木质素的微生物降解_secret
微生物分解木质素的机制和应用

微生物分解木质素的机制和应用当人们想到微生物,往往会联想到细菌和病毒。
然而,微生物还有另一个十分重要的作用,那就是分解木质素。
木质素是一种非常复杂的有机化合物,它是构成木材主要部分的聚合物。
木材中的木质素影响了木材的颜色、形状和硬度。
由于木质素的结构复杂,其降解也非常困难。
微生物的出现和进化,使得这一难题得到了一定程度的解决。
一、微生物分解木质素的机制1. 溶菌酶的作用溶菌酶是一种水解木质素的酶类,与细菌和真菌都有密切关系。
在存在溶菌酶的微生物中,木质素产生的底物可以通过微生物的代谢途径,转化为有机酸和气体等形式释放出来。
因此,溶菌酶的存在可以促进木质素的生物降解。
2. 氧化酶的作用氧化酶是一类氧化还原酶,可以用于将木质素中的芳香环酚类以及羟丙基、羟甲基等短链糖类转化为各类醛酮。
这些产物是微生物能够利用的底物,从而促进木质素的分解。
3. 木素脱甲基酶的作用木素脱甲基酶是一类针对木素分子中的甲基进行去除的酶。
这类酶主要存在于真菌和细菌中。
经过这种酶的处理,木质素中的芳香环甲基被去掉,从而使木质素更容易分解。
二、微生物分解木质素的应用1. 软木板软木板是以树皮为原料,经过加工处理得到的一种材料。
在制备过程中,木质素被微生物分解,从而使软木更加柔软、耐用。
2. 生物燃料生物燃料是以生物质为原料生产的一种燃料,它可以是来自植物、动物或者微生物的有机废弃物,如纤维素、木质素等。
通过微生物分解木质素,可以产生甲烷、CO2等气体,这些气体可以用于发电和供暖,从而成为一种清洁、可再生的能源。
3. 污染物降解一些化工废弃物和污染物,如苯、甲苯等芳香环化合物,由于分子结构复杂,难以通过传统的化学方法进行降解。
微生物通过分解木质素的作用,可以分解这些污染物,从而提供一种清洁的化学降解方法。
总的来说,微生物分解木质素机制的研究,不仅可以增加对微生物本身生态环境的理解,还可以为人们提供多种有益的工业应用,使得木质素等有机废弃物得到更加有效的利用。
木质素生物降解研究进展

2000年 12月河南农业大学学报Dec. 2000第34卷 第4期Journal of Henan Agricultural University V ol.34N o.4文章编号:1000-2340(2000)04-0349-06木质素生物降解研究进展吴坤,张世敏,朱显峰(河南农业大学生物工程学院,河南郑州450002)摘要:评述了木质生物降解的最新研究进展,主要包括木质素的生物降解机制、降解木质素的微生物种类及其产生的相关酶类、微生物的代谢调控和分子生物学.此外,对木质素降解生物的实际应用和应用前景也进行了评论.关键词:木质素;生物降解;秸杆;白腐真菌;环境保护中图分类号:Q591.4 文献标识码:AR ecent research advances on the lignin biodegradationW U K un1,ZH ANG Shi2min2,ZH U X ian2feng2(C ollege of Biological Engineering,Henan Agricultural University,Zhengzhou450002,China)Abstract:A review is given of the m ost recent research advances on the lignin biodegrading,including the mechanism of the lignin2biodegrading,ligninolytic organisms and the Enzymes they secrete,regulation and control of the microbe metabolism and m olecular biology.Finally the practical utilization and the prospects of lignin2biodegrading are dis2 cussed.K ey w ords:lignin;biodegradation;straw;white2rot fungi;environmental protection 绿色植物占地球陆地生物量的95%,其化学物质组成主要是木质素、纤维素和半纤维素,它们占植物干重的比率分别为15%~20%,45%和20%[1].农作物秸杆是这类生物质资源的重要组成部分,全世界年产量为20多亿吨,而我国为5亿多吨.但是,要充分、有效地利用这类资源却相当困难,这是由于秸秆产量随季节变化,且量大、低值、体积大、不便运输,大多数动物都不能消化其木质纤维素,自然降解过程又极其缓慢,导致大部分秸秆以堆积、荒烧等形式直接倾入环境,造成极大的环境污染和浪费.存在于秸秆中的非水溶性木质纤维素很难被酸和酶水解,主要是因纤维素的结晶度、聚合度以及环绕着纤维素与半纤维素缔合的木质素鞘所致.木质素与半纤维素以共价键形式结合,将纤维素分子包埋在其中,形成一种天然屏障,使酶不易与纤维素分子接触,而木质素的非水溶性、化学结构的复杂性,导致了秸秆的难降解性.所以,要彻底降解纤维素,必须首先解决木质素的降解问题.因此,秸秆利用的研究从过去的降解纤维素的研究转向了木质的降解研究,作者对此进行了综述.1 木质素的结构特点木质素是由苯丙烷单元(θ—Cα—Cβ—Cγ)通过醚键和碳碳键联接的复杂的无定形高聚物,难以被酸水解,是天然高聚物中最难搞清楚的一个领域.其原因有两个方面,一是木质素的结构单元之间除醚键联接外还有C-C键;另一方面是不可能把整个木质素分子以其完整状态分离出来[2].典型木质素是由松柏醇(C oniferyl alcohol),芥子醇(Sinapyl alcohol)和对—香豆醇(p2C oumaryl alcohol)这3种不同的醇作为先体结构物质组成基本的结构单元,这些木质素结构单元常常以它们的糖苷形式贮存在细胞中,一旦需要合成木质素,它们便从这些糖苷中释放出来.这些木质素结构单元之间主要是通过醚收稿日期:2000-06-14基金项目:国家科委生物工程中心“九五”攻关项目(962C01202201)作者简介:吴坤(1963-),男,河南平舆人,河南农业大学副教授,在读博士,从事食品加工方面的研究. 350河 南 农 业 大 学 学 报第34卷键和碳碳键的方式联接.醚键包括酚醚键、烷醚键和二芳醚键.在酚醚键中愈创木基2甘油2β2芳基醚键(β2024)数量最多,占酚醚键的1/2左右,其次是愈创木基2甘油2α2芳基醚键(α2024).而木质素的C 2C 键的连接类型主要有β25,β2β,β21,β22和525等形式(图1).图1 木质素C 2C 键的连接类型2 木质素降解微生物的种类在自然界中,能降解木质素并产生相应酶类的生物只占少数.木质素的完全降解是真菌、细菌及相应微生物群落共同作用的结果,其中真菌起着主要作用.降解木质素的真菌根据腐朽类型分为:白腐菌———使木材呈白色腐朽的真菌;褐腐菌———使木材呈褐色腐朽的真菌和软腐菌.前两者属担子菌纲,软腐菌属半知菌类.白腐菌降解木质素的能力尤于其降解纤维素的能力,这类菌首先使木材中的木质素发生降解而不产生色素.而后两者降解木质素的能力弱于其降解纤维素的能力,它们首先开始纤维素的降解并分泌黄褐色的色素使木材黄褐变,而后才部分缓慢地降解木质素.白腐菌能够分泌胞外氧化酶降解木质素,因此被认为是最主要的木质素降解微生物[3,4]. 目前,研究最多的白腐菌有:黄孢厚毛平革菌(Phanerochete chrysosporium )、彩绒草盖菌(Coridus ver si 2color )、变色栓菌(Thametes ver sicolor )、射脉菌(Phlebia ra 2diata )、凤尾菇(Pleurotus pulmononanus )、朱红密孔菌(Pycnoporus cinnabarinus )等[3,4,5].这些菌多属于担子菌亚门,无隔担子菌亚纲、无褶菌目的多孔菌科.王佳铃等曾评述过1981~1995年间研究报道过的主要高产漆酶白腐真菌(表1)[6].表1 主要的高产漆酶微生物属 名 菌 种 名Polyporus P.ver sicolor ,P.hir sutus ,P.anceps ,P.sanguineus Coriolus (C.ver sicolor ),(C.hir sutus ),C.consor Pleurotus P.ostreatus ,P.sajor 2caju ,P.erynigii ,P.pulmonarius Lentinus L.edodes ,L.tigrinus Pycroporus P.cinnabarinus ,P.sanguineus ,P.coccineus Polystictus P.ver sicolor ,P.sanguineus Ganoderma G.luciolum ,G.valesiocim other s Agaricus bisporus ,Botrytis cinera ,Panus tigrinus ,Irpex lacuts ,Daedalea flavida ,Fomes annos ,Rhizoctonia practicola ,Neurospora crassa ,Lenzites be 2tulina ,Sporotrichum pulerulentum ,Pholiota mutabilis ,Schinus molle ,Sterumpurpureum ,Coprinus congregatus ,Cerrena unicolor ,Ceriporiopsis subvermis 2pora ,Bjerkaradera adusta ,Poliporus pinsitus ,Dichomitus sgualenss ,Pyricu 2laria oryzae ,Fyathus bulleri 褐腐菌的软腐菌中的有些种类也可以分泌一些降解木质素的酶类,但它们分解木质素的能力不是很强,因此,研究报道较少.降解木质素的原核生物以放线菌为主,如链霉菌属(Streptomyces )、节杆菌属(Arthrobaeter )、小单孢菌属(Micromonospora )和诺卡氏菌(Nocardia )等.细菌中有厌氧梭菌(Clostridum xylanoyticum )、假单孢菌(P seudomonas )、不动杆菌(Acineto 2bacter )和芽孢杆菌(Bacillus )等原核生物对于在土壤中木质素的转化和降解具有重要作用,但土壤中木质素的降解据报导是土壤微生物区系多种类型微生物协同进行的结果,而且进程缓慢,单一微生物纯培养的商业用途前景不很清晰,且这类原核生物的多酚氧化酶是胞内酶,这也决定了其在木质素降解菌的研究中处于一个相对从属的地位[5]. 此外,很多植物病原微生物如Rhizoctonia s olani ,R.praticola 等也具有分解木质素的能力,但这类微生物在处理秸秆上的应用前景不大.在木质素降解微生物中,研究得最多最彻底和最具应用前景的是黄孢原毛平革菌.这种微生物的特点第4期吴坤等:木质素生物降解研究进展351 是培养温度高(37℃左右),无性繁殖迅速,菌丝生长快且分泌木质素降解酶能力强,它已经成为研究白腐真菌的一种模式微生物.它的培养条件、代谢调控、分子生物学和遗传学及其在生产实践上的应用等均已被研究的相当仔细[5~8].3 木质素降解酶的种类及特性目前,关于木质素降解酶的研究工作主要集中在白腐菌所产生的酶系.研究得较多,并认为最为重要的木质素降解酶有3种,即木质素过氧化物酶(Lignin peroxidases/LiP)、锰依赖过氧化物酶(Manganese perxi2 dases/MnP)和漆酶(Laccase).LiP,MnP和Laccase只是分别代表一系列的同功酶.但各种微生物所产生的酶的种类和一些理化特性是有所不同的.其中LiP是第一个从黄孢原毛平革菌发现的木质素降解酶,以后又分别从射脉菌,彩绒革盖菌,变色栓菌中发现.Laccase没有在黄孢原毛平革菌中发现,而在Phlebia,Tram2 etes,Coriolus等菌中发现[7~9].木质素的降解酶系是非常复杂的一个体系,很多问题至今还不十分清楚.除了上述3种重要的酶外,其它的一些酶如芳醇氧化酶(AAO)、酚氧化酶、葡萄糖氧化酶、过氧化氢酶及一些还原酶、甲基化酶和蛋白酶等都参与或对木质素的降解产生一定的影响.LiP和MnP都是带有糖基的胞外血红素蛋白.又称血红素过氧化物酶(Heme Peroxidase),它们在反应中从苯酚或非酚类的苯环上夺取一个电子而使后者形成一个阳离子基团,从而导致木质素分子中主要键的断裂.LiP主要是氧化苯酚使之成为苯氧残基.MnP主要是从Mn2+和H2O2的氧化中得到Mn3+,然后Mn3+氧化苯酚使之成为苯氧残基.而每种酶在木质素降解中的具体作用还不甚清楚[5~7,9].漆酶(Laccase)是含铜的多酚氧化酶,主要来源于生漆和真菌.由于Laccase的含糖量高,难得到X2衍射分析用的漆酶单晶,它的三维空间结构尚不清楚.但已经证实,漆酶中的铜离子在催化氧化反应中起决定作用.根据磁学和光谱性质可将漆酶中的4个铜离子分为3类:Ⅰ型Cu2+和Ⅱ型Cu2+各1个,是单电子受体,呈顺磁性,可以用核磁共振控测;Ⅲ型Cu2+2个,是双电子受体,反磁性,用核磁共振不能检测.Ⅰ型Cu2+呈蓝色,在λ614nm处有特征吸收峰;Ⅱ型Cu2+为非蓝色,没用特征吸收峰;Ⅲ型Cu24+是偶合的离子对(Cu2+…Cu2+),在λ330nm处有宽的吸收峰[6,7,9,10].漆酶在木质素降解过程的机制和作用至今人们还不完全清楚.作为一种多酚氧化酶,它可催化氧化酚类或芳胺类等多种底物的氧化作用.氧化酚或芳胺先失去一个电子生成自由基,后者发生一系列非酶反应,氧化成醌,在有O2存在时,还原态漆酶被氧化,O2还原成水.底物自由基不稳定,可进一步发生键的断裂或生成,导致裂解或聚合反应.因此,底物分子可进一步聚合生成复杂的产物,也可催化芳香环支链Cα2 Cβ键断裂[6,7,10,11].G A LI ANO等发现,当Laccase和MnP分别单独存在时,都不能有效地降解木质素,而2种酶同时存在时则木质素得到有效的降解,这表明2种酶在催化木质素生物降解反应中具有协同作用.此外还发现在反应中加入其它的酶,如葡萄糖氧化酶时,可显著提高木质素的降解程度.他们认为,这是由于Laccase既有解聚,又有聚合木质素的能力,加入葡萄糖氧化酶可通过其还原Laccase氧化多酚产生的醌等阻止木质素的聚合作用,从而提高Laccase的降解能力[6].确定每一种酶在木质素降解过程中的作用和功能是困难的,因为至少目前木质素被微生物所产生的单一胞外酶彻底、完全地催化解聚和降解的情况还没被发现.每一种白腐真菌所产生的都不只是一种能降解木质素的酶,在缺陷任一种酶的体系中木质素仍能够较好的降解,而这些酶之间又相互作用和影响,使其存在于一个协同作用体系中.虽然现在已能用14C标识的合成木质素来研究木质素降解酶在木质素降解中的作用机理,但由于木质素的结构复杂,木质素的彻底降解需依赖微生物区系中多种微生物在较长时间内相互交替的生长繁殖才能完成.4 木质素的降解微生物的酶活性调控机理在白腐菌产酶的培养方式上,研究最多的是液体培养.国内外曾利用多种不同有机、无机或复合碳源和氮源来研究它们对微生物的木质素降解能力和产酶能力的影响.还研究了碳源或氮源限制对它们的影响.结果表明,碳源和氮源是微生物降解木质素和产酶的一个极为重要的影响因素[6~18]. 352河 南 农 业 大 学 学 报第34卷 在用不同的单糖、双糖作碳源的矿质培养基培养豹斑革菌时,用麦芽糖生物的生长量最高.在标准培养条件下,以葡萄糖作对照,甘露糖和麦芽糖使MnP 活性分别增2倍和318倍.酵母汁、牛肉膏等复合氮有利于LiP 活性的提高,而甘氨酸、柠檬酸铵、乙醇胺等则抑制LiP 的生成和活性[6~9].黄孢原毛平革菌等白腐菌的木质素降解和木质素降解酶的产生是发生在氮源或其它营养被消耗的次级代谢过程中.而细菌则与之相反,高浓度的有机或无机氮源都不能抑制木质素降解,所产生的过氧化物酶也具有与生长相联和不受高水平氮抑制的特点.我国学者李越中等曾对黄孢原平革菌合成木质素过氧化物酶的营养调控进行过较为详细的研究报道(表2、表3)[19].表2 不同碳源对黄孢原毛苹革菌木素酶活力的影响碳源/(2g ・L -1)酶活力/(U ・L -1)碳源/(2g ・L -1)酶活力/(U ・L -1)三碳糖 蕈糖26512 甘油12613非还原性三糖戊醛糖 D 2松三糖7518 D 2核糖16810 上述糖等比混合40010 D 2木糖22713吡喃型戊聚糖 L 2阿拉伯糖12211 木聚糖1216己醛糖呋喃型己聚糖 D 2葡萄糖32412 葡糖4613己酮糖吡喃型己聚糖 L 2山梨糖10110 糊精42914脱氧己糖 可溶性淀粉4211 D 2岩藻糖 0 微晶纤维素17216 L 2岩藻糖17618 地衣多糖19317 鼠李糖 412 海带多糖16412还原二糖其他 纤维二糖28616 葡甘聚糖2915非还原二糖 果胶14714 D 2松二糖 0 麸皮浸汁(1%)25618 蔗糖5417表3 不同氮源对黄孢原毛平革菌酶活力的影响氮源 菌丝重/mg 酶活力/(U ・L -1)无机氮 NH 4NO 3141223711 (NH 4)2S O 4141328316 (NH 4)2HPO 417119810 尿素19120有机氮 乙醇胺20110 丙烯酰铵181524718 石酸铵131735016 柠檬酸铵16190 甘氮酸21140 L 2谷氨酸201325715复杂氨 酵母浸汁181338912 牛肉膏201743619 注:氮源质量浓度为12mm ol ・L -1. 有些微量元素对于白腐菌相关酶类的产生是有较大影响的.漆酶是一种含铜的多酚氧化酶,因此,Cu 对于漆酶的产生和酶活性是相当重要的.Mn 对LiP 和MnP 都具有重要的作用,对漆酶的影响也是显著的.BI NNARME 等认为,Mn 2+对LiP 有抑制作用,而对MnP 则有诱导作用,其作用的大小与Mn 2+的浓度及添加时间有关. C.subuermispora 在Mn 2+浓度为11×10-6m ol ・L -1时,木质素矿化速率最大,而MnP 和Laccase 的活性在410×10-6时最高.不添加Mn 2+可使Agaricus campestris 的Laccase 活性显著降低,但Mn 2+对其影响是以Cu 2+的存在为前提,如果没有Cu 2+存在,那么Laccase 的活性则与Mn 2+浓度无关且酶活性始终很低.有报道,高浓度的Mn 2+会提高Phlebia breispora 的Laccase 活性,但对Dichomitus 来说,Laccase 活性都不受Mn 2+的影响. 结构和木质素有关的低分子芳香化合物或木质素降解的碎片化合物,可诱导Laccase 产生并提高其酶产量,这类化合物如香草酸、藜芦醇、愈创木酚、吐温、甲苯胺等.在豹斑革耳菌上还发现了专一的MnP 诱导物32甲基苄醇.培养温度、通气状况,pH 值乃至环境湿度等,也是影响木质素降解酶产生的因素.如黄孢原毛平革菌的最适生长温度为37℃,最适产酶温度为30℃.豹斑革耳菌增加通气可使木质素矿化率有所提高,而对于Phlebia radiata 来说,通氧则抑制其酶活性.培养温度和pH 值对Laccase 的影响因具体菌种而异,不同的菌种有不同的适宜培养温度和pH 值.5 木质素降解的分子生物学特性黄孢原毛平革菌(P.chrysosporium )已演变成为白腐真菌研究的模式生物,因此,有关这类微生物的研究中,对P.chrysosporium 的研究是最为详细的,当然,在分子生物学领域的研究也不例外[7].511 基因组在P.chrysosporium 菌丝体细胞中,可以随机分散多个细胞核,担孢子则含有2个等同的核.有性孢子的形成可以是同宗配合或异宗配合.黄孢原毛平革菌ME 2446菌株基因组大小估计为414×107bp ,其中20%~30%是线粒体和核糖体DNA ,其担孢子呈现染色体长度多态性.第4期吴坤等:木质素生物降解研究进展353 512 相关的基因家族51211 LiP 现在已知编码P.chrysosporium LiP的至少10个相近基因的家族,定名从lip A至lip J.从其它真菌如Trametes ver sicolor,Bjerkandera adusta和Phlebia adusta中也克隆出了几个LiP基因.LiP基因序列是高度保守的,其氨基酸序列有53%~9819%的相似性.每个LiP基因编码一个由22~28个残基的引导肽的343~345个氨基酸的成熟蛋白质产物.成熟蛋白质的分子量为36360~36607,表现分子量38000~43000,表现分子量的6%~13%推测为糖基部分.lip编码区含有大约60%~65%G+C,而非编码区G+C大约为44%~49%.P.chrysosporium lip均含有8个或9个内含子排列,每个大小为49~78bp.已测序的lip基因的5′端非编码区包含一个T AT A框(-66~-81bp)和一个C AAT顺序(-107~-228bp)的调节序列.51212 MnP 和LiP一样,MnP同功酶也是由多基因编码的.菌株OG C101的MnPⅠ编码一个357个氨基酸的成熟蛋白及一个21个氨基酸的导肽.它也具有糖基化位点,因此MnP也是糖蛋白.MnPⅠ的内含子为6个,大小在57~72bp之间;MnPⅡ的内含子为7个,大小在50~55bp之间.mnp 5′非翻译区包括一个T AT AA单元(-81bp)及3个反向CC AAT单元.51213 Laccase 漆酶也是由一个结构相近的基因家族所编码,目前至少有17个真菌的Laccase基因已经被克隆和测序,如Rhivoctonia solani,Trametes villosa,Agericu bisporus,Neurospora crassa,Coriclcls hir sutus,Cry2 phonectria parasitica,Aspergillus nidulan和Pleurctus ostleatus等.总的来说,在2个真菌Laccase之间的顺序等同性较低,但在铜结合区保守性却很高[9,20~24].513 相关基因的表达木质素降解酶基因的转录和翻译情况比较复杂,如菌株BK M2F21767LiP基因的转录明显地受C,N水平的调节,而MnP的产生赖于Mn的浓度,且Mn的调节是在转录水平上的.漆酶的表达调节在种间有显著的不同[7,9,20~24].在异源表达上,总的来说表达水平均不是太高.但是,一个构巢曲霉(Aspergillus nidulan)漆酶基因在米曲霉(A.aryzae)上的重组和成功表达,已使这个重组菌株用于实际的商业生产.6 木质素的生物降解的应用木质素的生物降解目前成功地用于生产实践的实际应用尚不多见,但在有些方面的研究已经显现出诱人的前景.1)造纸工业 分解木质素的酶类在造纸工业上的应用有两个方面,一是用改造旧的造纸工艺,用于生物制浆、生物漂白和生物脱色.黄孢原毛平革菌和P.brevispora等在国外已经得到成功利用.如用P.brevis2 pora进行生物制浆预处理可降低47%的能耗并增加了纸浆的张力,但它们的木质素降解率和产酶量都还是极为有限的,处理时间过长,距大规模推广应用尚有一定的距离.二是木质素分解菌或酶类用于造纸废水的处理,这方面的国内外研究报告已有很多且已取得了一定的实效[9].2)饲料工业 木质素分解酶或分解菌处理饲料可提高动物对饲料的消化率.实际上,木素酶和分解菌的应用已经突破了秸秆仅用于反刍动物饲料的禁地,已有报道饲养猪、鸡的实验效果.目前,以木素酶、纤维素酶和植酸酶等组成的饲料多酶复合添加剂已达到了商品化的程度.3)发酵与食品工业 木质纤维素中木质素的优先降解是制约纤维素进一步糖化和转化的关键,已有很多实验偿试使用秸秆进行酒精发酵或有机酸发酵,但看来这还有很长的路要走.在食品工业如啤酒的生产中,可使用漆酶等进行沉淀和絮凝的脱除,使酒类得到澄清.4)生物肥料 传统上曾使用高温堆肥的办法来使秸秆转化为有机肥料,但这些操作劳动强度大,近年来不为农民所欢迎.最近,秸秆转化为有机肥料的简单而行之有效的办法是秸秆就地还田.但是,还田秸秆在田间降解迟缓并带来了一系列的耕作问题,而解决这些问题的关键是加速秸秆的腐熟过程,因此,以白腐菌为代表的木质素降解微生物为这种快速腐熟提供了理论上的可能性.在国内,已有几家科研单位在进行相关的研究与探索.5)环境保护[2,6,9,25] 降解木质素的主要酶系是过氧化物酶系,它们作用于底物的机制是夺取电子和自由基的形成,这些特点决定了它们的底物非专一性,即它们的底物不是一种而是一类或几类有机化合 354河 南 农 业 大 学 学 报第34卷物.以漆酶为例,它是含铜的多酚氧化酶,它的底物涵盖了多种芳胺、酚类、二茂铁及共衍生物.季立才等人最近又确证了40多个漆酶的新发现底物,如1152萘二胺、1152二氨基蒽醌、邻和对乙酰氨基苯酚、22氨基242硝基苯酚、32氨基242羟基苯磺酸、32羟基242氨基萘磺酸、原儿苯酸硝类、茜素红、茜素紫、醌茜素、苏木精等.鉴于木质素降解菌和它们产生的相关酶类对多种有机化合物的降解能力,因此它们也是化工废水处理研究最为活跃的领域之一,这方面已不乏成功的范例.另外,在国外,黄孢原毛平革菌已被成功地用于有机氯农药污染土壤的生物修复.参考文献:[1] CRAWFORD R L.Lignin biodegradation and trans formation[M].New Y ork:John Wiley&S ons,Inc,1981.[2] HIG UCHI T.Lignin biochemistry:biosynthesis and biodegradation[J].W ood Sci T echnol,1990,24:23-63.[3] BUSWE LL J A,ODIER E.Lignin biodegradation[J].CRC Crit Rev Biotechnol,1987,6:1-60.[4] KIRK T K,FARRE LL R L.Enzymatic“combustion”:The microbial degradation of lignin[J].Ann Rev M icrobiol,1987,41:465-505.[5] 管莜武,张甲耀,罗宇煊.木质素降解酶及其调控机理研究的进展[J].上海环境科学,1998,17(11):46-49.[6] 王佳玲,余惠生,付时雨,等.白腐菌漆酶的研究进展[J].微生物学通报,1998,25(4):233-235.[7] MICH AE L H G,A LIC M.M olecular biology of the lignin2degrading Basidiomycete Phanerochaete chrysosporium[J].M icrobiol Rev,1993,57(3):605-622.[8] 卢雪酶,李越中,王蔚,等.黄孢原毛平革菌木素过氧化物酶类在天然木素降解中作用的研究[J].菌物系统,1998,17(2):179-184.[9] C U LLE N D.Recent advances on the m olecular genetic fungi[J].J Biotechnol,1997,53:273-289.[10]季立才,胡培植.漆酶催化氧化反应研究进展[J].林产化学与工业,1997,17(1):79-83.[11]Y OUN H,H AH Y,K ANG S.R ole of laccase in lignin degradation by white2rot fungi[J].FE MS M icrobiol Lett,132:183-188.[12]FAIS ON B D.,KIRK T K.Factors inv olved in the regulation of a ligninase activity in Phanerochete chrysosporium[J].Appl EnvironM icrobiol,1985,49(2):299-304.[13]TIE N M,KIRK T K.Lignin2degrading enzyme from Phanerochaete chrysosporium:Purification,characterization,and catalytic propertiesof a unique H H2O22requiring oxygenase[J].Proc Natl Acad Sci US A,1984,81:2280-2284.[14]FREIT AG M.,M ORRE LL J.Changes in slected activities during growth of pure and mixed cultures of the white2rot decay fungusTrametes ver sicolor and potential biocontrol fungus Trichoderma har zianum[J].Can J,1992,38:317-323.[15]SH UTT LEW ORTH K,BO LLAGJ.Production of induced laccase by the fungus Rhizoctonia practicola[J].Can J M icrobiol,1986,32:867-870.[16]ROTHSCHI L D N,LE VK OWITZ A,H ADAR Y,et al.Manganese dificiency can replace high oxygen levels needed for lignin peroxi2dase formation by Phanerochaete chrysosporium[J].Appl Environ M icrobiol,1999,65(2):483-488.[17]ORTH A,ROY SE D,TIE N M.Ubiguity of lignin2degrading peroxidases am ong various w ood2degrading fungi[J].Appl Environ M icro2biol,1993,59(12)4017-4023.[18]M AS APHY S,LE VANON D.The effect of lignocellose on lignocellulolytic activity of Puleurtus pulmonrius in submerged culture[J].Appl M icrobiol Biotechnol,1992,36:828-832.[19]李越中,高培基,王祖农.黄孢原毛平革菌合成木质素过氧化物酶的营养调控[J].微生物学报,1994,34(1):29-39.[20]LIWICKI R,PATERS ON A,M AC DONA LD M,et al.Phenotypic classes of phenoloxidase2negative mutants of the lignin2degrading fun2gus Phanerochaete chrysosporium[J].J Bacteriol,1985,162(2):641-644.[21]Y AVER D,OVER J ERO M,X U F,et al.M olecular characterization of laccase gene from the Basidiomycete cinereus and heterolog ousexpress of the laccase lcc l[J].Appl Envirom M icrobiol,1999,65(11):4943-4948.[22]GE LPKE M,M ARY M,CEREG HI NO G,et al.H om olog ous expression of recombinant lignin peroxidase in Phanerochaete chrysospori2um[J].Appl Environ M icrobiol,1999,65(4):1670-1674.[23]BERK A R,SCH NEI DER P,G O LIG HT LY E,et al.Characterization of the gene encoding on extractcellar laccase of Mecsliophthorathermophila and analysis of the recombinant enzyme expressed in Aspergillus oryzae[J].Appl Environ M icrobiol,1997,63(8):3151 -3157.[24]K O J I M A Y,TS UK UDA Y,K AW AI Y,et al.Cloning,sequence analysis,and expression of ligninolytic phenoloxidase genes of thewhite2rot Basidiomycete Coriplus hir sutus[J].J Biological Chem,1990,265(25):15224-15230.[25]季立才,王光辉,胡培植.漆树漆酶的新底物[J].自然杂志,1990,13(3):189-190.。
木质生物质的生物分解及生物转化

木质生物质的生物分解及生物转化木质生物质指的是植物通过光合作用生成的有机资源,主要由纤维素、半纤维素和木质素3种高分子物质构成,如树木、农作物秸秆等。
全世界每年由光合作用生成的木质生物质约2 000亿t,相当于全世界每年消费能源的10倍,其中树木固定的木质生物质占整体的90%~95%(赵广杰,2004)。
然而,我国目前却存在着对木质生物质资源的严重浪费。
如我国木材加工工业采伐、加工剩余物每年就达到1 000万t(刘家建,1995);农业生产中的剩余物(农作物秸秆)每年也多于5亿t(吴坤等,2000),有效利用的只占很少一部分,大部分被浪费掉。
同时在人们的日常生活中还会有大量弃置不用的木质材料,我国年产废弃木质材料360万t,如废旧家具、木制包装,建筑施工时遗弃的废木料(每年300万m3),道路及城市园林绿化系统更新的树木等(王珊珊等,2004)。
这些废弃木质材料的处理要花费大量的资金,占用大量的土地。
因此有专家提出关于废弃木质材料回收及再生利用的建议(崔积常等,2004)。
在美国,把废弃的木质材料称为“第四种森林”,是倒在地上的森林(王珊珊等,2004)。
这些材料如果能被再次利用,势必产生巨大的经济效益和社会效益。
随着经济和社会的发展以及人口的增长,对粮食和能源的需求也在逐渐增长,面对着不可更新的化石资源面临枯竭,粮食和能源危机成了当前人类发展所面临的重要问题。
用微生物酶可以将木质生物质材料分解转化,得到葡萄糖和各种低聚糖作为人类的营养品和保健品,得到菌体蛋白可以作为人类食品和动物饲料,葡萄糖继续发酵还可得到乙醇、甲烷等清洁燃料。
因此,木质生物质作为一种可更新的有机资源,对于解决粮食和能源问题以及城市废弃木质资源有双重意义。
同时这种木质材料的生物转化过程又不会对环境造成污染,有很好的发展前景,近些年来愈来愈受到人们的关注和重视。
1·1多糖的生物分解木材中的多糖指的是纤维素和半纤维素。
纤维素是一切植物体的主要组成部分,由β-D-葡萄糖通过β-1,4糖苷键相结合而形成的链状高分子聚合物,聚合度在7 000~10 000之间,占植物体干质量的50%左右,地球上每年由光合作用生成的纤维素约 1 100亿t(张全福等,1994;Kubiceket al.,1993),是最丰富的木质生物质资源。
木质素降解研究

木质素降解技术还可应用于生物柴油 的生产,通过将木质素转化为脂肪酸 酯,可获得具有高能量密度的生物柴 油。
有机肥料的制备
有机肥料的生产
木质素降解过程中产生的酚类物 质可以与氨基酸结合,形成腐殖 质,这种物质具有较高的肥效, 可以作为有机肥料使用。
有机肥料的生产
腐殖质不仅可以提供植物所需的 营养元素,还具有改善土壤结构 、提高土壤保水保肥能力的作用 。
高分子材料
利用木质素降解产物合成高分子材料,如聚合物、 树脂等。
生物农药
木质素降解产物中的某些化合物具有生物活性, 可用于开发新型生物农药。
木质素降解过程中的环境影响
有机物排放
木质素降解过程中会产生一些有 机物,如挥发性有机酸和酚类物 质,可能对环境造成一消耗一定 的营养物质,如氮、磷等,可能 对水体和土壤环境产生一定影响。
木质素降解研究
$number {01}
目 录
• 木质素降解概述 • 木质素降解的微生物种类 • 木质素降解的机理研究 • 木质素降解的应用前景 • 木质素降解的挑战与展望
01
木质素降解概述
木质素定义与特性
总结词
木质素是一种天然高分子聚合物,主要由苯丙烷单元通过醚 键和碳-碳键连接而成,具有复杂的结构。
优化微生物种类
01
通过筛选和驯化具有高效降解木质素的微生物,提高木质素的
降解效率。
基因工程技术
02
利用基因工程技术对微生物进行改造,增强其木质素降解能力。
物理化学预处理
03
通过物理或化学方法对木质素进行预处理,降低其结晶度和聚
合度,提高微生物对其的接触和利用。
木质素降解产物的利用
生物质能源
木质素的结构与生物降解性能

木质素的结构与生物降解性能木质素是一种存在于植物细胞壁中的复杂有机高分子化合物,它在植物界中广泛存在,尤其是在木质植物中的细胞壁中含量较高。
木质素的结构与生物降解性能是研究木质素功能和利用的重要内容。
首先,我们需要了解木质素的结构。
木质素具有多种不同结构的类似物,但最常见的木质素是由三个苯环通过间苯基和间甲基(3,4-二羟基苯基)连接在一起的共轭骨架构成。
这些苯环可以是单体或杂环,而共轭骨架可以使木质素具有很强的稳定性和抗降解性能。
除了共轭骨架,除去的羟基和甲基功能基团也对木质素的结构和性能发挥重要作用。
羟基基团在木质素中可承担自由基抗氧化作用,增加结构稳定性。
而甲基基团则与其他细胞壁聚合物(如纤维素和半纤维素)交互作用,增强细胞壁的机械强度。
然而,木质素的结构也限制了其生物降解性能。
由于共轭骨架和稳定的结构,纯净的木质素难以被微生物降解。
这使得木质素在环保工业和可再生能源领域的利用受到限制。
为了提高木质素的生物降解性能,研究人员通过多种途径进行了改性。
一种方法是使用化学手段改变其结构,如通过氧化、酚醛解聚等方法。
这些方法可以改变木质素的化学键和功能基团,使其更易于微生物降解。
然而,这些方法通常需要使用高温和强酸等条件,且效果不佳。
另一种方法是利用酶类来催化木质素的降解。
许多微生物,包括真菌和细菌,可以分泌特定的木质素酶来降解木质素。
这些酶可以打断木质素的结构,使其更易于微生物利用。
此外,还可以利用基因工程技术来改造微生物,使其能够高效降解木质素,并产生有经济价值的产物,如乙醇和生物柴油。
除了改变木质素本身的结构,还可以通过改变植物的基因来影响木质素的含量和结构。
一些研究发现,通过调控特定基因的表达,可以增加或减少木质素的积累。
此外,改变基因表达还可以调控木质素的组成,进一步提高其降解性能。
总的来说,木质素的结构与生物降解性能密切相关。
其共轭骨架和稳定的结构使得纯净的木质素难以被微生物降解,限制了其在环保工业和可再生能源领域的应用。
1微生物对木质纤维素类物质的降解

纤维素的可及性(Accessibility )
在不溶性纤维素水解之前,纤维素酶必 须结合到底物颗粒的表面。
纤维素颗粒的三维结构以及纤维素酶的 大小和形状,决定了纤维素酶能否进攻 到β–1,4–葡萄糖苷键,即纤维素酶能 否发挥其水解纤维素的作用。
纤维素颗粒的内表面取决于毛细作用的结构和粒子 内的孔道(1-10nm)及粒子间的空间(>5um) 。 而 纤维素颗粒的外表面积则与其形状和颗粒大小相关。
在酸处理过程中,半纤维素脱乙酰基产生乙 酸和糖类降解产生的少量糠醛具有毒性,对 后期的发酵有一定的抑制,通常采用离子交 换、过量石灰中和等措施脱毒,或选育和使 用能抗毒性的发酵菌。
纤维素DP对酶解速率的作用主要表现在对外切酶 的影响。外切酶作用于链端,因此聚合度较低的纤 维素底物具有更多的外切酶作用位点。
水解过程中DP的变化则取决于外切和内切酶的比 例以及纤维素的性质。由于外切酶在链端依次水解 纤维素链,因此只能有限降低DP,而内切酶作用 于链内,能快速降低DP,导致链端的大量增加。
在不同植物纤维原料中,木质素的结构不同,即使 在同一原料不同部位,木质素的结构也不相同。
木质素在结构上具有庞大性和复杂性,在化学上具 有极不稳定性,当受到化学试剂、温度、酸度影响 时,都会发生化学变化,即便在较温和的条件下, 也会发生缩合作用。
迄今为止,没有一种方法能得到天然木质素,这给 木质素研究造成一定困难。
纤维素的聚合度(Degree of Polymerization, DP )
纤维素的水溶性随DP的升高而迅速降低。聚合度 为2-6的纤维糊精可溶于水,而聚合度为7-13或 更高的纤维糊精微溶于热水。DP为30的葡聚糖 已经具有纤维素聚合物的结构和性质特点。
根据底物的来源和预处理方法的不同,纤维素底 物的DP变化非常大。
木质素的微生物降解
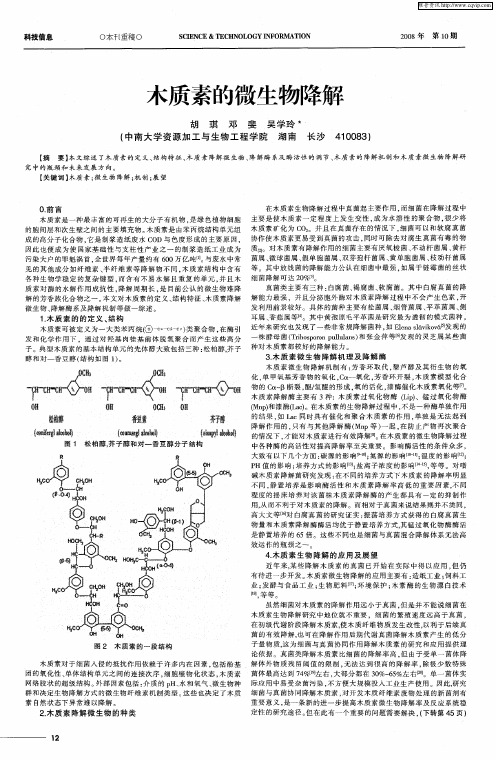
图 1 松 柏 醇 , 子 醇 和对 一 香 豆 醇 分 子 结 构 芥
一
醇 和 对 一 香 豆 醇 ( 构如 图 1 。 结 )
3木 质 素 微 生物 降解 机 理 及 降解 酶 .
木 质 素 微 生 物 降 解 机 制 有 : 香 环 取 代 , 芦 醇 及 其 衍 生 物 的 氧 芳 黎 化 , 甲 氧 基芳 香 物 的 氧 化 . d 氧化 , 香 环 开 裂 , 质 素模 型 化 合 单 C一 芳 木
在 木 质 素 生 物 降解 过程 中真 菌 起 主要 作 用 , 而细 菌 在 降解 过 程 中 主 要 是 使 木 质 素 一 定 程 度 上 发 生 变 性 , 为 水 溶 性 的 聚 合 物 , 少 将 成 很 木质素矿化为 C O 。并 且 在 真 菌存 在 的情 况 下 , 菌 可 以和 软 腐 真 菌 细 协 作 使 木 质 素 更 易受 到 真菌 的攻 击 , 同时 可 除 去 对 腐 生 真菌 有 毒 的 物 质 。对 木 质 素 有 降 解 作 用 的 细菌 主要 有 厌 氧 梭 菌 、 动 杆 菌属 、 杆 不 黄 菌 属 、 球 菌 属 、 单 胞 菌属 、 芽孢 杆 菌 属 、 单 胞 菌属 、 动 杆 菌 属 微 假 双 黄 枝 等 。其 中放 线 菌 的 降解 能 力公 认 在 细 菌 中最 强 , 如属 于链 霉 菌 的 丝 状 细 菌 降 解 可 达 2 %口 0 】 。 真菌类主要有三种 : 白腐 菌 、 腐 菌 、 腐 菌 。其 中 白 腐 真 菌 的 降 褐 软 解 能 力 最 强 .并 且 分 泌 胞外 酶对 木 质 素 降解 过程 中 不会 产 生 色素 , 开 发 利 用 前 景 较 好 。 具 体 的 菌种 主要 有 栓 菌 属 、 管菌 属 、 革 菌 属 、 烟 平 侧
木质素的生物降解及其应用

第20卷第6期2010年12月皮革科学与工程LEATHER SCIENCE AND ENGINEERINGVol.20,No.6Dec.2010文章编号:1004-7964(2010)06-0027-05木质素的生物降解及其应用李海涛,姚开*,何强,贾冬英(四川大学轻纺与食品学院,四川成都610065)收稿日期:2010-04-18基金项目:国家公益性农业科研专项基金(200803033-A004C )第一作者简介:李海涛(1985-),男,河南永城人,硕士研究生,研究方向为食物资源化学,E-mail :lihaitaoyong@ 。
通讯联系人:姚开,教授,E-mail :yaokai555@ 。
摘要:综述了具有降解木质素能力的微生物和酶的种类及其降解特性,阐述了木质素生物降解在生物化学制浆、纸浆生物漂白、造纸废水生物处理、饲料工业、生物肥料等领域的应用现状,展望了木质素生物降解技术研究的发展趋势。
关键词:木质素;生物降解;微生物;酶;应用中图分类号:Q 948.12文献标识码:ABiodegradation and Applications of LigninLI Hai-tao ,YAO Kai *,HE Qiang ,JIA Dong-ying(College of Light Industry and Food Engineering ,Sichuan University ,Chengdu 610065,China )Abstract :In this paper ,the types and the degradation characteristics of ligninolytic microorganisms and lignin-degrading enzymes are introduced.In addition ,the practical applications of lignin biodegradation in biochemical pulping ,biological bleaching of pulp ,biological treatment of papermaking wastewater ,feed industry and bio-fertilizer are summarized.And tech-nology trends in lignin biodegradation are outlook.Key words :lignin ;biodegradation ;microbes ;enzymes ;applications木质素资源十分丰富,是植物光合作用制造的总量仅次于纤维素的有机化合物,估计全球的木质素年产量可达1500亿t 。
细胞中的纤维素和木质素的生物降解机制

细胞中的纤维素和木质素的生物降解机制纤维素和木质素是植物细胞壁中的主要组成部分,其对植物的结构和功能至关重要。
但是,由于其高度的复杂性和纤维素结晶的强度,造成它们的降解相当困难,需要各种酶类蛋白的协同作用。
在大多数情况下,纤维素和木质素的生物降解主要是由微生物完成的。
其中最重要的为真菌、细菌、古菌等在环境中存在的微生物。
而微生物中则包括了各种能够分解木质素和纤维素的菌种,这些菌种具有高度的特异性,根据不同的微生物分类、生长条件,木质素和纤维素的生物降解机制也会发生变化。
纤维素的生物降解纤维素是由许多β-葡聚糖链相互交织而成的复杂生物高分子物质,是植物细胞壁中最丰富的组分之一,其结晶区晶胞面阵列堆叠排布,大量的交联构造使其热稳定性和机械强度都非常高,极其难以被降解。
微生物中,具有降解纤维素能力的菌只有极小的一部分。
通常,能够降解纤维素的微生物可以产生细胞外酶来降解周围环境中的纤维素。
纤维素酶主要分为三种,即聚糖酶、催化酶和酯酶,其中聚糖酶和催化酶主要针对纤维素分子中的β-葡聚糖链进行效果显著,而酯酶则主要作用于纤维素和木质素中的脂肪酰基。
在细菌和古菌中,分解纤维素的主要酶包括细菌纤维素酶(Cellulases)、纤维素乙酰化酶(Acetylesteretse)、纤维素结合蛋白(Cellulose-binding protein)等。
真菌能够分泌Cellulases、Acetylesteretse、β-glucosidases、xylanases、hemicellulases等降解酶类。
由此可见,虽然纤维素作为植物细胞壁的主要成分,但是被真菌、细菌等微生物降解时,需要多种复杂酶从分子链中逐渐降解出单糖分子才能真正被分解,其过程相当复杂。
木质素的生物降解与纤维素相比,木质素是含氧的天然高分子化合物,具有独特的结构和环节,是植物细胞壁中不可分割的一部分。
不同于纤维素,由于木质素有一定的芳香环结构和立体构型,其降解极其复杂。
木质素的微生物降解

木质素的微生物降解木质素是三种苯丙烷单元通过醚键和碳碳键相互连接形成的具有三维网状结构的生物高分子,存在于木质组织中,主要作用是通过形成交织网来硬化细胞壁,为次生壁主要成分。
木质素主要位于纤维素纤维之间,起抗压作用。
在木本植物中,木质素占25%,是世界上第二位最丰富的有机物(纤维素是第一位)。
由于自然界中木质素与纤维素、半纤维素等往往相互连接,形成木质素-碳水化合物复合体(Lignin-Carbohydrate Complex),故目前没有办法分离得到结构完全不受破坏的原本木质素。
木质素是自然界中含量丰富的三大生物质之一,占陆生植物干重的15%−40%,其结构为由苯丙烷单元结构通过醚键和碳碳键组成的具有三维空间结构无定型芳香类化合物,与纤维素和半纤维素构成植物骨架的主要成分。
木质素与纤维素、半纤维素结合十分稳定,不易降解,是生物质能源利用的关键瓶颈之一,同时木质素也是木材水解工业和造纸工业的副产物以及城市生活垃圾中一种较难降解的物质,采用化学试剂处理这些废弃物会造成严重的环境污染。
尽管针对木质素的降解方法已经有了几十年的研究,但是仍然存在许多的问题,例如化学法不能够解决污染问题,而物理法又有高能耗的挑战,生物法对环境友好但是效率较低,因此仍然是生物能源领域研究的热点和难点。
木质素在自然界中的完全降解是真菌、细菌及相应微生物群落共同作用的结果,其中真菌降解木质素的研究最为广泛和深入,但是到目前为止仍未开发出具有商业价值的生物利用木质素工业途径,以真菌为主要模式菌株开展的木质素生物降解研究长期未能突破。
由于广泛的生长条件和良好的环境适应能力,细菌在木质素降解方面深受研究人员的关注,成为国际上的研究热点之一。
在降解机制、新种属筛选、调控机理以及工业化的可能性等方面进行了大量深入的研究,也取得了显著的成就。
要想知道什么是微生物降解请到来进行了解,我们每期都会为大家介绍更多的固体废弃物安全小知识。
微生物酶解木质素的研究进展

微生物酶解木质素的研究进展木质素是植物细胞壁中的一种关键组分,由于其坚硬的结构和高度的抵抗性,对于生物高效的利用一直是一个难题。
微生物酶解木质素是一种有效的途径,可以将其转化为有用的化学品、生物能源等,对于提高可持续生产和环境保护等方面具有广泛的应用前景。
本文将从微生物酶解木质素的产物、酶学机制、应用前景等方面进行论述。
一、微生物酶解木质素的产物微生物酶解木质素通常可以产生多种木质素单体和多酚化合物,这些产物在生物能源、化学品、医药等方面具有广泛的应用前景。
典型的产物包括:单体的糖苷、芳香族化合物和羟基化产物(如羟基苯甲酸、羟基苯乙酸等)、酚类产物(如2,6-二甲基苯酚、2-甲基-1,4-苯二酚等)、酮类、甲酰基产物等。
同时,微生物酶解木质素还能转化为乙醇、氢气等生物能源,这些产物都有很高的经济价值和环境意义。
二、酶学机制微生物酶解木质素的酶学机制比较复杂,涉及多种不同的酶类和协同作用。
其中,laccase酶是对木质素的氧化解聚最具代表性的酶类之一,可以将木质素分解为较小的单体和多聚体的氧化产物。
另外,极微生物产生的黑汁酸酶(dark fermentative acidogenic bacteria)可以将木质素通过酸解作用转化为醋酸、甲酸和氢气等物质,这些产物是开展糖醇生产和生物能源开发的重要原料。
目前,在酶学机制的深入研究中,同时也有多种基因工程技术和合成生物学技术的应用,为生产高效酶和构建全新的木质素酶解系统提供技术支持。
三、应用前景微生物酶解木质素的应用前景非常广泛,具体表现在以下几个方面。
1. 生物能源方面:随着生物燃料的发展和需求的增加,微生物酶解木质素所产生的乙醇、氢气等生物能源成为了替代石化能源的有力选择,具有很好的经济效益和环境效益。
2. 化学品方面:微生物酶解木质素所产生的多重羟基产物、酚类产物等都是重要的有机化学品原料,可广泛应用于高性能涂料、聚合物等化学品的生产与加工等领域。
木质素低温降解总结

木质素低温降解总结木质素是天然生物质中的主要组成部分之一,它们在生物质能源的转化和生物降解中起着重要的作用。
木质素的低温降解是指在相对较低的温度下,木质素发生化学反应,从而引起其结构和性质的改变。
这一过程可以通过多种方式来实现,如酸催化、碱催化、生物降解等等。
本文将对木质素低温降解的一些主要反应和机理进行总结,并讨论其应用前景。
首先,木质素的低温降解反应可以通过酸催化来实现。
酸催化反应是指在酸性条件下,木质素发生裂解和转化的反应。
酸催化反应主要涉及到木质素结构中的酚羟基和酯基等官能团的裂解,从而产生醛、酮和酸等化合物。
这些裂解产物可以用于合成有价值的化合物,如酮和醛可以用作溶剂、表面活性剂和胶粘剂等。
其次,碱催化也是一种常用的木质素降解方法。
碱催化反应是指在碱性条件下,木质素的酸性官能团发生转化的反应。
碱催化反应主要涉及到酸性官能团的去酸化和酯化等反应,从而产生酯和醚等化合物。
这些产物可用于合成润滑剂、树脂和染料等。
此外,生物降解也是一种重要的木质素低温降解途径。
生物降解是指在生物体内,通过微生物和酶的作用,将木质素降解为小分子的有机化合物。
这一过程主要涉及到木质素的醛基、酮基和羧基的还原和氧化等反应。
生物降解可以在较温和的条件下进行,并且产物多为有机酸、醇和酮等物质,具有较高的降解效率和产物利用率。
综上所述,木质素的低温降解主要涉及到酸催化、碱催化和生物降解等反应。
这些反应可通过调控温度、酸碱性条件和降解体系中的微生物和酶的选择来实现。
木质素低温降解不仅对于生物质能源转化有重要意义,也对于木质素的高值利用具有重要意义。
通过降解产物的分离和提纯,可以获得多种有机化合物,如生物柴油、生物塑料和生物活性物质等。
因此,进一步研究木质素低温降解的反应机理和优化降解条件,对于提高木质素资源利用率和开发新型木质素材料具有重要意义。
木质素的微生物降解_secret

木质素的微生物降解摘要:综述了木质素微生物降解酶系及酶活性调控措施,介绍了木质素降解机制及木质素微生物降解的应用。
关键字:木质素微生物降解木素降解酶系木质素是植物的主要成分之一,占植物细胞化学组成的15%~30%。
在植物体中,木质素是包裹在纤维素的外面,功能之一就是保护植物细胞不受外界微生物的侵蚀。
从生物合成过程研究得知,木质素首先是由葡萄糖发生芳环化反应形成莽草酸(Shikirnic acid),然后由莽草酸合成三种具有苯丙烷结构的基本单元,分别为:对香豆醇、松柏醇、芥子醇。
从化学结构看,针叶树的木质素主要由松柏醇的脱氢聚合物构成愈创木基木质素;阔叶树的木质素由松柏醇和芥子醇的脱氢聚合物构成愈创木基—紫丁香基木质素;草本植物则由松柏醇、芥子醇和对香豆醇的脱氢聚合物和对香豆酸组成[1]。
木质素是结构复杂、稳定、多样的无定形三维体型大分子,在自然界中,降解缓慢,成为地球生物圈中碳循环的障碍。
1木质素微生物降解酶系及酶活性调控自然界参与降解木质素的微生物的种类有真菌、放线菌和细菌。
其中,真菌能把木质素彻底降解为CO和水。
2降解木质素的真菌主要分为三类:白腐菌、褐腐菌和软腐菌。
白腐菌在木质素的生物降解中占有十分重要的地位。
白腐菌多数是担子菌(Basidiomycetes[2],少数为子囊菌。
黄孢原毛平革菌是研究最多的木质素降解菌。
Tien[3]和Glenn 两个研究小组几乎同时发现木质素被降解的关键是黄孢原毛平革菌(Phanerochaete chrysosporium)产生的胞外木质素过氧化物酶系的作用。
该酶系包括木质素过氧化物酶(Lignin peroxidase,简称Lip)、锰过氧化物酶(Mn-dependent peroxidase ,简称MnP),除此之外,还有虫漆酶、HRP、CDH 等酶类[4]。
云芝(Corilus versicolor)是一种非常重要的白腐菌,对木质素的降解能力较强。
能分解木质素的微生物03

白腐菌在木质素的生物降解中占有十分重要的地位。白腐菌多数是担子菌(Basidiomycetes[2],少数为子囊菌。黄孢原毛平革菌是研究最多的木质素降解菌。Tien[3]和Glenn 两个研究小组几乎同时发现木质素被降解的关键是黄孢原毛平革菌(Phanerochaete chrysosporium)产生的胞外木质素过氧化物酶系的作用。该酶系包括木质素过氧化物酶(Lignin peroxidase,简称Lip)、锰过氧化物酶(Mn-dependent peroxidase ,简称MnP),除此之外,还有虫漆酶、HRP、CDH 等酶类[4]。云芝(Corilus versicolor)是一种非常重要的白腐菌,对木质素的降解能力较强。丁少军等 研究了云芝漆酶的培养和分离纯化研究,发现在云芝的木质素降解过程中,漆酶活力较高而木素过氧化物酶、锰过氧化物酶活力较低,它们对木质素的降解率比黄孢原毛平革菌提高近一倍,并认为漆酶在云芝的木质素降解过程中起非常重要的作用。林鹿等[6]研究了白腐菌木云芝和黄孢原毛平革菌对制浆黑液中硫酸盐木质素的降解作用和影响因素。他们发现分子量在1 500~3 000 kD 之间的硫酸盐木质素降解最为显著;两种白腐菌的降解能力不同,培养10 天后,木云芝Lu-11 的降解率达74.5%,降解产物只有一种主要组分,而黄孢原毛平革菌的降解率为65.6%,降解产物有两种,培养条件如:碳源、氮源、pH 值、温度对白腐菌降解硫酸盐木质素的作用有明显影响;在白腐真菌胞内中,除含有利用H2O2 的过氧化物酶系外,还分泌利用O2 的多酚氧化酶系,主要为漆酶(laccase),该酶需要依赖氧和助剂的存在才能有明显的脱甲氧基和脱木质素作用。Andrzej[7]综述了漆酶在木质素降解的作用与活性,认为虫漆酶在木质素降解中起非常关键的作用。付时雨等[8,9] 用漆酶/助剂通入氧气处理苇浆残余木素,通过甲氧基的测定、元素分析、13C-NMR 、IR 分析苇浆残余木素与漆酶/助剂反应前后结构的变化,研究发现苇浆残余木素在反应后甲氧基、酚羟基减少,醇羟基增多,木素结构中羧基增多,木素的苯环结构发生了开环反应。除上述酶类外,白腐菌还产生胞内H2O2 产生酶,因胞内还有过氧化物水解酶,因此胞内不会积累;纤维二糖脱氢酶和纤维二糖/醌还原酶系(CBQ)也是白腐菌产生的重要的木素降解酶[10]。黄峰等[11,12] 报道用裂褶菌(Schizophllum commune)所产的纤维二糖脱氢酶(CDH)加入在黄胞原毛平革菌锰过氧化物酶解硫酸盐浆木质素,结果发现木质素溶出物质明显增加;木质素中甲氧基、羧基、酚羟基和总羟基含量减少,CDH 促进了锰过氧化物酶(MnP)的降解作用。表明CDH 不仅能促进纤维素的降解,而且也是木质素降解体系中的重要组成部分。纤维二糖/醌还原酶(CBQ) 是1974 年由Westermark 和Erikssion 发现的。木质素在氧化酶作用下转变为芳环自由基和醌类,CBQ 能防止活性中间体再聚合,将醌还原为氢醌,CBQ 在纤维素和木素降解中起到了协同作用,即纤维素降解产物为木质素的降解提供还原力[13]。谢君等[14]利用白腐侧耳菌(P. sp2)和粗毛栓菌(T. gallica )在含有麦草的液体培养基中生产木质纤维素降解酶的研究,发现侧耳sp2 和粗毛栓菌在液体培养基中产木质纤维素酶类的能力强、产酶较快,且首先降解木质素,侧耳菌和粗毛栓菌比较,粗毛栓菌(T. gallica )具有更强的分解麦草木质素的能力,60 天后麦草中70.14%的木质素被降解。
木材的生物降解与防腐

生物降解与防腐的平衡点
生物降解:木材的自然 分解过程,受环境因素
影响
防腐处理:防止木材腐 烂、虫蛀等,延长使用
寿命
平衡点:在保证木材 正常使用的同时,尽 量减少对环境的影响
解决方案:选择环保的 防腐剂,合理使用防腐 技术,确保木材的生物
降解与防腐达到平衡
木材的生物降解 与防腐的研究进 展
国内外研究现状
防腐剂的添加方式和浓度也会 影响防腐效果
木材防腐的方法
化学防腐:使用化学药剂处理木材,使其具有防腐性能 物理防腐:通过改变木材的物理结构,提高其防腐性能 生物防腐:利用生物来防治木材的腐朽和虫害 综合防腐:结合多种方法进行木材防腐,提高防腐效果
木材防腐的应用
建筑行业:木材防腐剂用于建筑木材,如地板、门窗、屋顶等,以延长使用寿命和防 止腐烂。
国外研究进展:主要关注木材生物降解和防腐剂的开发和应用
国内研究进展:注重木材生物降解和防腐剂的研发,以及木材保护技术的研究
研究热点:生物降解、防腐剂、木材保护技术 研究挑战:如何提高木材生物降解和防腐性能,降低环境污染
研究热点与难点
生物降解:研究木材生物降解的机理和 影响因素,以开发高效、环保的生物降 解技术。
的侵害。
木材防腐方法:包括涂刷、 浸泡、压力处理等,可以 根据具体情况选择合适的
方法。
木材防腐效果:可以有效 延长木材使用寿命,防止 木材腐烂、变形、变色等。
木材防腐的原理
木材中的水分和营养物质是微 生物生长的必要条件
防腐剂可以杀死或抑制微生物 的生长,防止木材腐烂
防腐剂的种类包括:水溶性防 腐剂、油溶性防腐剂、气溶性 防腐剂等
防腐处理可以降低木材的维护 成本和更换频率
木质素的生物降解及其应用
第20卷第6期2010年12月皮革科学与工程LEATHER SCIENCE AND ENGINEERINGVol.20,No.6Dec.2010文章编号:1004-7964(2010)06-0027-05木质素的生物降解及其应用李海涛,姚开*,何强,贾冬英(四川大学轻纺与食品学院,四川成都610065)收稿日期:2010-04-18基金项目:国家公益性农业科研专项基金(200803033-A004C )第一作者简介:李海涛(1985-),男,河南永城人,硕士研究生,研究方向为食物资源化学,E-mail :lihaitaoyong@ 。
通讯联系人:姚开,教授,E-mail :yaokai555@ 。
摘要:综述了具有降解木质素能力的微生物和酶的种类及其降解特性,阐述了木质素生物降解在生物化学制浆、纸浆生物漂白、造纸废水生物处理、饲料工业、生物肥料等领域的应用现状,展望了木质素生物降解技术研究的发展趋势。
关键词:木质素;生物降解;微生物;酶;应用中图分类号:Q 948.12文献标识码:ABiodegradation and Applications of LigninLI Hai-tao ,YAO Kai *,HE Qiang ,JIA Dong-ying(College of Light Industry and Food Engineering ,Sichuan University ,Chengdu 610065,China )Abstract :In this paper ,the types and the degradation characteristics of ligninolytic microorganisms and lignin-degrading enzymes are introduced.In addition ,the practical applications of lignin biodegradation in biochemical pulping ,biological bleaching of pulp ,biological treatment of papermaking wastewater ,feed industry and bio-fertilizer are summarized.And tech-nology trends in lignin biodegradation are outlook.Key words :lignin ;biodegradation ;microbes ;enzymes ;applications木质素资源十分丰富,是植物光合作用制造的总量仅次于纤维素的有机化合物,估计全球的木质素年产量可达1500亿t 。
《木质素降解研究》PPT课件

2021/4/25
4
1 简介
1.1基本概念 1.2单体与结构 1.3发展前景
2021/4/25
5
木质素(lignin)是由四种醇单体(对香豆醇、松柏醇、5-羟基 松柏醇、芥子醇)形成的一种复杂酚类聚合物。木质素是构成植 物细胞壁的成分之一,具有使细胞相连的作用。在植物组织中具 有增强细胞壁及黏合纤维的作用。其组成与性质比较复杂,并具 有极强的活性。不能被动物所消化,在土壤中能转化成腐殖质。 如果简单定义木质素的话,可以认为木质素是对羟基肉桂醇类的 酶脱氢聚合物。它含有一定量的甲氧基,并有某些特性反应。木 质素是由聚合的芳香醇构成的一类物质,存在于木质组织中,主 要作用是通过形成交织网来硬化细胞壁。木质素主要位于纤维素 纤维之间,起抗压作用。在木本植物中,木质素占25%,是世界上 第二位最丰富的有机物(纤维素是第一位)。
2021/4/25
9
3 木质素降解微生物的种类
3.1三种重要木质素降解酶 I. 木质素过氧化物酶(LiP II. 锰过氧化物酶(MnP III .漆酶
3.2 三种木质素降解酶协同作用
2021/4/25
10
木质素是造纸工业排放黑液COD 和色度形成的主要原因, 其结构 是由甲氧基取代的对- 羟基肉桂酸聚 合而成的异质多晶三维多聚体, 分子间多为稳定的醚键、C- C 键, 是目前公认的微生物难降解芳香 化合物之一。自1934 年Boruff 和Buswell 首次发现能降解木质素的 微生物种群, 人们对木质素的生物降解进行了大量研究, 1983 年和 1984 年发现了木质素过氧化物酶(LiP) 和锰过氧化物酶( MnP) , 由日 本吉田首次在生漆中发现的漆酶( Laccase) , 也始终引起着人们的关 注。这三种酶被公认为是木质素重要的降解酶。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
木质素的微生物降解摘要:综述了木质素微生物降解酶系及酶活性调控措施,介绍了木质素降解机制及木质素微生物降解的应用。
关键字:木质素微生物降解木素降解酶系木质素是植物的主要成分之一,占植物细胞化学组成的15%~30%。
在植物体中,木质素是包裹在纤维素的外面,功能之一就是保护植物细胞不受外界微生物的侵蚀。
从生物合成过程研究得知,木质素首先是由葡萄糖发生芳环化反应形成莽草酸(Shikirnic acid),然后由莽草酸合成三种具有苯丙烷结构的基本单元,分别为:对香豆醇、松柏醇、芥子醇。
从化学结构看,针叶树的木质素主要由松柏醇的脱氢聚合物构成愈创木基木质素;阔叶树的木质素由松柏醇和芥子醇的脱氢聚合物构成愈创木基—紫丁香基木质素;草本植物则由松柏醇、芥子醇和对香豆醇的脱氢聚合物和对香豆酸组成[1]。
木质素是结构复杂、稳定、多样的无定形三维体型大分子,在自然界中,降解缓慢,成为地球生物圈中碳循环的障碍。
1木质素微生物降解酶系及酶活性调控自然界参与降解木质素的微生物的种类有真菌、放线菌和细菌。
其中,真菌能把木质素彻底降解为CO和水。
2降解木质素的真菌主要分为三类:白腐菌、褐腐菌和软腐菌。
白腐菌在木质素的生物降解中占有十分重要的地位。
白腐菌多数是担子菌(Basidiomycetes[2],少数为子囊菌。
黄孢原毛平革菌是研究最多的木质素降解菌。
Tien[3]和Glenn 两个研究小组几乎同时发现木质素被降解的关键是黄孢原毛平革菌(Phanerochaete chrysosporium)产生的胞外木质素过氧化物酶系的作用。
该酶系包括木质素过氧化物酶(Lignin peroxidase,简称Lip)、锰过氧化物酶(Mn-dependent peroxidase ,简称MnP),除此之外,还有虫漆酶、HRP、CDH 等酶类[4]。
云芝(Corilus versicolor)是一种非常重要的白腐菌,对木质素的降解能力较强。
丁少军等[5]研究了云芝漆酶的培养和分离纯化研究,发现在云芝的木质素降解过程中,漆酶活力较高而木素过氧化物酶、锰过氧化物酶活力较低,它们对木质素的降解率比黄孢原毛平革菌提高近一倍,并认为漆酶在云芝的木质素降解过程中起非常重要的作用。
林鹿等[6]研究了白腐菌木云芝和黄孢原毛平革菌对制浆黑液中硫酸盐木质素的降解作用和影响因素。
他们发现分子量在1 500~3 000 kD 之间的硫酸盐木质素降解最为显著;两种白腐菌的降解能力不同,培养10 天后,木云芝Lu-11 的降解率达74.5%,降解产物只有一种主要组分,而黄孢原毛平革菌的降解率为65.6%,降解产物有两种,培养条件如:碳源、氮源、pH 值、温度对白腐菌降解硫酸盐木质素的作用有明显影响;在白腐真菌胞内中,除含有利用H2O2的过氧化物酶系外,还分泌利用O2的多酚氧化酶系,主要为漆酶(laccase),该酶需要依赖氧和助剂的存在才能有明显的脱甲氧基和脱木质素作用。
Andrzej[7]综述了漆酶在木质素降解的作用与活性,认为虫漆酶在木质素降解中起非常关键的作用。
付时雨等[8,9]用漆酶/助剂通入氧气处理苇浆残余木素,通过甲氧基的测定、元素分析、13C-NMR 、IR 分析苇浆残余木素与漆酶/助剂反应前后结构的变化,研究发现苇浆残余木素在反应后甲氧基、酚羟基减少,醇羟基增多,木素结构中羧基增多,木素的苯环结构发生了开环反应。
除上述酶类外,白腐菌还产生胞内H2O2产生酶,因胞内还有过氧化物水解酶,因此胞内不会积累;纤维二糖脱氢酶和纤维二糖/醌还原酶系(CBQ)也是白腐菌产生的重要的木素降解酶[10]。
黄峰等[11,12]报道用裂褶菌(Schizophllum commune)所产的纤维二糖脱氢酶(CDH)加入在黄胞原毛平革菌锰过氧化物酶解硫酸盐浆木质素,结果发现木质素溶出物质明显增加;木质素中甲氧基、羧基、酚羟基和总羟基含量减少,CDH 促进了锰过氧化物酶(MnP)的降解作用。
表明CDH 不仅能促进纤维素的降解,而且也是木质素降解体系中的重要组成部分。
纤维二糖/醌还原酶(CBQ)是1974 年由Westermark和Erikssion 发现的。
木质素在氧化酶作用下转变为芳环自由基和醌类,CBQ 能防止活性中间体再聚合,将醌还原为氢醌,CBQ 在纤维素和木素降解中起到了协同作用,即纤维素降解产物为木质素的降解提供还原力[13]。
谢君等[14]利用白腐侧耳菌(P. sp2)和粗毛栓菌(T. gallica )在含有麦草的液体培养基中生产木质纤维素降解酶的研究,发现侧耳sp2 和粗毛栓菌在液体培养基中产木质纤维素酶类的能力强、产酶较快,且首先降解木质素,侧耳菌和粗毛栓菌比较,粗毛栓菌(T. gallica )具有更强的分解麦草木质素的能力,60 天后麦草中70.14%的木质素被降解。
不同的生理条件对微生物降解木质素的重要影响成为木质素微生物降解研究的一个热点问题[15,16],王宜磊[17]研究了碳源与氮源对彩绒革盖菌Coridus versicolor 木质素酶分泌的影响,发现淀粉含量丰富的物质作碳源有利于木质素降解酶的分泌,而尿素对木质素降解酶的分泌有抑制作用,具体原因有待于进一步研究;王佳玲等[18,19]研究了氮源、碳源及Mn2+浓度对Panus conchatus 分泌木质素降解酶的影响,认为高氮条件可能是这种类型的白腐菌生产木素降解酶的重要条件,同时发现紫丁香醛和香豆酸对其锰过氧化物酶和漆酶分别起显著的诱导作用,碳源的种类对MnP 和漆酶(laccase)产生的影响有明显的差异,Mn2+对P. conchatus MnP 和laccase 的产生有明显的调节作用:不加Mn2+条件下,P. conchatus 几乎不分泌MnP,加入Mn2+并提高Mn2+浓度,MnP 活力随之提高,而laccase 活力却相应降低。
余惠生等[20]研究了铜(Ⅱ)离子浓度的变化对白腐菌Panus conchatus 产漆酶的影响,结果表明漆酶的产生显著受Cu2+的调控,没有铜离子存在,漆酶的活性很低,增加Cu2+浓度,漆酶的活性增大,但过多的Cu2+又会对漆酶产生抑制作用,使酶活降低。
2 木质素生物降解机制[21](1)木质素模型化合物的Ca-Cb断裂现已基本确定,通过单电子转移机制,LiP 催化氧化b-1 非酚型木质素模型化合物为其芳香正离子自由基,经Ca-Cb 断裂形成3,4-二甲氧基苯乙醇自由基和质子化形式的藜芦醛。
有氧存在时,前者加氧后再释放超氧离子形式羰基,或形成醇,无氧存在时则溶剂水参与而形成醇。
付时雨等[22]利用色—质联用技术分析了稻草木质素经白腐菌Panusconchatus 生物作用后木质素降解产物中低分子酸性组分的主要变化及其结构,通过质谱对其主要11 种化合物结构进行分析,证实木质素生物降解过程发生木质素侧链Ca-Cb链氧化断裂。
(2)Ca-氧化机制 LiP 催化b-O-4 木质素模型化合物的主要反应是Ca-Cb 断裂形成的藜芦醛和2-甲氧基苯酚,后者在反应条件下易于聚合,同时还有相当一部分形成Ca氧化产物。
这种氧化产物是正离子自由基中间体失去质子或直接失去氢形成的。
在活性氧存在时,后者更容易发生。
(3)芳环取代机制在催化反应中形成的芳香正离子自由基与溶剂水或其他亲和试剂作用,随底物的不同而发生不同类型的反应,例如,当苯环上的取代基是甲氧基时,就发生脱甲氧基反应,氧化形成相应的醌。
(4)氧的活化在木质素模型化合物氧化中一种普遍现象是分子氧与羟基取代的苯自由基间的反应中能引起氧的活化,氧被还原成超氧离子,又与氢质子反应生成H2O2与O2,这个过程是发生在分子氧与苯基中间体之间的纯化学过程。
这个反应也出现在木质素过氧化物酶的催化循环中,其结果是O2成为最终的电子受体。
(5)藜芦醇及其衍生物的氧化藜芦醇是黄孢原毛平革菌的一个次级代谢产物,又被木质素过氧化物酶所分解,分解产物主要是藜芦醛,其次是开环产物和醌,产物的分布受pH 值控制。
(6)芳香环开裂利用标记底物和反应物的方法对许多模型化合物进行芳香环的开裂反应研究表明,藜芦醇开环反应的机制也适用于木质素单体、二聚体和低聚体模型化合物,在大多数请况下都得到了多种产物的混合物。
(7)单甲氧基芳香物的氧化对于3,4-二甲氧基芳香环,非但可被Phanerochaete chrysosporium 代谢,而且可被LiP 氧化,单甲氧基芳香环虽然可被Phanerochaete chrysosporium 代谢,却不被LiP 氧化,然而加入少量的藜芦醇或其他的二甲氧基芳香物,则能大大提高单甲氧基芳香物的反应速度。
(8)醌/氢醌的形成漆酶是一种含铜的酚氧化酶,它能催化酚型二聚体模型物b-1 和b-O-4 结构经过Ca-芳烃断裂,产生甲氧基取代的醌/氢醌。
辣根过氧化物酶(HRP)也能得到类似的结果。
单体木质素模型物如香草酸、香草醛、香草醇等也能被氧化成甲氧基取代的醌/氢醌。
余惠生等[23]用定量13C-NMR 技术研究了稻草经白腐菌Panus conchatus 降解后木质素的高分子部分的结构特征变化,发现发酵稻草木质素的13C-NMR 图谱出现新的吸收峰以及某些吸收峰的强度有明显变化,这些变化证实了在生物作用过程中木质素大分子中形成了醌类。
(9)漆酶催化木质素氧化的机制漆酶以O2作电子受体催化多酚化物经四次单电子传递形成醌及自由基的含铜的酚氧化酶,而且不需要H2O2参与,对于木质素而言,其反应包括脱甲氧基、脱羟基、C-C 键断裂过程等。
3 木质素微生物降解的意义木质素是自然界可再生的含量丰富的天然高分子聚合物,也是化学制浆工业中重要的废弃物和污染物。
(1)应用于污染治理与环境保护传统的化学制浆会带来大量的工业废水,污染环境,将木质素的生物降解应用于生物制浆及造纸废水的处理,不仅可以极大地改善造纸工业的环境污染问题,还有利于减少能耗创造经济价值。
Gary 利用木素过氧化物酶处理2,4二溴苯酚等工业废水,在氧化的过程中伴随Ward[24]二聚体、三聚体和四聚体的形成,消除了废水的毒性。
我国的纺织业和印染业非常发达,印染废水的污染成为水污染的一个重要来源,陈叶福等[25]综述了木质素生物降解与纸浆工业废水脱色,认为白腐真菌能降解木质素和相关化合物,有可能成为传统处理方法的替代方法。
据文献报道,黄孢原毛平革菌在产酶条件下可以降解偶氮染料、蒽醌染料以及三苯甲烷等多种类型的染料。
张朝晖等[26,27]研究了黄孢原毛平革菌所产木质素过氧化物酶系对染料降解的研究,建立了木质素过氧化物酶降解偶氮染料卡布龙红的动力及过程模型。
(2)应用于造纸工业木素酶的生物漂白技术是通过生物木素酶直接作用于纸浆的残余木质素,并使之发生降解溶出,从而有利于纸浆的进一步漂白。