5第五章 果蝇胚轴形成
03-6第三章 动物胚胎的早期发育6——果蝇胚轴形成

23
母源性Bicoid蛋白控制hunchback的表达
4
二、果蝇前-后轴的形成
到末端区域以外,保证tor基因只 在末端区域活化。
二、果蝇前-后轴的形成
¾ TOR与配体结合后,引起自身磷酸化,经一系列 信号传递,最终激活合子靶基因的表达。
¾ tor基因的活化可激活合子缺口基因hkb和tll在末 端区域的表达,这两个基因均编码转录调节因子, 进一步调节其他基因的表达。
35
36
6
末端系统:Torso信号途径
子的极性。 ¾ 调节果蝇胚胎前-后轴的形成有4个重要的形态发
生素: 9 Biciod(BCD)和Hunchback(HB)——前端 9 Nanos(NOS)和Caudal(CDL)——后端
14
二、果蝇前-后轴的形成
¾ 形态发生素调节首先表达合子基因——缺口基因 (gap gene),缺口基因表达区呈带状,带宽 约相当于3个体节的宽度,不同缺口基因的表达 区之间有部分重叠;
前端原头区和后端尾节的缺失。
32
二、果蝇前-后轴的形成
¾ 在末端系统中起关键作用的是torso(tor)基因。 9 失活突变体:缺失原
头区和尾节; 9 显性突变体:中部区
域(头、胸、腹)的 分节缺失,而末端结 构增大。
失活突变体
33
二、果蝇前-后轴的形成
¾ tor基因编码一种跨膜受体酪氨酸激酶(receptor tyrosine kinase,RTK),在整个合胞体胚胎的表 面表达,其NH2-位于细胞膜外,COOH-位于细胞膜 内。
果蝇的发育过程及其调控机制

果蝇的发育过程及其调控机制果蝇作为模式生物,其发育过程及调控机制已经被广泛研究。
果蝇的发育过程主要包括卵母形成、卵的受精、胚胎发育、幼虫期和蛹期等阶段。
这些阶段都受到不同的调控机制影响。
一、卵母形成和受精卵母形成过程在果蝇体内发生,一般从卵巢的端部开始,向基部发展。
在卵母形成早期,睾丸激素和卵泡刺激素对生殖细胞的发育起着重要作用。
随着卵母不断生长发育,卵母细胞核的复制和分裂也不断进行,最终形成完整的卵母。
成熟的卵母受到雄性精子的受精作用,形成受精卵。
在受精卵中,父本和母本细胞贡献的基因发挥不同的作用,影响胚胎的发育命运和表型表达。
二、胚胎发育受精卵受到一系列的调控因素作用,经过不同的胚胎发育阶段,最终形成成熟的幼虫体。
胚胎发育过程受到多个信号通路和基因网络的调控。
在果蝇的胚胎发育中,Wnt信号通路通过不同的基因调节细胞分裂和定向移动,对胚胎的头尾轴和背腹轴形成起着重要作用。
Hedgehog信号通路则调节胚胎前期的背腹轴形成。
同时,一个非常重要的因子是胰岛素样生长因子信号通路,其对幼虫的大小和发育有关键调节作用。
胚胎发育过程中也涉及到了一些胚胎基因调控系统,如TGFβ/Activin和Notch信号通路,调节着胚胎的细胞命运和分化。
三、幼虫期和蛹期在幼虫期和蛹期,果蝇表现出不同的形态和功能,同时也涉及到不同的代谢途径和调控机制。
在幼虫期,果蝇需要做出重要的生长和分化决策。
通过对食物和营养的感知和代谢途径的调控,果蝇可以根据生长的需要积累能量和物质,维持其生命活动的正常进行。
例如调节能量代谢的AMPK和Sirt1,以及调节食欲和代谢途径的Insulin/IGF1和TOR信号通路,都对幼虫的生长分化起着重要作用。
蛹期是果蝇生命周期中的一个转折点,幼虫期积累的营养和物质被重新配置为成虫身体的各种组织和器官。
在成虫器官分化和重构的过程中,也涉及到不同的调控机制。
例如调节上皮细胞形成和器官增殖的Notch信号通路,调节生殖腺发育的JAK/STAT信号通路,以及影响翅膀和大脑发育的Wnt和Hedgehog信号通路,都对蛹期的发育起着至关重要的作用。
果蝇体轴形成的分子机制果蝇的卵、胚胎、幼虫、成虫都有明确的前后和

果蝇体轴形成的分子机制果蝇的卵、胚胎、幼虫、成虫都有明确的前后和背腹轴,在果蝇最初的发育中,由母源效应基因及其编码蛋白构成位置信息的基本网络,激活合子基因的表达,控制果蝇躯体模式的建立。
1、果蝇胚胎的极性果蝇早期胚轴形成设计由母源效应产物构成的位置信息网络,其中有3组与前-后轴形成有关,为前端系统;1组决定胚胎的背腹轴,即后端系统;还有一组决定背腹轴形成。
2、果蝇前-后轴的形成在果蝇前-后轴的形成中,有四个非常重要的形态发生素:Biociod(BCD)和Hunchback(HB)调节胚胎前端结构的形成;Nanos(NOS)和Candal(CDL)调节胚胎后端结构的形成。
1)前端组织中心: BCD蛋白浓度梯度前端系统至少包括4个主要的基因,其中起关键作用的是BCD,bcd是一种母源效应基因,在卵子发生时,bcd mRNA于滋养细胞中转录,再转运至卵子中并定位于卵子前极。
受精后迅速翻译,BCD具有决定胚胎极性和组织空间图示的功能。
受精后BCD蛋白在前端积累并向后端弥散,形成从前向后稳定的浓度梯度,主要覆盖胚胎前2/3区域。
bcd 基因也是同源异型框基因,BCD蛋白是一种转录调节因子,可与DNA特异性结合并激活合子靶基因的表达。
BCD蛋白浓度梯度可以同时特意新启动不同基因的表达,从而将胚胎划分为不同的区域。
2)后端组织中心:Nanos蛋白和Candal蛋白浓度梯度后端系统在控制图式形成中起到作用与前端系统相似。
决定胚胎后端的最初信息也是母源效应基因转录产物,在卵子发生过程中,后端决定子Nanos(NOS)的mRNA在卵室前端的滋养细胞中转录,通过转运定位到卵子后极,在成熟卵中定位于生殖质。
后端系统是通过抑制转录因子HB的翻译起作用,该系统包括约10个基因,这些基因都是腹部图示形成所必须的基因。
NOS活性从后端向前弥散形成浓度梯度,在胚胎后部抑制hb mRNA的翻译。
HB蛋白的分布区域主要位于胚胎前半部分。
果蝇胚胎发育和体轴形成的分子机制

果蝇胚胎发育和体轴形成的分子机制果蝇是一种十分常见的昆虫,它的胚胎期相当短暂,只需要不到两天的时间就能完成发育。
在这个过程中,许多重要的分子和信号通路都在起作用,对于了解发育和形态建成的分子机制具有重要意义。
本文将着重讨论果蝇胚胎发育和体轴形成的分子机制。
一、胚胎发育的基本过程果蝇的生命周期很短,在一般的实验室条件下,一只果蝇的寿命约为两个月。
雌性果蝇通常会在食物和水的混合物中产卵,大约在24小时内就会孵化。
在这之后的几天中,果蝇的发育非常迅速,逐渐从卵到幼虫,然后到成虫。
而在这个发育过程中,果蝇胚胎发育是非常关键的一步。
胚胎发育的过程可以分为五个连续的阶段。
第一阶段为卵细胞核形成,包括以交配卵产生的核和父母亲体细胞内孤雌生殖卵产生的核。
第二阶段是卵剖面形成,包括老二系统等。
通过定期锯切进行紧密环绕,则表明胚胎发展正常。
长时间僵滞不进则可能会在胚胎发展的后期出现显著的异常。
第三阶段是胚胎心脏形成,包括神经脑、背板神经织物、翼的最初生长和原胚内的胚芽形成。
第四阶段胚胎形成,则大半是身体器官、肌肉、皮肤等的形成。
卵子在此时分化为头部,胸部和腹部。
第五阶段是成虫形成,也就是最后的阶段。
它涉及更广泛的组织分化,包括瓢虫和蜜蜂等其他昆虫几乎都会经历的阶段。
以上这些阶段综合起来,构成了果蝇胚胎发育过程中的基本过程。
接下来我们将要探究果蝇胚胎形态的建立和维持所涉及的分子机制。
二、体轴形成的分子机制在果蝇发育的过程当中,体轴形成似乎是整个过程中最显著的阶段之一。
而体轴形成涉及到许多基因和信号通路的调控。
1. Wnt信号通路Wnt信号通路是胚胎发育中非常重要的一个通路。
Wnt分子在体轴形成中具有特殊作用,因为它们能够影响Dorsal基因的表达。
Dorsal基因在果蝇胚胎发育中发挥着严格的调控作用,在形成体胚层时非常重要。
Wnt拮抗剂能够使得胚胎中Dorsal基因表达出现异常,从而导致体轴形成不良。
2. Hedgehog信号通路Hedgehog信号通路也是在胚胎发育中重要的信号通路。
果蝇的胚胎发育与影响因素

生命科学学院学院20 -20 学年第 学期《 发育生物学 》课程论文 课程号:2522080任课教师 成绩 论文题目:(可指定题目,也可说明题目范围。
)果蝇的发育及其发育的影响因素论文要求:(对论文题目、内容、行文、字数等作出判分规定。
) 1. 论文题目:准确得体,简短精炼,醒目2. 摘要:文字简练,字数不超过正文的5%;关键词不少于三个,关键词之间用分号间隔3. 正文:内容充实,论据充分、可靠,论证有力,主题明确语言流畅,条理清晰,字数不少于3000字4.字体:摘要、关键词宋体5号字;题目黑体三号字;正文宋体四号字 10分 教师评语:教师签字:年 月 日果蝇的发育及其发育的影响因素摘要:果蝇(Drosophila melanogaster)由于具有容易饲养、产卵多、生命周期短以及具有粗大的多线染色体而便于进行基因定位等优点,已成为遗传学和发育生物学等研究领域中重要的模式生物之一。
本文主要介绍了果蝇的发育过程及樟树叶片挥发物,紫外线照射,酵母粉和甲醛对果蝇生长发育的影响。
Abstract : Drosophila melanogaster is easy feeding, spaen many , shorter lifecycle and has athick multi-line chromosome.It has become such an important model organisms of Genetics and Developmental Biology field. This article mainly introduced the growth process of Drosophila melanogaster and some influence factor, such as essential oil produced by the leaves of camphor, ultraviolet rays, yeast powder and Formaldehyde. 关键词:果蝇;发育;樟树;紫外线;酵母粉;甲醛一、果蝇简介果蝇易于培养,生活周期短,基因组也比较小(1.4×108bp/每个单倍体基因组),只有4对染色体,且存在多线染色体,有关果蝇的遗传学知识已积累了80多年,遗传背景十分清楚,这些都是其重要的优势,故人们选择果蝇作为研究发育的模式动物。
果蝇卵子发生

末端系统: TORSO信号途径在前端和后端系统失活时,果蝇胚胎仍可产生某些前后模式,形成具有两个尾节的胚胎。
说明存在第三个与前后轴建立有关的系统。
现证明其中包括9个母体效应基因。
如果这个系统失活导致胚胎不分节的部分消失,即缺少原头区和尾节。
该系统起主要作用的是torso(tor)基因。
它有两类突变体,一类失活突变体,胚胎缺少原头区和尾节;另一类为显性突变体,胚胎中部分节消失,末端结构增大。
TOR蛋白在整个合胞体胚胎表面表达(下图),其NH2-端序列位于膜外,COOH-端位于膜内。
当胚胎前后端细胞外存在某种信号分子(配体)时,可使TOR特异性活化而导致胚胎前后端细胞命运的特化。
tor基因编码一种跨膜酪氨酸激酶受体(PTK)。
PTK的活化需要一种配体与其受体部位结合,可能torso-like(tsl)基因编码这个配体。
tsl基因在两组特异滤泡细胞中表达,TSL蛋白被分泌到两极的卵周隙中,可能通过与卵黄膜结合固定于两极,直到TOR蛋白表达它才释放。
Tsl基因的异位表达引起胚胎末端膨大。
除TSL外,末端途径其余成分在胚胎中均匀分布,TOR在整个合胞体表面表达,为跨膜蛋白(NH2-端位于膜外,COO-端膜内)TSL为配体。
在卵子发生的过程中,tsl基因在两组特异性滤泡细胞中(卵子前极的边缘细胞和卵室后端与卵子后极相对的极性滤泡细胞)表达表达。
TSL蛋白被分泌到卵子两极处的围卵隙中,与卵黄膜结合而维系在两极区域(配体两极局部定位)。
TOR蛋白表达,TSL释放。
由于TOR表达过量,TSL不会扩散到末端区域外,保证TOR 蛋白只在两极被活化。
TOR蛋白表达,TSL释放。
由于TOR表达过量,TSL不会扩散到末端区域外,保证TOR 蛋白只在两极被活化。
TOR被活化后,再经过一系列信号传导,最后激活合子靶基因的表达。
卵子发生过程:tsl在两组细胞(前极边缘、后端)中表达——通过卵黄膜维系位置——TOR 蛋白表达时释放——活化表达。
果蝇胚胎发育与前-后轴

卵表面 纺锤体 分裂沟 星状体 细胞核 沟渠 微管
卵黄膜
表面卵裂
动画果蝇早期发育,自卵中央行核有丝分裂(无膜), 核边移,细胞膜形成。
3、时间
• 核到达外围后,分裂所需的时间不断延长。第1-10个周期 平均每周期8min,第13周期(合胞体胚层的最后一个周 期)需要25min完成,第14次分裂才形成细胞。第14次分 裂是不同步的,有的细胞需要75分钟才能完成分裂,有的 细胞需要175分钟。
后轴和背-腹轴的细胞类型特化(the specification of cell types)
都是在一个多核细胞当中完成的。而且,前-后轴和背-腹轴的形 成是由卵细胞在母体当中的位置决定的,卵细胞和它周围的滤 泡细胞细胞共同作用所决定行成。海鞘和线虫当中,轴是由精 子进入的位点所决定。
During Drosophila development, however, cellular membranes do not form until after the thirteeth nuclear division. Prior to this time, all the nuclei share a common cytoplasm,and material can diffuse throughout the embryo.
Moreover, the initiation of the anterior-posterior and dorsalventral differences is controlled by the position of the egg within the mother's ovary. Whereas the sperm entry site may fix the axes in ascidians and nematodes, the fly's anteriorposterior and dorsal-ventral axes are specified by interactions between the egg and its surrounding follicle cells.
5第五章果蝇胚轴形成

35
在果蝇中,大多数同源异型基因是在第3号染色 体上,排成两簇。一簇叫触角足复合体(AntpC),另一簇叫双胸复合体(BX-C),这两簇基因 一起构成同源异型复合体(HOM-C), HOM-C 基因各自发挥独特的功能。
除pb基因外,HOM-C基因在染色体上的前后 排列顺序与早期胚胎体轴的头尾顺序和发育次 序一致,表达早的基因对应于胚胎的头部基因, 表达晚的基因对应于胚胎的尾部。
36
基因的突变会引起同源异形转化现象。
37
同源异型基因的表达图式受缺口基因和成 对控制基因的调控。
14
末端系统:Torso信号途径
◆Torso 是决定前后轴两个端部的核心 蛋白。torso突变体缺少前端原头区和 后端尾节区。
◆ Torso是受体酪氨酸激酶(RTK),未受 精前,Torso已均匀分布在卵膜上; 受精后,两极的Torso被配体结合而 活化,开启端部系统发育的信号传导。
◆其配体可能是torsolike (tsl) 基因的产 物。tsl基因只定位于卵子前极的边缘 细胞和卵室后端的极性滤泡细胞中。 TSL蛋白被分泌到卵子两极处的卵周 隙(与torso的活化位置相同),可能通 过与卵黄膜的结合而维系在两极局域。
第五章 果蝇胚轴形成 (Embryonic
axes)
1
图式形成(Pattern formation):
动物发育中,胚胎不但要产生不同类别的细 胞,而且要由这些细胞构成功能性的织织和器官 并形成有序空间结构的形体模式(body plan)。
胚胎细胞形成不同组织、器官、构成有序空 间结构的过程称为图式形成。在动物胚胎发育中 最初的图式形成主要涉及胚轴(embryonic axis)形 成、体节形成(segmentation)、肢芽和器官原基 (anlage)及其一系列相关的细胞分化过程。
果蝇的胚胎发育(PPT文档)

3. 后端组织中心:
NANOS蛋白和CAUDAL蛋白浓度梯度
后端系统包括约10个基因,这些基因的突 变都会导致胚胎腹部的缺失。在这一系统
缺口基因、成对控制基因以及体节极性基 因共同调节同源异型基因(homeotic gene) 的表达,决定每个体节的发育命运。
2. 前端组织中心
BICOID(BCD)蛋白浓度梯度
前端系统至少包括4个主要基因,其中bicoid
(bcd)基因对于前端结构的决定起关键的作
用。BCD具有组织和决定胚胎极性与空间图 式的功能。
如果前端和后端系统都失活,果蝇胚胎仍 可产生某些前后图式,形成具有两个尾节 的胚胎。
Torso系统基因的失活会导致胚胎不分节的部分,即前 端原头区和后端尾节,缺失
三、果蝇背 – 腹轴的形成
与果蝇胚轴形成有关的4组母体效应基因中, 背–腹系统最为复杂,涉及约20个基因。其
中dorsal(dl)等基因的突变会导致胚胎背
现已筛选到与胚胎前后轴和背腹轴形成有 关的约50个母体效应基因(maternal effect gene)和120个合子基因(zygotic gene)。 目前,对果蝇胚轴形成的调控机制已有了 一个较为清晰的认识。
在果蝇最初的发育中,由母体效应基因构 建位置信息的基本网络,激活合子基因的 表达,控制果蝇形体模式的建立。
在动物胚胎发育中,最初的图式形成主要 涉及胚轴(embryonic axes)形成及其一系 列相关的细胞分化过程。胚轴指胚胎的前 -后轴(anterior -posterior axes)和背 – 腹 轴(dorsal -ventral axis)。
果蝇前—后轴的形成PPT文档共34页

16、业余生活要有意义,不要越轨。——华盛顿 17、一个人即使已登上顶峰,也仍要自强不息。——罗素·贝克 18、最大的挑战和突破在于用人,而用人最大的突破在于信任人。——马云 19、自己活着,就是为了使别人过得更美好。——雷锋 20、要掌握书,莫被书掌握;要为生而读,莫为读而生。——布尔沃
END
果蝇前—后轴的形成
31、别人笑我太疯癫,我笑他人看不 穿。(名 言网) 32、我不想听失意者的哭泣,抱怨者 的牢骚 ,这是 羊群中 的瘟疫 ,我不 能被它 传染。 我要尽 量避免 绝望, 辛勤耕 耘,忍 受苦楚 。我一 试再试 ,争取 每天的 成功, 避免以 失败收 常在别 人停滞 不前时 ,我继 续拼搏 。
33、如果惧怕前面跌宕的山岩,生命 就永远 只能是 死水一 潭。 34、当你眼泪忍不住要流出来的时候 ,睁大 眼睛, 千万别 眨眼!你会看到 世界由 清晰变 模糊的 全过程 ,心会 在你泪 水落下 的那一 刻变得 清澈明 晰。盐 。注定 要融化 的,也 许是用 眼泪的 方式。
35、不要以为自己成功一次就可以了 ,也不 要以为 过去的 光荣可 以被永 远肯定 。
果蝇的发育和成虫盘解剖
实验一果蝇的发育和成虫盘解剖1、果蝇胚胎发育和成虫盘自1910年遗传学鼻祖Morgan发现第一个突变体白眼果蝇以来,生物学家将果蝇作为遗传学研究的模式动物已经近百年,并广泛应用于现代遗传学和发育生物学研究的模型。
果蝇生命周期如图1。
果蝇胚胎发育速度快,前13次卵裂每次只间隔9min,细胞核成倍增加成为一个合胞体(syncytim),发育过程中的胚胎为观察分析卵裂、早期胚胎发生和躯体模式形成等发育调控机制的提供了很好的材料。
果蝇胚胎发育的梯度假说被证实,在1997年和1998年连续两年被《Science》杂志评为当年十大突破成就之一。
现已鉴定出在一些卵子中形成梯度、调节细胞定位和分化并决定胚胎发育方式的形态发生素(morphogen)。
果蝇存在变态过程,由幼虫(larval)经蛹(pupal)转变为成虫。
成虫盘(imaginal discs)是果蝇早期胚胎发育期间保留下来的而在幼虫期开始分化的。
变态期间,幼虫表皮转成蛹壳而成虫盘则发育成为成虫外层。
成虫盘首先出现在20~40细胞群的胚胎中,由胚胎上皮内陷形成,最初通过各种标记基因的表达而被确认。
每一个成虫盘都由单层上皮构成片状或囊状,并有一个细茎与胚胎和幼虫的上皮连接。
最终每一个液囊展平,液囊的两端具有不同的特征,并执行不同的发育任务。
一端形成较厚而高度折叠的成虫盘上皮,另一端形成较薄而展开的周膜。
大部分成虫外胚层结构由成虫盘上皮演化而来。
成虫盘在一、二、三龄幼虫期都有不同程度的细胞增殖,在晚期三龄幼虫后,即变态之前,每个成虫盘形成了数万个细胞。
在变态之前,大约产卵后108h(AEL),25℃放置,三龄幼虫即蠕动——“爬”出培养基,生活在培养瓶的瓶壁上,这个阶段约12h,在这个阶段末期,果蝇幼虫前端的呼吸孔外翻,幼虫停止蠕动并开始蛹化。
上皮形成蛹的桶状体壁——蛹壳,此时为白色的蛹前期阶段(WPP)。
大约1h后,蛹壳变为深褐色。
蛹化开始的第一个5-6h后,果蝇仍然处于蛹前期阶段,成虫细胞仍然分泌形成蛹的表皮。
果蝇胚轴的形成

分节基因与果蝇胚胎体节的形成
分节基因的功能是把早期胚胎沿前—后轴分为一 系列重复的体节原基。分节基因的突变可使胚胎 缺失某些体节或体节的某些部分。 分节基因的可分为三类:缺口基因、成对控制基 因和体节极性基因。 这三类基因的调控是逐级进行的,首先由母体效 应基因控制缺口基因的活化,其次缺口基因之间 互相调节彼此的转录并且共同调节成对控制基因 的表达,然后成对控制基因之间相互作用把胚体 分隔成为一系列重复的体节,并且进一步控制体 节极性基因的表达。缺口基因和成对控制基因再 共同调控同源异型基因的表达。
节的区域,后端系统 (posterior system)
决定分节的腹部,末端系统 (terminal
system) 决定胚胎两端不分节的原头区和
尾节。另一组基因决定胚胎的背 — 腹轴, 即背腹系统 (dorsoventral system) 。
果蝇前 — 后轴的形成
A-P轴线由三类母体效应基因控制: 突变鉴定
果蝇胚胎的极性
胚胎躯体轴线的建立首先由母体效应基因决定
卵子发生中,母体效应基因的mRNA由滋养细胞 合成后迁移进卵子,分别定位于一定区域。这些 mRNA编码转录因子或翻译调控蛋白因子,它们 在受精后立即翻译且分布于合胞体胚盘中,激活 或抑制一些合子基因的表达,调控果蝇胚轴的形 成。 这些母体效应基因的蛋白质产物又称为形态发生 素(morphogen)
红色显示突变影响 的部位
Nature 424:935-938
Bicoid基因提供A-P轴线形态发生素梯度
为什么不 能完全恢 复?
Bicoid mRNA和蛋白质的分布
Bicoid是控制头胸发育 的一个关键母体效应基 因,其不同浓度开启不 同合子基因的表达。 在未受精卵中,bicoid mRNA定位在胞质前端; 其受精后翻译出的蛋白 质沿AP轴扩散,形成浓 度梯度,为胚胎的后续 分化提供位置信息。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
果蝇形体模式的分子机制:发育生物学研究的重大成
果之一
2
胚轴指胚胎的前后轴(anterior - posterior axis) 和背腹轴(dorsal- ventral axis)。所有多细胞机体至 少具有一种主要的胚轴。两侧对称的动物还具有中 侧轴(mediolateral axis)或左右轴(left-right axis)。
具有正常形态特征的一类关系密切的基因。这些基因 都含有一段高度保守的180 bp 的DNA片段,该片段称 为同源异型框 (homebox)。
功能:它将决定产生哪一类体节(前胸部、中胸部或 后胸部)。在每一个体节确定后,调控进一步的结构 特化。
35
在果蝇中,大多数同源异型基因是在第3号染色 体上,排成两簇。一簇叫触角足复合体(Antp-
22
表达特点:
(1). 都编码转录因子;
(2). 都在合胞胚期开始 表达; (3). 其产物的半衰期一 般较短,仅数分钟,因 而它们的扩散距离较短; (4). 其表达局限在一定 的区域,其突变会导致 胚胎在该区域及附近区 域的缺失。
lose of Kruppel
23
功能:
1. 彼此调节确定表达界限;
末端系统:Torso信号途径,决定胚胎两端不分节的原头区和尾 节
背腹轴
TOLL降解CS :Dorsal(dl)-DL在细胞核中
腹部
DL和CS结合,Toll无活性: Dorsal(dl)-DL在细胞质中
背部:
二、分节基因和胚胎体节的形成(研究 缺失突变体):
缺口基因
成对控制基因
体节极性基因
同源异型基因
13
NOS 蛋白和CDL 蛋白浓度控制后端区结构:
nanos mRNA 定位在卵子后极,受精后翻译, NOS蛋白形成从后到前的浓度梯度。这种浓度 梯度为胚胎的后端分化提供位置信息。 NOS 蛋白是翻译抑制子,不直接调节缺口基因 的表达。 母源型hunchback mRNA 在卵子中均匀分布; 受精后, NOS蛋白抑制母源型hunchback mRNA在后端区的翻译,使HB蛋白只在前端 区高浓度表达。
26
2. 成对控制基因(Pair-rule gene)
成对控制基因的表达区域以两个体节为单位, 具有周期性,在相互间隔的一个副体节中表 达。成对控制基因的表达是胚胎出现分节的 最早标志,并确定副体节的界限。
副体节(parasegments) :
在体节沟真正形成之前,基因表达带并不是刚好 与未来的体节吻合,而是逆着可见体节向前移动 半个体节。果蝇学家将其称之为副体节。一个副 体节包括一个未来体节的后半部和后面相邻体节 的前半部分。
由单一的浓度梯度来控制,而是由多个 转录因子控制。
32
3. 体节极性基因 (segment polarity gene)
体节极性基因产生每一副体节内部的发育图式。 主要有engrailed (en)、wingless(wg)、hedgehog(hh) 及patched等。 表达特点:
在成对控制基因之后表达;
of seven strips);
eve在奇数副体节表达,ftz在偶数副体节表
达 同一副体节中的每一排细胞可能表达不同 的成对控制基因
主要为转录因子; 合胞体胚胎期表达。
30
成对基因的缺失
31
功能:
建立副体节的边界;
成对控制基因的每个表达条纹由一组缺
口基因控制。
成对控制基因的间隔性重复表达,无法
9
影响背腹极性的基因:
2. 果蝇前—后轴的产生:
存在于卵质中调节果蝇胚胎前-后轴的形成有4个 非常重要的形态发生素(morphogen):Bicoid (BCD)、 Hunchback (HB)、Nanos (NOS)和Caudal (CDL); Bicoid (BCD)和Hunchback (HB)控制胚胎前端结构 的形成,决定头胸部分节的区域. Nanos (NOS)和 Caudal (CDL)调节胚胎后端结构 (腹部)的形成。决定分节的腹部 末端系统:Torso信号途径,决定胚胎两端不分节 的原头区和尾节
37
同源异型基因的表达图式受缺口基因和成 对控制基因的调控。 HOM-C基因的表达区域多相互重叠,表 达区域靠后的基因可部分和全部抑制表达 区域靠前的基因。
通过突变鉴 定确立影响 果蝇前后轴 的三类基因, 即前端系统、 后端系统和 末端系统。
bicoid mutant
nanos mutant
torso mutant
8
影响前后极性的基因:
以bicoid (BCD) 和hunchback(HB)为核心的 前端系统,调节胚胎头胸结构的形成; 以 nanos (NOS) 和caudal(CDL)为核心的后 端系统,调节胚胎腹部结构的形成; 以torso为核心的末端系统,调节胚胎前端原 头区和后端尾节区的发育; gurken、dorsal 和 toll
DL是一种转录因子。 DL蛋白的核内浓度梯度成 为背腹轴产生区域特异性的位置信息。
18
小结: 前后轴
Bicoid (BCD)和Hunchback (HB)控制胚胎前端结构的形成,决定 头胸部分节的区域.
Nanos (NOS)和 Caudal (CDL)调节胚胎后端结构(腹部)的形成。 决定分节的腹部
10
BCD是母体效应因子;bicoid 是母性极性基因 (转录因子)
11
前端bicoid 控制基因hunchback, caudal 的表达
12
Bicoid(BCD)蛋白和 HB 蛋白控制前部区结构:
biciod 是母体效应基因,mRNA 定位于卵子前极,受 精后翻译,BCD蛋白形成从前到后的浓度梯度。这种 浓度梯度为胚胎的前部区分化提供位置信息。
21
1. 缺口基因 (gap gene)
缺口基因是指那些在受精后最 早沿AP轴线呈区域性表达的 合子基因,缺口基因的表达使 胚胎沿AP轴线形成一些较宽 的区域,每个区域约相当于3 个体节,表达之间可有部分重 叠。 主要有hb、 giant 、Krüppel (kr)、Knirps (kni) 和tailless
BCD蛋白是一种转录调节因子。受精后,前部区高浓 度的BCD蛋白激活合子hunchback基因的表达。
HB蛋白在前部区形成高浓度表达。 hunchback基因的 表达受到BCD蛋白浓度梯度的控制,只有BCD蛋白浓
度达到一定临界值才能启动hunchback基因的转录。
母源型 caudal mRNA在卵中均匀分布,受精后BCD蛋白抑 制母源型 caudal mRNA在前区的表达,因而CDL蛋白形成 从后到前的浓度梯度。
14条纹,在每一副体节的特定区域细胞中表达, 发生 突变时,幼虫的每个体节就会缺失一个特定的区域。
表达持续时间长,有些维持一生。
产物为分泌蛋白、受体、细胞内信号及转录因子 。
33
功能:
建立并维持副体节/体节的界限
en在每一副体节最前端的一列细胞中表达,wg基 因的表达区域刚好位于en表达带之前,即每一副 体节的最后一列细胞。因此,这两个基因表达区 域的界线正好确定副体节的界线。
2. 调节成对控制基因和体节极性基因的
表达,建立基本体节模式;
3. 调节同源异型基因的表达,确定体节
命运;
24
缺口基因的表达调控:
其表达受母源bicoid基因产物的调节; 且呈浓度依赖性; 相互调节(主要为抑制),确定表达区域
(界限)。
25
The Nobel Prize in Physiology or Medicine 1995
27
果蝇胚胎 的体节和 副体节
未来的体节 副体节
28
果蝇胚胎的体节和副体节
体节
副体节
副体节仅在原肠作用后一个短暂的时期内能观察到
这些区域通常与基因活性(paired-rule gene)区域一致
29
初级成对控制基因的表达直接受缺口基因的 调控;
表达特点:
相间的7条条纹(patterns
建立体节内部细胞发育命运的差异
体节出现后,en表达于每一体节的最后,把体节 分为前后两个区域,两个区域内的细胞不再交换, 每个区域有各自的发育命运。
34
4. 同源异型基因 (homeotic gene)
同源异型基因最终决定身体体节的发育类别,为体节 决定基因。
同源异型基因是指控制动物体节发育前后次序使动物
15
A-P轴形成总结:
蓝色表示受精前
红色表示受精后
16
3. 果蝇背—腹轴的形成:
(1) 背腹系统:
Dorsal(dl)-DL在细胞核中
Dorsal(dl)-DL在细胞质中
腹部;
背部;
dl mRNA和DL蛋白在卵子中均匀分布。第9 次细胞核分裂之后,胚胎细胞核迁移到达合 胞体胚盘的外周皮质层,在胚胎腹侧DL蛋白 开始往核内聚集,而背侧DL蛋白仍位于胞质 中,从而使DL蛋白在细胞核内的分布沿背- 腹轴形成一种浓度梯度。
3
一、果蝇胚轴形成
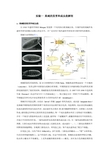
果蝇胚胎沿前- 后轴分为头节、 3个胸节和8个腹 节,在幼虫的两 末端又特化为前 面的原头和后端 的原尾。沿背- 腹轴分化为4个 区域,分别为背 侧外胚层、腹侧 外胚层、中胚层 和羊浆膜。
4
羊浆膜
原头
原尾