 推测出了对应的氨基酸序列 蛋白质产物 ⁄ 与秀丽新小杆线虫相应的氨基酸序列比较 发现线虫和果蝇 的所有结构都很相似 主要都是由 重复区域构成 但没有发现纤连蛋白 样结构域和激酶结构域 这表明 在结构和功能上与肌联蛋白家族的其它成员截然不同≈ ∀所有的 的一级结构在进化过程中都是相当保守的 在果蝇 个氨基酸中 由 个免疫球蛋白结构域组成 结构域间有间隔肽分开 其中 Π 的间隔肽长度保持不变 并且都有 个相同的保守区域 推测是肌动蛋白的结合位点 惟一特别的是 端的 个氨基酸非重复序列≈ ∀的 个等位基因突变体的遗传学分析表明 对于肌节!肌腱正常结构的形成和保持非常重要∀所以 缺少了 活性 或 突变的杂合体 都不能形成肌节≈ ∀∂ ≥ 等提出 通过与细肌丝末端相连 促进了细肌丝的反向平行伸长 从而加固了细肌丝在 盘的锚定位点≈ ∀ 是 盘的形成和维持的基础 基因的连续表达对于肌肉的正常生长非常重要≈ ∀免疫显微镜观察发现 在很多类型的肌肉中都位于 盘区域≈ 标记有 抗体的飞行肌肌节的显微照片显示 盘的外围被染色 在提取了与 相连的肌动蛋白后 带边缘也褪色了 相反 带的另外一种蛋白的染色并不因为肌动蛋白的消失而褪色 免疫印迹分析发现 的≤ 端与肌球蛋白相连 由此推断 不只与肌动蛋白相连 还与粗肌丝的末端相连≈ 为肌动蛋白和肌球蛋白≤ 端提供与肌节的连接≈ ∀ 和凸出蛋白可能组成了昆虫飞行肌的弹性纤丝系统 使肌纤维具有弹性 这 种蛋白可能通过改变长度而调节肌肉纤维的弹性≈∀图2果蝇间接飞行肌中肌动蛋白!肌球蛋白!连锁蛋白和凸出蛋白的排列模型[27] 214⁄ 肌联蛋白⁄ 肌联蛋白 ⁄2 端位于 盘 长度大概是脊椎动物中× 长度的一半 具有脊椎动物的× 区域 即°∞∂ 区域!ƒ 区域和 重复序列 但没有× 激酶区域 × 的激酶功能似由凸出蛋白提供 ⁄2 和凸出蛋白共同行使或者同样具有脊椎动物肌肉× 的功能≈ ∀插入等位基因的⁄2 突变体分析表明 ⁄2 在肌肉发育过程中起重要作用 首先⁄2 在肌细胞生成期对于成肌细胞的融合有毋庸质疑的作用 其次⁄2 以后把肌原纤维组织成高度有序的排列 形成骨骼肌中的条纹≈ ∀目前仍不清楚⁄2 是如何调节肌原纤维和成肌细胞融合的 根据已知数据 有一种模型认为⁄2 与最初表达的细胞骨架蛋白相互作用 免疫化学并不认为⁄2 在肌肉发育初期独自形成组装的模版或骨架 更确切地说 ⁄2 是作为有弹性和粘性的带 上面有很多蛋白结合位点 比如 所结合的肌动蛋白结合位点∀在肌细胞生成过程中 ⁄2 粘附在 盘 把附着其上的细胞骨架拉开 没有⁄2 很多肌节成分比如肌球蛋白和肌动蛋白就不能组成高度有序的结构≈ ∀3其它蛋白还有一些蛋白质对于肌原纤维结构和组装具有重要作用 下面作一简要叙述∀311原肌球调节蛋白原肌球调节蛋白 ∏ 是肌动蛋白末端的封端蛋白 它调节横纹肌中细肌丝尖端肌动蛋白的动态组装 通过控制细肌丝的长度调节肌原纤维的组装 它结合于细肌丝的一端形成/帽子0 防止肌动蛋白的增加或减少 调节肌动蛋白纤维的长度∀细肌丝的 2 端被× 封闭 端被≤ 2 封闭∀为了了解肌肉发育过程中× 对间接飞行肌的作用 2⁄∏ 和ƒ 用果蝇进行研究 得出以下结论 肌原纤维组装时细肌丝末端延长 细肌丝末端处于动态的增长 封端蛋白在细肌丝末端从形成动态的/帽子0到变为永久的/帽子0封住细肌丝的末端 肌原纤维组装时细肌丝末端封顶对于细肌丝的最终长度!肌原纤维的结构以及肌肉的功能具有重要作用≈ ∀312Α 辅肌动蛋白Α 辅肌动蛋白 Α2 是一种重要的肌动蛋白交联蛋白 可以将肌动蛋白纤丝交联成平行排列成束的结构 在肌节中具有结构作用∀用钙蛋白酶 一种钙激活的蛋白酶 处理的肌原纤维 被钙蛋白酶消化 释放出Α 辅肌动蛋白 并且 盘失去了致密的物质≈ ∀此外 肌节中起结构作用的蛋白质还有 ∏ 从 盘伸出 与肌动蛋白纤维伴行 可能参与调节肌动蛋白纤维的组装≈ ∀4结语在所有已知的化合物中 蛋白质是最复杂的 同时也是生物体最重要的特征≈ ∀遗憾的是只有极少的昆虫飞行肌蛋白获得纯化 因此关于昆虫飞行肌蛋白质的化学结构的资料很少∀本文尽可能地就昆虫飞行肌蛋白质的结构和功能进行讨论 但是由于对昆虫蛋白质的结构和顺序的研究只在少数昆虫中进行 虽然参考文献比较丰富 但是这些文献大多数是通过某一个蛋白质等位基因的单核苷酸突变体进行该蛋白质的功能研究 因此在一定程度上限制了详细全面地叙述昆虫飞行肌的蛋白质∀如何将蛋白质分子的行为与肌原纤维的动态变化 蛋白质分子的结构与肌细胞的功能结合起来 将是以后研究的重点∀飞行肌蛋白质的结构分析!基因表达调节及肌丝的动态装配成为飞行肌研究的重要内容 并且蛋白质在飞行肌中的位置及其功能研究仍有待进行∀参考文献陈明1生物化学与生物物理进展 21 ∗≤ ⁄ ϑ ΧελλΒιολ 151 ∗≠ ƒ ⁄ ⁄ √ • ∏ ∞ ϑ ΧελλΣχι 113 ∗∏ ⁄ • ∂ ∏¬ ΝεωσΠηψσιολ Σχι14 ∗罗克斯坦 见 罗克斯坦 主编 昆虫生物化学 北京 科学出版社 ∗ƒ ∏ ≤ ∏ ∏ • ∏ ϑ ΧελλΣχι 107 ∗∏ƒ ∏ ≤ ≤ ≤ ≥ ƒ ∞ ƒ ϑ ΧελλΒιολ 140 ∗≤ ∏ ∂ ∏¬ ϑ ΧελλΒιολ 151 ∗∏ 2⁄∏ ≥ ≥ × . ≤ ≥ ϑ ΧελλΒιολ 160 ∗ ° ∞ ≥ ⁄ ≠ ∂ ∞ ≤ ° ƒ 2 ≤ ϑ Μολ Βιολ 331 ∗≥ ⁄ ⁄ √ ƒ 2 ≤ ∞ ≤ ° ϑ Μολ Βιολ 265 ∗• .⁄ ° × ƒ ∂ ∏¬ εταλ ϑ Μολ Βιολ 249 ∗• ∏ ϑ Βιολ Χηεµ 273 ∗∂ ≥ ∏ ⁄ ≤ ∏ ϑ Μολ Βιολ 285 ∗ 张蕾 国外医学 儿科学分册 30 ∗ ≥ ∏ 2≥ ∏ ϑ Μολ Βιολ 313 ∗⁄ ≥ ∏ 2≥ ∏ ϑ Μολ Βιολ 279 ∗∏ ƒ∏ ⁄ • ∏ Ευρϑ ΧελλΒιολ78 ∗≥ ∞ ≥ ≥ ϑ ΧελλΒιολ 148 ∗2≥ ∏ × × Ρεϖ Πηψσιολ Βιοχηεµ Πηαρµαχολ 138 ∗ƒ ƒ ΙνσεχτΒιοχη Μολ Βιολ 32 ∗ ƒ ∞ Βιοχ Βιοπηψσ Αχτα 1569 ∗ ∗° ∏ ≥ ⁄ ∂ √ √≥ √≥ εταλ ΙνσεχτΒιοχηεµ Μολ Βιολ 33 ∗≥∏ ∏ ≥ × Ηερεδιτψ 82 ° ∗2≥ ∏ ∂ ∏¬ ° ∏ Προχ Νατλ Αχαδ Σχι ΥΣΑ 88 ∗ ≤ ∂ × ƒ ∏ ≤ εταλ ϑ Μολ Βιολ 296 ∗∏ ≤ √ εταλ ϑ ΧελλΒιολ 154 ∗≤ • ⁄ ∏ ≥ ΦΕΒΣΛεττερσ 535 ∗ ∗。