第二节 微管及其功能
细胞概念图:第2节:微管及其功能

千里之行 始于足下1微管组成蛋白(右图)α-微管蛋白、β-微管蛋白微管组装的结构单位二者结合形成αβ-微管蛋白二聚体(图)α-微管蛋白有一个GTP 结合位点,GTP 不水解,称为不可交换位点(负极)β-微管蛋白有一个GTP 结合位点,GTP 可水解,称为可交换位点(正极)γ-微管蛋白位于中心体外周物质(PCM),用于诱导微管的成核与组装微管组装过程(右下图)二聚体→原纤丝一个二聚体的β亚基不断加聚到另一个二聚体的α亚基13根原纤丝→片层相邻原纤丝错位1nm ,13根形成一个平行四边形片层→成核片层弯曲缝合成微管→组装、去组装踏车行为当一端组装的速度和另一端解聚的速度相同时,微管的长度保持不变,即踏车行为组装与去组装取决于二聚体的浓度是否高于临界浓度微管结合蛋白/药物stathmin(微管去稳定蛋白)机体中二聚体的浓度远高于临界浓度,需要与其结合妨碍组装二者结合受本身磷酸化调控stathmin 磷酸化失去活性stahmin 去磷酸化恢复活性秋水仙素秋水仙素可与二聚体结合而加载到微管负极端,妨碍微管继续组装紫杉醇与微管结合后阻止微管去组装细胞内微管起源胞体起源于→中心体中心粒中心粒外周物质γ-微管蛋白γ-微管蛋白与二聚体α-微管蛋白结合(负极),微管沿正极组装纤毛、鞭毛起源于基体千里之行 始于足下2微管的功能对网格结构的调节微管结合蛋白(右图)MAP →1,2,3,4tau 蛋白MAP2、tauC 端具有微管结合域(带正电荷),可与微管表面(带负电荷)结合,稳定微管(右图)对细胞结构的组织作用细胞器在细胞内具有特定的空间分布,线粒体的运输等依赖的是微管的作用表现(解聚微管后)内质网回缩到细胞核周围高尔基体解体成小膜泡细胞分裂停止依赖于微管的物质运输驱动蛋白(左图)第三种分子马达组成马达结构域两个重要功能ATP 结合位点微管结合位点位于N 端→负极向正极移动位于C 端→正极向负极移动杆状区轻链(尾部)货物结合域沿微管运动的分子机制下图①动力蛋白(右下图)独特之处已知马达蛋白中最大、速度最快细胞质动力蛋白与胞内体/溶酶体、高尔基体及其他一些膜泡运输,动粒和有丝分裂纺锤体的定位,染色体分离等密切相关轴丝动力蛋白下页讲述纤毛与鞭毛的摆动(下面简述)纺锤体和染色体运动参与的蛋白细胞质动力蛋白结合着丝粒,驱动着丝粒沿微管移动驱动蛋白13位于着丝粒,作用于微管正极端,促进微管解聚驱动蛋白5作用于交错重叠的微管,介导驱动纺锤体距离的加长下图②3 千里之行始于足下千里之行 始于足下45 千里之行始于足下图① 图②6千里之行始于足下千里之行 始于足下 7①驱动蛋白沿微管移动的分子机制:当驱动蛋白沿微管行走时,两个马达结构域中位于前面的那个(L)与ATP 结合,导致驱动蛋白发生构象变化,该马达结构域(L)与微管紧密结合,并使后面的马达结构域(T)向前移动(带ADP),越过L ,至微管正极一侧,与微管的结合位点结合(此时移)动了16nm),该马达结构域随即释放ADP ,同时现在位于后面的马达结构水解ATP ,使驱动蛋白二聚体恢复到原来开始时的状态鞭毛的摆动纤毛的结构9+2大多为动纤毛9+0缺乏中央微管,大多为不动纤毛,与感受器有关纤毛的组装(发生)4阶段(右图)高尔基体膜泡包裹成熟的母中心粒的顶端,形成中心粒膜泡(CV)CV 随着新的膜泡融合逐渐变大,成为次级中心粒膜泡(SCV),与此同时母中心粒延伸并获取成为基体的零件,初生轴丝开始显现母中心粒锚定在细胞质膜的纤毛组装位点,SCV 与质膜融合形成杯状结构在"鞭毛内运输复合物"介导下,原生鞭毛延长千里之行始于足下8 图④千里之行 始于足下 9中间丝简介围绕细胞核开始组装,并延伸到细胞质膜与膜蛋白相连形成核纤层,与分裂期细胞核膜消失重组有关体外合成无需ATP 、GTP ,无踏车行为并不是所有真核细胞都具有中间丝基因组家族庞大,赋予不同细胞特殊的细胞骨架,被认为是区分细胞类型的身份证组装结构单位四聚体(下图)过程中间丝蛋白分子(二聚体)→四聚体→中间丝(图解⑤)影响组装的因素中间丝蛋白亚基的磷酸化与去磷酸化磷酸化→网络解体去磷酸化→重新组装核纤层核纤层结构(图⑥)作用支持核膜、连接核膜与染色质分裂前期→核纤层解聚、核膜瓦解(磷酸化)核纤层蛋白A 弥散在细胞千里之行 始于足下1011 千里之行始于足下12 千里之行始于足下。
第九章细胞骨架
体外实验表明,微丝正极与负极都能组装,组 装快的一端为正极,慢的一端为负极;去组装 时,同样正极比负极快。
在微丝组装时,若G-actin 添加到F-actin上的速率正 好等于G-actin 从F-actin失 去的速率时,微丝净长度 没有改变,这种过程称为 踏车行为。它是由G-actin 单体的临界浓度决定的。
七、细胞内依赖于微管的物质运输
有规则地沿微管运输货物的分子马达主要有驱动蛋
细胞中能利用水解 ATP将化学能转变为机械能, 白(kinesin)和胞质动力蛋白(cytoplasmic dynein)。
驱动蛋白及其功能:通常朝微管的正极运动
驱动蛋白沿微管运动的分子机制
胞质动力蛋白及其功能:朝微管的负极运动
八、纤毛和鞭毛的结构与功能
九、纺锤体和染色体运动
微管的类型
单管:大部分细胞质微管是单管微管,它在低温、Ca2+ 和秋水仙素作用下容易解聚,属于不稳定微管。 二联管:是构成纤毛或鞭毛的轴丝微管,是运动类型的 微管,它对低温、Ca2+和秋水仙素都比较稳定。 三联管:存在于中心粒和基体,它对低温、Ca2+和秋水
微管组装是一个动态不稳定的过程
微管组装的动力学不稳定性是指微管组装生长与快 速去组装的交替变换现象。
动力学不稳定性产生的原因
决定微管正端是GTP帽还是GDP帽,又受两种因素
影响,一是结合GTP的游离微管蛋白二聚体的浓度,
二是GTP帽中GTP水解的速度。 当一端组装的速度和另一端解聚的速度相同时,微 管的长度保持稳定,即所谓的踏车行为。
微丝的组装及动力学特性
MF是由G-actin单体形成的多聚体,肌动蛋白单体具有极性, 装 配时呈头尾相接, 故微丝具有极性,有正极与负极之分。 组装可分为成核反应、延长期和稳定期三个阶段。
细胞生物学第四版(细胞骨架2)

一、微管的结构组成与极性 二、微管的组装和去组装 三、微管组织中心 四、微管的动力学性质 五、微管结合蛋白对微管网络结构的调节 六、微管对细胞结构的组织作用 七、细胞内依赖于微管的物质运输 八、纤毛和鞭毛的结构与功能 九、纺锤体与染色体运动
一、微管的结构组成与极性
• 微管(microtubule, MT):一种内/外径分别为 15/24 nm的中空的管状细胞骨架纤维,由α/β微管蛋白 形成的异二聚体组装而成。大部分微管在细胞质内形 成暂时性的结构。
• 微管的动态不稳定性依赖于微管末端β-微管蛋白上 GTP的有无:当体系中α/β-微管蛋白浓度大于临界浓度 时,微管末端新的微管蛋白加入的速度大于GTP水解 的速度,末端的β-微管蛋白上带有GTP,组装快于解 聚;反之,则发生原纤丝弯曲,微管末端倾向于解聚。
• 踏车行为:当微管一端组装的速度与另一端解聚(去 组装)的速度相等时,微管的长度保持稳定,即所谓 的“踏车行为”。
• 原纤丝(protofilament):微管的横截面是由13个球 形蛋白亚基构成的环状结构。微管的管壁是由α/β-微 管蛋白异二聚体纵向排列而成的13根原纤丝合拢而成。 由于相邻的原纤丝之间在排列上存在1nm左右的交错, 以至微管蛋白沿微管的圆周呈螺旋状排列,在微管合 拢的位置微管蛋白构成的螺旋被终止,出现α-微管蛋 白和β-微管蛋白之间的横向结合,并产生纵贯长轴的 “接缝”。每一根原纤丝的两端都是不对称的,它们 在微管的某一端都是α-微管蛋白,而在另一端都是β微管蛋白,从而使得整根微管在结构上呈极性状态。 人们通常将微管组装较快的一端称为正极(拥有β-微 管蛋白),而另一端称为负极。
(一)驱动蛋白及其功能 (二)细胞质动力蛋白及其功能
在轴突内部的微管和膜性细胞器之间有马达 蛋白构成的横桥(箭头所指)相连(图1025)
细胞生物学学习资料(第9-11章)

第九章第一节细胞骨架是指真核细胞内由一些特异蛋白质构成的纤维网架结构。
广义的细胞骨架包括细胞外基质、膜骨架、细胞质骨架和核骨架。
细胞质骨架包括微丝、微管和中间丝3种结构。
一、微丝的组成及其组装(一)结构与组成微丝的主要结构成分是肌动蛋白。
肌动蛋白细胞内有两种存在形式,即肌动蛋白单体(又称球状肌动蛋白,G-ac-tin)和由单体组装而成的纤维状肌动蛋白。
肌动蛋白单体外观呈蝶状,中央有一个裂口,可结合ATP和Mg2+。
每条微丝由二股螺旋状相互盘绕而成,每条丝由肌动蛋白单体头尾相连呈螺旋状排列而成。
(二)微丝的组装及动力学特性当溶液中含有适当浓度的Ca2+,而Na+和K+的浓度很低时,微丝趋向于解聚。
当溶液中含有A TP、Mg+以及较高浓度的Na+和k+时,G-actin趋向于组装成微丝。
组装时,正极组装速度比负极快。
解聚时,正极解聚速度比负极更快。
A TP-G-actin处于临界浓度时,微丝组装和去组装过达到平衡,此时称踏车行为。
体外组装过程中具有踏车行为。
组装过程:成核(Arp2和Arp3等蛋白形成起始复合物)--纤维的延长(ATP帽与ADP帽)--进入稳定期(三)影响微丝组装的特异性药物细胞松弛素可将微丝切断,但对微丝解聚没有明显影响。
鬼笔环肽对微丝的解聚有抑制作用,可使肌动蛋白丝保持稳定状态。
二、微丝网络动态结构的调节与细胞的运动(一)非肌肉细胞内微丝的结合蛋白微丝结合蛋白的类型:成核蛋白、单体-隔离蛋白、单体-聚合蛋白、成束蛋白、封端蛋白、纤维-解聚蛋白、网络-形成蛋白、纤维切割蛋白、膜结合蛋白等。
维丝结合蛋白对微丝组装具有调节作用,如隔离、聚合和成束等。
(二)细胞皮层在紧贴细胞质膜的细胞质区域由微丝结合蛋白将微丝交联形成了凝胶状三维网格结构,这个区域通常称为细胞皮层。
细胞的多种运动方式(如胞质环流、阿米巴运动、变皱膜运动、细胞吞噬)都与细胞凝胶态或胶态的转化有关。
(三)应力纤维在紧贴黏合斑的质膜内侧由微丝紧密排列成束,这种微丝束称为应力纤维。
微管及其功能 2

分类
MAP1、 MAP2、 MAP3 、MAP4、tau蛋白
10
相同点:
•分子结构相似 •C端具有3~4个由18个氨基酸残基构成的结构域 •N端形成横桥,决定了相邻微管的间距
不同点:
MAP2
•存在部位:
tau蛋白
是神经细胞内含量最高 的微管相关蛋白
• 存在部位:
胞体和树突内
•氨基酸残基多(1820个) •诱导产生的微管束内相邻
适温度37℃)、pH(最适pH=6.9)、
微管蛋白临界浓度(critical
concentration)。
8
动力学不稳定性产生的原因:
微管两端具GTP帽(取决于微管蛋
白浓度),微管将继续组装,反之, 无GTP帽则解聚。
9
五、微管结合蛋白对微管网络结构的 调节作用
微管结合蛋白的定义:
始终伴随着微管的组装与去组装的一类蛋白称为微 管结合蛋白 (microtubule associated protein MAP)
动态不稳定性模型 (dynamic instability)
当微管末端有微管蛋白GTP帽,微管继续组 装;而具有微管蛋白-GDP 帽时,将改变异二聚体的 构象使原纤维弯曲而不能 形成微管的管壁,微管则 趋于解聚。
4
1、微管的动态不稳定性
① 在间期细胞内,大部分微
管的一端与中心体相连, 呈辐射状向细胞四周延伸; 一些微管向中心体方向缩 短,另一些不断伸长,到 达一定程度有开始缩短或 伸长,还有一些新的微管 从中心体部位产生。
1、与细胞器的分布以及细胞的形态 发生与维持有关。 2、充当物质运输的轨道
15
6
② 进入分裂期,细胞质微管几乎全部裂解, 而装配形成星微管和纺锤体微管。
第十章-2细胞骨架
秋水仙素、长春花碱、紫杉酚可破坏微管
功能。
二 、微管的化学组成
1 微管蛋白:
Tubulin α
tubulin β
tubulin γ
存在于微管组织中 (MTOCs)即PCM,对微管 的形成和微管极性的确
定有重要作用 。
微管结合蛋白P211
微管相关蛋白MAP 微管聚合蛋白Tau
(二)踏车模型(treadmilling model)
在微管组装达到稳定态时,微
管正端持续加入新的二聚体与微
管负端持续减去已组装的二聚体 达到一种平衡,并且需要GTP水
解供给能量来维持这一平衡
稳定期
踏车现象
+
(一)微管组装的过程
1、成核期 2、延长期
3、稳定期
成核期
(一)微管的动力学特征
由α和β异二聚体构成的微管其两个
末端的结构和动力学特性不同
β亚单位暴露的一端(+)能较快生长。 α亚单位暴露的另一端(-)生长较慢。
在细胞内,微管的负端靠近中心体,位
于细胞中心,而正端则位于细胞质周 围。
(四)影响微管组装的因素
GTP浓度温度、Ca++浓度、pH、压力等,
秋水仙素和秋水仙胺能结合和稳定游离微
管蛋白,引起微管解聚。
长春花碱和长春新碱能结合微管蛋白异二
聚体,抑制它们的聚合作用。
数字通信原理 微管

数字通信原理微管
数字通信原理中,微管是一种非常重要的元件。
微管是一种半导体器件,也称为场效应管(FET)。
它是一种具有放大和开关功能的器件,可被用来实现各种数字电路。
微管有许多独特的特性,例如高输入阻抗、低噪声水平和高频特性,这使得它成为数字通信系统中使用频率较高的元器件之一。
在数字通信系统中,微管通常被用来放大信号或变幻信号。
另外,在数字通信系统的接收器中,微管也被广泛应用。
接收器的任务是接收来自信源的数字信号,并将它转换为电流或电压信号。
微管可以被用来放大并增强这些信号。
在数字通信系统中,微管的选择至关重要。
微管的性能特点,如最大频率、动态范围、输出电阻等,都与其物理结构有关。
系统设计师必须选择适合特定应用程序的微管,以确保系统性能得到最优化。
微管在数字通信原理中的作用十分重要。
只有了解微管的工作原理,才能更好地理解数字通信系统的原理和概念。
对于那些希望从事数字通信系统研究或从事数字通信系统工程的人来说,熟悉微管的原理和应用是非常必要的。
总之,数字通信原理是一个非常复杂的领域,微管作为其中的一个重要元件,它的工作原理和应用是非常需要深入了解的。
了解微管
使用的方法和特性,将有助于提高数字通信系统的性能,并可以为数字通信领域的其他相关研究提供帮助。
微管[微管]细胞学概念
![微管[微管]细胞学概念](https://img.taocdn.com/s3/m/17f2e916a7c30c22590102020740be1e640ecc55.png)
微管[微管]细胞学概念微管的功能支架作用以细胞核为中心向外放射状排列的微管纤维细胞中的微管就像混凝土中的钢筋一样,起支撑作用,在培养的细胞中,微管呈放射状排列在核外,(+)端指向质膜(图9-20),形成平贴在培养皿上的形状。
在神经细胞的轴突和树突中,微管束沿长轴排列,起支撑作用,在胚胎发育阶段微管帮助轴突生长,突入周围组织,在成熟的轴突中,微管是物质运输的路轨。
细胞内运输微管起细胞内物质运输的路轨作用,破坏微管会抑制细胞内的物质运输。
与微管结合而起运输作用的马达蛋白有两大类:驱动蛋白kinesin,动力蛋白dynein,两者均需ATP提供能量。
Kinesin发现于1985年,是由两条轻链和两条重链构成的四聚体(图9-21),外观具有两个球形的头(具有ATP酶活性)、一个螺旋状的杆和两个扇子状的尾。
通过结合和水解ATP,导致颈部发生构象改变,使两个头部交替与微管结合,从而沿微管'行走',将'尾部'结合的'货物'(运输泡或细胞器)转运到其它地方。
据估计哺乳动物中类似于kinesin的蛋白(KLP, kinesin-like protein or KRB, kinesin-related protein)超过50余种,大多数KLP能向着微管(+)极运输小泡,也有些如Ncd蛋白(一种着丝点相关的蛋白)趋向微管的(-)极。
Dynein发现于1963年,因与鞭毛和纤毛的运动有关而得名。
dynein分子量巨大(接近1.5Md),由两条相同的重链和一些种类繁多的轻链以及结合蛋白构成(鞭毛二联微管外臂的动力蛋白具有三个重链)。
其作用主要有以下几个方面:在细胞分裂中推动染色体的分离、驱动鞭毛的运动、向着微管(-)极运输小泡(图9-22)。
形成纺锤体纺锤体是一种微管构成的动态结构,其作用是在分裂细胞中牵引染色体到达分裂极。
纤毛与鞭毛的运动纤毛与鞭毛是相似的两种细胞外长物,前者较短,约5~10um;后者较长,约150um,两者直径相似,均为0.15~0.3um。
细胞骨架(4)
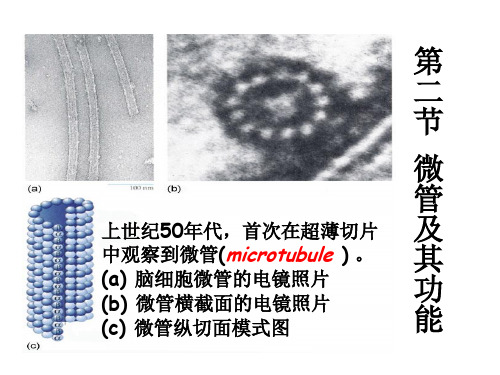
第 二 节 微 管 及 其 功 能
上世纪50年代,首次在超薄切片 中观察到微管(microtubule ) 。 (a) 脑细胞微管的电镜照片 (b) 微管横截面的电镜照片 (c) 微管纵切面模式图
①微管生长时,正端以开口片状存在, 结合GTP的二聚体添加其上。
②快速生长阶段,微管蛋白二聚体添加 速率快于微管蛋白上GTP水解的速率。 微管末端形成GTP帽,有利于添加更 多的亚基使微管生长。但带有开放末 端的微管会产生自发反应,导致管口 闭合。 ③管口闭合,迫使结合的GTP水解,改 变微管蛋白二聚体的构像。 ④当原纤维向外翻卷并发生级联去组装 时,GTP水解形成的张力被释放。
原生纤毛的形成分四个阶段: 2. ① 一个从高尔基体上分离的膜泡形成中心粒膜泡 (CV),包裹在成熟的母中心粒的顶端,一些中 纤 心体蛋白会从母中心粒的顶端移除。 毛 的 ② 母中心粒开始延伸并获取成为基体所需的附属结 构,初生轴丝开始显现。随着新膜泡的融合,CV 组 慢慢变大,最终成为次级中心粒膜(SCV) 装 ③ 母中心粒随同scv向质膜下迁移,当母中心粒锚定 (发 在细胞质膜下的纤毛组装位点时,scv与质膜融合 生) 形成一个杯状结构,称为纤毛项链。 ④ 在鞭毛内运输(IFT)复合物的介导 下,原生鞭毛进一步装配并延长。
(二)基体和其他微管组织中心
基体位于鞭毛和纤毛根部的结构
四、微管的动力学性质
1.微管的动态不稳定性: 在间期细胞,大部分微管与中心体相连,呈辐射状想 细胞四周延伸。一些微管向中心体方向缩短,另一些不断 伸长,达到一定程度又开始缩短或再次伸长,还有一些新 的微管从中心体部位产生。 进入分裂期,细胞质微管几乎全部解聚,而装配形成 星微管和纺锤体微管。 2.微管的稳定性差异: 源于中心体的微管时刻处于动态平衡中,并对各种理 化因素和化学药物敏感; 源于基体的微管,结构稳定; 神经突起内的微管,结构稳定
第2节:微管及其功能

微管组成蛋白(右图)α-微管蛋白、β-微管蛋白微管组装的结构单位二者结合形成αβ-微管蛋白二聚体(图)α-微管蛋白有一个GTP 结合位点,GTP 不水解,称为不可交换位点(负极)β-微管蛋白有一个GTP 结合位点,GTP 可水解,称为可交换位点(正极)γ-微管蛋白位于中心体外周物质(PCM),用于诱导微管的成核与组装微管组装过程(右下图)二聚体→原纤丝一个二聚体的β亚基不断加聚到另一个二聚体的α亚基13根原纤丝→片层相邻原纤丝错位1nm ,13根形成一个平行四边形片层→成核片层弯曲缝合成微管→组装、去组装踏车行为当一端组装的速度和另一端解聚的速度相同时,微管的长度保持不变,即踏车行为组装与去组装取决于二聚体的浓度是否高于临界浓度微管结合蛋白/药物stathmin(微管去稳定蛋白)机体中二聚体的浓度远高于临界浓度,需要与其结合妨碍组装二者结合受本身磷酸化调控stathmin 磷酸化失去活性stahmin 去磷酸化恢复活性秋水仙素秋水仙素可与二聚体结合而加载到微管负极端,妨碍微管继续组装紫杉醇与微管结合后阻止微管去组装细胞内微管起源胞体起源于→中心体中心粒中心粒外周物质γ-微管蛋白γ-微管蛋白与二聚体α-微管蛋白结合(负极),微管沿正极组装纤毛、鞭毛起源于基体微管的功能对网格结构的调节微管结合蛋白(右图)MAP →1,2,3,4tau 蛋白MAP2、tauC 端具有微管结合域(带正电荷),可与微管表面(带负电荷)结合,稳定微管(右图)对细胞结构的组织作用细胞器在细胞内具有特定的空间分布,线粒体的运输等依赖的是微管的作用表现(解聚微管后)内质网回缩到细胞核周围高尔基体解体成小膜泡细胞分裂停止依赖于微管的物质运输驱动蛋白(左图)第三种分子马达组成马达结构域两个重要功能ATP 结合位点微管结合位点位于N 端→负极向正极移动位于C 端→正极向负极移动杆状区轻链(尾部)货物结合域沿微管运动的分子机制下图①动力蛋白(右下图)独特之处已知马达蛋白中最大、速度最快细胞质动力蛋白与胞内体/溶酶体、高尔基体及其他一些膜泡运输,动粒和有丝分裂纺锤体的定位,染色体分离等密切相关轴丝动力蛋白下页讲述纤毛与鞭毛的摆动(下面简述)纺锤体和染色体运动参与的蛋白细胞质动力蛋白结合着丝粒,驱动着丝粒沿微管移动驱动蛋白13位于着丝粒,作用于微管正极端,促进微管解聚驱动蛋白5作用于交错重叠的微管,介导驱动纺锤体距离的加长下图②图① 图②①驱动蛋白沿微管移动的分子机制:当驱动蛋白沿微管行走时,两个马达结构域中位于前面的那个(L)与ATP 结合, 导致驱动蛋白发生构象变化,该马达结构域(L)与微管紧密结合,并使后面的马达结 构域(T)向前移动(带ADP),越过L ,至微管正极一侧,与微管的结合位点结合(此时移) 动了16nm),该马达结构域随即释放ADP ,同时现在位于后面的马达结构水解ATP , 使驱动蛋白二聚体恢复到原来开始时的状态纤毛与鞭毛的摆动纤毛的结构9+2大多为动纤毛9+0缺乏中央微管,大多为不动纤毛,与感受器有关纤毛的组装(发生)4阶段(右图)高尔基体膜泡包裹成熟的母中心粒的顶端,形成中心粒膜泡(CV)CV 随着新的膜泡融合逐渐变大,成为次级中心粒膜泡(SCV),与此同时母中心粒延伸并获取成为基体的零件,初生轴丝开始显现母中心粒锚定在细胞质膜的纤毛组装位点,SCV 与质膜融合形成杯状结构在"鞭毛内运输复合物"介导下,原生鞭毛延长鞭毛内运输复合物复合物B 由驱动蛋白2驱动,负责运输组装所需物质到纤毛顶端复合物A 由动力蛋白驱动,把完成使命的蛋白从顶端运往胞体纤毛/鞭毛的运动机制原理二联管间的纤毛动力蛋白的行走产生的动力,由于“微管的固定”,而转化成纤毛的局部弯曲过程右图下中间丝简介围绕细胞核开始组装,并延伸到细胞质膜与膜蛋白相连形成核纤层,与分裂期细胞核膜消失重组有关体外合成无需ATP 、GTP ,无踏车行为并不是所有真核细胞都具有中间丝基因组家族庞大,赋予不同细胞特殊的细胞骨架,被认为是区分细胞类型的身份证组装结构单位四聚体(下图)过程中间丝蛋白分子(二聚体)→四聚体→中间丝(图解⑤)影响组装的因素中间丝蛋白亚基的磷酸化与去磷酸化磷酸化→网络解体去磷酸化→重新组装核纤层核纤层结构(图⑥)作用支持核膜、连接核膜与染色质核纤层的解体与组装分裂前期→核纤层解聚、核膜瓦解(磷酸化)核纤层蛋白A 弥散在细胞核纤层蛋白B 与核膜小泡保持结合分裂末期带有核纤层蛋白B 的核膜小泡重新形成新的核膜。
第十章-细胞骨架

非肌细胞前缘肌动蛋白的聚合和伪足的形成
(五)微绒毛(microvillus) 是肠上皮细胞的指状突起,用以增加肠上皮细胞 表面积,以利于营养的快速吸收。 微丝束功能:支撑微绒毛,无收缩功能
微绒毛中的微丝和微丝结合蛋白
(六)胞质分裂环
胞质分裂环由大量反向平行排 列的微丝组成,其收缩机制是肌动 蛋白和肌球蛋白相对滑动。
2、Ca2+的释放:肌质网→肌浆
3、原肌球蛋白位移:暴露出肌动蛋白与肌球蛋白 结合位点 4、肌动蛋白丝与肌球蛋白丝的相对滑动:肌球蛋 白沿肌动蛋白丝滑动,水解ATP 5、Ca2+的回收 :收缩停止
结合
分开
-端
+端
+端抬升
构象改变
Pi释放
-端 Ga2+
+端
第二节、微 管(Microtubules)及其功能
促进微管的装配,并使已形成的微管稳定。
三、微管组织中心(MTOC)
◆概念:在生理状态或实验处理时,能够起始微管的 成核作用并使之延伸的细胞结构称为微管组织中心 (microtubule organizing center, MTOC)。
◆常见微管组织中心:
肌肉可看作一种特别富含细胞骨架的效力 非常高的能量转换器,它直接将化学能转变为 机械能。
(一)肌纤维的结构
骨骼肌及肌纤维的结构
粗肌丝与细肌丝的分子结构
(二)肌肉收缩系统中的有关蛋白
①肌球蛋白(myosin) 头部具ATP酶活力,沿微丝从负极到 正极进行运动。 主要分布于肌细胞,有两个球形头部结构域(具有 ATPase活性)和尾部链,多个Myosin尾部相互缠绕,形成 myosin filament,即粗肌丝。 ②原肌球蛋白(tropomyosin, Tm)由两条平行的多肽链形
《微管及其应用》课件

微管的功能
细胞骨架:构成细胞骨架,维持细 胞形态和结构
微管在细胞信号传 导、基因表达调控 等方面具有重要作 用
微管在医学、生物 学等领域具有广泛 的应用前景,如药 物研发、基因治疗 等
微管研究的挑战与机遇
挑战:微管结构的复杂性和动态性 挑战:微管功能的多样性和复杂性 机遇:微管在细胞分裂、细胞运动和细胞信号传导中的重要作用 机遇:微管在疾病治疗和药物开发中的潜在应用
04 微管的应用领域
医学领域
细胞生物学研究:微管在细胞分裂、细胞运动、细胞形态维持等方面具有重要作用
药物研发:微管在细胞信号传导、细胞增殖、细胞凋亡等方面具有重要作用,可用于药 物研发
癌症治疗:微管在细胞分裂、细胞运动、细胞形态维持等方面具有重要作用,可用于癌 症治疗
神经科学:微管在神经细胞形态维持、神经细胞信号传导等方面具有重要作用,可用于 神经科学研究
3D打印技术:通 过3D打印技术制 造微管,提高生 产效率和精度
生物技术:利用 生物技术制造微 管,提高其生物 相容性和可降解 性
智能技术:利用 智能技术制造微 管,提高其智能 化和自动化程度
微管在各领域的发展趋势
生物医学领域:微管在细胞分裂、细胞运动、细胞信号传 导等方面的应用前景
纳米技术领域:微管在纳米材料、纳米器件、纳米机器人 等方面的应用前景
微管在细胞分裂、 细胞运动、细胞 形态维持等方面 发挥重要作用
微管稳定性受到多 种因素影响,如微 管蛋白的磷酸化、 去磷酸化等
研究微管分子的机械性质及其生物学功能

研究微管分子的机械性质及其生物学功能微管作为一种重要的细胞骨架蛋白,广泛存在于生物体内。
除了在细胞分裂、细胞运动、信号传递等功能中扮演重要角色外,微管还具有一系列独特的力学特性和生物学功能。
本文将着重探讨微管分子的机械性质及其生物学功能。
一、微管分子的结构与功能微管是由α-和β-微管蛋白(microtubule-associated proteins)组成的纤维状蛋白体系,具有亚微米级的直径和数微米到数百微米不等的长度。
它们具有于骨架支撑细胞、调节细胞形态和运动、分子转运等重要功能。
微管由α和β微管蛋白分子相互组装而成,每一个微管蛋白分子有一个α和一个β两个可交换的亚基,且这两个亚基的朝向不对称,使得微管有了极性性质。
微管通过它们的独特结构和动态重组能力,参与细胞架构的多种功能。
二、微管的力学特性1. 微管的强度微管由蛋白分子多层环状编织而成,这种编织方式使得微管的结构非常牢固。
一般来说,微管的强度是它的初始结构、构象、分支数目和比例、以及液体环境等因素的综合结果。
2. 微管的刚度微管的刚度取决于其组成蛋白分子的性质,包括α和β微管蛋白、微管相关蛋白等。
同时,刚度也受到活性调控和剪切等外力的影响。
3. 微管的抗弯性微管的抗弯性是指它们在受到弯曲扭曲等外力作用时的力学响应特性。
除了与微管分子特有的管中空结构相关的因素外,还受到微管内部的生物大分子和像ATP、GTP等化学小分子的影响。
三、微管的生物学功能1. 物质输送微管与马达蛋白相互作用,能够在细胞内迅速转移分子。
例如,微管参与许多神经递质的运输作用,并驱动细胞质包含线粒体、蛋白质酶等器官组织的移动。
失去微管的细胞则会导致慢性或急性的能量不足和代谢异常。
2. 细胞运动细胞骨架的重要组成部分之一就是微管。
在细胞活动中,微管通常呈现出异向分布的构象特征,从而驱动细胞的伸缩和变形。
3. 细胞分裂微管在细胞周期的不同阶段发挥着不同的作用。
在细胞分裂前期,微管会重新定位细胞内的各种器官,由此引导着新细胞的分裂和细胞核的分离。
第九章_细胞骨架

2. 装配
3. 微管特异性药物
4. 微管组织中心(MTOC)
5. 微管结合蛋白(MAP) 6. 微管功能
1.微管结构与组成
微管可装配成单管, 二联管(纤毛和鞭毛 中),三联管(中心粒 和基体中)。
2. 装配
① 装配方式 ② 所有的微管都有确定的极性 ③ 微管装配是一个动态不稳定过程
成核蛋白(nucleating protein),使游离的actin核化开始组装 单体聚合蛋白-------使肌动蛋白单体组装到纤维。如:profilin 微丝解聚蛋白-------使微丝快速解聚,例如:cofilin 此外,封端蛋白、隐蔽蛋白、纤维切断蛋白
成束 成网
封端蛋白
交联蛋白
单体隐蔽蛋白
核化蛋白
③ 中心体(centrosome)
④ 基体(basal body)
① 微管组织中心(MTOC): 微管在生理状态或实 验处理解聚后重新装配的发生处称为微管组 织中心(microtubule organizing center, MTOC)。 ② 常见微管组织中心 a) 间期细胞MTOC: 中心体(动态微管) b) 分裂细胞MTOC: 有丝分裂纺锤体极 (动态 微管) c) 鞭毛纤毛细胞MTOC:基体(永久性结构)
(五) 微绒毛
微丝束
肌动蛋白纤维 绒毛蛋白 作为横桥起稳定、维持微丝束, 使微丝束与细胞膜连接起来。 毛缘蛋白
端网结构
(六)胞质分裂环: 有丝分裂末期,两个即将分离的子细胞内产生收缩 环,收缩环由平行排列的微丝和myosin II组成。随 着收缩环的收缩,两个子细胞的胞质分离,在细胞松 驰素存在的情况下,不能形成胞质分裂环,因此形 成双核细胞。
ATP- G-actin与F-actin末端亲和力强 V(+)大于V(-) ADP- G-actin与F-actin末端亲和力弱
师生共讲-微管的功能

松鼠鱼的一个红色素细胞中色素颗粒运动的电镜观察
沿微管运输 的囊
谢谢
微管的功能
目录
支持和维持细 胞形态
为细胞内物质 形成中心粒、 运输提供轨道, 纺锤体、基体、 也为细胞器的 纤毛、鞭毛 分布提供结合
位点
支持和维持细胞形态
动蛋白与内质网膜的胞质面结合,使胞质 微管与膜平行排列
ER
秋水仙处理的成纤维细胞形态变化
支持和维持细胞形态:神经元细胞
秋水仙素使微管解体 神经元的突起停止生长,甚至缩回胞体
形成鞭毛
形成纤毛
形成纤毛
形成中心粒
中心体:微管组织中心
中心体:微管组织中心
中心体:在细胞分裂期指导纺锤丝排列和 染Байду номын сангаас体的移动
形成纺锤体
纺锤体:染色体的运动 ::
形成基粒
物质运输:马达蛋白
驱动蛋白 动力蛋白
物质运输:驱动蛋白
物质运输:动力蛋白
比较:反向运动
物质运输
神经轴突中的蛋白和膜结构的转运; 鱼色素细胞中色素颗粒的运输; 微管可作为高尔基体和其他小泡及颗粒的
支持和维持细胞形态:早老性痴呆与tau蛋白
Tau过度磷酸化 从微管上解离 形成神经元缠结 微管稳定性下降,结构紊乱 神经细胞死亡
形成纤毛、鞭毛
轴心含“9+2”排列的平行微管
形成纤毛、鞭毛
动力蛋白臂(dynein arms)水解ATP产生能 量,驱动相邻轴心外 围的微管二联体间 (outer doublet MTs) 的相对滑动
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
作用于微管的特异性药物
• 1、一此药物如秋水仙素和紫杉醇等可以影 响细胞内微管的组装和去组装。①秋水仙 素,它阻断微管蛋白装配成微管; ②紫杉 酚能促进微管装配,并使已形成的微管稳 定。
• 2、微管组装和去组装的动态行为还与温度 有关。温度高于20℃时微管蛋白亚基可以 组装成微管,而当温度较低时微管会发生 去组装,但冷稳定微管不会去组装 。
基体和其他微管组织中心
• 1、鞭毛和纤毛内部的微管起源于其基体的 结构。基体在结构上与中心粒基本一致, 其壁由9组三联体微管构成,微管A完全微 管,微管B和C为不完全微管 。
• 2、中心粒体基体都具有自我复制的性质, 新的中心粒是由原来的中心粒在S期复制而 来。
微管的动力学性质
• 细胞处于生长状态时,微管的组装与去组 装也并不是同步进行的,往往可以观察到 一部分微管正在组装,而另一部分的微管 却处在解聚过程中。
2、从低等的单细胞真核生物到高等哺乳动物,微管蛋白在生物进化过程 可能是最稳定的蛋白分子之一(P281,L7,W1) 。
3、微管管壁是由αβ-微管蛋白二聚体纵向排列而成的原纤丝构成,13 根原纤丝合拢后构成微管的管壁(P328,LR11,W1) 。
4、每一根原纤丝,它们在微管的某一端都是α-微管蛋白,而在另一端 都是β-微管蛋白。人们通常将组装较快的一端(β-微管蛋白端)称 为正极,而另一端(α-微管蛋白端)为负极(P282,L9,WR8) 。
• 细胞内的微管组装通常都起源于某一特殊 位点,如间期、G0期细胞内的微管以及有 丝分裂时的纺锤体微管大都起源于中心体, 纤毛和鞭毛内部的微管起源于基体。
间期细胞:细胞质微管与微管亚蛋白库处于相对平 衡状态; 分裂期:胞质微管装配和去装配动态受细胞周期控,
前期:胞质微管网络中的微管去组装,游离 的微管蛋白 亚单位装配为纺锤体;
• ①tau蛋白(神经轴突中,具有热稳定性)、 ② 突M)A、P③1(M对A热P2敏(感见,于见神于经神元经胞轴体突和和树树突, 具 高热 度稳热定稳性定)性、 )④。MAP4(各种细胞中,具
• MAPS与微管的结合有助于提高微管的稳定 性,用秋水仙素处理一般间期细胞可以阻
止微管的组装,但并不影响解聚,在神经
5、比结构上看,细胞内的微管有3种类型,它们是单管(细胞质微管和 纺锤体微管)、二联管(纤毛或鞭毛的轴丝微管),三联管(中心粒 或基体的微管) (P282,LR6,W17) 。
13
13+10
13+10+10
二、微管的组装和去组装
• 微管的体外组装与踏车行为 • 作用于微管的特异性药物
微管在体外的组装过程可分为成核和延伸两个阶段。
• 2、动物细胞的间期微管通常都是从中心体开始生长。中 心体含有一对桶状的中心粒,彼此垂直分布,外面被无定 形的中心粒外周物质所包围 。
• 3、每个中心粒含有9组等间三联体微管(13根原纤丝的 微管A,10根原纤丝的微管B,10根原纤丝的微管C) 。
• 4、微管并不起源于中心粒,而是在中心粒外周物质区域 成核。
①一些微管蛋白(α微管蛋白和β微管蛋白)二聚体( ①αβ二聚体) 首先纵向聚合形成短的丝状结构( ②原纤维),即所谓的成核反应 ②然后通过两端以及侧面增加二聚体而扩展成片状,当片状聚合物加宽到大致 13根原纤丝时,即合拢成为一段微管( ③微管)。新的微管蛋白二聚体不断地 组装到这段微管的两端,使之延长
• 通常持有α微管蛋白的负极端组装较慢,而 持有β微管蛋白的正极端组装较快。微管的 组装同样与其底物(携带GDP αβ二聚体) 的浓度有关(微管两端具GTP帽,微管将 继续装配,具GDP帽则解聚)
• 当一端组装的速度和另一端解聚的速度相 同时,微管的长度保持稳定,即所谓的踏 车行为
• 细胞内微管的组装和去组装在时间和空间 上是高度有序的
• 有时整根微管解聚后又从中心体的同一个 微管蛋白的环状模板上从新组装。
• 甚至在同一根微管的末端,其组装和去组 装可以反复进行。
• 以上微管所表现的这种动力学不稳定性通 常发生在正极或中心体远端
五、微管结合蛋白对微管网络结
构的调节
• 始终伴随着微管的组装和去组装而存在的 这类蛋白称之为身微管结合蛋白 (microtubule-associated protein, MAP)。主要有
柔韧性铰链
转运泡 驱动蛋白受体
驱动蛋白
球形头部
驱动蛋白介导转运泡由微管负极向正极移动ຫໍສະໝຸດ 八、鞭毛与纤毛的结构与功能
一、纤毛与鞭毛的结构
• 1、鞭毛与纤毛这种轴丝微管的排列方式被 称为“9+2”排列,与中心体微管一样,所 有的轴丝微管的极性都是正级端向外,即 指向纤毛或鞭毛的顶端(P296,LR18, W6) [微管(-)极指向MTOC,(+)极背 向MTOC]。
一、微管的结构组成与极性
1、微管由微管蛋白亚基组装而成。每个微管蛋白亚基都是由2个非常相 似的球状蛋白(α微管蛋白和β微管蛋白结合而成的导二聚体,这种 αβ-微管蛋白二聚体是细胞质内游离态微管蛋白的主要存在形式, 也是微管组装的基本结构单位(P280,LR2,W1)。 α微管蛋白含 450个氨基酸残基,β微管蛋白含455个氨基酸残基。两者均含有酸性 C端序列(P281,L4,W3) 。
• 2、位于纤毛和鞭毛基部的基体在结构上与 中心粒类似,呈“9+0”模式(P296, LR9,W1) 。
二、纤毛与鞭毛的运动机制
• 1、A管动力蛋白头部B管接触促使动力蛋白 结合ATP水解,产物释放,同时造成头部角 度改变
• 2、新的ATP结合使动力蛋白头部与B管脱 离
• 3、ATP水解,其释放的能量使头部的角度 复原
三、微管组织中心
• (一)中心体 • (二)基体和其他微管组织中心
中心体
• 1、在活细胞中,能够起始微管的成核作用,并使之延伸 的细胞结构称为微管组织中心(microtubule organizing center, MTOC),除中心体以外,细胞内起微管组织中 心作用的类似结构还有位于纤毛和鞭毛基部的基体等细胞 器。
• 分子马达:能利用水解ATP将化学能转变为 机械能,有规则的沿微管运输货物
• 分子马达主要有驱动蛋白和胞质动力蛋白
驱动蛋白及其功能
• 它能运载膜性细胞器沿着微管向轴突末梢 移动
• 通常由2条重链(KHC)和2条轻链(KLC) 组成
驱动蛋白沿微管运动的分子机制
• 驱动蛋白马达结构有两个重要的功能位点: 1、ATP结合位点2、微管结合位点
• 驱动蛋白沿微管运动的分子模型有两种: 步行模型和(inchworm)爬行模型
• 步行模型像“人走路一样”每步移动16nm, (inchworm)爬行模型每步移动8nm。
步行模型过程
• 两个头部交替与微管结合,以确保在移动 过程中不会从微管上掉下来,马达结构域 每水解一个ATP分子,整个分子就向前移动 一步(16nm)
元的树突或轴突内部,这些药物同样能与
微管蛋白二聚体结合而影响微管的组装,
但神经突起内部的微管似乎并不那么容易 解聚,原因是大量的MAPS与微管结合,从 而发挥稳定微管作用
六、微管对细胞结构的组织作用
• 微管与细胞器的分布以及细胞的形态发生 与维持有很大的关系
七、细胞内依赖于微管的物质运输
• 观察活细胞,可以看到许多细胞器或膜状 小泡在细胞内或轴突内部沿微管作定向运 动,在微管和膜泡之间存在一类既能与微 管结合,又能与膜泡特异性结合的是分子 马达
第二节 微管及其功能
生技31103 许瑶
• 一、微管的结构组成与极性 • 二、微管的组装和去组装 • 三、微管组织中心 • 四、微管的动力学性质 • 五、微管结合蛋白对微管网络结构的调节 • 六、微管对细胞结构的组织作用 • 七、细胞内依赖于微管的物质运输 • 八、纤毛和鞭毛的结构与功能 • 九、纺锤体和染色体运动
• 4、带有水解产物动力蛋白头部与B管上另 一位点结合,开始又一次循环
九、纺锤体和染色体运动
• 1、纺锤体微管可分类为:①动粒微管; ② 极微管; ③星体微管 。