eagle现象及细菌天然耐药
铜绿耐药机制

铜绿耐药机制
铜绿耐药是指革兰氏阴性菌铜绿假单胞菌(Pseudomonas aeruginosa)对抗生素的抗药性。
具体的耐药机制可以包括以
下几个方面:
1. 基因变异:铜绿假单胞菌可以通过基因突变或水平基因传递获得耐药相关基因,从而导致对抗生素的耐药性。
这些基因可以编码产生药物降解酶(如β-内酰胺酶)、药物排出蛋白
(如外膜孔道蛋白)、药物的修饰酶(如磷酸转移酶)等。
2. 外膜通道改变:铜绿假单胞菌的外膜由脂多糖组成,可以起到阻挡抗生素的作用。
耐药菌株可能通过改变外膜通道结构或降低通透性来减少抗生素的进入。
3. 药物靶标改变:铜绿假单胞菌能够通过改变药物的靶标结构,使其失去对抗生素的敏感性。
例如,可能发生药物靶标的突变,导致抗生素无法与其结合或结合力降低。
4. 药物外排泵:铜绿假单胞菌含有多种药物外排泵,这些泵能够将抗生素从细胞内排出,减少药物在菌体内的积累,从而降低抗生素的效果。
综上所述,铜绿耐药机制是多种因素共同作用的结果。
这些机制使得铜绿假单胞菌对多种抗生素具有较高的耐药性,增加了其治疗的难度。
acutalibacteraceae 中文

acutalibacteraceae 中文Acutalibacteraceae(真杆菌科)Acutalibacteraceae是真杆菌目(Acidimicrobiales)中一科细菌,该科细菌的特点和分类地位受到科学界的广泛关注。
本文将介绍Acutalibacteraceae的分类、形态特征、生态学意义和研究进展。
一、分类Acutalibacteraceae是真杆菌目中的一科,目前已发现的属包括Acutalibacter、Georgenia和Zygomonas三个。
这些属的分类地位仍有争议,有学者根据16S rRNA基因序列进行系统发育分析,提出了将Acutalibacter和Zygomonas合并为一个属的建议。
未来的研究将进一步明确Acutalibacteraceae的分类地位。
二、形态特征Acutalibacteraceae的细菌是革兰氏阳性的厌氧菌,细胞呈直杆状。
细胞壁主要由肽聚糖和胆固醇构成,使得细菌在某些环境条件下具有高温和低pH的耐受能力。
在培养基上观察到的菌落通常呈乳黄或灰白色。
此外,Acutalibacteraceae的细胞内还发现有颗粒状的聚合物,可能与其生活方式和生态学意义相关。
三、生态学意义Acutalibacteraceae的细菌在自然环境中广泛存在,包括土壤、水体和肠道等生境。
它们对环境中的有机物分解和矿物质转化具有重要作用。
研究发现Acutalibacteraceae中的一些细菌能够利用复杂的多糖类物质,如纤维素和半纤维素,参与到生物地球化学循环中。
此外,Acutalibacteraceae的细菌还能够产生一些有益的代谢产物,如酸和酶等,对环境中其他微生物的生长和代谢有一定影响。
四、研究进展对Acutalibacteraceae的研究主要集中在菌株的分离与鉴定、生物学特性的研究和生态功能的解析等方面。
近年来,随着高通量测序技术的发展,基于其16S rRNA基因的环境DNA测序分析为Acutalibacteraceae的研究提供了新的方法和思路。
什么叫天然耐药?了解天然耐药有什么意义?

什么叫天然耐药?了解天然耐药有什么意义?
细菌耐药可分为天然耐药(也称固有耐药)和获得性耐药。
天然耐药是由染色体决定的,菌种整体上(几乎全部分离株)都表现为耐药特性。
不同细菌细胞结构与化学组成不同,使其本身对某些抗生素天然不敏感,比如肠杆菌科细菌大肠埃希菌、肺炎克雷伯菌、阴沟肠杆菌等对于万古霉素就天然耐药。
天然耐药以外其他的耐药,往往属于获得性耐药。
细菌获得性耐药的原因可以是敏感细菌在某些环境下自身发生了基因突变产生的耐药性,或者从外源获得耐药基因所产生。
比如,对苯嗖西林敏感的金黄色葡萄球菌获得mecA基因,则会对内酰胺类抗生素产生耐药性。
细菌的天然耐药通常是固定的,可以长期稳定遗传。
实验室不必测试天然耐药。
如果测试,而且在体外试验条件下没有检测出耐药,导致假敏感,向临床报告则将严重误导临床。
比如铜绿假单胞菌对于复方磺胺、头抱嚷的天然耐药,但是有部分实验室采用仪器法可能得出敏感的结果。
因此了解细菌天然耐药知识,可以有效规避这些错误(天然耐药信息可以在医学专业书籍、文件内查询,比如C LSIM100文件,在附录部分会提供临床常见细菌的天然耐药表)。
天然耐药是感染性疾病临床医师、临床药师的必备知识。
临床微生物学从业人员应该积极宣传,避免错误。
铜绿假单胞菌的抗生素耐药性与抗菌治疗策略

铜绿假单胞菌的抗生素耐药性与抗菌治疗策略铜绿假单胞菌(Pseudomonas aeruginosa)是一种常见的革兰阴性细菌,被广泛认为是院内感染的主要致病菌之一。
它能够造成多种感染,特别是对于免疫功能低下的患者,如机械通气、烧伤、固定术后等患者,感染的风险更高。
然而,铜绿假单胞菌的抗生素耐药性日益成为一个严重的问题,给治疗带来了困难。
因此,针对铜绿假单胞菌的抗菌治疗策略需要得到详细的研究和指导。
一、铜绿假单胞菌的抗生素耐药性铜绿假单胞菌的抗生素耐药性主要归因于其先天性耐药性基因的存在以及后天性的耐药机制的获得。
先天性耐药性基因包括外膜通道蛋白质的相关基因、多药泵的基因等,可以降低抗生素进入细菌细胞的能力。
后天性耐药机制则源于铜绿假单胞菌细胞的遗传变异和外源性基因的水平传递。
1.多重抗药 (Multi-drug resistance, MDR):MDR是指铜绿假单胞菌对两种或更多不同类别的抗菌药物耐药。
这种耐药性的主要机制是多药泵的过度表达,它能从细菌细胞中主动排出抗生素,从而降低抗生素在细菌内的有效浓度。
2.广谱β-内酰胺酶 (Extended-spectrum β-lactamases, ESBLs):铜绿假单胞菌产生的β-内酰胺酶能够水解多种β-内酰胺类抗生素,使得这些药物失去抗菌效果。
此外,铜绿假单胞菌也能产生金属酶,使得抗生素大环内酯类和氨基糖苷类产生耐药性。
3.碳青霉烯酶 (Carbapenemases):碳青霉烯酶是一种能够水解碳青霉烯类抗生素的β-内酰胺酶,是目前对碳青霉烯类抗生素最为重要的耐药机制。
碳青霉烯酶主要分为A、B、D三类,其中KPC、NDM和OXA是临床上较为常见的。
二、铜绿假单胞菌的抗菌治疗策略铜绿假单胞菌的抗生素耐药性给其抗菌治疗带来了一定的挑战,因此合理选择抗菌药物和正确使用抗菌药物是至关重要的。
以下是一些常用的抗菌治疗策略:1.组合用药:针对临床上难以治疗的铜绿假单胞菌感染,可以考虑使用两种或更多抗菌药物的组合疗法。
革兰阴性杆菌的主要耐药机制

革兰阴性杆菌的主要耐药机制今天我给大家讲解的是革兰阴性杆菌的主要耐药机制。
首先我们复习一下细菌耐药性的分类,细菌耐药性的分类主要分为天然耐药和获得性耐药,天然耐药是指某微生物种的所有菌株具有相同的内在特性,由染色体介导,通常直接传给子代,是该菌的特征,可以用于鉴定细菌。
例如铜绿假单胞菌,对复方西诺敏是天然耐药,克雷伯菌对氨苄西林天然耐药等等。
而获得性耐药是指发生在一个菌种或菌属中的部分菌株,发生的比例会随时间的变化而改变,通常由可转移的DNA 比如质粒、转座子、整合子等介导,并可水平传播,也可以垂直传播,在缺少抗菌药物的选择压力时,耐药性有时会消失。
而获得性耐药是我们今天讲解的重点。
细菌对抗菌药物的耐药性主要分为下面四种。
第一产生灭活酶;第二,细胞通透性的变化,特别是细胞膜通透性的变化;第三,主动泵出;第四,对抗菌药物作用靶位的修饰改变。
而前面三种主要是革兰阴性杆菌的耐药机制。
最后一种常常表现在革兰阳性菌中。
我们今天讲解的也是以前三种耐药机制为主。
首先我们讲解一下灭活酶。
这是一张所有耐药机制的简易图,我们可以看到右上方有两个耐药机制,第一个是由于某孔蛋白兴奋性的障碍导致抗菌药物不能够顺利地通过细菌的细胞膜,而直接在细胞膜外游走,不能发挥它的作用。
另一个就是泵出机制,由于细胞膜上的泵将细胞膜内的抗菌药物泵出细胞外,使抗菌药物在细胞内的浓度大大降低,本图右下方所指的就是灭活酶的作用机制,细菌对进入细胞内的抗菌药物有两种灭活酶的作用方式,第一是将抗菌药物水解掉,第二是将抗菌药物进行修饰或者将其自身的靶位进行修饰,使抗菌药物不能发挥正常的作用。
这张图的左上方指的是细菌的细胞对于抗菌药物作用的靶位发生了改变,因此,抗菌药物不能够顺利地结合在作用的靶位上,从而失去了自身发挥作用的能力。
左下方这一张图表示的是抗菌药物进入细胞内,应当通过正常的途径发挥其作用,而此时细菌对因此产生一种常用通路使抗菌药物不能发挥其作用,以上机制是细菌的主要耐药机制的简介图。
常见细菌的天然耐药

大肠埃希菌
伤寒沙门菌 志贺菌 奇异变形杆菌 普通变形杆菌/潘氏变形杆菌 克雷伯菌属(肺炎克雷伯菌) 摩根菌属(摩根摩根菌)
阴沟肠杆菌复合群* 赫氏埃希菌 黏质沙雷菌
弗劳地枸椽酸杆菌 科泽枸椽酸杆菌 产气肠杆菌 雷氏普罗威登斯菌/斯氏普罗威登 斯 菌 蜂房哈夫尼亚菌 小肠结肠炎耶尔森菌
பைடு நூலகம்
肠杆菌的天然耐药
注:* .阴沟肠杆菌复合群包含阴沟肠杆菌、溶解肠杆菌、霍氏肠杆菌、阿氏肠杆菌。①普罗威登斯菌对除了阿米 素之外的所有氨基糖苷类耐药,对四环素耐药;斯氏普罗威登斯菌对氨基糖昔类的庆大霉素、奈替米星和妥布霉素 阿米卡星无天然耐药。②三代头抱菌素头孢吡月亏、氨曲南、替卡西林-克拉维酸、哌拉西林-他哩巴坦和碳青霉 没有列出,因为肠杆菌科菌对它们无天然耐药。③肠杆菌科同样对克林霉素、达托霉素、夫地西酸、糖肽类抗生 素、替考拉宁)、利奈唑胺、达福普丁、利福平和大环内酯类抗生素(红霉素、克拉霉素和阿奇霉素)天然耐药 大环内酯类抗生素例外(如沙门菌属及志贺菌属对阿奇霉素)
肠杆菌的天然耐药
天然耐药
克林霉素,达托霉素,夫地西酸,糖肽类抗菌药物(万古霉素、替考拉宁),利奈唑胺, 奎奴普丁-达福普汀,利福平和大环内酯类抗菌药物(红霉素、克拉霉素)等
一代、二代氨基糖首类(庆大霉素、阿米卡星等) 一代、二代头泡菌素(头泡吠辛),头霉素类(头弛西丁),氨基糖苷类(庆大霉素、 阿米卡星) 四环素,替加环素,多黏菌素B/黏菌素,咲喃妥因 氨苄西林,头孢唑林,头孢噻吩,头孢呋辛,四环素,替加环素,多黏菌素B/黏菌素,呋 喃妥因 氨苄西林,替卡西林 氨苄西林,阿莫西林-克拉维酸,头孢唑啉,头孢噻吩,头孢呋辛,四环素,替加环素,多 黏菌素B/ 黏菌素,呋喃妥因 氨苄西林,氨苄西林-舒巴坦,阿莫西林-克拉维酸,头孢唑啉,头孢噻吩,头孢西丁,头孢 替坦,头孢呋辛 氨苄西林,替卡西林 氨苄西林,氨苄西林-舒巴坦,阿莫西林-克拉维酸,头孢噻吩,头孢唑啉,头孢西丁, 头孢替坦,头孢呋辛,多黏菌素B/黏菌素,呋喃妥因 氨苄西林,氨苄西林-舒巴坦,阿莫西林-克拉维酸,头孢噻吩,头孢唑啉,头孢西丁, 头孢替坦,头孢呋辛 氨苄西林,哌拉西林,替卡西林 氨苄西林,阿莫西林-克拉维酸,氨苄西林-舒巴坦,头孢噻吩,头孢唑啉,头孢西丁,头 孢替坦,头孢呋辛 氨苄西林,阿莫西林-克拉维酸,头孢唑啉,头孢噻吩,四环素,替加环素,多黏菌素B/黏 菌素,呋喃妥因,氨基糖苷类(除了阿米卡星和链霉素) 氨苄西林,氨苄西林-舒巴坦,阿莫西林-克拉维酸,头孢噻吩,头孢唑啉,头孢西丁,头孢 替坦 氨苄西林,阿莫西林-克拉维酸,替卡西林,头孢噻吩,头孢唑啉,头孢西丁
鸡和人志贺菌gyrA、parC基因突变与喹诺酮类耐药的关系

Lu H n yn , u h ag C e u Y n i, n iwe, u Lnu Z n h n eg i , n h a q i o g i X e Su n , hn L , a g Xa Wag X n i aj ,e g C eg Z n qn Wa gC u mg g X g n
( CR) a d P , n
q ioo e e i a td t mii e i ( DR wi i yA g n s a ay e .No m t in fb s a r o n e e u l — s tn e r nn rgo QR ) t n g r e e wa n lz d n n r s e g n h ua o s o ae p i we fu d b t n t r e we te Q D e u n e f c i e n u a hg l o di hc a o s t tw t s se t it etrs l I pt o o h R R sq e c s o h k n a d h n S ie a b y i c m l ,w i w s c ni e i u c pi l ts eut n s i fn h sn h bi y . e
( l g fAnma ce c n tr ay M e iie He a r utrlUnv ri Col e o i lS in e a d Vee n r dcn , e i n n Ag i l a iest Z e g h u 4 0 0 ) c u y, h n z o 5 0 2
狐狸源大肠埃希菌TEM1型耐药基因检测和耐药质粒消除研究

狐狸源大肠埃希菌TEM1型耐药基因检测和耐药质粒消除研究高光平;朱利霞;王洪彬;史秋梅【摘要】研究大肠埃希菌耐药性和耐药基因和十二烷基硫酸钠(SDS)对大肠埃希菌耐药质粒消除作用.以大肠埃希菌临床分离株为试验菌株,采用K-B药敏纸片法对6种抗菌药物进行药物敏感性检测,并对大肠埃希菌耐药质粒TEM1型耐药基因进行PCR检测,采用SDS消除大肠埃希菌临床分离菌株的耐药质粒,采用琼脂糖凝胶电泳检测质粒条带,药敏试验检测大肠埃希菌耐药质粒消除前后的药物敏感性.结果表明,大肠埃希菌对庆大霉素敏感,对大观霉素中敏,对氨苄西林、阿莫西林、头孢拉定、环丙沙星耐药;大肠埃希菌耐药质粒检测出TEM1型耐药基因;试验菌株经7.5 g/L的SDS处理至第3代时,质粒条带有明显的减少,TEM1型耐药基因消失,并且恢复了对β内酰胺类抗生素的敏感性,说明耐药质粒已经成功消除.【期刊名称】《动物医学进展》【年(卷),期】2019(040)003【总页数】5页(P40-44)【关键词】大肠埃希菌;耐药基因;耐药质粒;十二烷基硫酸钠;消除【作者】高光平;朱利霞;王洪彬;史秋梅【作者单位】河北科技师范学院/河北省预防兽医学重点实验室,河北秦皇岛066604;河北科技师范学院/河北省预防兽医学重点实验室,河北秦皇岛 066604;河北科技师范学院/河北省预防兽医学重点实验室,河北秦皇岛 066604;河北科技师范学院/河北省预防兽医学重点实验室,河北秦皇岛 066604【正文语种】中文【中图分类】S852.612随着毛皮动物养殖业迅速发展,养殖规模化、集约化程度加深,使得中国成为养殖毛皮动物最多的国家之一,然而大肠杆菌病等细菌性疾病呈现出逐年流行的趋势,给毛皮动物养殖造成了巨大的经济损失。
大肠埃希菌的抗原性和血清型复杂多样,临床症状、病理变化较复杂,对人类和动物的健康造成了严重的威胁。
此外,抗生素的泛用、滥用,导致细菌的耐药情况更加严重[1-2],菌株出现多重耐药性或者交叉耐药等一系列问题,不仅降低了抗生素对细菌的治疗效果,甚至出现部分新药刚研制出不久,病原菌就对其产生了耐药性,使大肠杆菌病在临床上的防治变得更加困难,最终给养殖业的健康发展以及人类自身的健康带来巨大的威胁[3-4]。
志贺氏菌耐药基因

志贺氏菌耐药基因
志贺氏菌(Shigella)是引起细菌性痢疾的病原菌,严重威胁人类健康。
近年来,志贺氏菌出现了多种抗药性,使得治疗变得更加困难。
志贺氏菌的耐药性主要是由其基因所决定的,其中包括下列几种常见的耐药基因:
1.TEM-1基因:能够使细菌产生β-内酰胺酶,使得抗生素青霉素、羟苄青霉素等失去有效性。
2.CTX-M基因:也是产生β-内酰胺酶的基因,耐药性强,往往和TEM-1基因联合作用。
3.OXA基因:能够使细菌产生产羟基苄青霉素酶、环丙沙星酶等,耐药性强。
4. Sul基因:使细菌产生磺胺类耐药基因,使得抗生素磺胺甲噁唑等失去有效性。
5. Tet基因:使细菌产生四环素酶,耐药性强,常与其他耐药基因联合作用。
以上仅为常见的几种志贺氏菌耐药基因,随着时间的推移,还会出现新的耐药基因。
因此,对志贺氏菌进行耐药性检测,了解其耐药基因的存在情况,对于制定针对性治疗方案具有重要意义。
常见革兰阴性菌的天然耐药
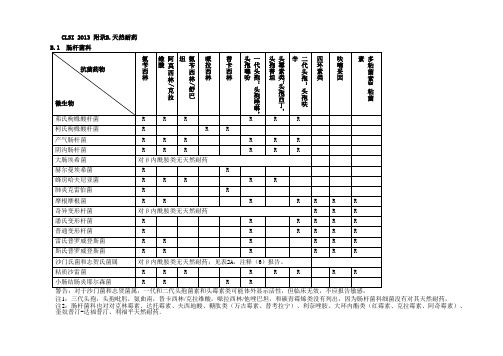
CLSI 2013 附录B.天然耐药
注1:三代头孢,头孢吡肟,氨曲南,替卡西林/克拉维酸,哌拉西林/他唑巴坦,和碳青霉烯类没有列出,因为肠杆菌科细菌没有对其天然耐药。
注2:肠杆菌科也对对克林霉素、达托霉素、夫西地酸、糖肽类(万古霉素、替考拉宁)、利奈唑胺、大环内酯类(红霉素、克拉霉素、阿奇霉素)、奎奴普汀-达福普汀、利福平天然耐药。
CLSI 2013 附录B (续)
+ 嗜麦芽窄食单胞菌对四环素天然耐药,而对多西环素或米诺环素不是天然耐药。
注:非发酵革兰阴性菌也对一代头孢(头孢噻吩、头孢唑林),二代头孢(头孢呋辛),头霉素类(头孢西丁、头孢替坦),克林霉素,达托霉素,夫西地酸,糖肽类(万古霉素、替考拉宁),利奈唑胺,大环内酯类(红霉素、阿奇霉素、克拉霉素),青霉素,奎奴普汀-达福普汀、利福平天然耐药。
CLSI 2013 附录B (续)
B.3 葡萄球菌
注1:革兰阳性菌也对氨曲南、多粘菌素B/粘菌素和萘啶酸天然耐药。
注2:苯唑西林耐药的金黄色葡萄球菌和凝固酶阴性葡萄球菌(耐甲氧西林葡萄球菌[MRS]),被认为对其他β-内酰胺类耐药,如,青霉素类,β-内酰胺/β-内酰胺酶抑制剂复合物,头孢类(除外具有抗MRSA活性的头孢菌素类),和碳青霉烯类。
因为大多数病例证明MRS感染对β-内酰胺类治疗反应差,或因为证明这些药物临床有效性的令人信服的临床资料还没有出现。
CLSI 2013 附录B (续)
警告:对于肠球菌属,头孢菌素类,氨基糖苷类(除外高浓度氨基糖苷类耐药检测),克林霉素,和复方新诺明可能体外显示活性,但临床无效,不应报告敏感。
注:革兰阳性菌也对氨曲南、多粘菌素B/粘菌素和萘啶酸天然耐药。
常见细菌和真菌的天然耐药性知识分享

常见细菌和真菌的天然耐药性常见细菌和真菌的天然耐药性(一)肠杆菌科天然耐药表1、弗氏柠檬酸杆菌对氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)、头孢菌素II代(头孢呋辛)天然耐药。
2、克氏柠檬酸杆菌对氨苄西林、哌拉西林、替卡西林天然耐药。
3、产气肠杆菌和阴沟肠杆菌:氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)、头孢菌素II代(头孢呋辛)。
4、大肠埃希菌:此菌对β-内酰胺类药物无天然耐药。
5、肺炎克雷伯菌和赫氏埃希菌:氨苄西林、替卡西林。
6、蜂房哈夫尼菌:氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)。
7、摩根摩根菌:氨苄西林、阿莫西林/克拉维酸、头孢菌素I代(头孢唑啉、头孢噻吩)、头孢菌素II代(头孢呋辛)、四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
8、普通变形杆菌和彭氏变形杆菌:氨苄西林、头孢菌素I代(头孢唑啉、头孢噻吩)、头孢菌素II代(头孢呋辛)、四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
9、奇异变形杆菌:四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
此菌对青霉素和头孢菌素没有天然耐药性。
10、粘质沙雷氏菌:氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)、头孢菌素II代(头孢呋辛)、呋喃妥因、多粘菌素B、黏菌素。
11、小肠结肠炎耶尔森菌:氨苄西林、阿莫西林/克拉维酸、替卡西林、头孢菌素I代(头孢唑啉、头孢噻吩)。
12、沙门氏菌和志贺氏菌:此菌对β-内酰胺类药物无天然耐药,一代、二代头孢菌素和头霉素在体外可显示活性,但临床无效,不能报告为敏感。
13、雷氏普罗维登斯菌和斯图普罗威登斯菌:氨苄西林、阿莫西林/克拉维酸、头孢菌素I代(头孢唑啉、头孢噻吩)、四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
天然耐药细菌.(优选)

细菌天然耐药一、革兰氏阳性菌*警告:对于肠球菌,头孢菌素、氨基糖苷类(除了高水平耐药筛选外)、克林霉素和复方新诺明在体外有活性,但在临床上无效,因此,不应该报告敏感。
注:革兰氏阳性细菌对氨曲南、多粘菌素B/黏菌素和萘啶酸也存在天然耐药。
苯唑西林耐药的金黄色葡萄球菌和凝固酶阴性的葡萄球菌(耐甲氧西林葡萄球菌MRS)被认为对其他β-内酰胺类药物,即青霉素类、β-内酰胺类/β内酰胺酶抑制剂复合物、头孢烯类(除具有抗MRSA活性的头孢菌素,如头孢噻吩、头孢氨苄、头孢羟氨苄等)和碳青霉烯类耐药。
这是因为大多数情况下对MRS引起的感染用 -内酰胺类治疗疗效不佳,或因为没有令人信服的临床数据证明这些药物的临床疗效。
二、革兰氏阴性菌*由于舒巴坦对不动杆菌具有抗菌活性,因此,鲍曼/醋酸钙不动杆菌可出现氨苄西林/舒巴坦敏感。
+嗜麦芽窄食单胞菌对四环素存在天然耐药,但对对喜欢苏和米诺环素不存在注:非发酵革兰阴性菌对青霉素、1代头孢菌素(头孢噻吩、头孢唑林)、2代头孢菌素(头孢呋喃)、头孢菌素(头孢西丁、头孢替坦)、克林霉素、达托霉素、夫西地酸、糖肽类(万古霉素、替考拉宁)、利奈唑胺、大环内酯类(红霉素、克拉霉素和阿奇霉素)、奎奴普丁、达福普丁和利福平存在天然耐药。
*变形菌属、普罗威登斯菌属和摩根菌属有可能由于非碳青霉烯酶导致的亚胺培南MIC升高的情况,报告检测出敏感的结果。
+斯氏普罗威登斯菌应该对庆大霉素、奈替米星和妥布霉素天然耐药,但对阿米卡星没有天然耐药。
注:3代头孢菌素,头孢吡肟,氨曲南、替卡西林/克拉维酸,哌拉西林/他唑巴坦和碳青霉烯类抗生素没有天然耐药,肠杆菌科细菌同样对克林霉素、达托霉素、夫西地酸、糖肽类(万古霉素、替考拉宁)、利奈唑胺、奎奴普丁-达福普丁,利福平和大环内酯类(红霉素、克拉霉素和阿奇霉素)天然耐药,然而,大环内酯类也有例外(沙门菌属和志贺菌属对阿奇霉素没有天然耐药)。
最新文件---------------- 仅供参考--------------------已改成-----------word文本--------------------- 方便更改。
常见细菌和真菌的天然耐药性

常见细菌和真菌的天然耐药性(一)肠杆菌科天然耐药表1、弗氏柠檬酸杆菌对氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)、头孢菌素II代(头孢呋辛)天然耐药。
2、克氏柠檬酸杆菌对氨苄西林、哌拉西林、替卡西林天然耐药。
3、产气肠杆菌和阴沟肠杆菌:氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)、头孢菌素II代(头孢呋辛)。
4、大肠埃希菌:此菌对β-内酰胺类药物无天然耐药。
5、肺炎克雷伯菌和赫氏埃希菌:氨苄西林、替卡西林。
6、蜂房哈夫尼菌:氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)。
7、摩根摩根菌:氨苄西林、阿莫西林/克拉维酸、头孢菌素I代(头孢唑啉、头孢噻吩)、头孢菌素II代(头孢呋辛)、四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
8、普通变形杆菌和彭氏变形杆菌:氨苄西林、头孢菌素I代(头孢唑啉、头孢噻吩)、头孢菌素II 代(头孢呋辛)、四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
9、奇异变形杆菌:四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
此菌对青霉素和头孢菌素没有天然耐药性。
10、粘质沙雷氏菌:氨苄西林、阿莫西林/克拉维酸、氨苄西林/舒巴坦、头孢菌素I代(头孢唑啉、头孢噻吩)、头霉素类(头孢西丁、头孢替坦)、头孢菌素II代(头孢呋辛)、呋喃妥因、多粘菌素B、黏菌素。
11、小肠结肠炎耶尔森菌:氨苄西林、阿莫西林/克拉维酸、替卡西林、头孢菌素I代(头孢唑啉、头孢噻吩)。
12、沙门氏菌和志贺氏菌:此菌对β-内酰胺类药物无天然耐药,一代、二代头孢菌素和头霉素在体外可显示活性,但临床无效,不能报告为敏感。
13、雷氏普罗维登斯菌和斯图普罗威登斯菌:氨苄西林、阿莫西林/克拉维酸、头孢菌素I代(头孢唑啉、头孢噻吩)、四环素类/替加环素、呋喃妥因、多粘菌素B、黏菌素。
铜绿假单胞菌 耐药基因

铜绿假单胞菌耐药基因
铜绿假单胞菌(Pseudomonas aeruginosa)是一种常见的耐药细菌。
它具有多种机制来抵抗常规抗生素的作用。
以下是一些铜绿假单胞菌中常见的耐药基因:
1. β-内酰胺酶基因(β-lactamase genes):铜绿假单胞菌产生各类β-内酰胺酶,如TEM、SHV和CTX-M等,这些酶能降解β-内酰胺类抗生素,如青霉素和头孢菌素。
2. 美罗培南酶基因(Metallo-beta-lactamase genes):铜绿假单胞菌中也存在一类特殊的β-内酰胺酶,称为金属β-内酰胺酶(MBL),如NDM、IMP和VIM等。
这些酶能降解包括美罗培南在内的广谱抗生素。
3. 氨基糖苷酶基因(Aminoglycoside-modifying enzyme genes):铜绿假单胞菌可产生氨基糖苷酶,例如AAC、ANT和APH等。
这些酶能够修饰和降解氨基糖苷类抗生素,如庆大霉素和阿米卡星。
4. 磺胺类药物耐药基因(Sulfonamide resistance genes):铜绿假单胞菌可以通过携带sul等基因来获得对磺胺类药物的耐药性。
5. 氟喹诺酮耐药基因(Fluoroquinolone resistance genes):铜绿假单胞菌中常见的氟喹诺酮耐药基因包括gyrA和parC,突变这些基因会导致细菌对氟喹诺酮类抗生素产生耐药性。
请注意,这只是一些常见的耐药基因,并不代表全部。
铜绿假单胞菌具有高度的遗传多样性,因此可能存在其他未列出的耐药基因。
铜绿假单胞菌的基因组分析及耐药机制研究

铜绿假单胞菌的基因组分析及耐药机制研究铜绿假单胞菌(Pseudomonas aeruginosa)是一种广泛存在于自然环境中的革兰氏阴性杆菌,是人类与动植物的常规共生菌之一。
然而,铜绿假单胞菌也是一种常见的致病菌,引发了许多医院获得性感染和呼吸道感染,对人类健康构成了威胁。
除此之外,铜绿假单胞菌还具有很高的耐药性,尤其是多重耐药性,使得其治疗变得困难。
为了更好地了解铜绿假单胞菌的基因组及耐药机制,科学家们进行了深入的研究。
在基因组分析方面,科学家们采用了高通量测序技术,通过测序铜绿假单胞菌的基因组,获得了该菌株的完整基因组序列。
同时,通过进行基因组比对和功能注释,科学家们发现了一些与耐药性相关的基因。
首先,科学家们发现了一些铜绿假单胞菌耐药性的基因。
这些基因编码了一系列的药物转运蛋白,能够将抗生素从菌体内外排除,从而减少了抗生素的作用。
此外,还发现了一些编码抗生素降解酶的基因,这些酶能够将抗生素分解为无活性的代谢产物,从而降低抗生素的疗效。
其次,科学家们还发现了一些与铜绿假单胞菌耐药性相关的突变。
这些突变可能是导致菌株耐药性增加的重要因素。
例如,一些突变可能导致菌株产生特定蛋白质的点突变,从而改变了该蛋白的结构或功能,使得抗生素难以对其起效。
此外,还发现了一些突变可能导致铜绿假单胞菌耐药性突变的调控基因的突变。
这些调控基因在细菌的基因表达调控中起到重要的作用,突变会导致整个基因网络的改变,进而影响耐药性的形成。
此外,铜绿假单胞菌的耐药性还与其生物膜形成密切相关。
铜绿假单胞菌具有形成生物膜的能力,生物膜能够保护细菌免受外界环境的侵害,从而增加了菌株对抗生素的耐药性。
科学家们发现了一些与生物膜形成相关的基因,并对这些基因进行了研究。
通过研究生物膜形成相关的基因,科学家们希望能够找到新的药物靶点,从而提供新的耐药性干预策略。
综上所述,铜绿假单胞菌的基因组分析及耐药机制研究为我们深入了解该菌株的抗药性提供了重要的线索。
eagle现象及细菌天然耐药

Eagle现象的机制和影响
Eagle现象的产生机制涉及到 细菌细胞壁的通透性、外排泵 的表达以及DNA修复等多个方
面。
Eagle现象的存在使得一些原 本对抗生素敏感的细菌逐渐 获得耐药性,从而对临床治
疗造成极大的挑战。
了解和掌握Eagle现象的机制 有助于开发新的抗菌药物和治 疗方法,以应对细菌耐药性的
对这一全球性的健康问题。
04
应对Eagle现象和细菌天然耐药性的
策略
研发新型抗菌药物
抗菌药物创新
通过研究新的抗菌药物靶点、药 物设计和合成方法,开发出具有 新作用机制和抗菌谱的新型抗菌 药物。
抗菌药物组合
研究抗菌药物的联合使用,通过 药物间的协同作用增强抗菌效果, 降低耐药性的产生。
抗菌药物修饰
细菌天然耐药性对Eagle现象的影响
耐药细菌更容易发生Eagle现象
具有天然耐药性的细菌可能更容易受到Eagle现象的影响,因为这些细菌的细胞膜通透性和代谢状态 可能与Eagle现象的发生和发展有关。
耐药基因可能影响Eagle现象的表现
细菌的耐药基因可能通过影响细胞膜成分或细胞内代谢途径等方式,影响Eagle现象的表现和严重程 度。
对现有抗菌药物进行结构修饰, 以提高其抗菌活性、降低耐药性 产生和改善药物代谢动力控制抗菌药物使用
制定和实施抗菌药物使用指南,规范临床医 生用药行为,减少不必要的抗菌药物使用。
实施抗菌药物分级管理
根据抗菌药物的疗效、安全性、耐药性等特点,将 抗菌药物分为不同级别,限制高级别抗菌药物的滥 用。
• 总结:在使用抗菌药物时,应根据药敏试验结果选择最敏感的药物,避免盲目 使用广谱抗菌药物。同时,需要控制抗菌药物的用量和使用时间,以降低耐药 性的发生和发展。
香港海鸥型菌四环素耐药性及其耐药分子机制研究

保存 , 并作为药敏试验的质控菌株。 1 . 2 方 法 1 . 2 . 1 香 港海 鸥 型菌 的分 离 与鉴 定 参 照文 献 『 7 ] ,用选择性培养基分离 、生化鉴定和 1 6 S r R N A 基因鉴定 的方法分离出香港海鸥型菌 。 1 . 2 . 2 四环素类抗 生素 的耐药性测定 采用纸 片 扩散法测定香港海鸥型菌分离株对 四环素 、 多西环 素和米诺环素的耐药性 , 其耐药性判断按照文献 [ 8 ] 的方法 , 参考肠杆菌科判定标准进行 。 1 . 2 . 3 四环素耐药基 因 t e t A、 t e t B 、 t e t C 、 t e t M和 t e t O 的检测 参照文献 [ 9 ] , 分别合成 5 种 四环 素耐药 基 因 的引物 。用 煮沸 法提 取 四环 素耐 药株 的总 D N A, P C R反应 条件 为 : 9 4 ℃预变 性 5 m i n : 9 4 c c 变 性3 0 s , 5 2 c c 退火 3 0 S , 7 2 %延伸 1 m i n , 3 5 个循环 : 最后 7 2 %延伸 5 m i n 。取 5 L扩增产物于 1 . 5 %琼 脂糖凝胶 电泳后在凝胶成像 系统中检测。 1 . 2 . 4 完整 t e t A基 因及其阻遏基因 t e t R的扩增和 测序 将 t e t A基 因检 i 贝 4 阳性 的 四 环 素耐 药 株 进 行 完整 t e t A基 因及 其阻遏基 因 t e t R的扩增和测 序 根据 文献 [ 报道 的 p H L H K 2 2质 粒 上 的 t e t A 和 t e t R基 因序列设计 引物 , t e t A基 因的引 物为 t e t A .
与 1 6 S r R N A 基 因 鉴 定 的 阳 性 对 照 。 大 肠 杆 菌 A T C C 2 5 9 2 2和金 黄 色葡 萄 球 菌 A T C C 2 5 9 2 3由本 系
eagle现象及细菌天然耐药正式版

▪ 交叉或协同耐药检测失败 ➢ 肠杆菌科对奈替米星I或R,但对妥布霉素为S,协同耐药检测失败 ➢ 链球菌对青霉素G为I或R,但头孢菌素一代=S,交叉耐药失败
如下情况出现要复查
▪ 罕见表型
罕见耐药
▪ 金黄色葡萄球菌对糖肽类抗生素的耐药株,
➢ 目前中国尚未发现
▪ 金黄色葡萄球菌对利萘唑胺和奎奴普丁的耐药株及凝固酶阴性葡萄球 菌对万古霉素或对利萘唑胺耐药, 均应予以高度注意。
▪ 肺炎链球菌对美洛培南、万古霉素、替考拉宁和利萘唑胺任一药物均 可产生耐药。
▪ A 、B 、C、G群β-溶血链球菌对青霉素、万古霉素、替考拉宁、利 萘唑胺任一药物均可产生耐药。
▪ mecillinam and Providencia stuartii
➢ Kerry, Hamilton-Miller & Brumfitt, 1976
▪ cefotaxime and Staph.aureus and Pseudomonas aeruginosa
➢ Shah,Troche & Stille, 1979
eagle现象及细菌天然耐药
J. Exp. Med., Jul 1948; 88: 99 - 131
Paradoxical effect (Eagle phenomenon)
▪ 指抗菌药物在高浓度的时候反而降低杀菌活性 ▪ 这种现象第一次由Eagle和Musselman两位科学 Stille & Uffelman,1973; Yourassowsky, Van der Linden & Schoutens, 1976)
acutalibacteraceae 中文 -回复

acutalibacteraceae 中文-回复acutalibacteraceae(刺味杆菌科)是细菌界疣状球菌纲的一科,包括一系列产酸菌属,这些菌属能够在特定环境中引起酸性反应。
刺味杆菌科的成员广泛分布于自然环境中,包括土壤、水体和食品中,它们对人类和动物的健康产生了重要的影响。
刺味杆菌科菌属的特点:1. 菌落形态:一般为乳白色,并产生酸状物质,可以使培养基pH值下降。
2. 革兰氏染色:菌体为革兰氏阳性菌。
3. 形态特征:通常为杆状,有时呈细长双钩状。
4. 代谢特点:为产酸菌,能够通过代谢过程产生大量有机酸,如乳酸、丙酸等。
刺味杆菌科有着广泛的生态和生理特性。
它们存在于许多环境中,包括土壤、水体、植物、动物和人体等。
某些菌属如Acidobacterium,能够利用根系分泌的有机酸作为能源,从而与植物形成共生关系。
同时,刺味杆菌科中的一些菌属如Acetobacter、Gluconacetobacter、Komagataeibacter等,能够参与食品和饮料的发酵过程,产生有利于食物品质和口感的有机酸,如醋酸和柠檬酸等。
此外,刺味杆菌科中的一些菌属也与人类和动物的健康产生重要的关联。
例如,Achromobacter xylosoxidans是人类常见的呼吸道和泌尿道感染病原菌之一,能够导致肺部感染和尿路感染。
而Acetobacter genus有一些菌株被发现在蜜蜂肠道中产酸,帮助维持蜜蜂肠道微生物平衡。
刺味杆菌科菌属的酸性代谢机制有着广泛的研究。
这些菌属通过代谢过程中产生的有机酸调节周围环境的pH值,从而改变其生态环境,获得生存竞争的优势。
酸性环境对许多其他菌属的生长和代谢具有抑制作用,因此刺味杆菌科成员的代谢特点为其提供了适应不同环境和竞争的能力。
刺味杆菌科在农业、食品工业和医学领域具有广泛应用。
举例来说,酸性乳酸菌用于发酵制作酸奶和乳酸饮料,具有益生菌功能,有助于肠道健康;酸性杆菌在食品保鲜和储藏过程中,通过酸性环境抑制一些食源性病原菌的生长;在医学研究中,刺味杆菌科的代谢特点和与宿主的共生关系被广泛研究,为研发新的药物和治疗方法提供了理论基础。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
析,为临床医师提供合理的抗感染治疗建议。因此,了解国际 上未曾报道或极为罕见的耐药谱型具有重要意义。
罕见耐药
金黄色葡萄球菌对糖肽类抗生素的耐药株,
目前中国尚未发现
金黄色葡萄球菌对利萘唑胺和奎奴普丁的耐药株及凝固酶阴性葡萄球 菌对万古霉素或对利萘唑胺耐药, 均应予以高度注意。 肺炎链球菌对美洛培南、万古霉素、替考拉宁和利萘唑胺任一药物均 可产生耐药。 A 、B 、C、G群β -溶血链球菌对青霉素、万古霉素、替考拉宁、利 萘唑胺任一药物均可产生耐药。 不动杆菌属和铜绿假单胞菌对多粘菌素E耐药 淋病奈瑟菌对第3 代头孢菌素耐药 肠杆菌科细菌对美洛培南耐药 如在常规药敏试验中发现上述耐药谱型, 应对分离菌种行进一步鉴定并 重复药敏 试验, 如得出的结果仍相同,应将菌株送到参考实验室进行确证。下情况出现要复查
葡萄球菌:苯唑西林
R 红霉素 S 全部氨基糖甙类 S 喹诺酮 S 肠杆菌对亚胺培南或美罗培南耐药
如下情况出现要复查
天然耐药检测失败
阴沟肠杆菌、产气肠杆菌和弗劳地枸掾酸
杆菌对头孢西丁=S 奇异变形杆菌对呋喃妥因和四环素S 小肠结肠炎耶尔森菌对先锋1号、氨基青霉 素=S
阿米卡星对大肠埃希菌的抑菌现象
J. Exp. Med., Jul 1948; 88: 99 - 131
Paradoxical effect (Eagle phenomenon)
指抗菌药物在高浓度的时候反而降低杀菌活性
这种现象第一次由Eagle和Musselman两位科学 家发现并报道。
青霉素类抗菌药物在作用于葡萄球菌、β溶血性 链球菌以及肠球菌时出现药物低浓度(仍超过 MIC值)时抗菌活性好于高浓度。
J. Exp. Med., Jul 1948; 88: 99 - 131
有关Eagle现象的报道
ampicillin and Streptococcus faecalis
Stille & Uffelman,1973; Yourassowsky, Van der Linden &
这是一株大肠杆菌的药敏结果 请大家分析其耐药机制
这份报告是否存在问题?
谢谢大家
Thank you
交叉或协同耐药检测失败 肠杆菌科对奈替米星I或R,但对妥布霉素为S,协同耐药检测失败 链球菌对青霉素G为I或R,但头孢菌素一代=S,交叉耐药失败
如下情况出现要复查
罕见表型
葡萄球菌对苯唑西林S,但对红霉素、四
环素和林可霉素I或R。多重耐药常耐苯唑 西林。 葡萄球菌:
-内酰胺酶(-)
cefotaxime and Staph.aureus and Pseudomonas aeruginosa
Shah,Troche & Stille, 1979
Aminoglycosides and Gram-negative bacilli
Lorian el ai,1979
药物:青霉素 细菌:化脓性链球菌
药物:青霉素G 细菌:B群β溶血性链球菌
药物: 青霉素G 细菌: B群β溶血性链球菌
药物:青霉素G 细菌:粪肠球菌
药物:青霉素G 细菌:粪肠球菌
药物:青霉素
细菌:肺炎链球菌
药物:青霉素
细菌:金葡菌
小结
对于大部分药物和细菌来说,随着药物浓度的升高,其抗菌活性也相 应增强。最高药物浓度的抗菌活性比最低有效浓度(敏感浓度)增强 2-20倍。甚至有的能增加32000倍。 抗菌药物对少数菌株在高浓度时的抗菌活性反而不如低浓度,提示抗
菌药物对这些细菌的最佳有效浓度范围处于一个相对窄的空间。
不同种类细菌之间有较大的差异,同一菌种的不同亚种之间也会有差 异。
意义
天然或固有耐药的菌属或菌种
有些菌属和菌种对某些抗菌药物天然耐药或固有耐药, 该耐
药特性具有种属特异性。因此,若药敏试验的结果为“敏感” 即应予以怀疑, 有必要重复药敏试验和重新鉴定菌种。 罕见耐药谱型
Schoutens, 1976)
carbenicillin and Proteus mirabilis
Yourarsowsky, Van der Linden & Schoutens, 1976
mecillinam and Providencia stuartii
Kerry, Hamilton-Miller & Brumfitt, 1976
如下情况出现要复查
天然耐药检测失败 鹑鸡肠球菌或铅黄肠球菌对万古霉素=S 交叉耐药检测失败 假单胞菌属对哌拉西林I或R,但对氨基青霉素 =S 替卡西林=S 在革兰阴性卡中,对头孢三代、四代为I或R, 但对二代头孢菌素、头霉菌素、头孢泊肟、氨 基青霉素为S
如下情况出现要复查
革兰阴性杆菌,除外绿脓,对哌拉西林I或R,但对氨基青 霉素和羧基青霉素S 待测细菌对常规测试药物全部耐药 矛盾的结果: 革兰阴性杆菌对第三代、四代头孢菌素=R,但对一代和 二代头孢、头孢克肟、头孢泊肟、氨基青霉素为S 肠杆菌对单剂抗生素为S,但对含酶抑制剂的复方制剂为R 革兰阴性杆菌对第一代喹诺酮为S,但对环丙沙星等为R