C3、C4和CAM植物的光合特性比较..
东北林业大学植物生理学思考题及答案

植物生理学思考题及答案第一章1.植物细胞和土壤溶液水势的组成有何异同点?答案:2一个细胞放在纯水中水势和体积如何变化?答案:水势升高,体积变大。
3植物体内水分的存在形式及其与植物代谢强度、抗逆性有何关系?答案:存在形式:束缚水,自由水;与植物代谢强度、抗逆性关系:自由水与束缚水比值较高时,植物代谢活跃,但抗逆性差;反之,代谢活性低,但抗逆性较强。
4.气孔运动的机制及其影响因素。
答案:机制:淀粉-糖转化学说,无机离子吸收学说,苹果酸代谢学说;影响因素:凡能影响光合作用和叶子水分状况的各种因素:①光照(主要因素)②温度③二氧化碳(影响显著)④叶片含水量。
5水分进出植物体的途径及动力。
答案:途径:质外体途径,跨膜途径,共质体途径;动力:①上端原动力—蒸腾拉力;②下端原动力—根压;③中间原动力—水分子间的内聚力及导管壁附着力。
6.如何区别主动吸水与被动吸水?答案:第二章1.溶液培养法有哪些类型?用溶液培养植物时应注意的事项?答案:s2如何确定植物必需的矿质元素?植物必须的矿质元素有哪些生理作用?答案:植物必须元素的三个标准:①由于该元素缺乏,植物生育发生障碍,不能完成生活史;②去除该元素则表现出专一缺乏症,且这种缺乏症可以预防恢复;③该元素在植物营养生理上应表现直接的效果,绝不是因土壤或培养基的物理化学微生物条件的改变而产生的间接效果。
生理作用:①是细胞结构物质的组成成分;②是植物生命活动的调节者,参与酶的活动;③起电化学作用,即离子浓度的平衡,胶体的稳定和电荷中和。
3植物细胞通过哪些方式吸收矿质元素?答案:离子通道运输,载体运输,离子泵运输,胞饮作用。
4.试述植物从土壤中吸收的硝酸盐是如何进行还原和氨基酸的同化?答案:硝酸盐的还原:①硝酸盐还原成亚硝酸盐的过程是由细胞质中的硝酸还原酶催化的。
硝酸盐还原的步骤:②亚硝酸盐还原成氨是由叶绿体中的亚硝酸还原酸催化的,其酶促过程如下式:氨基酸的同化:谷氨酸脱氢酶途径,氨基交换作用,酰胺合成酶途径。
光合作用—C3、C4、CAM区别

光合作用光合作用可分为:光反应和碳反应(旧称暗反应)两个阶段。
光反应条件:光照、光合色素、光反应酶。
场所:叶绿体的类囊体薄膜。
(蓝细菌等微生物的反应场所在细胞膜:色素所在地)过程:①水的光解:2H2O→4[H]+O2(在光和叶绿体中的色素的催化下)。
②ATP的合成:ADP+Pi+能量→ATP(在酶的催化下)。
影响因素:光照强度、CO2浓度、水分供给、温度、酸碱度、矿质元素等。
意义:①光解水,产生氧气。
②将光能转变成化学能,产生ATP,为碳反应提供能量。
③利用水光解的产物氢离子,合成NADPH(还原型辅酶Ⅱ),为碳反应提供还原剂NADPH(还原型辅酶Ⅱ)。
碳反应条件:多种酶。
场所:叶绿体基质。
过程:①碳的固定:C5+CO2→2C3(在酶的催化下)②C3+[H]→(CH2O)+C5(在ATP供能和酶的催化下)影响因素:温度、CO2浓度㊣卡尔文循环(Calvin Cycle)可分为三个阶段: 羧化(碳的固定)、还原(3-磷酸甘油醛(G3P(PGAL))的合成)、二磷酸核酮糖的再生。
卡尔文循环,又称还原磷酸戊糖循环(以对应呼吸作用中的氧化磷酸戊糖途径)、C3循环(CO2固定的第一产物是三碳化合物)、光合碳还原,还是光合作用的暗反应的一部分。
反应场所为叶绿体内的基质。
㊣固碳作用:※碳三植物(小麦、水稻、大豆、棉花等)二氧化碳经气孔进入叶片后,直接进入叶肉进行卡尔文循环。
而C3植物的维管束鞘细胞很小,不含或含很少叶绿体,卡尔文循环不在这里发生。
※碳四植物(玉米、甘蔗、高粱、苋菜等)碳四植物维管束周围有两种不同类型的细胞:靠近维管束的内层细胞称为鞘细胞,围绕着鞘细胞的外层细胞是叶肉细胞。
①叶肉细胞里的磷酸烯醇式丙酮酸(PEP)经PEP羧化酶的作用,与CO2结合,形成苹果酸或天门冬氨酸。
②这些四碳双羧酸转移到鞘细胞里,通过脱羧酶的作用释放CO2,后者在鞘细胞叶绿体内经核酮糖二磷酸(RuBP)羧化酶作用,进入光合碳循环(卡尔文循环。
C4植物及C3植物的光合作用曲线比较

C4植物与C3植物的光合作用曲线比较1、光合作用强弱变化的指标光合作用强弱变化的指标通常是光合速率和光合生产率光合速率是指单位时间、单位叶面积吸收CO2的量或放出O2的量或有机物的消耗量。
一般测定光合速率的方法都没有把叶片的呼吸作用考虑在内,所以测定的结果实际是光合作用减去呼吸作用的差数,称为表观光合速率或净光合速率。
如果把表观光合速率加上呼吸速率,则得到总(真正)光合速率。
光合生产率又称净同化率,是指植物在较长时间(一昼夜或一周)内,单位叶面积生产的干物质量。
光合生产率比光合速率低,因为已去掉呼吸等消耗。
2、影响光合作用的因素内因1)叶龄:叶片的光合速率与叶龄密切相关。
从叶片发生到衰老凋萎,其光合速率呈单峰曲线变化。
新形成的嫩叶由于组织发育不健全、叶绿体片层结构不发达、光合色素含量少、光合酶含量少、活性弱、气孔开度低、细胞间隙小、呼吸细胞旺盛等原因,净光合速率很低,需要从其它功能叶片输入同化物。
随着叶片的成长,光合速率不断提高。
当叶片伸展至叶面积最大和叶厚度最大时,光合速率达最大值。
通常将叶片充分展开后光合速率维持较高水平的时期,称为叶片功能期,处于功能期的叶叫功能叶。
功能期过后,随着叶片衰老,光合速率下降2)光合产物的运输:光合产物从叶片中输出的快慢影响叶片的光合速率。
例如,摘去花或果实使光合产物的输出受阻,叶片的光合速率就随之降低。
反之,摘除其他叶片,只留一个叶片和所有花果,留下叶片的光合速率就会增加。
如对苹果枝条进行环割,光合产物会积累,则叶片光合速率明显下降。
叶肉细胞中蔗糖的积累会促进叶绿体基质中的淀粉合成和淀粉粒形成,过多的淀粉粒一方面会压迫和损伤叶绿体,另一方面,由于淀粉粒对光有遮挡,从而阻碍光合膜对光的吸收。
(1)光照:光是光合作用的能量来源,是形成叶绿素的必要条件。
此外,光还调节着光合酶的活性和气孔开度,因此光是影响光合作用的重要因素1)光强:在暗中叶片无光合作用,只进行细胞呼吸释放CO2。
C3植物、C4植物与CAM植物

C3植物、C4植物与CAM植物标题: C3植物、C4植物与CAM植物摘要: [C3植物、C4植物与CAM植物]在高等植物中,光合碳同化主要有3种类型:C3途径,C4途径和景天酸代谢途径(CAM)。
C3植物中,CO2的固定主要取决于1,5-二磷酸核酮糖羧化酶(RuBPCase)的活化状态,因为该酶是光合碳循环的入口钥匙。
它催化1,5-二磷酸核酮糖(RuBP)羧化,将大气中的CO2同化,产生两分子磷酸甘油酸,可见RuBPCase在C3植物中同化CO2的重要性。
C4植物是从C3植物进化而来的一种高光效种类。
与C3植物相比,它具有在高光强……[关键词:叶绿体活性基因外稃细胞质叶片大豆]……关键词: 叶绿体活性基因外稃细胞质叶片大豆在高等植物中,光合碳同化主要有3种类型:C3途径,C4途径和景天酸代谢途径(CAM)。
C3植物中,CO2的固定主要取决于1,5-二磷酸核酮糖羧化酶(RuBPCase)的活化状态,因为该酶是光合碳循环的入口钥匙。
它催化1,5-二磷酸核酮糖(RuBP)羧化,将大气中的CO2同化,产生两分子磷酸甘油酸,可见RuBPCase在C3植物中同化CO2的重要性。
C4植物是从C3植物进化而来的一种高光效种类。
与C3植物相比,它具有在高光强,高温及低CO2浓度下,保持高光效的能力。
C4植物固定CO2的酶为磷酸烯醇式丙酮酸羧化酶(PEPCase),与C3作物中RuBPCase相比,PEPCase对CO2的亲和力高。
C4植物的细胞分化为叶肉细胞和鞘细胞,而光合酶在两类细胞中的分布不同,如PEPCase在叶肉细胞固定CO2,生成草酰乙酸(OAA),OAA进一步转化为苹果酸(Mal),Mal进入鞘细胞,脱羧,被位于鞘细胞内的RuBPCase羧化,重新进入卡尔文循环。
这种CO2的浓缩机理导致了鞘细胞内的高浓度的CO2,一方面提高RuBPCase的羧化能力,另一方面又大大抑制了RuBPCase的加氧活性,降低了光呼吸,从而使C4植物保持高的光合效率。
C3C4CAM植物光合作用之比较
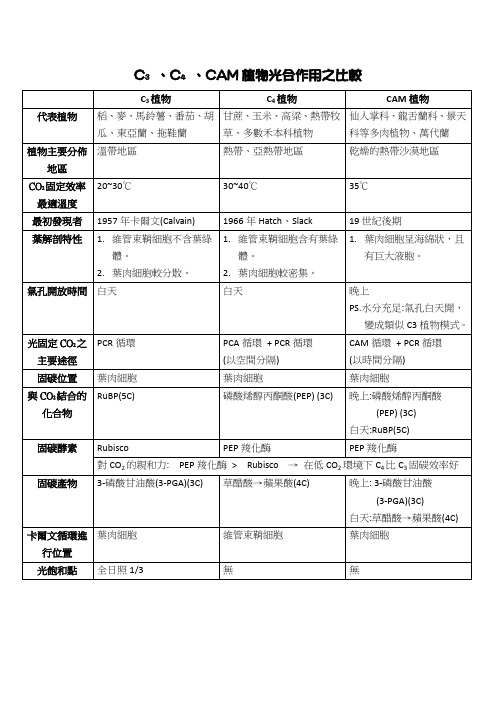
C3 植物 代表植物 瓜、東亞蘭、拖鞋蘭 植物主要分佈 溫帶地區 地區 CO2 固定效率 最適溫度 最初發現者 葉解剖特性 1957 年卡爾文(Calvain) 1. 維管束鞘細胞不含葉綠 體。 2. 葉肉細胞較分散。 氣孔開放時間 白天 1966 年 Hatch、Slack 1. 維管束鞘細胞含有葉綠 體。 2. 葉肉細胞較密集。 白天 晚上 PS.水分充足:氣孔白天開, 變成類似 C3 植物模式。 光固定 CO2 之 主要途徑 固碳位置 與 CO2 結合的 化合物 固碳酵素 PEP 羧化酶 PEP 羧化酶 > Rubisco 草醋酸→蘋果酸(4C) 葉肉細胞 RuBP(5C) PCR 循環 PCA 循環 + PCR 循環 (以空間分隔) 葉肉細胞 磷酸烯醇丙酮酸(PEP) (3C) CAM 循環 + PCR 循環 (以時間分隔) 葉肉細胞 晚上:磷酸烯醇丙酮酸 (PEP) (3C) 白天:RuBP(5C) Rubisco 對 CO2 的親和力: 固碳產物 3-磷酸甘油酸(3-PGA)(3C) PEP 羧化酶 → 在低 CO2 環境下 C4 比 C3 固碳效率好 晚上: 3-磷酸甘油酸 (3-PGA)(3C) 白天:草醋酸→蘋果酸(4C) 卡爾文循環進 葉肉細胞 行位置 光飽和點 全日照 1/3 無 無 維管束鞘細胞 葉肉細胞 19 世紀後期 1. 葉肉細胞呈海綿狀,且 有巨大液胞。 20~30℃ 30~40℃ 35℃ C4 植物 草、多數禾本科植物 熱帶、亞熱帶地區 CAM 植物 科等多肉植物、萬代蘭 乾燥的熱帶沙漠地區 稻、麥、馬鈴薯、番茄、胡 甘蔗、玉米、高粱、熱帶牧 仙人掌科、龍舌蘭科、景天
C3、C4和CAM植物的光合特性比较

C3还原 的场所 叶肉细胞叶 绿体 维管束鞘细 胞叶绿体
暗反应 途径 C3途径 C3途径 C4途径
二、生理特性
在生理上,C4植物一般比C3植物具有较强的
光合作用,这是与C4植物的磷酸烯醇式丙酮 酸羧化酶活性较强,光呼吸很弱有关。
1、羧化酶种类和位置 2、PEP羧化酶对CO2的亲和力强
1、羧化酶种类和位置
C3(PEP) C3 C5
多种酶参加 催化
NADP+ ATP ADP+Pi
C3ADP+Pi 4植物光合途径的比较 和C ATP (丙酮酸)
2、C3植物和C4植物光合作用途径比较
项目 种类
(CH2O)
CO2固定 CO2的受体 后产物 C5 PEP ( C3 ) 2C3 C4
C3植物 C4植物
CO2固定 的场所 叶肉细胞叶 绿体 叶肉细胞叶 绿体
C4植物
PEP 草酰乙 酸 16~18
CAM植物
光下:RuBP ; 暗中:PEP 光下:PGA;暗 中:草酰乙酸 0.2
8、CO2固定的最初产 PGA 物 0.30~0.35 9、PEP羧化酶活性 [μ mol/(mg Chl. min)]
10、光合速率 [CO2mg/(dm2.h)]
12、光饱和点
C4植物光合作用特点:
C4途径(叶肉 细胞中的叶绿 体) CO2 酶 C3(PEP) ADP+Pi C5 C4 C3途径和CO2的还 原(维管束鞘细胞 中的叶绿体)
C4
CO2
2C3 多种酶 参加催化
NADPH NADP+ ATP ADP+Pi
C3 ATP (丙酮酸) (CH2O)
C4植物光合作用特点示意图
某农业大学《植物生理学》考试试卷(768)

某农业大学《植物生理学》课程试卷(含答案)__________学年第___学期考试类型:(闭卷)考试考试时间:90 分钟年级专业_____________学号_____________ 姓名_____________1、判断题(10分,每题5分)1. 蛋白质的可逆磷酸化是生物体内一种普遍的翻译后修饰方式。
()[扬州大学2019研]答案:正确解析:蛋白质磷酸化是指由蛋白质激酶催化的把ATP的磷酸基转移到底物蛋白质氨基酸残基(丝氨酸、苏氨酸、酪氨酸)上的过程,或者在信号作用下结合GTP,是生物体内一种普通的调节方式,在细胞信号转导的过程中起重要作用。
蛋白质磷酸化是调节和控制蛋白质活力和功能的最基本、最普遍,也是最重要的机制。
2. 极性运输是生长素的唯一运输方式。
()[扬州大学2019研]答案:错误解析:生长素在植物体内运输方式有两种:在未成熟的组织中,由形态学上端向形态学下端运输,即极性运输;在成熟的组织中,生长素进行非极性运输。
2、名词解释(55分,每题5分)1. 胞饮作用答案:胞饮作用是指吸附在质膜上的物质,通过膜的内折而转移到细胞内以获取物质的过程。
解析:空2. 长日植物[沈阳农业大学2019研]答案:长日照植物是指在一定的发育时期内,每天的光照时间只有大于某一时数(临界日长),并经过一定的天数才能开花的植物。
例如大麦、小麦、菠菜、向日葵等。
这类植物生长在小于临界日长的光照下,就会保持营养生长状态。
用人工方法延长光照时间,可提早开花。
解析:空3. 受精作用答案:受精作用是指开花后,经过花粉在柱头上萌发、花粉管伸长进入胚囊,完成雄性生殖细胞(精子)与雌性生殖细胞(卵细胞)融合的过程。
解析:空4. 大量元素答案:大量元素指植物生命活动必需的、且需要量较多的一些元素。
它们占植物体干重的0.01~10,有碳、氢、氧、氮、磷、钾、钙、镁、硫等9种元素。
5. 伤呼吸答案:伤呼吸是指植物组织受伤后增强的呼吸。
C3、C4、CAM植物的某些光合特征和生理特征比较
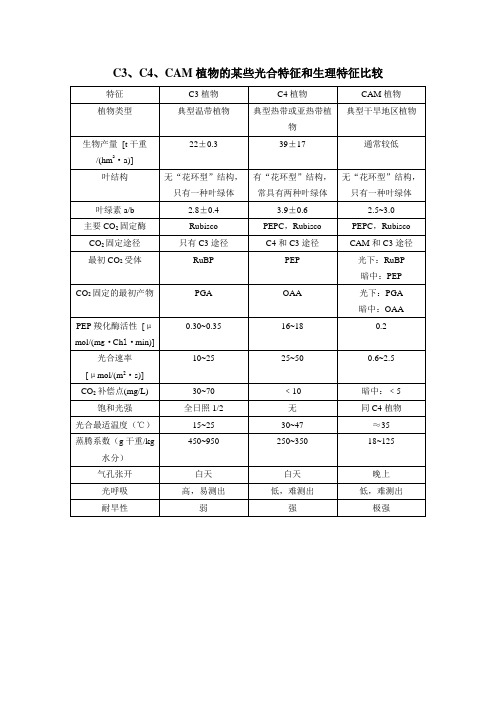
白天
晚上
光呼吸
高,易测出
低,难测出
低,难测出
耐旱性
弱
强
极强
0.30~0.35
16~18
0.2
光合速率
[μmol/(m2·s)]
10~25
25~50
0.6~2.5
CO2补偿点(mg/L)
30~70
﹤10
暗中:﹤5
饱和光强
全日照1/2
无
同C4植物
光合最适温度(℃)
15~25
30~47
≈35
蒸腾系数(g干重/kg水分)
450~950
250~350
18~125
气孔Байду номын сангаас开
叶绿素a/b
2.8±0.4
3.9±0.6
2.5~3.0
主要CO2固定酶
Rubisco
PEPC,Rubisco
PEPC,Rubisco
CO2固定途径
只有C3途径
C4和C3途径
CAM和C3途径
最初CO2受体
RuBP
PEP
光下:RuBP
暗中:PEP
CO2固定的最初产物
PGA
OAA
光下:PGA
暗中:OAA
PEP羧化酶活性[μmol/(mg·Chl·min)]
C3、C4、CAM植物的某些光合特征和生理特征比较
特征
C3植物
C4植物
CAM植物
植物类型
典型温带植物
典型热带或亚热带植物
典型干旱地区植物
生物产量[t干重/(hm2·a)]
22±0.3
39±17
通常较低
叶结构
无“花环型”结构,只有一种叶绿体
C3植物、C4植物和CAM植物的比较

1、C3植物、C4植物和CAM植物 的定义和基本特点
C3植物是指通过卡尔文循环进行光合作用的植物,它们在二氧化碳固定时使 用酶 Rubisco。C4植物是指通过C4途径进行光合作用的植物,它们在二氧化碳固 定时使用酶 PEPC。而CAM植物则是指通过CAM途径进行光合作用的植物,它们在 二氧化碳固定时使用酶 PEP羧化酶。
C4植物的进化
C4植物是指通过C4途径进行二氧化碳固定的植物。这类植物的光合作用效率 较C3植物更高,且受环境因素影响更小。其进化历程大致如下:
C4植物可能最初出现在约6千万年前,当时地球大气中的二氧化碳浓度开始 逐渐降低,导致全球气候逐渐变得干旱。为了适应这种环境,一部分C3植物开始 尝试进行C4途径固定二氧化碳。成功适应这一转变的植物逐渐繁衍开来,形成了 现在约20%的全球植被。典型的C4植物包括玉米、甘蔗等。
分利用效率适应性最强。此外这三种类型的植物在基因组方面也存在差异具 有各自的独特性和进化发展历程。
未来发展方向:针对三种不同类型的植物的特点和优劣我们可以得出以下发 展方向:首先,针对C3植物的光合作用过程可以通过基因工程技术手段来提高它 们的二氧化碳固定能力和光能利用率从而提高其产量;其次针对C4植物在恶劣环 境下的适应能力可以通过深入探究它们的生理机制和基因表达模式来提高它们的 环境适应性并培育出耐旱、耐盐等新品种;最后针对CAM植物的
C3植物的进化
C3植物是指通过卡尔文循环进行二氧化碳固定的植物。这类植物的光合作用 效率较高,但受环境因素影响较大。其进化历程大致如下:
最初的C3植物可能出现在约2.5亿年前,当时地球大气中的二氧化碳浓度较 高,导致全球气候温暖湿润,适合这种植物生长。随着时间的推移,地球气候逐 渐变得干燥,C3植物为了适应这种环境,开始在自身生理代谢上做出调整,例如 增加气孔开度、加强水分吸收等。这些调整使得C3植物能够在干旱条件下生存并 逐渐繁衍开来。现在,C3植物已经成为了地球上最为普遍的一类植物,包括了如 水稻、小麦、大豆等重要的农作物。
c3c4cam植物的区别

c3c4cam植物的区别
C3C4CAM植物在光合作用和碳固定方面存在一些显著的区别。
1. 碳固定:C3途径是植物通过C3途径,利用光反应产生的能量,将二氧化碳固定到3个碳原子中,生成富含三个碳原子的三碳酸分子。
而CAM途径是C4途径,这种途径中,植物通过CAM循环将二氧化碳固定到四个碳原子中,生成富含四个碳原子的四碳二碳酸分子。
2. 光反应:在光反应阶段,C3植物是将二氧化碳固定到3个碳原子上的,而CAM植物则是将二氧化碳固定到四个碳原子上的。
3. 温度适应:CAM植物在温度方面比C3植物更具适应性。
CAM植物在夜间进行二氧化碳的吸收和固定,这有助于降低夜间温度,减少水分蒸发。
4. 水分利用:CAM植物在水分利用方面也较C3植物更为高效。
CAM植物在夜间进行光合作用,这有助于减少水分蒸发,从而更高效地利用水分。
总的来说,CAM植物在碳固定、温度适应和水分利用方面相比C3植物具有更高的效率。
然而,CAM植物的生物量通常比C3植物低,且CAM植物的种植和管理更为复杂。
光合作用—C3、C4、CAM区别

光合作用光合作用可分为:光反应和碳反应(旧称暗反应)两个阶段。
光反应条件:光照、光合色素、光反应酶。
场所:叶绿体的类囊体薄膜。
(蓝细菌等微生物的反应场所在细胞膜:色素所在地)过程:①水的光解:2H2O→4[H]+O2(在光和叶绿体中的色素的催化下)。
②ATP的合成:ADP+Pi+能量→ATP(在酶的催化下)。
影响因素:光照强度、CO2浓度、水分供给、温度、酸碱度、矿质元素等。
意义:①光解水,产生氧气。
②将光能转变成化学能,产生ATP,为碳反应提供能量。
③利用水光解的产物氢离子,合成NADPH(还原型辅酶Ⅱ),为碳反应提供还原剂NADPH(还原型辅酶Ⅱ)。
碳反应条件:多种酶。
场所:叶绿体基质。
过程:①碳的固定:C5+CO2→2C3(在酶的催化下)②C3+[H]→(CH2O)+C5(在ATP供能和酶的催化下)影响因素:温度、CO2浓度㊣卡尔文循环(Calvin Cycle)可分为三个阶段: 羧化(碳的固定)、还原(3-磷酸甘油醛(G3P(PGAL))的合成)、二磷酸核酮糖的再生。
卡尔文循环,又称还原磷酸戊糖循环(以对应呼吸作用中的氧化磷酸戊糖途径)、C3循环(CO2固定的第一产物是三碳化合物)、光合碳还原,还是光合作用的暗反应的一部分。
反应场所为叶绿体内的基质。
㊣固碳作用:※碳三植物(小麦、水稻、大豆、棉花等)二氧化碳经气孔进入叶片后,直接进入叶肉进行卡尔文循环。
而C3植物的维管束鞘细胞很小,不含或含很少叶绿体,卡尔文循环不在这里发生。
※碳四植物(玉米、甘蔗、高粱、苋菜等)碳四植物维管束周围有两种不同类型的细胞:靠近维管束的内层细胞称为鞘细胞,围绕着鞘细胞的外层细胞是叶肉细胞。
①叶肉细胞里的磷酸烯醇式丙酮酸(PEP)经PEP羧化酶的作用,与CO2结合,形成苹果酸或天门冬氨酸。
②这些四碳双羧酸转移到鞘细胞里,通过脱羧酶的作用释放CO2,后者在鞘细胞叶绿体内经核酮糖二磷酸(RuBP)羧化酶作用,进入光合碳循环(卡尔文循环。
C4植物与C3植物的光合作用曲线比较

C4植物与C3植物的光合作用曲线比较1、光合作用强弱变化的指标光合作用强弱变化的指标通常是光合速率和光合生产率光合速率是指单位时间、单位叶面积吸收CO2的量或放出O2的量或有机物的消耗量。
一般测定光合速率的方法都没有把叶片的呼吸作用考虑在内,所以测定的结果实际是光合作用减去呼吸作用的差数,称为表观光合速率或净光合速率.如果把表观光合速率加上呼吸速率,则得到总(真正)光合速率。
光合生产率又称净同化率,是指植物在较长时间(一昼夜或一周)内,单位叶面积生产的干物质量.光合生产率比光合速率低,因为已去掉呼吸等消耗。
2、影响光合作用的因素内因1)叶龄:叶片的光合速率与叶龄密切相关。
从叶片发生到衰老凋萎,其光合速率呈单峰曲线变化。
新形成的嫩叶由于组织发育不健全、叶绿体片层结构不发达、光合色素含量少、光合酶含量少、活性弱、气孔开度低、细胞间隙小、呼吸细胞旺盛等原因,净光合速率很低,需要从其它功能叶片输入同化物。
随着叶片的成长,光合速率不断提高。
当叶片伸展至叶面积最大和叶厚度最大时,光合速率达最大值.通常将叶片充分展开后光合速率维持较高水平的时期,称为叶片功能期,处于功能期的叶叫功能叶.功能期过后,随着叶片衰老,光合速率下降2)光合产物的运输:光合产物从叶片中输出的快慢影响叶片的光合速率。
例如,摘去花或果实使光合产物的输出受阻,叶片的光合速率就随之降低.反之,摘除其他叶片,只留一个叶片和所有花果,留下叶片的光合速率就会增加。
如对苹果枝条进行环割,光合产物会积累,则叶片光合速率明显下降.叶肉细胞中蔗糖的积累会促进叶绿体基质中的淀粉合成和淀粉粒形成,过多的淀粉粒一方面会压迫和损伤叶绿体,另一方面,由于淀粉粒对光有遮挡,从而阻碍光合膜对光的吸收。
(1)光照:光是光合作用的能量来源,是形成叶绿素的必要条件.此外,光还调节着光合酶的活性和气孔开度,因此光是影响光合作用的重要因素1)光强:在暗中叶片无光合作用,只进行细胞呼吸释放CO2.随着光强的增高,光合速率相应提高,当达到某一光强时,叶片的光合速率与呼吸速率相等,净光合速率为零,这时的光强称为光补偿点在一定范围内,光合速率随着光强的增加而呈直线增加;但超过一定光强后,光合速率增加转慢;当达到某一光强时,光合速率就不再随光强增加而增加,这种现象称为光饱和现象。
C4植物与C3植物的光合作用曲线比较

C4植物与C3植物的光合作用曲线比较1、光合作用强弱变化的指标光合作用强弱变化的指标通常就是光合速率与光合生产率光合速率就是指单位时间、单位叶面积吸收CO2的量或放出O2的量或有机物的消耗量。
一般测定光合速率的方法都没有把叶片的呼吸作用考虑在内,所以测定的结果实际就是光合作用减去呼吸作用的差数,称为表观光合速率或净光合速率。
如果把表观光合速率加上呼吸速率,则得到总(真正)光合速率。
光合生产率又称净同化率,就是指植物在较长时间(一昼夜或一周)内,单位叶面积生产的干物质量。
光合生产率比光合速率低,因为已去掉呼吸等消耗。
2、影响光合作用的因素内因1)叶龄:叶片的光合速率与叶龄密切相关。
从叶片发生到衰老凋萎,其光合速率呈单峰曲线变化。
新形成的嫩叶由于组织发育不健全、叶绿体片层结构不发达、光合色素含量少、光合酶含量少、活性弱、气孔开度低、细胞间隙小、呼吸细胞旺盛等原因,净光合速率很低,需要从其它功能叶片输入同化物。
随着叶片的成长,光合速率不断提高。
当叶片伸展至叶面积最大与叶厚度最大时,光合速率达最大值。
通常将叶片充分展开后光合速率维持较高水平的时期,称为叶片功能期,处于功能期的叶叫功能叶。
功能期过后,随着叶片衰老,光合速率下降2)光合产物的运输:光合产物从叶片中输出的快慢影响叶片的光合速率。
例如,摘去花或果实使光合产物的输出受阻,叶片的光合速率就随之降低。
反之,摘除其她叶片,只留一个叶片与所有花果,留下叶片的光合速率就会增加。
如对苹果枝条进行环割,光合产物会积累,则叶片光合速率明显下降。
叶肉细胞中蔗糖的积累会促进叶绿体基质中的淀粉合成与淀粉粒形成,过多的淀粉粒一方面会压迫与损伤叶绿体,另一方面,由于淀粉粒对光有遮挡,从而阻碍光合膜对光的吸收。
(1)光照:光就是光合作用的能量来源,就是形成叶绿素的必要条件。
此外,光还调节着光合酶的活性与气孔开度,因此光就是影响光合作用的重要因素1)光强:在暗中叶片无光合作用,只进行细胞呼吸释放CO2。
C3植物、C4植物与CAM植物
C3植物、C4植物与CAM植物标题: C3植物、C4植物与CAM植物摘要: [C3植物、C4植物与CAM植物]在高等植物中,光合碳同化主要有3种类型:C3途径,C4途径和景天酸代谢途径(CAM)。
C3植物中,CO2的固定主要取决于1,5-二磷酸核酮糖羧化酶(RuBPCase)的活化状态,因为该酶是光合碳循环的入口钥匙。
它催化1,5-二磷酸核酮糖(RuBP)羧化,将大气中的CO2同化,产生两分子磷酸甘油酸,可见RuBPCase在C3植物中同化CO2的重要性。
C4植物是从C3植物进化而来的一种高光效种类。
与C3植物相比,它具有在高光强……[关键词:叶绿体活性基因外稃细胞质叶片大豆]……关键词: 叶绿体活性基因外稃细胞质叶片大豆在高等植物中,光合碳同化主要有3种类型:C3途径,C4途径和景天酸代谢途径(CAM)。
C3植物中,CO2的固定主要取决于1,5-二磷酸核酮糖羧化酶(RuBPCase)的活化状态,因为该酶是光合碳循环的入口钥匙。
它催化1,5-二磷酸核酮糖(RuBP)羧化,将大气中的CO2同化,产生两分子磷酸甘油酸,可见RuBPCase在C3植物中同化CO2的重要性。
C4植物是从C3植物进化而来的一种高光效种类。
与C3植物相比,它具有在高光强,高温及低CO2浓度下,保持高光效的能力。
C4植物固定CO2的酶为磷酸烯醇式丙酮酸羧化酶(PEPCase),与C3作物中RuBPCase相比,PEPCase对CO2的亲和力高。
C4植物的细胞分化为叶肉细胞和鞘细胞,而光合酶在两类细胞中的分布不同,如PEPCase在叶肉细胞固定CO2,生成草酰乙酸(OAA),OAA进一步转化为苹果酸(Mal),Mal进入鞘细胞,脱羧,被位于鞘细胞内的RuBPCase羧化,重新进入卡尔文循环。
这种CO2的浓缩机理导致了鞘细胞内的高浓度的CO2,一方面提高RuBPCase的羧化能力,另一方面又大大抑制了RuBPCase的加氧活性,降低了光呼吸,从而使C4植物保持高的光合效率。
C4植物及C3植物的光合作用曲线比较
C4植物与C3植物的光合作用曲线比较1、光合作用强弱变化的指标光合作用强弱变化的指标通常是光合速率和光合生产率光合速率是指单位时间、单位叶面积吸收CO2的量或放出O2的量或有机物的消耗量。
一般测定光合速率的方法都没有把叶片的呼吸作用考虑在内,所以测定的结果实际是光合作用减去呼吸作用的差数,称为表观光合速率或净光合速率。
如果把表观光合速率加上呼吸速率,则得到总(真正)光合速率。
光合生产率又称净同化率,是指植物在较长时间(一昼夜或一周)内,单位叶面积生产的干物质量。
光合生产率比光合速率低,因为已去掉呼吸等消耗。
2、影响光合作用的因素内因1)叶龄:叶片的光合速率与叶龄密切相关。
从叶片发生到衰老凋萎,其光合速率呈单峰曲线变化。
新形成的嫩叶由于组织发育不健全、叶绿体片层结构不发达、光合色素含量少、光合酶含量少、活性弱、气孔开度低、细胞间隙小、呼吸细胞旺盛等原因,净光合速率很低,需要从其它功能叶片输入同化物。
随着叶片的成长,光合速率不断提高。
当叶片伸展至叶面积最大和叶厚度最大时,光合速率达最大值。
通常将叶片充分展开后光合速率维持较高水平的时期,称为叶片功能期,处于功能期的叶叫功能叶。
功能期过后,随着叶片衰老,光合速率下降2)光合产物的运输:光合产物从叶片中输出的快慢影响叶片的光合速率。
例如,摘去花或果实使光合产物的输出受阻,叶片的光合速率就随之降低。
反之,摘除其他叶片,只留一个叶片和所有花果,留下叶片的光合速率就会增加。
如对苹果枝条进行环割,光合产物会积累,则叶片光合速率明显下降。
叶肉细胞中蔗糖的积累会促进叶绿体基质中的淀粉合成和淀粉粒形成,过多的淀粉粒一方面会压迫和损伤叶绿体,另一方面,由于淀粉粒对光有遮挡,从而阻碍光合膜对光的吸收。
(1)光照:光是光合作用的能量来源,是形成叶绿素的必要条件。
此外,光还调节着光合酶的活性和气孔开度,因此光是影响光合作用的重要因素1)光强:在暗中叶片无光合作用,只进行细胞呼吸释放CO2。
C3植物、C4植物和CAM植物的比较

叶绿体
水在光下分解
[H] 供氢
叶绿体
还
中的色 素
ATP 供能 原
2c3 固
定
co2
多种酶
参加催化 C5
酶
ADP+Pi
(CH2O) [有机物]
光反应阶段
(类囊体膜)
暗反应阶段
.
(叶绿体基质)
C4植物光合作用特点:
C4途径(叶肉 细胞中的叶绿 体)
C3途径和CO2的 还原(维管束鞘细
胞中的叶绿体)
CO2
5、光合速率 [CO2mg/(dm2.h)]
15~35
C4 40~80
光下:C3;暗中: C4
1~4
.
为什么C4植物比C3植物生物产量高?
1、C4植物CO2固定酶活性比C3植物的 高,对CO2的结合力强。 2、C4植物比C3植物光呼吸低 3、C4植物耐高光强,耐高温,耐干旱、 耐高氧。 4、C4植物光合产物运输快
C4 酶
C4
CO2
2C3
C3(PEP) ADP+Pi
ATP
C5 多种酶 参加催化
C3
(丙酮酸) (CH2O)
C4植物光合作. 用特点示意图
NADPH NADP+
ATP ADP+Pi
C3指的是含三个碳原子的化合物 3-磷酸甘油酸 C4指的是含四个碳原子的化合物 草酰乙酸
.
CAM植ism,简称CAM)的植物,多为多浆液植物。在 夜间通过开放的孔吸收CO2,形成草酰乙酸,然后在还原 成苹果酸;第二天光照后苹果酸从液泡中转运回细胞质和 叶绿体中脱羧,释放CO2被进入C3循环。CAM代谢是植 物为了在干旱热带地区生存下来,发展出的一套生存策略。 CO2的固定在时间上分开,这样就可以避免水分过快的流 失,因为气孔只在夜间开放以摄取CO2。
CC和CAM植物的光合特性比较

CC和CAM植物的光合特性比较植物皆以光合作用取得能量,但不同的植物有不同的光合途径和机制。
CC(C3)和CAM(Crassulacean Acid Metabolism)植物是两种常见的光合途径,它们在光合作用中的过程不同,本文将从光合作用和二氧化碳浓度利用等方面比较和分析两种植物的光合特性。
一、光合作用光合作用是植物利用光能将二氧化碳和水转化为有机物质和氧气的过程。
CC植物是最古老的光合途径,主要特征是在叶绿素通道进行光合作用,同时存在最广泛的植物中。
CAM植物是一种较为特殊的光合途径,它通过早晨或夜间吸收二氧化碳,但通过逆向的气孔关闭,使二氧化碳在白天被释放出来用于光合作用,因此叶片上气孔在白天关闭,在夜间开放。
CC植物与CAM植物的光合作用有很大的差异。
CC植物光合速率较高,因为它的光合作用始终处于光合作用状态。
而CAM植物光合速率相对较低,因为它需要在夜间吸收二氧化碳才能在白天完成光合作用。
因此,CC植物在光照期内光合速率较高,而CAM植物则在长时间光照下光合速率较快。
二、二氧化碳浓度利用光合作用不仅关注二氧化碳的运用,而且会受到二氧化碳含量的影响。
在理想状态下,CC植物常常存在二氧化碳饱和,CAM植物却因其气孔时间的不同而产生不同的二氧化碳分压。
CC植物的光合作用发生在近似固定的二氧化碳分压下(近似于大气中的差不多380ppm)拉动,即二氧化碳分压会影响到CC植物的光合速率,低二氧化碳分压会影响到产生光化学反应中的细胞色素f的供应,同时减少了碳的利用能力,影响到光合速率。
因此,二氧化碳含量升高可以增加CC植物的光合速率。
CAM植物与CC植物不同,夜间吸收的二氧化碳会存储在一个细胞器中,而不是被分配至光合作用。
当早上阳光到来时,CAM植物才开始进行光合作用,光合作用才会开始消耗之前吸收的二氧化碳。
由于 CAM植物是在夜间收集二氧化碳并在白天固定二氧化碳,因此其光合速率与二氧化碳浓度有关,低二氧化碳浓度会限制其光合速率。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
C3植物、C4植物和CAM植物主要光合特征和生理特征 特征 1、植物类型 C3植物 C4植物 CAM植物
典型温带植 典型热带或亚 典型干旱地区 物 热带植物 植物 39±17 有Kranz型结 构,常具有两 种叶绿体 3.9±0.6 PEPC, Rubisco C4和C3途径 通常较低 无Kranz型结 构,只有一种 叶绿体 2.5~3.0
C4植物光合作用特点:
C4途径(叶肉 细胞中的叶绿 体) CO2 酶 C3(PEP) ADP+Pi C5 C4 C3途径和CO2的还 原(维管束鞘细胞 中的叶绿体)
C4
CO2
2C3 多种酶 参加催化
NADPH NADP+ ATP ADP+Pi
C3 ATP (丙酮酸) (CH2O)
C4植物光合作用特点示意图
C3(PEP) C3 C5
多种酶参加 催化
NADP+ ATP ADP+Pi
C3ADP+Pi 和 C4 植物光合途径的比较 ATP (丙酮酸)
2、C3植物和C4植物光合作用途径比较
项目 种类
(CH2O)
CO2固定 CO2的受体 后产物 C5 PEP ( C3 ) 2C3 C4
C3植物 C4植物
CO2固定 的场所 叶肉细胞叶 绿体 叶肉细胞叶 绿体
①羧化酶种类和所在位置
②卡尔文循环固定的CO2来源
③进行卡尔文循环的叶绿体位置
④同化CO2和进行卡尔文循环
C3、C4、CAM植物的羧化酶种类和 位置的比较
C3植物
C4植物
CAM植物
2、PEP羧化酶对CO2的亲和力强
C4植物PEP羧化酶对CO2的亲合力高 PEP羧化酶对CO2的亲合力高,Km=7μ mol,而 Rubisco对CO2的亲合力弱,Km=450μ mol,前者 对CO2的亲和力比后者强很多,因此C4植物的PEP 羧化酶就可把外界低浓度的CO2运到鞘薄壁细胞中, 起到CO2泵的作用,增加维管束鞘薄壁细胞的 [CO2]/[O2]比率,改变Rubisco的作用方向,羧化 大于加氧,因C4植物在光照下只产生少量的乙醇酸, 光呼吸非常低。
C3、C4和CAM植物 的光合特性比较
主讲人:张文平 刘成 准备人员:曾子爽 赵媛媛
人们根据光合作用碳素同化的最初光合产物
的不同,把高等植物分成两类: (1)C3植物。这类植物的最初产物是3-磷酸甘 油酸(三碳化合物),这种反应途径称为C3途 径,如水稻、小麦、棉花、大豆等大多数植 物。 (2) C4植物。这类植物以草酰乙酸(四碳化合 物)为最初产物,所以称这种途径为C4途径, 如甘蔗、玉米、高粱等。
叶绿体 含有 含有
植物类型
C3植物
C4植物
叶片的解剖 结构
叶绿体的类 型
无“花环型”结构
维管束鞘细胞及周围的一部分叶 肉细胞构成“花环型”结构
有两种类型的叶绿体,叶肉细胞 的叶绿体正常,维管束鞘细胞的 叶绿体没有基粒
有一种类型的叶绿体 ,主要位于叶肉细胞 中
2、淀粉粒形成的场所
C4植物通过磷酸烯醇式丙酮酸固定二氧化碳的反应是在叶肉 细胞中进行的,生成的四碳双羧酸转移到维管束鞘薄壁细胞 中,放出二氧化碳,参与卡尔文循环,形成糖类,所以甘蔗 、玉米等C4植物进行光合作用时,只有维管束鞘薄壁细胞形 成淀粉,在叶肉细胞中没有淀粉。而水稻等C3植物由于仅有 叶肉细胞含有叶绿体,整个光合过程都是在叶肉细胞里进行 ,淀粉亦只是积累在叶肉细胞中,维管束鞘薄壁细胞不积存 淀粉。
有少量CO2放出,即可被周围排列紧密的叶 肉细胞俘获,被高亲合力的PEP羧化酶固定 ,重新运往鞘细胞。 在维管束鞘细胞中的基粒缺乏PSII,因此不 能产生氧,使光呼吸不易进行。因此,C4植 物又称为低光呼吸植物。
C3、C4 植物的一些条件比较
C4植物耐高光强,耐高温,耐干旱。
C4植物的光饱合点高,达全日照10万lux, C3植物的光饱合点低,4-5万lux。 C4植物起源于热带,耐高温,PEP羧化酶 以HCO3-为底物,高温对其溶解度影响不 大, C3植物的Rubisco以CO2为底物,高 温下溶解度降低。 C4最适温度30~40℃,C3最适温度10~25℃。
22±0.3 2.生物产量 [t干重/(ha2.a)] 3、叶结构 无Kranz型 结构,只有 一种叶绿体 4、叶绿素a/b 2.8±0.4 5、主要CO2固 Rubisco 定酶 6、CO2固定途 只有C3途径 径
C3指的是含三个碳原子的化合物 3-磷酸甘油酸 C4指的是含四个碳原子的化合物 草酰乙酸
一般来说,C4植物比C3植物具有较强的 光合作用
原因可从两方面来讨论
一、叶片结构 二、生理特性
一、叶片结构
1、叶片的显微结构——重点比较维管束鞘细胞结构
C4植物叶片的维管束薄壁细胞较大,其中含有许多较大的叶绿体,叶绿体没 有基粒或基粒发育不良;维管束鞘的外侧密接一层成环状或近于环状排列的 叶肉细胞,组成了“花环型”结构。这种结构是C4植物叶片所特有的特征。 叶肉细胞内的叶绿体数目少,个体小,有基粒。 C3植物的维管束鞘薄壁细胞较小,不含或很少叶绿体,没有“花环型”结构, 维管束鞘周围的叶肉细胞排列松散。
C4植物CO2补偿点低
C4植物的CO2补偿点比较低,为0~10mg/L, 而C3植物CO2补偿点比较高,为50~150mg/L。 因此C4植物成为低补偿植物,C3植物称为高补 偿植物。当外界干旱时,C4植物就能利用低含 量的CO2继续生长,所以在干旱环境中,C4植 物比C3植物生长好。
C4作用光呼吸在维管束鞘细胞中进行,一旦
C3还原 的场所 叶肉细胞叶 绿体 维管束鞘细 胞叶绿体
暗反应 途径 C3途径 C3途径 C4途径
二、生理特性
在生理上,C4植物一般比C3植物具有较强的
光合作用,这是与C4植物的磷酸烯醇式丙酮 酸羧化酶活性较强,光呼吸很弱有关。
1、羧化酶种类和位置 2、PEP羧化酶对CO2的亲和力强
1、羧化酶种类和位置
栅栏组织 维管束 鞘细胞 海绵组织
维 ห้องสมุดไป่ตู้ 束
维管束 鞘细胞 一部分叶 肉细胞
维 管 束
C 4 植物的叶片结构
维管束 鞘细胞 海绵组织
束
鞘细胞 一部分叶 肉细胞
C 4 植物的叶片结构
比较 类型
C 3 植物 C 4 植物
维管束鞘细胞 叶肉细胞 排 列 细胞大小 叶绿体 小 不含 排列疏松 “花环状 ”环绕在维管 大 含有 束鞘细胞外