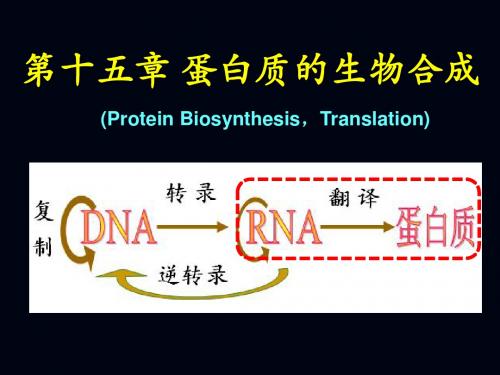
第十五章 DNA、RNA和蛋白质的生物合成
第十五章蛋白质的生物合成

第一位
(5ˊ)
U
U
C
A
G
遗传密码字典 第二位
C
A
G
第三位
(3ˊ)
U C A G
U C A G
U C A G
U C A G
6 4 组 密 码 子 中 , AUG 既 是 甲 硫氨酸的密码,又是起始密码; 有三组密码不编码任何氨基酸, 而是多肽链合成的终止密码子: UAG、UAA、UGA。
• 在原核生物和真核生物中,均存在另一 种携带蛋氨酸的tRNA,识别非起动部位 的蛋氨酸密码,AUG。
第十五章蛋白质的生物合成
核糖体的研究历史
1.早在本世纪30年代后期就发现细胞质和细胞核中都有 核酸存在,不过用1924年福尔根发明的染色法只能使细胞核 中的核酸染色。但两种核酸在260nm的吸收非常相似。
第十五章 蛋白质合成及转运
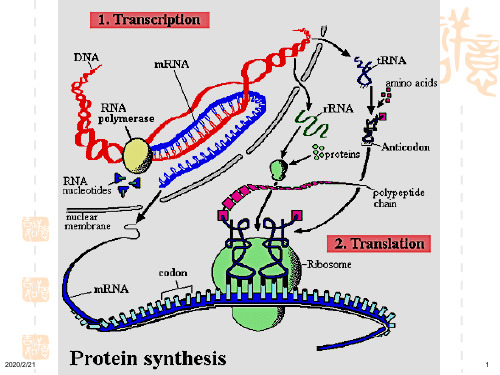
蛋白质的生物合成
中心法则指出,遗传信息的表达最终是合成出 具有特定氨基酸顺序的蛋白质,这种以mRNA上所 携带的遗传信息,到多肽链上所携带的遗传信息的传 递,就好象以一种语言翻译成另一种语言时的情形 相似,所以称以mRNA为模板的蛋白质合成过程为 翻译(translation)。
2.Crick, F.比较了核酸和氨基酸的大小和形状后,认为不可 能在空间上互补,因此预测:(1) 存在一类分子转换器,使信 息从核酸序列转换成氨基酸序列;(2) 这种分子很可能是核酸; (3) 它不论以何种方式进入蛋白质翻译系统的模板,都必须与 模板形成氢键(即配对);(4) 有20种分子转换器,每种氨基 酸一个;(5) 每种氨基酸必定还有一个对应的酶,催化与特定 的分子转换器结合。
2.1941年,细胞学家J.Brachet和T.Caspersor注意到细 胞质中的核酸与蛋白质的合成有密切的关系。
第十五章 蛋白质的生物合成-翻译

二、tRNA
tRNA是氨基酸的搬运工具。 tRNA是多肽链和mRNA之间的 重要转换器。 每一种氨基酸可以有一种以上 tRNA作为运载工具。 能够携带相同氨基酸而反密码子 不同的一组tRNA分子称为氨基 酸的同工受体tRNA (isoaccepting tRNA) 。
tRNA须具备的功能 • 与氨基酸结合(3’末端) • 识别特异的氨酰-tRNA合成酶(D环) • 识别mRNA链上密码子 • 与核糖体结合,使延长中的肽链附着于核糖体上(TψC环)
蛋白质生物合成过程包括: 1. 氨基酸的活化; 2. 合成起始; 3. 肽链延伸:进入、转肽、移位; 4. 终止合成。
一、氨基酸的活化
二、合成的起始阶段
核糖体大小亚基分离; mRNA在小亚基定位结合; 起始氨酰tRNA的结合; 核糖体大亚基结合。
1. 核糖体大、小亚基分离 IF1和IF3与30S小亚基结合,促进核糖体大、小
翻译过程实际上就是由tRNA携带着氨基酸,逐一识别 mRNA上的密码子,并将氨基酸依密码子的排序相互 连接的过程。核糖体是翻译的场所。
一、mRNA模板和遗传密码
• mRNA是翻译的直接模板。 (一)遗传密码的破译
mRNA上四种核苷酸→组成蛋白质的20种aa
核苷酸与氨基酸对应关系?
3个相邻的核苷酸→1个aa, 有43种排列→64种密码子
• 核糖体可以看作是一个大分子的机构,它具有许多精密的 配合部分,来挑选并管理参与蛋白质合成的各个组分。它 参与多肽链的启动、延伸和终止的各种因子的识别。
原核生物核糖体
5S rRNA, 23S rRNA 50S
34种蛋白质 70S
16S rRNA 30S
21种蛋白质
真核生物核糖体
5SrRNA,5.8SrRNA,28SrRNA 60S
RNA生物合成

1、转录的不对称性
转录(transcription)的不对称性就是 指以双链DNA中的一条链作为模板进行 转录,从而将遗传信息由DNA传递给 RNA。 对于不同的基因来说,其转录信息可以 存在于两条不同的DNA链上。
能够转录RNA的那条DNA链称为模板链, 也称作反义链或负链。
与模板链互补的另一条DNA链称为编码链 (coding strand),也称为有义链或正链。
4、有特定的起始和终止位点
RNA转录合成时,只能以DNA分子中的某 一段作为模板,故存在特定的起始位点和特 定的终止位点。 特定起始点和特定终止点之间的DNA链构 成一个转录单位,通常由转录区和有关的调 节顺序构成。
二、参与RNA转录合成的物质
➢原料:NTP (ATP, UTP, GTP, CTP)。 ➢模板:单链DNA。 ➢酶:RNA聚合酶(DDRP,RNA-pol)。 ➢其他蛋白质因子:如转录因子、终止因子等。
1. 5’端加帽
5’端:m7GpppGpN(甲基化三磷酸双鸟苷) 部位:细胞核 作用: 1)使mRNA免受磷酸酶和核酸酶的攻击,
稳定mRNA分子的一级结构; 2)提供核蛋白体识别位点,促进翻译起
始复合物形成,增强mRNA翻译效率; 3)有利于mRNA前体的剪接。
mRNA 的5’帽子结构
5 pppGp…
帽 子 结 构 的 生 成
磷酸酶 5 ppGp…
Pi
pppG
鸟苷酸
转移酶
ppi
5 GpppGp…
SAM
甲基转移酶
5 m7GpppGp…
2.3’末端的产生和多聚腺甘酸化
3’端:20-200个腺苷酸残基 部位:细胞核 作用: 1)维持mRNA的稳定;
2)增加翻译效率; 3)与mRNA运输有关。
分子生物学第十五章 蛋白质的生物合成

1.遗传密码种类:
• mRNA 分子有4种碱基:A、G、C、U,可组合成64个密 码子,其中61个分别代表20种不同氨基酸
• 遗传密码共有43=64种, 64: UAA、UAG、UGA 终止密码 61: AUG 起始密码 代表Met 60: 代表19种氨基酸
9
遗传密码表 第一碱基
(5/-端)
第二碱基
43
一、起始阶段
(一)原核生物翻译起始复合物的形成
参与的物质: 核糖体50S和30S大小两类亚基 mRNA 起始作用的fMet-tRNAfMet GTP供能 起始因子(IF1、IF2、IF3)
44
起始过程:
• 核糖体大小亚基解离 • 小亚基与mRNA结合:
16sRNA识别mRNA的SD序列
• fMet-tRNAfMet与mRNA起始密码子AUG结合 • 核糖体大小亚基形成起始复合物
tRNAphe Phe-tRNAphe
• 肽链起始和延长的甲硫氨酰-tRNA:
真核生物:起始: Met-tRNAiMet 延长: Met-tRNAMet
原核生物: 起始:fMet-tRNAfMet 延长:Met-tRNAMet
39
40
第三节 肽链的合成过程
41
整个翻译过程可分为 :
• 翻译的起始 • 翻译的延长 • 翻译的终止
tRNA反密码子 第1位碱基
mRNA密码子 第3位碱基
I
U
G AC
U, C, A A, G U, C U G
17
摆 动 配 对
32 1
U
123
18
• ⑤遗传密码的通用性
从原核生物到人类都共用同一套遗传密码,被称为遗传密码的通用性。 密码的通用性进一步证明各种生物进化自同一祖先。
蛋白质的生物合成习题与参考答案

第十五章蛋白质生物合成一、填空题:1.三联体密码子共有64 个,其中终止密码子共有 3 个,分别为UAA 、UAG 、UGA ; 2.密码子的基本特点有四个分别为从5′→3′无间断性、简并性、变偶性、通用性; 3.次黄嘌呤具有广泛的配对能力,它可与U 、 C 、 A 三个碱基配对,因此当它出现在反密码子中时,会使反密码子具有最大限度的阅读能力;4.原核生物核糖体为70 S,其中大亚基为50 S,小亚基为30 S;而真核生物核糖体为80 S,大亚基为60 S,小亚基为40 S;5.原核起始tRNA,可表示为tRNA f甲硫,而起始氨酰tRNA表示为f Met-tRNA f甲硫;真核生物起始tRNA可表示为tRNA I甲硫,-tRNA表示为Met-tRNA f甲硫;6.肽链延伸过程需要进位、转肽、移位三步循环往复,每循环一次肽链延长 1 个氨基酸残基,原核生物中循环的第一步需要EF-Tu 和EF-Ts 延伸因子;第三步需要EF-G 延伸因子;7.原核生物mRNA分子中在距起始密码子上游约10个核苷酸的地方往往有一段富含嘌呤碱基的序列称为Shine-Dalgrano序列,它可与16S-rRNA 3′-端核苷酸序列互补;8.氨酰-tRNA的结构通式可表示为:OtRNA-O-C-CH-RNH2,与氨基酸键联的核苷酸是A腺嘌呤核苷酸;9.氨酰-tRNA合成酶对氨基酸和相应tRNA都具有较高专一性,此酶促反应过程中由ATP 水解提供能量;10.肽链合成的终止阶段, RF1因子和RF2因子能识别终止密码子,以终止肽链延伸,而RF3因子虽不能识别任何终止密码子,但能协助肽链释放;11.蛋白质合成后加工常见的方式有磷酸化、糖基化、脱甲基化、信号肽切除; 12.真核生物细胞合成多肽的起始氨基酸为甲硫氨酸,起始tRNA为tRNA I甲硫,此tRNA 分子中不含T C 序列;这是tRNA家庭中十分特殊的;二、选择题只有一个最佳答案:1.下列有关mRAN的论述,正确的一项是 CA、mRNA是基因表达的最终产物B、mRNA遗传密码的阅读方向是3′→5′C、mRNA遗传密码的阅读方向是5′→3′D、mRNA密码子与tRNA反密码子通过A-T,G-C配对结合E、每分子mRNA有3个终止密码子2.下列反密码子中能与密码子UAC配对的是 DA、AUGB、AUIC、ACUD、GUA3.下列密码子中,终止密码子是 BA、UUAB、UGAC、UGUD、UAU4.下列密码子中,属于起始密码子的是 AA、AUGB、AUUC、AUCD、GAG5.下列有关密码子的叙述,错误的一项是 CA 、密码子阅读是有特定起始位点的B 、密码子阅读无间断性C 、密码子都具有简并性D 、密码子对生物界具有通用性6.密码子变偶性叙述中,不恰当的一项是 AA 、密码子中的第三位碱基专一性较小,所以密码子的专一性完全由前两位决定B 、第三位碱基如果发生了突变如A G 、C U,由于密码子的简并性与变偶性特点,使之仍能翻译出正确的氨基酸来,从而使蛋白质的生物学功能不变C 、次黄嘌呤经常出现在反密码子的第三位,使之具有更广泛的阅读能力,I-U 、I-C 、I-A 从而可减少由于点突变引起的误差D 、几乎有密码子可用U C XY 或U C XY 表示,其意义为密码子专一性主要由头两个碱基决定7.关于核糖体叙述不恰当的一项是 BA 、核糖体是由多种酶缔合而成的能够协调活动共同完成翻译工作的多酶复合体B 、核糖体中的各种酶单独存在解聚体时,同样具有相应的功能C 、在核糖体的大亚基上存在着肽酰基P 位点和氨酰基A 位点D 、在核糖体大亚基上含有肽酰转移酶及能与各种起始因子,延伸因子,释放因子和各种酶相结合的位点8.tRNA 的叙述中,哪一项不恰当 DA 、tRNA 在蛋白质合成中转运活化了的氨基酸B 、起始tRNA 在真核原核生物中仅用于蛋白质合成的起始作用C 、除起始tRNA 外,其余tRNA 是蛋白质合成延伸中起作用,统称为延伸tRNAD 、原核与真核生物中的起始tRNA 均为fMet-tRNA9.tRNA 结构与功能紧密相关,下列叙述哪一项不恰当 DA 、tRNA 的二级结构均为“三叶草形”B 、tRNA3′-末端为受体臂的功能部位,均有CCA 的结构末端C 、T C 环的序列比较保守,它对识别核糖体并与核糖体结合有关D 、D 环也具有保守性,它在被氨酰-tRNA 合成酶识别时,是与酶接触的区域之一10.蛋白质生物合成中多肽的氨基酸排列顺序取决于 CA 、相应tRNA 的专一性B 、相应氨酰tRNA 合成酶的专一性C 、相应mRNA 中核苷酸排列顺序D 、相应tRNA 上的反密码子11.下列有关氨酰- tRNA 合成酶叙述中,哪一项有误 CA 、氨酰-tRNA 合成酶促反应中由ATP 提供能量,推动合成正向进行B 、每种氨基酸活化均需要专一的氨基酰- tRNA 合成酶催化C 、氨酰-tRNA 合成酶活性中心对氨基酸及tRNA 都具有绝对专一性OD 、该类酶促反应终产物中氨基酸的活化形式为R -CH -C -O -ACC -tRNANH 212.原核生物中肽链合的起始过程叙述中,不恰当的一项是 DA 、mRNA 起始密码多数为AUG,少数情况也为GUGB 、起始密码子往往在5′-端第25个核苷酸以后,而不是从mRNA5′-端的第一个苷酸开始的C 、在距起始密码子上游约10个核苷酸的地方往往有一段富含嘌呤的序列,它能与16SrRNA3′-端碱基形成互补D、70S起始复合物的形成过程,是50S大亚基及30S小亚基与mRNA自动组装的13.有关大肠杆菌肽链延伸叙述中,不恰当的一项是 CA、进位是氨酰-tRNA进入大亚基空差的A位点B、进位过程需要延伸因子EFTu及EFTs协助完成C、甲酰甲硫氨酰-tRNA f进入70S核糖体A位同样需要EFTu-EFTs延伸因子作用D、进位过程中消耗能量由GTP水解释放自由能提供14.移位的叙述中哪一项不恰当 CA、移位是指核糖体沿mRNA5′→3′作相对移动,每次移动的距离为一个密码子B、移位反应需要一种蛋白质因子EFG参加,该因子也称移位酶C、EFG是核糖体组成因子D、移位过程需要消耗的能量形式是GTP水解释放的自由能15.蛋白质生物合成的方向是: BA、从C端到N端B、从N端到C端C、定点双向进行D、从C端、N端同时进行16.在蛋白质合成过程中,下列哪些说法是正确的 CA、氨基酸随机地连接到tRNA上去B、新生多肽链的合成都是从C-端向N-端方向延伸的C、通过核糖核蛋白体的收缩,mRNA不断移动D、肽键形成是由肽酰转移酶作用下完成的,此种酶不属于核糖体的组成成分17.70S起始复合物的形成过程的叙述,哪项是正确的 DA、mRNA与30S亚基结合过程需要超始因子IF1B、mRNA与30S亚基结合过程需要超始因子IF2C、mRNA与30S亚基结合过程需要超始因子IF3D、mRNA与30S亚基结合过程需要超始因子IF1、IF2和IF318.mRNA与30S亚基复合物与甲酰甲硫氨酰-tRNA f结合过程中起始因子为 AA、IF1及IF2B、IF2及IF3C、IF1及IF3D、IF1、IF2及IF319.原核细胞中氨基酸掺入多肽链的第一步反应是: DA、甲酰蛋氨酸-tRNA与核蛋白体结合B、核蛋白体30S亚基与50S亚基结合C、mRNA与核蛋白体30S亚基结合D、氨酰tRNA合成酶催化氨基酸活化20.假设翻译时可从任一核苷酸起始读码,人工合成的AACnn为任意整数多聚核苷酸,能够翻译出几种多聚氨基酸 CA、一种B、二种C、三种D、四种21.绝大多数真核生物mRNA5’端有 AA、帽子结构B、PolyAC、起始密码D、终止密码22.能与密码子ACU相识别的反密码子是DA、UGAB、IGAC、AGID、AGU23.原核细胞中新生肽链的N-末端氨基酸是CA、甲硫氨酸B、蛋氨酸C、甲酰甲硫氨酸D、任何氨基酸24.tRNA的作用是 DA、把一个氨基酸连到另一个氨基酸上B、将mRNA连到rRNA上C、增加氨基酸的有效浓度D、把氨基酸带到mRNA的特定位置上25.细胞内编码20种氨基酸的密码子总数为: DA、16B、64C、20D、6126.下列关于遗传密码的描述哪一项是错误的CA、密码阅读有方向性,5'-端开始,3'-端终止B、密码第3位即3′-端碱基与反密码子的第1位即5′-端碱基配对具有一定自由度,有时会出现多对一的情况C、一种氨基酸只能有一种密码子D、一种密码子只代表一种氨基酸27.蛋白质合成所需的能量来自CA、ATPB、GTPC、ATP和GTPD、CTP28.下列关于氨基酸密码的描述哪一项是错误的 AA、密码有种属特异性,所以不同生物合成不同的蛋白质B、密码阅读有方向性,5′-端起始,3′-端终止C、一种氨基酸可有一组以上的密码D、一组密码只代表一种氨基酸29.mRNA的5′-ACG-3′密码子相应的反密码子是 CA、5′-UGC-3′B、5′-TGC-3′C、5′-CGU-3′D、5′-CGT-3′30.下列哪一个不是终止密码 BA、UAAB、UACC、UAGD、UGA三、是非题在题后括号内打√或×:1、蛋白质生物合成所需的能量都由ATP直接供给; ×2、反密码子GAA只能辨认密码子UUC; ×3、生物遗传信息的流向,只能由DNA—→RNA而不能由RNA—→DNA; ×4、原核细胞新生肽链N端第一个残基为fMet,真核细胞新生肽链肽链N端第一个氨基酸残基为Met; √5、DNA复制与转录的共同点在于都是以双链DNA为模板,以半保留方式进行,最后形成链状产物; ×6、依赖DNA的RNA聚合酶叫转录酶,依赖于RNA的DNA聚合酶即反转录酶; √7、密码子从5’-端至3’-端读码,而反密码子则从3’-端至5’-端读码; ×8、一般讲,从DNA的三联体密码子中可以推定氨基酸的顺序,相反从氨基酸的顺序也可毫无疑问地推定DNA顺序; ×9、DNA半不连续复制是指复制时一条链的合成方向是5′→3′而另一条链方向是3′→5′; ×10、真核生物蛋白质合成起始氨基酸是N-甲酰甲硫氨酸; ×11、原核细胞的DNA聚合酶一般都不具有核酸外切酶的活性; ×12、在具备转录的条件下,DNA分子中的两条链在体内都可能被转录成RNA; ×13、核糖体是细胞内进行蛋白质生物合成的部位; √14、mRNA与携带有氨基酸的tRNA是通过核糖体结合的; √15、核酸是遗传信息的携带者和传递者; √16、RNA的合成和DNA的合成一样,在起始合成前亦需要有RNA引物参加; ×17、真核生物mRNA多数为多顺反子,而原核生物mRNA多数为单顺反子; ×18、合成RNA时,DNA两条链同时都具有转录作用; ×19、在蛋白质生物过程中mRNA是由3’-端向5’-端进行翻译的; ×20、蛋白质分子中天冬酰胺,谷氨酰胺和羟脯氨酸都是生物合成时直接从模板中译读而来的; ×21、逆转录病毒RNA并不需要插入寄主细胞的染色体也可完成其生命循环; ×四、问答题:1.氨酰-tRNA合成酶在多肽合成中的作用特点和意义;答:氨基酰-tRNA合成酶具有高度的专一性:一是对氨基酸有极高的专一性,每种氨基酸都有一种专一的酶,它仅作用于L-氨基酸,不作用于D-氨基酸,有的氨基酸-tRNA合成酶对氨基酸的专一性虽然不很高,但对tRNA仍具有极高专一性;这种高度专一性会大大减少多肽合成中的差错;2.原核细胞与真核细胞蛋白质合成起始氨基酸起始氨基酰—tRNA及起始复合物的异同点有那些答:为了便于比较列表如下3.原核生物与真核生物mRNA的信息量及起始信号区结构上有何主要差异;答:为了便于比较列表如下:4.简述三种RNA在蛋白质生物合成中的作用;答:1 mRNA:DNA的遗传信息通过转录作用传递给mRNA,mRNA作为蛋白质合成模板,传递遗传信息,指导蛋白质合成;2 tRNA:蛋白质合成中氨基酸运载工具,tRNA的反密码子与mRNA上的密码子相互作用,使分子中的遗传信息转换成蛋白质的氨基酸顺序是遗传信息的转换器;3 rRNA:核糖体的组分,在形成核糖体的结构和功能上起重要作用,它与核糖体中蛋白质以及其它辅助因子一起提供了翻译过程所需的全部酶活性;五、名词解释:1.遗传密码与密码子多肽链中氨基酸的排列次序mRNA分子编码区核苷酸的排列次序对应方式称为遗传密码;而mRNA分子编码区中每三个相邻的核苷酸构成一个密码子;由四种核苷酸构成的密码子共64个,其中有三个不代表任何氨基酸,而是蛋白质合成中的终止密码子;2.起始密码子与终止密码子蛋白质合成中决定起始氨基酸的密码子称为起始密码子,真核与原核生物中的起始密码子为代表甲硫氨酸的密码子AUG和代表缬氨酸的密码子GUG;3.密码的简并性和变偶性一种氨基酸可以具有好几组密码子,其中第三位碱基比前两位碱基具有较小的专一性,即密码子的专一性主要由前两位碱基决定的特性称为变偶性;4.核糖体与多核糖体生物系统中合成蛋白质的部侠,称为核糖体;多聚核糖体:一条mRNA模板链可附着10-100个核糖体,这些核糖体依次结合起始密码子,沿5-3方向读码移动,同时进行肽链合成,这种mRNA与多个核糖体形成的聚合物称为多聚核糖体;5.同功tRNA、起始tRNA、延伸tRNA用于携带或运送同一种氨基酸的不同tRNA称同功tRNA,能特异识别mRNA上起始密码子的tRNA,称为起始tRNA;在肽链延伸过程中,用于转运氨基酸的tRNA称为延伸tRNA;6.EFTu-EFTs循环,移位,转肽肽键形成EF-Tu与EF-T S称为延伸因子,参与氨基酰-tRNA进位,每完成一次进位需要EF-Ts-EF-Tu循环一周,其过程如下:移位:就是核糖体沿着mRNA从5′向3′-端移动一个密码子的距离:转肽则是位于核糖体大亚基P 位点的肽酰基在转肽酶的作用下,被转移到A位点,氨在酰-tRNA的氨基上形成肽键的过程;7.信号肽几乎所有跨膜运送的蛋白质结构中,多数存在于N-末端的肽片段称为信号肽,其长度一般为15—35个氨基酸残基;它在蛋白质跨膜运送中起重要作用;少数信号肽位于多肽中间某个部位,称为“内含信号肽;”8.移码突变在mRNA分子编码区内插入一个或删除一个碱基,就会使这点以后的读码发生错误,这称为移码;由于移码引起的突变称为移码突变;。
第十五章 蛋白质的合成-答案

一、选择题1.与原核生物核糖体小亚基结合的蛋白质合成抑制剂是(A )A.链霉素B.氯霉素C.利福霉素D.放线菌素E.青霉素3.大肠杆菌合成的所有未修饰的多肽链,其N末端的氨基酸残基应是(C)A.甲硫氨酸B.丝氨酸C.甲酰甲硫氨酸D.甲酰丝氨酸E.谷氨酸4.某5岁儿童突发咽痛,呼吸急促,表现烦躁。
体温测得为38.5T,咽部检查见扁桃体肿大,甚至观察有循环系统衰竭的症状,诊断为咽白喉,白喉病产生的外毒素(白喉毒素)为致病的主要因素。
白喉毒素抑制蛋白质合成的主要机制是(E)A.降解核糖体大亚基的28SrRNA使其失活B.特异性结合40S亚基的A位干扰延长阶段的进位C.抑制肽酰转移酶从而抑制肽链延长D.与40S亚基结合影响翻译准确性E.使eEf-2失活从而阻断肽链延长6.小明因为受伤需要抹点红霉素,红霉素作为一种抗生素其作用原理是(C)A.阻止翻译起始复合物的形成B.抑制氨基酸tRNA与小亚基结合C.抑制肽酰转移酶D.引起读码错误E.抑制EF-G10.多肽链的延长与下列物质无关的是(B)A.肽酰转移酶B.甲酰甲硫氨酰-tRNAC.GTPD.mRNAE.EF-Tu、EF-Ts和EF-G11.肽键形成部位是(B)A.核糖体大亚基P位B.核糖体大亚基A位C.两者都是D.两者都不是E.核糖体大亚基E位13.氨基酰-tRNA合成酶的特点是(E)A.存在于细胞核内B.只对氨基酸的识别有专一性C.只对tRNA的识别有专一性D.催化反应需GTPE.对氨基酸、tRNA的识别都有专一性15.多数氨基酸都有两个以上密码子,下列氨基酸只有一个密码子的是(D)A.苏氨酸、甘氨酸B.脯氨酸、精氨酸C.丝氨酸、亮氨酸D.色氨酸、蛋氨酸E.天冬氨酸、天冬酰胺19.翻译起始复合物的组成(C)A.DNA模板+RNA+RNA聚合酶B.Dna蛋白+开键DNAC.核糖体+甲硫氨酰tRNA+mRNAD.翻译起始因子+核糖体E.核糖体+起始者tRNA20.下列关于核糖体的叙述,正确的是(B)A.是遗传密码的携带者B.由rRNA与蛋白质构成C.由snRNA与hnRNA构成D.由引物、DNA和蛋白质构成E.由tRNA与蛋白质构成21,下列关于密码子的叙述,正确的是(C)A.由DNA链中相邻的三个核苷酸组成B.由tRNA中相邻的三个核苷酸组成C.由mRNA上相邻的三个核苷酸组成D.由rRNA中相邻的三个核苷酸组成E.由多肽链中相邻的三个核苷酸组成22.遗传密码的简并性是指(C)A.密码子的第3位碱基决定编码氨基酸的特异性B.一个密码子可代表多个氨基酸C.多个密码子可代表同一氨基酸D.密码子与反密码子之间不严格配对E.所有生物可使用同一套密码23.下列关于遗传密码的叙述,正确的是(E)A.遗传密码只代表氨基酸B.一种氨基酸只有一个密码子C.一个密码子可代表多种氨基酸D.密码子与反密码子遵守严格的碱基配对原则E.密码子的简并性降低了基因突变的效应24.一个tRNA的反密码子为5,UGC3,,它可识别的密码子是(A )A.5'GCA3'B.5,ACG3'C.5'GCU3'D.5'GGC3'E.5'AUG3,25.氨基酸通过下列哪种化学键与tRNA进行特异结合(B)A.糖苷键B.酯键C.酰胺键D.磷酸酯键E.氢键26.蛋白质生物合成中氨基酸的活化与tRNA的结合需要(B )A.氨基酸tRNA合成酶B.氨基酰tRNA合成酶C.ATP合成酶D.转位酶E.GTP27.参与新生多肽链正确折叠的蛋白质是(A)A.分子伴侣B.G蛋白C.转录因子D.释放因子E.p因子28.原核生物起始tRNA是 (C)A.甲硫氨酰-tRNAB.缬氨酰-tRNAC.甲酰化的甲硫氨酰-tRNAD.氨酰-tRNAE.乙酰化的甲硫氨酰-tRNA29.新生肽链合成的方向是 (A )A.从N端到C端合成B.从C端到N端合成C.没有固定的方向D.先从N端到C端合成小片段,再连接成一条多肽链E.从中间部位向N端和C端两个方向同时进行30.蛋白质磷酸化修饰的潜在修饰位点是(B)A.甘氨酸B.酪氨酸C.苯丙氨酸D.谷氨酸E.赖氨酸34.蛋白质合成终止是 (B)A.核糖体到达mRNA分子的3'末端B.释放因子识别终止密码子进入A位C.释放因子进入P位D.mRNA出现发夹结构,核糖体无法移动E.特异的tRNA进入A位35.氯霉素对细菌蛋白质合成的生物学影响是(E )A.导致未成熟多肽链的释放B.抑制核糖体大小亚基的结合C.抑制tRNA功能D.抑制30S核蛋白体亚基的活化E.抑制50S核蛋白体亚基的肽酰转移酶活性38.若向mRNA的编码区插入一个核苷酸,则会出现(D)A.翻译出的蛋白质的第1个氨基酸残基改变B.翻译出的蛋白质的氨基酸残基序列不变C.插入处上游翻译出来的氨基酸序列完全改变D.插入处下游翻译出来的氨基酸序列完全改变E.仅在插入处一个氨基酸残基改变。
第15章 蛋白质的生物合成
tRNA反密码子 第1位碱基
I
U G AC
mRNA密码子 第3位碱基
U, C, A A, G U, C U G
2020/2/21
21
Robert W. Holley 1922~1993
Har Gobind Khorana
1922~
Marshall W. Nirenberg
1927~2010
The Nobel Prize in Physiology or Medicine 1968
• 原核细胞中数个结构基因常串联为一个转录 单位,转录生成的mRNA可编码几种功能相关
的蛋白质,为多顺反子(polycistron) 。
2020/2/21
5
原核生物的多顺反子
5 PPP
3
真核生物的单顺反子
5 mG - PPP
蛋白质
3
蛋白质
非编码序列
核蛋白体结合位点
编码序列
起始密码子
终止密码子
"for their interpretation of the genetic code and
2020/2/21
its function in protein synthesis"
22
二、核糖体是多肽链合成的装置
2020/2/21
23
2020/2/21
24
2020/2/21
27
原核、真核生物核蛋白体的组成
4. 通用性(universal)
▪ 蛋白质生物合成的整套密码,从原核生 物到人类都通用。
▪ 已发现少数例外,如动物细胞的线粒体、 植物细胞的叶绿体。
▪ 密码的通用性进一步证明各种生物进化 自同一祖先。
蛋白质的生物合成习题与参考参考答案

精心整理第十五章蛋白质生物合成一、填空题:1.三联体密码子共有64个,其中终止密码子共有3个,分别为UAA、UAG、UGA。
2.密码子的基本特点有四个分别为从5′→3′无间断性、简并性、变偶性、通用性。
3.次黄嘌呤具有广泛的配对能力,它可与U、C、A三个碱基配对,因此当它出现在反密码子中时,会使反密码子具有最大限度的阅读能力。
4.原核生物核糖体为70S,其中大亚基为50S,小亚基为30S;而真核生物核糖体为80S,大亚基为60S5tRNA 可表示为67列称为8.氨酰,NH29.氨酰能量。
101112T C序列。
这是1A、B、mRNA遗传密码的阅读方向是3′→5′C、mRNA遗传密码的阅读方向是5′→3′D、mRNA密码子与tRNA反密码子通过A-T,G-C配对结合E、每分子mRNA有3个终止密码子2.下列反密码子中能与密码子UAC配对的是(D)A、AUGB、AUIC、ACUD、GUA3.下列密码子中,终止密码子是(B)A、UUAB、UGAC、UGUD、UAU4.下列密码子中,属于起始密码子的是(A)A 、AUGB 、AUUC 、AUCD 、GAG5.下列有关密码子的叙述,错误的一项是(C )A 、密码子阅读是有特定起始位点的B 、密码子阅读无间断性C 、密码子都具有简并性D 、密码子对生物界具有通用性6.密码子变偶性叙述中,不恰当的一项是(A )A 、密码子中的第三位碱基专一性较小,所以密码子的专一性完全由前两位决定B 、第三位碱基如果发生了突变如AG 、CU ,由于密码子的简并性与变偶性特点,使之仍能翻译出正确的氨基酸来,从而使蛋白质的生物学功能不变C 、次黄嘌呤经常出现在反密码子的第三位,使之具有更广泛的阅读能力,(I-U 、I-C 、I-A )从而可减少由于点突变引起的误差D 、几乎有密码子可用U C XY 或U C XY7.关于核糖体叙述不恰当的一项是(B )A B C 、在核糖体的大亚基上存在着肽酰基(P D 结合的位点8.tRNA 的叙述中,哪一项不恰当(D )A 、tRNAB 、起始C D 9.tRNA D )A 、tRNAB CCA 的结构末端C 、T CD 、D10C )A 、相应C 、相应上的反密码子11C )A 、氨酰-tRNA 合成酶促反应中由ATP 提供能量,推动合成正向进行B 、每种氨基酸活化均需要专一的氨基酰-tRNA 合成酶催化C 、氨酰-tRNA 合成酶活性中心对氨基酸及tRNA 都具有绝对专一性OD 、该类酶促反应终产物中氨基酸的活化形式为R -CH -C -O -ACC -tRNANH 212.原核生物中肽链合的起始过程叙述中,不恰当的一项是(D )A 、mRNA 起始密码多数为AUG ,少数情况也为GUGB 、起始密码子往往在5′-端第25个核苷酸以后,而不是从mRNA5′-端的第一个苷酸开始的C 、在距起始密码子上游约10个核苷酸的地方往往有一段富含嘌呤的序列,它能与16SrRNA3′-端碱基形成互补D 、70S 起始复合物的形成过程,是50S 大亚基及30S 小亚基与mRNA 自动组装的13.有关大肠杆菌肽链延伸叙述中,不恰当的一项是(C )A 、进位是氨酰-tRNA 进入大亚基空差的A 位点B 、进位过程需要延伸因子EFTu 及EFTs 协助完成C 、甲酰甲硫氨酰-tRNA f 进入70S 核糖体A 位同样需要EFTu -EFTs 延伸因子作用D 、进位过程中消耗能量由GTP 水解释放自由能提供14.移位的叙述中哪一项不恰当(C )A 、移位是指核糖体沿mRNA (5′→3′)作相对移动,每次移动的距离为一个密码子B 、移位反应需要一种蛋白质因子(EFGC 、EFG 是核糖体组成因子D 、移位过程需要消耗的能量形式是GTP 15.蛋白质生物合成的方向是:(B )A 、从C 端到N 端B 、从N 端到C 端C 、定点双向进行D 、从C 端、N 端同时进行16A 、氨基酸随机地连接到tRNA 上去B 、?新生多肽链的合成都是从C -端向N -C 、?D 、?17.70S D )A 、mRNA 1B 、mRNA 2C 、mRNA 3D 、mRNA 1、IF 2和IF 318.mRNA 与f 结合过程中起始因子为(A )A 、IF 1IF 2及IF 319D )A B C 、mRNA 与核蛋白体30S 亚基结合D 、氨酰tRNA 合成酶催化氨基酸活化20.假设翻译时可从任一核苷酸起始读码,人工合成的(AAC )n (n 为任意整数)多聚核苷酸,能够翻译出几种多聚氨基酸?(C )A 、一种B 、二种C 、三种D 、四种21.绝大多数真核生物mRNA5’端有(A )A 、帽子结构B 、PolyAC 、起始密码D 、终止密码22.能与密码子ACU 相识别的反密码子是(D )A 、UGAB 、IGAC 、AGID 、AGU23.原核细胞中新生肽链的N-末端氨基酸是(C )A、甲硫氨酸B、蛋氨酸C、甲酰甲硫氨酸D、任何氨基酸24.tRNA的作用是(D)A、?把一个氨基酸连到另一个氨基酸上B、将mRNA连到rRNA上C、增加氨基酸的有效浓度D、把氨基酸带到mRNA的特定位置上25.细胞内编码20种氨基酸的密码子总数为:(D)A、16B、64C、20D、6126.下列关于遗传密码的描述哪一项是错误的?(C)A、密码阅读有方向性,5'-端开始,3'-端终止B、密码第3位(即3′-端)碱基与反密码子的第1位(即5′-端)碱基配对具有一定自由度,有时会出现多对一的情况C、一种氨基酸只能有一种密码子D、一种密码子只代表一种氨基酸27.蛋白质合成所需的能量来自(C)A、ATPB、GTPC、ATP和GTPD、CTP28AB、密码阅读有方向性,5′-端起始,3′-C、一种氨基酸可有一组以上的密码D、一组密码只代表一种氨基酸29.mRNA的A、5′30A、UAAB123RNA—→DNA。
中国海洋大学资料生物化学课件第十五章 蛋白质合成(新)

(一)遗传密码的破译
* 美国科学家 Nirenberg 等, 1968 年获诺贝尔生 理学奖。 * 1961 年, Nirenberg 等人,在大肠杆菌的无细 胞体系中外加 poly(U) 模板、 20 种标记的氨基 酸,经保温后得到了多聚 phe-phe-phe ,于是 推测UUU编码phe。利用同样的方法得到 CCC 编码pro,GGG编码gly,AAA编码lys。 * 如果利用 poly ( UC ),则得到多聚 Ser-LeuSer-Leu,推测UCU编码Ser,CUC编码Leu。 * 到1965年就全部破译了64组密码子。
蛋白质合成时,mRNA结合于核糖体小亚基 上,大亚基结合带氨基酸的tRNA,tRNA的 反密码子与mRNA密码子配对,ATP供能, 合成蛋白质。
三、遗传密码
遗传密码:mRNA中的核苷酸序列与蛋白质 中氨基酸序列之间的对应关系称为遗传密码 三个碱基编码一个氨基酸称为三联体密码或 密码子(Coden)。 密码子的发现。
(1)在进入A位点之前,新氨酰tRNA首先必须与延伸因子EF— TU—GTP结合。 (2)延伸因子 EF—Tu是一个GTP结合蛋白,参与氨酰 tRNA的 就位。 (3)氨酰tRNA入位后,EF—TU—GTP水解,EF—TU—GDP从 核糖体上释放下来,在第二个延伸因子 EF—Ts 帮助下 EF— Tu—GDP 释 放 掉 GDP 并 重 新 结 合 一 分 子 GTP 再 生 成 EF— Tu—GTP。
简并性的生物学意义
A、减少有害突变:假如每种氨基酸只有一个密 码子,那么剩下的44个密码子都成了终止子, 如果一旦哪个氨基酸的密码子发生了单碱基的 点突变,那么极有可能造成肽链合成的过早终 止。如GUN 编码 Ala ,由于简并性的存在,不 论第三位的U变成什么,都仍然编码Ala; B、可以使DNA上的碱基组成有较大的变化余地, 而仍然保持多肽上氨基酸序列不变(意思基本 同上)。
华中农业大学生物化学本科试题库_第15章__RNA生物合成

华中农业⼤学⽣物化学本科试题库_第15章__RNA⽣物合成第15章 RNA⽣物合成五单元⾃测题(⼀)名词解释或概念⽐较转录与逆转录单顺反⼦与多顺反⼦反意义链与有意义链启动⼦与终⽌⼦内含⼦与外显⼦RNA聚合酶全酶与核⼼酶操纵⼦与操纵基因顺式作⽤元件与反式作⽤因⼦。
阻遏物与辅阻遏物-10序列与TATA box11. 核酶(⼆)填空题1. 引物酶与转录中的RNA聚合酶之间的差别在于它对不敏感,并可以作为底物。
2. ⼤肠杆菌中DNA指导的RNA聚合酶全酶的亚基组成为,去掉因⼦的部分称为核⼼酶,这个因⼦使全酶能辩认DNA上的位点。
3. 利福平抑制细菌中转录的起始,因为。
4. 原核⽣物中各种RNA是催化⽣成的。
⽽真核⽣物基因的转录分别由种RNA聚合酶催化,其中rRNA基因由转录,hnRNA基因由转录,各类⼩分⼦量RNA则是的产物。
5. ⼀个转录单位⼀般应包括序列、序列和顺序。
6. 真核细胞中编码蛋⽩质的基因多为。
编码的序列还被保留在成熟mRNA中的是,编码的序列在前体分⼦转录后加⼯中被切除的是。
在基因中______被_____分隔,⽽成熟的mRNA中外显⼦转录的序列被拼接起来。
7. 真核⽣物与原核⽣物的tRNA前体⼀个重要的区别就是前者含有。
8. 在原核细胞中,由同⼀调控区控制的⼀群功能相关的结构基因组成⼀个基因表达调控单位,称为,其调控区包括和。
9. ⼤肠杆菌乳糖操纵⼦调节基因编码的与结合,对lac表达实施负调控;和的复合物结合于上游部分,对lac表达实施正调控。
10. ⼤肠杆菌⾊氨酸操纵⼦阻遏蛋⽩必须先与辅阻遏物相结合,才能结合于操纵基因。
在trp操纵基因与结构基因之间有⼀段能被转录的,可编码含有2个残基的14肽。
⾊氨酸充裕时,翻译迅速,⾊氨酸不⾜时,翻译迟滞,结构基因的转录得以继续进⾏,称为调节。
11. 乳糖操纵⼦的启动,不仅需要有诱导物乳糖存在,⽽且培养基中不能有,因为它的分解代谢产物会降低细胞中的⽔平,⽽使复合物不⾜,它是启动基因启动所不可缺少的调节因⼦。
生物化学 第十五章 DNA的生物合成

(二)与DNA复制有关的酶和蛋白质(原核生物)
5´ 3´
解旋酶 解链酶 引物酶和引 发体
SSB
RNA引物
DNA聚合 酶III DNA聚合 酶I
3´
5´
RNA引 物
3´
5´
1.DNA聚合酶 DNA聚合酶Ⅰ polⅠ
(1) 具5′
3′聚合酶活性
将脱氧核糖核苷三磷酸 dNTP 逐个添加到具有 3 ′ — OH 末端的 多核苷酸链上形成3, 5—磷酸二酯键。
4. 引发酶(引物合成酶)
引物的合成由引发酶催化完成,这些酶 在模板单链DNA上识别特殊序列,合成RNA引 物。它本身没有活性,需要与“引发前体” 结合在一起,形成“引发体”后才有活性。
复制早期易发生碱基参入错误,用 RNA较好,然后通过polⅠ的5′ 3′端 核酸外切酶的活性切去,代之以dNMP,可 消除最初阶段的错误。
• 2、方向 • DNA复制可以朝一个方向(单向复 制,unidirectional),也可以朝两个相反 方向进行(双向复制,bidirectional,主 要)。
•
DNA复制一般是对称的,两条链同时 进行,也有不对称的,一条链复制完后 再进行另一条链的复制。 • 在迅速生长的原核生物中,第一个染 色体DNA分子的复制还未完成,第二个 DNA分子就在同一个起始点上开始复制。
+
50 复制
转化率
功能
0 .05
修复
1999年发现聚合酶 和,它们涉及DNA的错误倾向修 复(errooune repair)
polⅠ不是DNA复制酶,理由:
A、该酶合成 DNA 速度太慢,只是细胞 内DNA复制速度的1%; B、持续合成能力较低,而细胞内 DNA 复制不会频繁中止; C、许多基因突变都会影响DNA复制, 但都与polⅠ无关。
蛋白质的生物合成习题和参考答案与解析

完美格式整理版第十五章蛋白质生物合成一、填空题:1.三联体密码子共有64 个,其中终止密码子共有 3 个,分别为UAA 、UAG 、UGA 。
2.密码子的基本特点有四个分别为从5′→3′无间断性、简并性、变偶性、通用性。
3.次黄嘌呤具有广泛的配对能力,它可与U 、 C 、 A 三个碱基配对,因此当它出现在反密码子中时,会使反密码子具有最大限度的阅读能力。
4.原核生物核糖体为70 S,其中大亚基为50 S,小亚基为30 S;而真核生物核糖体为80 S,大亚基为60 S,小亚基为40 S。
5.原核起始tRNA,可表示为tRNA f甲硫,而起始氨酰tRNA表示为f Met-tRNA f甲硫;真核生物起始tRNA可表示为tRNA I甲硫,而起始氨酰-tRNA表示为Met-tRNA f甲硫。
6.肽链延伸过程需要进位、转肽、移位三步循环往复,每循环一次肽链延长 1 个氨基酸残基,原核生物中循环的第一步需要EF-Tu 和EF-Ts 延伸因子;第三步需要EF-G 延伸因子。
7.原核生物mRNA分子中在距起始密码子上游约10个核苷酸的地方往往有一段富含嘌呤碱基的序列称为Shine-Dalgrano序列,它可与16S-rRNA 3′-端核苷酸序列互补。
8.氨酰-tRNA的结构通式可表示为:OtRNA-O-C-CH-RNH2,与氨基酸键联的核苷酸是A(腺嘌呤核苷酸)。
9.氨酰-tRNA合成酶对氨基酸和相应tRNA都具有较高专一性,此酶促反应过程中由ATP 水解提供能量。
10.肽链合成的终止阶段,RF1因子和RF2因子能识别终止密码子,以终止肽链延伸,而RF3因子虽不能识别任何终止密码子,但能协助肽链释放。
11.蛋白质合成后加工常见的方式有磷酸化、糖基化、脱甲基化、信号肽切除。
12.真核生物细胞合成多肽的起始氨基酸为甲硫氨酸,起始tRNA为tRNA I甲硫,此tRNA 分子中不含T C 序列。
这是tRNA家庭中十分特殊的。
蛋白质的生物合成

16
(二)氨基酰-tRNA合成酶 (aminoacyl-tRNA synthetase)
氨基酰-tRNA合成酶
氨基酸 + tRNA
氨基酰- tRNA
ATP
AMP+PPi
17
1. 氨基酰-tRNA合成酶对底物氨基酸和tRNA 都有高度特异性:每种氨基酸都只有1种氨基酰tRNA合成酶,生成特定的氨基酰-tRNA(aa-tRNA)
参与蛋白质生物合成的主要物质 蛋白质生物合成过程 翻译后的加工与靶向输送 蛋白质生物合成的抑制剂
3
第一节 参与蛋白质生物合成的 主要物质
蛋白质合成体系:20种氨基酸,mRNA、 tRNA、核蛋白体、酶和蛋白因子、无机离 子、ATP 、GTP 等。
4
一、mRNA:合成蛋白质பைடு நூலகம்模板
(一) mRNA是遗传信息的携带者 遗传学将编码一个多肽的遗传单位称为
14
密码子与反密码子除通过碱基互补结合外, 还具有摆动性,即密码子的第3位碱基与反 密码子的第1位碱基配对不严格,称为摆动 配对。
15
密码子、反密码子配对的摆动现象
tRNA反密码子 第1位碱基
I
U G AC
mRNA密码子 第3位碱基
U, C, A A, G U, C U G
密码子的第3位碱基发生突变时,并不影响tRNA带入 正确的氨基酸。
2.氨基酰-tRNA的表示方法:
Ser-tRNASer
Met-tRNAMet
真核生物起始因子辨认的Met-tRNAiMet 延长因子辨认Met-tRNAeMet
右上角字母可略
18
三、核糖体:合成蛋白质的场所
蛋白质的生物合成

第十五章蛋白质的生物合成蛋白质的生物合成:将mRNA分子中由碱基序列组成的遗传信息,通过遗传密码破译的方式转变成为蛋白质中的氨基酸排列顺序,因而称为翻译(translation)。
1、遗传密码1. 遗传密码和密码单位1) 密码子:mRNA中的核苷酸序列与多肽链中氨基酸序列之间的对应关系。
mRNA上每三个连续核苷酸对应一个氨基酸,这三个连续的核苷酸就称为一个密码子,或三联体密码。
2) 遗传密码:密码子的总和。
3) 64个密码子:其中61个代表20种氨基酸,3个代表终止密码子。
4) 遗传密码的破译5) 遗传密码字表起始密码:AUG(编码甲硫氨酸、甲酰甲硫氨酸),少数情况 GUG;终止密码:无义密码子,不编码氨基酸的密码子,它们单个或串联在一起用于多肽链翻译的结束,没有相应的tRNA存在,有UAA、UAG、UGA。
同义密码:编码相同氨基酸的不同密码子。
6) 遗传密码的基本特征方向性:5’到3’,AUG。
遗传密码的连续性:密码子之间没有任何起“标点”作用的空格,阅读是连续的,一次阅读3个核苷酸(碱基)。
遗传密码的不重叠性:在绝大多数生物中,阅读mRNA时是以密码子为单位,不重叠地阅读;但少数噬菌体的的遗传密码是重叠的。
兼并性:遗传密码中,除色氨酸和甲硫氨酸仅有一个密码子外,其余氨基酸有2~4个或多至6个密码子,这种由多个密码子编码同一种氨基酸的现象。
密码的简并性往往表现在密码子的第三位碱基上。
密码的偏爱性:在不同生物中使用同义密码子的频率是不相同的。
意义:减少有害突变,维持物种稳定。
密码的摆动性tRNA上的反密码子与mRNA上的密码子配对时,密码子的第一位、第二位碱基配对是严格的,第三位碱基可以有一定变动。
摆动性:tRNA上反密码子的第1位碱基与mRNA密码子的第3位碱基配对时,并不严格遵循碱基配对规律,可以在一定范围内变动的现象,又称变偶性。
密码子的表示法:码子的专一性基本取决于前两位碱基,第三位碱基起的作用有限。
华中农业大学生物化学本科试题库_第15章__RNA生物合成

第15章 RNA生物合成五单元自测题(一)名词解释或概念比较转录与逆转录单顺反子与多顺反子反意义链与有意义链启动子与终止子内含子与外显子RNA聚合酶全酶与核心酶操纵子与操纵基因顺式作用元件与反式作用因子。
阻遏物与辅阻遏物-10序列与TATA box11. 核酶(二)填空题1. 引物酶与转录中的RNA聚合酶之间的差别在于它对不敏感,并可以作为底物。
2. 大肠杆菌中DNA指导的RNA聚合酶全酶的亚基组成为,去掉因子的部分称为核心酶,这个因子使全酶能辩认DNA上的位点。
3. 利福平抑制细菌中转录的起始,因为。
4. 原核生物中各种RNA是催化生成的。
而真核生物基因的转录分别由种RNA聚合酶催化,其中rRNA基因由转录,hnRNA基因由转录,各类小分子量RNA则是的产物。
5. 一个转录单位一般应包括序列、序列和顺序。
6. 真核细胞中编码蛋白质的基因多为。
编码的序列还被保留在成熟mRNA中的是,编码的序列在前体分子转录后加工中被切除的是。
在基因中______被_____分隔,而成熟的mRNA中外显子转录的序列被拼接起来。
7. 真核生物与原核生物的tRNA前体一个重要的区别就是前者含有。
8. 在原核细胞中,由同一调控区控制的一群功能相关的结构基因组成一个基因表达调控单位,称为,其调控区包括和。
9. 大肠杆菌乳糖操纵子调节基因编码的与结合,对lac表达实施负调控;和的复合物结合于上游部分,对lac表达实施正调控。
10. 大肠杆菌色氨酸操纵子阻遏蛋白必须先与辅阻遏物相结合,才能结合于操纵基因。
在trp操纵基因与结构基因之间有一段能被转录的,可编码含有2个残基的14肽。
色氨酸充裕时,翻译迅速,色氨酸不足时,翻译迟滞,结构基因的转录得以继续进行,称为调节。
11. 乳糖操纵子的启动,不仅需要有诱导物乳糖存在,而且培养基中不能有,因为它的分解代谢产物会降低细胞中的水平,而使复合物不足,它是启动基因启动所不可缺少的调节因子。
第15章 蛋白质的生物合成
进 位
成肽 转 位
成肽
成肽是由转肽酶催化形成肽键的过程。
Tu GTP
Tu GDP
5'
AUG
3'
fMet fMet
Tu GTP
5'
AUG
3'
(三) 原核生物翻译终止
当mRNA上的终止密码子出现后,多肽链合成 停止,肽链从肽酰-tRNA中脱落,mRNA、核糖体 等解体,此时翻译过程终止。 终止密码子不被任何氨基酰-tRNA识别,只有 释放因子(RF)能够识别它并进入核糖体的A位。 1. 释放因子可引起转肽酶活性的改变,从而使 肽链从tRNA上脱落。 2. 释放因子可引起核糖体结构的改变,从而使 复合体趋于解体。
解码
反密码子(anti-codon) 反密码子是位于tRNA、可与mRNA的三联体密 码子配对的三个相邻核苷酸。在蛋白质的合成中, 起解读密码、辅助将特异的氨基酸引入合成位点。 tRNA
5'
AUG
反密码子
3'
mRNA
密码子
遗传密码的特点
翻译时遗传密码的阅读方向是 5’→3’,即读 码从mRNA的起始密码子AUG开始,按5’→3’ 的方向逐一阅读,直至终止密码子。
COORF
5'
UAG
3'
第四节
蛋白质翻译后加工和靶向输送
Posttranslational Modification and Targeting Transfer of Protein
蛋白质翻译后加工
第15章 RNA的生物合成

(NMP)n+1 + PPi
15.2.2 启动子和转录因子
启动子(promoter):RNA聚合酶结合模板DNA的部位。
一般位于转录起始位点(+1)的上游;
本身序列不被转录。
RNA聚合酶 与启动子的 结合模式图
原核生物启动子 RNA聚合酶与启动子结合的识别亚基 • σ 因子负责特异性识别启动子;
-10 区(Pribnow框)
TATAAT ATATTA
RNA-pol的识别位点
RNA-pol的结合位点
• -35区和-10区
不同基因的启动子中,二者序列和位置略有区别;
trp tRNA Trp lac rec A ara
最大一致性
35 - 区 TTGACA TT TACA TT TACA TTGATA CTGACG TTGACA
只以一条DNA链或其一段DNA为模板; 碱基互补配对的原则:A与U,G与C配对; 合成方向:5’→3’,并连续进行;
无校读功能;
需要Mg2+或Mn2+离子; 可与多种调节转录的蛋白因子相互作用。
种类: 原核生物的RNA聚合酶
真核生物的RNA聚合酶
原核生物的RNA聚合酶
核心酶(core enzyme)
• 不同σ 因子可以识别不同的启动子; • σ 因子与启动子亲和力的不同,对转录起始频率
(基因表达的程度)有重要的调控作用。
启动子的保守区域
RNA聚合酶结合区 结构基因
5 3 3 5 开始转录 5 3 -50 -40 -30 -20 -10 1 10 3 5
-35 区
TTGACA AAC T G T
转录 DNA的一条链
NTP(N=A、G、C、U)
第十五章 RNA的生物合成

5. 原核细胞基因转录的产物大多数为多顺反子mRNA,这 是由于原核转录系统中功能相关的基因共享一个启动子, 它们在转录时,以一个共同的转录单位进行转录。而真核 细胞,每一种蛋白质的基因都有自己独立的启动子,所以 真核细胞转录产物是单顺反子mRNA。
原核
DNA
A
B
C
P
转录
mRNA
A
B
C
真核
DNA P A P B P C
依赖ρ因子(rho factor)的终止:
(三)真核生物与原核生物转录的主要区别
1. 真核细胞RNApol种类较多,根据它们对α-鹅膏蕈碱的
敏感性不同分为RNA pol I 、II 、III(or A、B、C),
它们是高度分工的,不同的RNA聚合酶负责合成不同 的RNA。
2. 真核启动子比原核启动子更复杂和更多样性,不同的 RNA聚合酶有不同的启动子。
组成上类似于DNA(类似于DNA的RNA,D-RNA),代谢很 快,迅速合成和降解。 HnRNA分子很大,其中大约只有10%转变成mRNA,其 余在转录后的加工过程中被降解掉。
HnRNA转变成mRNA的加工过程包括
(1)戴帽 (2)加尾 (3)剪接(splicing) (4)修饰:主要是链内腺苷的甲基化
Qβ的基因次序:
5ˊ末端 成熟蛋白一外壳蛋白(或A1蛋白)一复制酶β亚基 3ˊ-末端
QβRNA复制酶由4种亚基组成:α、β、γ、δ亚 基,除了β亚基来自Qβ噬菌体外,其余的(α、γ和
δ)亚基都来自宿主即E.Coli。
当噬菌体Qβ侵入E.Coli后,先以Qβ的RNA为横
板合成β-亚基,然后再和宿主细胞中的α、γ、δ亚基 结合成复制酶,有了复制酶就可进行RNA的复制。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
一、半保留复制的实验证据
1958年M.Meselson和F.Stahl Radioisotope labelin(放 射性同位素标记)and density gradient(密度梯度 离心)centrifugation clearlydistinguishes replications of semiconservative from conservative.
α β γ ε δ
分布 聚合酶 活力 3`-5`外 切酶
功能
细胞核
+ 无 引物合 成
细胞核
+ 无 修复
线粒体
+ 有 线粒体 复制
细胞核
+ 有 修复
细胞核
+ 有 核DNA 合成
2. DNA 连接酶(DNA ligase)
连接DNA链3’-OH末端和相邻DNA链5’-P末端,使二者生成 3’ ,5’磷酸二酯键,从而把两段相邻的DNA链连成完整的链
15.1 DNA的复制
15.1.1 DNA的复制方式—半保留复制
1953年,Watson和Crick在DNA双螺旋 结构的基础上提出了半保留复制假说:
DNA在复制过程中,首先碱基之间的氢 键破裂,使两条链解旋并分开,然后以 碱基互补的方式,以每条单链为 模板, 按单链DNA的核苷酸顺序合成子链。在 此过程中,每个子代分子的一条链来自 亲代DNA,另一条链是新合成的,这种 复制方式称为半保留复制。
DNA聚合酶能催化四种脱氧核糖核苷三磷酸 合成DNA
四种脱氧核苷 三磷酸为底物
具有3`-OH末端的低聚 多核苷酸为引物
Mg
2+或Mn 2+
单链DNA为模板
① 原核细胞的DNA聚合酶
DNA聚合酶I(pol I)—Kornberg酶
pol I是一个多功能酶
DNA聚合酶活力:使DNA链 沿5`-3`方向延伸;
3.与DNA解旋和解链有关的酶和蛋白质:
• 解螺旋酶(helicase) • DNA拓扑异构酶(DNA topoisomerase) • 单链DNA结合蛋白(single stranded DNA
binding protein,SSB)
(1)解螺旋酶
解螺旋酶通过水解ATP供能,
使DNA两链间碱基对的氢键断 裂,从而解开双链DNA。 解螺旋酶可沿模板随复制叉 的伸展而移动
Байду номын сангаас
式称为半不连续复制。
随从链上不连续复制的DNA片段称为岗崎片段。每一
个岗崎片段5’-端都带有一个RNA引物。
随从链复制时必须等
待模板链解开足够长度时, 才能从5′→3′合成引物 后开始复制。延伸时,又 要等待下一段暴露出足够
长的模板,才能再次合成
岗崎片段
引物而延长。
15.1.2 参与DNA复制的酶类和蛋白质
第十五章 DNA、RNA和 蛋白质的生物合成
复 制 DNA
1958年Crick将生物
转 录 RNA
遗传信息的这种传 递方式称为中心法则。
反 录 转 蛋 质 白 翻 译
复 制
复制—以原来的DNA分子为模板,合成出相同 分子的过程。 转录—在DNA分子上合成出与其核苷酸顺序相 对应的RNA的过程。 翻译—在RNA的控制下,根据核酸链上每三个核 苷酸决定一个氨基酸的三联体密码规则,合成出 具有特定氨基酸顺序的蛋白质肽链的过程。
(2)单链结合蛋白(SSB)
SSB的作用是与分开
的两条DNA单链结合,
在复制中维持模板处
于单链状态并保护单
链的完整。
(3 )拓扑异构酶(I、II)
拓扑异构酶I:
切断DNA双链中的一条链,使DNA解旋中不 致打结,适当时再把切口封闭,反应不耗能。 拓扑异构酶II:
无ATP时,可切断超螺旋的两条链,使超螺旋 松弛,然后再把切口封上。 在利用ATP供能情况下,能将复制叉前方产生 的正超螺旋变成负超螺旋。
二、半保留复制的意义
DNA的半保留复制保证了DNA在代谢上的稳定性, 经过许多代的复制,DNA多核苷酸链仍可保持完整, 这种稳定性体现了DNA遗传过程的相对保守性。 遗传的保守性是相对的,而不是绝对的。自
然界还存在着普遍的变异现象。
三、复制的半不连续性
DNA复制时顺着解链方向而生成的子链,复制是连续 进行的,这股链称为前导链。另一条子链复制的方向与解 链方向相反,复制是不连续的,称为随从链。这种复制方
底物: dNTP( dATP、dGTP、dCTP、dTTP)
酶:DNA聚合酶 模板:解开成单链的DNA母链 引物:RNA,提供 3’-OH末端 其它酶和蛋白质因子:解螺旋酶、单链结合蛋白、
复制是酶催化下的核苷酸聚合过程,需要多种物质的共同参与:
拓扑异构酶、引物酶、DNA连接酶等
1.
DNA聚合酶(DNA-polymerase)
ε ε
DNA聚合酶Ⅲ(pol Ⅲ )
α亚基有DNA聚合酶活力; ε亚基有3`-5`核酸外切酶活力, 起校对作用; θ亚基起组建复合物的作用; DNA的错误倾向修复:修复 缺乏准确性,出现高突变率。
DNA聚合酶Ⅳ、Ⅴ
DNA聚合酶Ⅲ的β亚基二聚体 与DNA结合的空间关系
大肠杆菌中DNA聚合酶的性质比较
在复制过程中,DNA每复制10bp,复制叉前方的模 板DNA双螺旋就要绕其长轴旋转一周,产生正超螺旋。
三种酶催化生成磷酸二酯键的比较
提供核糖3’-OH
DNA聚合酶 在引物作用下 延长新链 连接酶 连接不连续的 两条单链 拓扑酶 切断两链再连接
结果
(dNMP)n+1 不连续→连续链
连接双链DNA的单链缺口
Arthur Kornberg
3`-5`核酸外切酶活力:校 对功能;
5`-3`核酸外切酶活力:切 除引物、嘧啶二聚体,修复;
焦磷酸解作用
焦磷酸基交换作用
3`-5` 核酸 外切酶 活力
5`-3` 核酸 外切酶 活力
DNA聚合酶Ⅱ(pol Ⅱ)
pol Ⅲ
φ
DNA聚合酶活力:需带有缺 口的DNA的双链作引物,主 要参与DNA的修复。 3`-5`核酸外切酶活力
pol I
功 DNA聚合酶 能 5`-3`外切酶 3`-5`外切酶 分子量 酶分子数/细胞 生物学活性 主要功能 + + + 109000 400 1 切除引物、修复
pol Ⅱ
+ - + 120000 100 0.05 修复
pol III
+ - + 900000 10 15 复制
② 真核细胞的DNA聚合酶