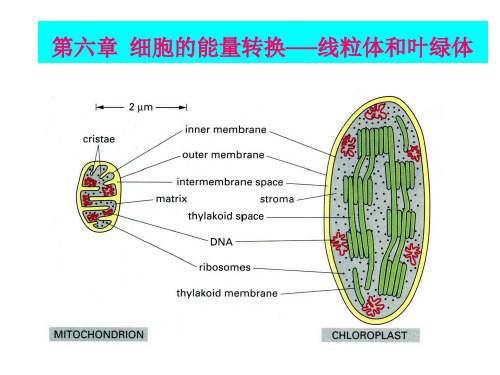
第六章 细胞的能量转换——线粒体和叶绿体
第六章细胞的能量转换

图5-4 化学渗透学说
ATP合酶的结构和作用机理 ATP合酶的结构和作用机理
ATP合酶的结构 ATP合酶的结构 合酶的
分为球形的F 头部)和嵌入膜中的F 基部) 分为球形的 1(头部)和嵌入膜中的 0(基部) F1由5种多肽组成 3β3γδε复合体,具有三个 种多肽组成α 复合体, 种多肽组成 复合体 具有三个ATP合成的催化位点 合成的催化位点 F0由三种多肽组成ab2c12复合体,嵌入内膜 。 复合体, F0由三种多肽组成ab 由三种多肽组成
第二节
叶绿体与光合作用
光合作用就是叶绿体利用光能同化二氧化碳和 水合成糖,同时产生氧的过程 的过程。 水合成糖,同时产生氧的过程。
一、形态与结构 二、光反应 三、暗反应
一、形态与结构
叶绿体形似凸透镜,由叶绿体外被、类囊体和基质三 叶绿体形似凸透镜,由叶绿体外被、类囊体和基质三部分组成 外被:由双层单位膜组成。 外被:由双层单位膜组成。外膜通透性大 ,内膜对物质有 很强的选择性。 很强的选择性。 类囊体:是单层膜围成的扁平小囊。 类囊体:是单层膜围成的扁平小囊。 许多类囊体象圆盘一样叠在一起,称为基粒 基粒。 许多类囊体象圆盘一样叠在一起,称为基粒。 组成基粒的类囊体,叫做基粒类囊体 基粒类囊体。 组成基粒的类囊体,叫做基粒类囊体。 贯穿在两个或两个以上基粒之间的没有发生垛叠的 类囊体称为基质类囊体 基质类囊体。 类囊体称为基质类囊体。 是内膜与类囊体之间的空间,主要成分叶绿体DNA DNA、 基质 :是内膜与类囊体之间的空间,主要成分叶绿体DNA、 蛋白质合成体系, 蛋白质合成体系,颗粒成分等 。
形态与分布
形态: 形态:线粒体一般呈粒状或杆状 大小:一般直径 大小:一般直径0.5~1µm,长1.5~3.0µm , 数量: 数量:数目一般数百到数千个 分布:分布在细胞功能旺盛的区域 分布:
南开大学细胞生物学-第六章 细胞的能量转换

底物水平的磷酸化
由相关的酶将底物分子上的磷酸基 团转移到ADP分子上,生成ATP。
1. ATP合成酶的结构与组成
2. 能量耦联与ATP合成酶的作用机制
◆解离与重建实验证明电子传递与ATP合成是由 两个不同的结构体系执行
◆化学渗透假说 ◆ATP合成机制
该实验证明了:
1. 颗粒是ATP合成必需的 2. 小泡是电子传递必需的 3. 电子传递和ATP合成是Ⅳ
在电子传递过程中,有几点需要说明
◆ 五种类型电子载体:黄素蛋白、细胞色素(含血红素辅基)、Fe-S中心、
铜原子、辅酶Q。前四种与蛋白质结合,辅酶Q为脂溶性醌。 ◆ 电子传递起始于NADH脱氢酶催化NADH氧化,形成高能电子(能量
转化), 终止于O2形成水。 ◆ 电子传递方向按氧化还原电势递增的方向传递(NAD+/NAD最低,
△P= △Ψ-2.3 —R△TpH F
(四)ATP形成机制——氧化磷酸化
将ADP转变为ATP的过程称为磷酸化(phosphorylation)。 底物水平的磷酸化 氧化磷酸化(oxidative phosphorylation) 光合磷酸化(photophosphorylation)
1. ATP合成酶的结构与组成 2. 能量耦联与ATP合成酶的作用机制
在胞质中的定位与迁移与微管有关。
线粒体的结构
膜间隙
嵴
外膜
内膜 基质
外膜(outer membrane)
厚约6nm 蛋白质、脂质各占50% 含孔蛋白(porin ),由β折叠链 形成桶状结构,中心为2~3nm的 小孔。可以可逆的开闭,完全打 开时,可通过分子量5×103的分 子。 外膜的通透性非常高 标志酶为单胺氧化酶
细胞的能量转换——线粒体和叶绿体

第六章细胞的能量转换——线粒体和叶绿体一、填空题1.能对线粒体进行专一染色的活性染料是____。
2.线粒体膜围腔有2个,它们是____和____,而叶绿体膜围腔有3个,自外向里分别是____,____和____。
3.线粒体各部分结构中有各自特殊的标记酶,它们分别在外膜是____,外腔是____,内膜是____,内腔是____。
4.三羧酸循环是在线粒体____中进行,氧化磷酸化过程是在____上,由____和____完成。
5.根据接受代谢物上脱下的氢的原初受体的不同,可以将细胞中的呼吸链分为两种典型的类型,分别为____和____。
6.ATP合成酶合成ATP的直接能量来自____。
7.在线粒体电子传递链的4种复合物中既是电子传递体又是质子位移体的是____、____和____。
8.在线粒体电子传递链中电子传递方向按氧化还原电势____的方向传递。
9.叶绿体在显微结构上主要分为____、____、____。
10.叶绿体的超微结构可分为____、____、____、____、____几部分。
11.叶绿体基质中的类囊体可以根据形态不同分为____和____。
12.氧是在植物细胞中____部位上所进行的____的过程中产生的。
13.叶绿体类囊体膜上色素分子按照其作用可以分为两大类,分别为____和____。
14.捕光色素和反应中心构成了____,它是进行光合作用的最小结构单位。
15.光合作用按照是否需要光可分为____和____两步,其中光反应又可分为____和____两步。
光反应发生在____,暗反应发生在____。
16.电子沿光合电子传递链传递时,根据最终电子受体的不同,光合磷酸化可分为____和____两条通路。
17.植物细胞中具有特异的质体细胞器,主要分为____、____、____。
18.线粒体的增殖是由____而来。
19.叶绿体的发育是由____分化而来。
20.线粒体和叶绿体的生长和增殖是受____两套遗传系统控制,所以称为半自主性细胞器。
细胞的能量转换--线粒体和叶绿体

第六章细胞的能量转换--线粒体和叶绿体线粒体和叶绿体是细胞内两个能量转换细胞器,它们能高效地将能量转换成ATP。
线粒体广泛存在于各尖真核细胞,而叶绿体仅存在于植物细胞中。
它们的形态结构都呈封闭的双层结构,内膜都演化为极其扩增的特化结构,并在能量转换中起主要作用。
线粒体和叶绿体以类似的方式合成ATP。
线粒体和叶绿体都是半自主性细胞器。
第一节线粒体与氧化磷酸化一、线粒体的形态结构(一)线粒体的形态与分布线粒体多呈线状和粒状,是由两层单位膜套叠而成的封闭囊状结构,由外膜、内膜、膜间隙及基质4部分构成。
(二)线粒体的结构与化学组成线粒体的化学成分主要是蛋白质和脂类。
线粒体的蛋白可分为可溶性与不溶性两类,可溶性蛋白质大多数是基质中的酶和膜外周蛋白;不溶性蛋白是膜的镶嵌蛋白、结构蛋白和部分酶蛋白。
线粒体脂类主要成分是磷脂。
蛋白质(线粒体干重的65~70%)脂类(线粒体干重的25~30%):磷脂占3/4以上,外膜主要是卵磷脂,内膜主要是心磷脂,线粒体脂类和蛋白质的比值:0.3:1(内膜);1:1(外膜)线粒体内外膜在化学组成上根本不同是脂类和蛋白质的比值不同。
内膜的脂类与蛋白质的比值低,外膜中比值较高。
1 外膜外膜:厚约6nm,含孔蛋白(porin),通透性高。
2 内膜厚约6-8nm,通透性低,只有不带电荷的小分子才能通过。
内膜向内褶叠形成嵴。
内膜和嵴的基质面上有许多排列规则的基粒,基粒由头部和基部组成,头部又叫F1,基部又叫F0。
3 膜间隙内外膜之间宽8nm的空隙,它延伸到嵴的轴心部(嵴内隙),内含许多可溶性酶类、底物和辅助因子。
4 线粒体基质内膜和嵴包围的空间,内含蛋白质性质的胶状物质。
基质中有催化三羧酸循环、脂肪酸 -氧化、氨基酸氧化、蛋白质合成等有关的酶类和其它成分,如环状DNA、RNA、核糖体及较大的致密颗粒,其作用主要是贮存Ca+。
线粒体酶的定位线粒体约有140种酶,分布在各个结构组分中,有的可作为某一部位所特有的标志酶,如外膜的单胺氧化酶,膜间隙的腺苷酸激酶,内膜的细胞色素氧化酶,基质中的苹果酸脱氢酶。
第六章 线粒体和叶绿体
1) 内膜结构
单层膜,比外膜厚; 通透性较外膜小; 向内折叠成嵴,表面积极大增加,代谢效率 极大提高;
嵴的形状与数量和细胞种类与生理状况有关
2) 线粒体分类(根据嵴的形态结构)
(a)板层形 (b)管形 (a),(b)为基本形 (c)羽冠形 (d)网膜形 (e)绒毛形 (f)平行形 (g)同心圆形
返回
第二节 叶绿体与光合作用
●一.叶绿体(Chloroplast)的形态结构
二.叶绿体的结构与化学组成
●三.叶绿体的功能—光合作用
(photosynthesis)
一、叶绿体(Chloroplast)的形态结构
●叶绿体与线粒体形态结构比较
叶绿体内膜并不向内折叠成嵴;内膜不含电
子传递链;除了膜间隙、基质外,还有类囊体; 捕光系统、电子传递链和ATP合成酶都位于类囊体 膜上。 ●叶绿体超微结构
1964年由Paul Boyer最先提出来的。此学
说认为电子沿电子传递链传递使线粒体 内膜蛋白质组分发生了构象变化,形成 一种高能形式。这种高能形式通过ATP 的合成而恢复其原来的构象。未能找到 有力的证据。
(3)化学渗透学说 1961 由英国生物化学家Peter Mitchell最先提出来
的。此学说认为电子传递释放出的自由能和ATP合成
+
个H 泵出内膜。
c 内膜对H+不能自由通过,泵出
膜外侧的H+不能自由返回膜内侧,
因而造成H+浓度的跨膜浓度梯度。
此梯度包含的能量可驱使ADP和Pi 生成ATP。
d.氢通过ATP合成酶上 的特殊的途径,返回基 质。H+浓度梯度所释放
的自由能,偶联ADP和
第六章 细胞的能量转换—线粒体和叶绿体 南开大学细胞生物学课件

1. 电子载体
电子传递链是由一系列特殊的电子载体构成的。 在电子传递过程中,与释放的电子结合并将电子传递 下去的化合物称为电子载体(electron carrier)。
电子载体有五种: 黄素蛋白(flavoprotein) 细胞色素(cytochrome) 泛醌(ubiquinone, UQ) 铁硫蛋白(iron-sulfur protein) 铜原子(copper atom)
Fis1和Mdv1 2. 参与作用
endophilin B1 Mff(mitochondrial fission factor) GDAP1(ganglioside-induced
fifferentiation associated protein 1)
2. 线粒体融合与分裂的细胞生物学基础
二、线粒体的超微结构
复合物Ⅱ
复合物Ⅲ
复合Ⅳ
在电子传递过程中,有几点需要说明
◆ 五种类型电子载体:黄素蛋白、细胞色素(含血红素辅基)、 Fe-S中心、铜原子、辅酶Q。前四种与蛋白质结合,辅 酶Q为脂溶性醌。
◆ 电子传递起始于NADH脱氢酶催化NADH氧化,形成高 能电子(能量转化), 终止于O2形成水。
◆ 电子传递方向按氧化还原电势递增的方向传递 (NAD+/NAD最低,H2O/O2最高)
叶肉细胞
维管束 维管束鞘细胞
第三节 线粒体和叶绿体的半自主性及其起源
线粒体和叶绿体的功能主要受细胞核 基因组调控,但同时又受到自身基因组的 调控,被称为半自主性细胞器。
线粒体和叶绿体以非孟德尔方式遗传。
一、线粒体和叶绿体的半自主性 二、线粒体和叶绿体的起源
一、线粒体和叶绿体的半自主性
(一)线粒体和叶绿体DNA (二)线粒体和叶绿体中的蛋白质 (三)线粒体和叶绿体基因组与细胞核
6-线粒体和叶绿体PPT课件

-
44
-
45
2)类囊体膜的化学组成
主要成分:蛋白质和脂质(比例约60:40)。 • 1.脂质
主要是磷脂和糖脂及色素、醌化合物等 不饱和的亚麻酸约87%,流动性大 • 2.蛋白质 (1)外在蛋白:CF1 、与光反应有关的酶 (2)内在蛋白:20余种多肽
-
46
3.叶绿体基质(stroma)
• 1)核酮糖-1,5-二磷酸羧化酶RuBPase • 2)环状DNA • 3)RNA:rRNA、tRNA、mRNA • 4)核糖体(蛋白质合成体系) • 5)脂滴(Lipiddroplet)或称嗜锇滴 • 6)植物铁蛋白、淀粉粒等
形成“转子”; 2)嵌入膜中的F0(基
部),组成“定子”;
-
25
F1:3:3:1:1:1
具3个ATP合成的催 化位点
F0: 1a:2b:12c
环形结构,具质 子通道
-
26
❖ F1因子具有ATP酶活性
-
27
b)ATP合成酶性质:是一种具有双向作用的装置
-
28
c)ATP合成酶的作用机制
Step 1: Proton gradient is built up as a result of NADH (produced from oxidation reactions) feeding electrons into electron transport system.
• 3由)1Fa、0偶1联b、因1子0-1:2c是亚内基膜组上成的,疏多水个蛋c亚白基复形合成体多,(形1成2)跨聚膜体质,子成通一道。 环 合状于结c亚构基,环和状转结子构结的合外。侧a,、并b亚通基过及δ亚F1的基δ和亚头基部形相成结“合定。子”,结
第六章:细胞的能量转换-线粒体和叶绿体

铜原子
电子传递链与电子传递
电子载体的排列顺序
复合物1-辅酶q-复合物111-复合物1111 复合物11-辅酶q-复合物111-复合物1111
线粒体的功能
电子转运复合物
复合物1=nadh-辅酶q还原酶=nadh脱氢酶
复合物11=琥珀酸-辅酶q还原酶=琥珀酸脱氢酶
复合物111=辅酶q-细胞色素c还原酶=细胞色素还 原酶=细胞色素bc1复合体=bc1
细胞的能量转换-线粒体 和叶绿体
线粒体与氧化磷酸化
线粒体的形态与分布
线粒体的形态结构
线粒体的结构与化学组成
外膜
单胺氧化酶
内膜
细胞色素氧化酶
膜间隙
腺苷酸激酶
线粒体基质
线粒体中的氧化代谢
黄素蛋白
电子载体
在电子传递过程中,与释放的电子结合并将电子 传递下去的复合物称为电子载体
细胞色素 泛醌=辅酶q 铁硫蛋白
由光照引起的电子传递与磷酸化相偶联而生成 ATP的过程,称为光合磷酸化
卡尔文循环
形成糖类
光合碳同化
c4途径
草酰乙酸
景天酸代谢
线粒体和叶绿体是半自主性细胞器
线粒体和叶绿体的DNA 线粒体和叶绿体的蛋白质合成 线粒体和叶绿体蛋白质的运送与组装
线粒体和叶绿体的增殖与起源
线粒体和叶绿体的增殖 线粒体和叶绿体的起源
复合物1111=细胞色素氧化酶=细胞色素c氧化酶
质子转移与质子驱动力的形成
质子动力势
高能电子释放的能量驱动线粒体内膜复合物将氢 离子从基质侧泵到膜间隙,形成跨线粒体内膜氢 离子梯度及电位差,在内膜两侧形成的电化学质 子梯度称为质子动力势
ATP合酶的结构与组成
f1头部 f0基部
细胞生物学第六章线粒体与叶绿体
ATP合酶的“结合变构模 型
三、线粒体与疾病
1、线粒体病:属母系遗传,如克山病、Leber遗 传性视神经病、肌阵挛性癫痫 2、线粒体与衰老 数目与体积的变化 mtDNA的损伤、缺失 线粒体是细胞内自由基的主要来源 3、线粒体与细胞凋亡
第二节 叶绿体与光合作用
一、叶绿体的形态、大小和数目 不同植物种类之间有很大差异。 一般形态为香蕉形,或者称为透镜形。宽2-4um, 长5-10um。叶肉细胞含50-200个叶绿体。 二、叶绿体的结构和化学组成 (一)叶绿体被膜 双层单位膜,6-8nm,之间为10-20nm的膜间 隙。外膜有孔蛋白,通透性大。
THE ESTABLISHMENT OF A PROTONMOTIVE FORCE
33
(四)ATP形成机制——氧化磷酸化
氧化磷酸化:在呼吸链上与电子传递相耦联的由 ADP被磷酸化生成ATP的酶促过程。 呼吸链上有三个部位是氧化还原放能与ADP磷酸 化生成ATP偶联的部位。这三个部位有较大的自 由能变化,足以使ADP与无机磷结合形成ATP。 部位Ⅰ在NADH至CoQ之间。部位Ⅱ在细胞色素 b和细胞色素c之间。部位Ⅲ在细胞色素a和氧之 间。
第二节 叶绿体与光合作用
内膜通透性低。 内膜上有特殊的转运载体,如磷酸交换载体、二 羧酸交换载体,都属于被动运输。 内膜上有与糖脂、磷脂合成有关的酶类。因此, 叶绿体被膜不仅是叶绿体脂合成的场所,也是整 个植物细胞脂合成的主要场所。
(二)类囊体 1、类囊体的结构 叶绿体基质中由单位膜包围形成的扁平小 囊称为类囊体。 基粒、基粒类囊体、基质类囊体。 一个叶绿体的全部类囊体是一个完整连续 的封闭膜囊。含有丰富的具半乳糖的糖脂。 流动性很大。类囊体膜上的成分有:PSI、 PSII、细胞色素bf、CF0-CF1ATP酶。其分 布不对称。
细胞生物学第六章细胞的能量转换器

3.细胞色素
分子中含有血红素铁(图7-7),以共价形式与蛋白结合,通Fe3+、Fe2+形式变化传递电子,呼吸链中有5类,即:细胞色素a、a3、b、c、c1,其中a、a3含有铜原子。
图7-7 血红素c的结构
5.Q:
4.铁硫蛋白:
在其分子结构中每个铁原子和4个硫原子结合,通过Fe2+、Fe3+互变进行电子传递,有2Fe-2S和4Fe-4S两种类型(图7-8)。 图7-8 铁硫蛋白的结构((引自Lodish等1999) 是脂溶性小分子量的醌类化合物,通过氧化和还原传递电子(图7-9)。有3种氧化还原形式即氧化型醌Q,还原型氢醌(QH2)和介于两者之者的自由基半醌(QH)。
线粒体的超微结构
1.2 线粒体的形态结构
图 7-3 肌细胞和精子的尾部聚集较多的线粒体, 以提供能量 图7-4 线粒体包围着脂肪滴,内有大量要被氧化的脂肪 图7-1线粒体的TEM照片
内膜向线粒体基质褶入形成嵴(cristae),嵴能显著扩大内膜表面积(达5~10倍),嵴有两种类型:①板层状(图7-1)、②管状(图7-3),但多呈板层状。 图7-3 管状嵴线粒体 嵴上覆有基粒(elementary particle),基粒由头部(F1偶联因子)和基部(F0偶联因子)构成,F0嵌入线粒体内膜。 图7-1线粒体的TEM照片
图7-9 辅酶Q
呼吸链的复合物
利用脱氧胆酸(deoxycholate,一种离子型去污剂)处理线粒体内膜、分离出呼吸链的4种复合物,即复合物Ⅰ、Ⅱ、Ⅲ和Ⅳ,辅酶Q和细胞色素C不属于任何一种复合物。辅酶Q溶于内膜、细胞色素C位于线粒体内膜的C侧,属于膜的外周蛋白。
即NADH脱氢酶,哺乳动物的复合物Ⅰ由42条肽链组成,呈L型,含有一个FMN和至少6个铁硫蛋白,分子量接近1MD,以二聚体形式存在,其作用是催化NADH的2个电子传递至辅酶Q,同时将4个质子由线粒体基质(M侧)转移至膜间隙(C侧)。电子传递的方向为:NADH→FMN→Fe-S→Q,总的反应结果为:
细胞生物学[第六章细胞的能量转换—一线粒体和叶绿体]课程预习
![细胞生物学[第六章细胞的能量转换—一线粒体和叶绿体]课程预习](https://img.taocdn.com/s3/m/7b24cc1c14791711cc791745.png)
第六章细胞的能量转换—一线粒体和叶绿体线粒体和叶绿体是细胞内的两种产能细胞器。
它们最初的能量来源有所不同,但却有着相似的基本结构,而且以类似的方式合成ATP。
线粒体是一种高效地将有机物转换为细胞生命活动的直接能源ATP的细胞器。
叶绿体通过光合作用把光能转换为化学能,并储存于糖类、脂肪和蛋白质等大分子有机物中。
线粒体和叶绿体都具有环状DNA及自身转录RNA与翻译蛋白质的体系。
线粒体和叶绿体都是半自主性的细胞器。
一、线粒体和氧化磷酸化线粒体通过氧化磷酸化作用,进行能量转换,为所需要的细胞进行各种生命活动提供能量。
(一)线粒体的形态结构1.线粒体的形态与分布线粒体一般呈粒状或杆状,但因生物种类和生理状态而异,可呈环形、哑铃形、线状、分权状或其他形状。
主要化学成分是蛋白质和脂类,其中蛋白质占线粒体干重的65%~70%,脂类占25%~30%。
一般直径0.5~1μm,长1.5~3.0μm,在胰脏外分泌细胞中可长达10~20μm,称巨线粒体。
数目一般数百到数千个,植物因有叶绿体的缘故,线粒体数目相对较少;肝细胞约1300个线粒体,占细胞体积的20%;单细胞鞭毛藻仅1个,酵母细胞具有一个大型分支的线粒体,巨大变形中达50万个;许多哺乳动物成熟的红细胞中无线粒体。
通常结合在维管上,分布在细胞功能旺盛的区域。
线粒体在细胞质中可以向功能旺盛的区域迁移,微管是其导轨,由马达蛋白提供动力。
2.线粒体的结构与化学组成线粒体的超微结构在电镜下观察到线粒体是由两层单位膜套叠而成的封闭的囊状结构。
主要由外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)、基质(matrix)或内室(inner chamber)四部分组成。
(1)外膜是包围在线粒体最外面的一层单位膜,光滑而有弹性,厚约6μm。
外膜上有排列整齐的筒状圆柱体,其成分为孔蛋白(porin),圆柱体上有小孔。
(2)内膜位于外膜内侧,把膜间隙与基质(内室)分开。
第六章 细胞的能量转换--线粒体和叶绿体

2) ATP合成酶的作用机理
ATP合成酶据认为是通过结合变化和旋转催化的机制来合成ATP的。 F1中的β亚基有三种构型: ① 紧密结合态(T态):可与ADP和Pi的结合很紧,能生成ATP,并能与 ATP牢牢结合。 ② 松散结合态(L态):同ADP和Pi的结合较强。
③ 空置状态(O态): 几乎不与ATP、ADP和Pi结合。
(二)线粒体和叶绿体的起源
1 内共生起源学说 2 非共生起源学说
(二)线粒体的化学组成及酶的定位
外膜主要是卵磷脂, 内膜主要是心磷脂。 线粒体不同部位标志酶: 1 外 膜-标志酶:单胺氧化酶。 2 内 膜-标志酶:细胞色素氧化酶。 3 膜间隙-标志酶:腺苷酸激酶。 4 基 质-标志酶:苹果酸脱氢酶。
(三)氧化磷酸化
1 线粒体的主要功能--氧化磷酸化。 2 有机物氧化、呼吸链(电子传递链)与ATP产生之间的关系。
光能→使叶绿体色素分子激发释放电子→电子传递和光合磷 酸化→产生ATP和NADPH2→通过卡尔文循环CO2和H2O被 ATP和NADPH2还原为糖类物质。 1 光反应-包括原初反应、电子传递和光合磷酸化。 2 暗反应-即碳同化。
1 原初反应
原初反应是指叶绿素分子从被光激发至引起第一个光化学反应为 止的过程。包括光能的吸收、传递与转换,即光能被捕光色素分 子吸收,并传至反应中心,在反应中心发生最初的光化学反应, 使电荷分离从而将光能转换为电能的过程。 (1)捕光色素:包括大部分叶绿素a,叶绿素b,胡萝卜素和叶黄 素等。叶绿素吸收的光主要是蓝色和红色而不是 绿色光。 (2)反应中心色素:由一种特殊状态的叶绿素a分子组成,可分为 P700(PⅠ的中心色素)和P680(PⅡ的中心色素). (3)反应中心:由一个中心色素分子和一个原初电子供体D及一个 原初电子受体A组成。 (4)光合作用单位:由捕光色素和反应中心构成。
细胞生物学 名词解释 第六章 线粒体和叶绿体

光合磷酸化数据
2H2O光解生成1分子O2和4H+,PSI和PSII分别吸收4个光子(共8个光子)传递4e-,造成12H+(水光解产生4H+,PQ释放4H+,PSI消耗基质中4H+)=3ATP,生成2NADPH
电子传递链
electron transport chain
膜上一系列由电子载体组成的电子传递途径。这些电子载体接受高能电子,并在传递过程中逐步降低电子的能量,最终将释放的能量用于合成ATP或以其他能量形式储存。
捕光复合体Ⅱ
light harvesting complex Ⅱ,LHCⅡ
位于光系统Ⅰ之外的色素蛋白复合,含有大量天线色素为光系统Ⅱ(PSⅡ)收集光子。
ATP合酶
ATP synthase
位于线粒体内膜或叶绿体的类囊体膜上,通过氧化磷酸化或光合磷酸化催化ADP和无机磷合成ATP的酶,由F1头部和嵌入膜内的F0基部组成,也常见于细菌膜上。
F1头部组成
α3β3γεδ,α3β3形成橘瓣,β亚基的结合位点具有催化ATP合成或水解的活性,γε形成转子,调节3个β亚基催化位点的开放和关闭;δ亚基为F1和Fo相连接所必须的。
电子从PS I开始经过Fd后传给Cytb6f,再经PC回到PS I,由PSI单独完成,电子传递呈现闭合的回路,只产生ATP,不产生NADPH和O2。
作用:当植物缺乏NADP+时,启动循环光合磷酸化,以调节ATP与NADPH的比例,适应碳同化对ATP与NADPH的比例需求(3:2)
细胞生物学第六章 细胞的能量转换-线粒体和叶绿体

线粒体的形态结构
线粒体的形态与分布
动态细胞器 外形:线状、粒状、哑铃状、环状、圆柱状等 大小:直径0.5-1.0μm,长为2-3μm,可达1020μm 数目:动物细胞植物细胞 人成熟红细胞中无线粒体 代谢旺盛的细胞 分布:细胞功能旺盛区域;定位与迁移与微管相关
线粒体可塑性
心肌
状如蘑菇,属F型质子泵。 分为球形的F1(头部)和嵌入膜中的F0(基部)。 F1由5种多肽组成α3β3γδε复合体,具有三个ATP合成 的催化位点(每个β亚基具有一个)。 F0由三种多肽组成ab2c12复合体,嵌入内膜,12个c亚基组 成一个环形结构,具有质子通道。
F1 particle is the catalytic subunit; The F0 particle attaches to F1 and is embedded in the inner membrane. F1: 5 subunits in the ratio 3:3:1:1:1 F0: 1a:2b:12c
2、复合物II:琥珀酸脱氢酶
组成:至少由4条肽链,含有一个FAD,2个铁硫蛋白。 作用:催化琥珀酸的低能量电子转至辅酶Q,但不转移质子。
琥珀酸→FAD→Fe-S→Q。 琥珀酸+Q→延胡索酸+QH2
3、复合物III:细胞色素c还原酶。 组成:至少11条不同肽链,以二聚体形式存在,每个单体包 含两个细胞色素b(b562、b566)、一个铁硫蛋白和一个 细胞色素c1 。 作用:催化电子从辅酶Q传给细胞色素c,每转移一对电子, 同时将4个质子由线粒体基质泵至膜间隙。 2还原态cyt c1 + QH2 + 2 H+M→2氧化态cyt c1 + Q+ 4H+C
细胞生物学[第六章细胞的能量转换—一线粒体和叶绿体]课程预习
![细胞生物学[第六章细胞的能量转换—一线粒体和叶绿体]课程预习](https://img.taocdn.com/s3/m/f716503f67ec102de2bd8951.png)
第六章细胞的能量转换—一线粒体和叶绿体线粒体和叶绿体是细胞内的两种产能细胞器。
它们最初的能量来源有所不同,但却有着相似的基本结构,而且以类似的方式合成ATP。
线粒体是一种高效地将有机物转换为细胞生命活动的直接能源ATP的细胞器。
叶绿体通过光合作用把光能转换为化学能,并储存于糖类、脂肪和蛋白质等大分子有机物中。
线粒体和叶绿体都具有环状DNA及自身转录RNA与翻译蛋白质的体系。
线粒体和叶绿体都是半自主性的细胞器。
一、线粒体和氧化磷酸化线粒体通过氧化磷酸化作用,进行能量转换,为所需要的细胞进行各种生命活动提供能量。
(一)线粒体的形态结构1.线粒体的形态与分布线粒体一般呈粒状或杆状,但因生物种类和生理状态而异,可呈环形、哑铃形、线状、分权状或其他形状。
主要化学成分是蛋白质和脂类,其中蛋白质占线粒体干重的65%~70%,脂类占25%~30%。
一般直径0.5~1μm,长1.5~3.0μm,在胰脏外分泌细胞中可长达10~20μm,称巨线粒体。
数目一般数百到数千个,植物因有叶绿体的缘故,线粒体数目相对较少;肝细胞约1300个线粒体,占细胞体积的20%;单细胞鞭毛藻仅1个,酵母细胞具有一个大型分支的线粒体,巨大变形中达50万个;许多哺乳动物成熟的红细胞中无线粒体。
通常结合在维管上,分布在细胞功能旺盛的区域。
线粒体在细胞质中可以向功能旺盛的区域迁移,微管是其导轨,由马达蛋白提供动力。
2.线粒体的结构与化学组成线粒体的超微结构在电镜下观察到线粒体是由两层单位膜套叠而成的封闭的囊状结构。
主要由外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)、基质(matrix)或内室(inner chamber)四部分组成。
(1)外膜是包围在线粒体最外面的一层单位膜,光滑而有弹性,厚约6μm。
外膜上有排列整齐的筒状圆柱体,其成分为孔蛋白(porin),圆柱体上有小孔。
(2)内膜位于外膜内侧,把膜间隙与基质(内室)分开。
第6章 线粒体与细胞的能量转换

线粒体
线粒体与叶绿体
能量转换:
叶绿体:通过光合作用把光能转换为化学能,储存于大分子有机物中
线粒体:将储存在生物大分子中的化学能转换为细胞可直接利用的能源 线粒体与叶绿体的共性:半自主性细胞器 1. 形态特征主要是封闭的双层单位膜结构,内膜经过折叠演化为表 面极大扩增的内膜特化结构系统,在能量转换过程中起重要作用。 2. 功能都是高效的产生ATP的精密装置。 3. 遗传物质都具有环状DNA及自身转录RNA与翻译蛋白质的体系(第二 遗传信息系统)
氧化磷酸化的耦联机制—化学渗透假说
◆化学渗透假说内容:(获1978诺贝尔化学奖)
由英国Michell提出,认为在电子传递过程中,由于 线粒体内膜的不通透性,形成了跨线粒体内膜的质子梯度驱动
ATP的合成 (实验证据)。
◆支持化学渗透假说的实验证据该实验表明: 质子动力势乃ATP合成的动力
膜应具有完整性
线粒体中的氧化代谢
线粒体中的三羧酸循环,是物质氧化的最终共同途径,氧 化磷酸化是生物体获得能量的主要途径。
Electrons carried via NADH Electrons carried via NADH and FADH2
Glycolsis Pyruvate Glucose
Citric acid cycle
线粒体的化学组成
◆蛋白质(线粒体干重的65~70%) ◆脂类(线粒体干重的25~30%): 磷脂占3/4以上,外膜主要是卵磷脂, 内膜缺乏胆固醇,富含主要是心磷脂(20%)。
线粒体内、外膜脂类和蛋白质的比值:
0.3:1(内膜);1:1(外膜)
线粒体主要酶的分布
部位 酶的名称 单胺氧化酶 NADH-细胞色素 c 还原酶 犬尿酸羟化酶 酰基辅酶 A 合成酶 细胞色素氧化酶 ATP 合成酶 琥珀酸脱氢酶 β-羟丁酸脱氢酶 肉毒碱酰基转移酶 丙酮酸氧化酶 NADH 脱氢酶 部位 酶的名称 腺苷酸激酶 二磷酸激酶 核苷酸激酶 苹果酸脱氢酶 柠檬酸合成酶 延胡索酸酶 异柠檬酸脱氢酶 顺乌头酸酶 谷氨酸脱氢酶 脂肪酸氧化酶系 天冬胺酸转氨酶 蛋白质和核酸合成酶系 丙酮酸脱氢酶复合物
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第六章细胞的能量转换--线粒体和叶绿体(本章内容详见生物化学)一、线粒体的形态结构(一)线粒体的形态与分布线粒体是一个动态细胞器,在生活细胞中具有多样性、易变性、运动型和适应性等特点,其形状、大小、数量与分布在不同细胞内变动很大,机制在同一细胞,随着代谢条件的不同也会发生变化。
线粒体的形状多种多样,但以线状和粒状最常见。
(二)线粒体的结构与化学组成线粒体只有外层彼此平行的单位膜套叠而成的封闭的囊状结构。
外膜(outer membrane)起界膜作用,内膜(inner membrane)向内折叠形成嵴(cristae)。
外膜和内膜将线粒体分割成两个区室:一个是内外膜之间的腔隙,称为膜间隙(intermembrane space);另一个是内膜所包围的空间,称为基质(matrix)。
1、外膜线粒体最外面的一层单位魔结构,厚约6nm,光滑而有弹性。
外膜中蛋白质和脂质约各占50%。
外膜含有孔蛋白(porin),孔蛋白是由β链形成桶状结构,中心是一直径为2~3nm的小孔,即内部通道。
外膜的标志酶是单胺氧化酶(monoamine oxidase)。
2、内膜内膜是位于外膜的内侧把膜间隙与基质分开的一层单位膜结构,厚6~8nm。
相对外膜而言,内膜有很高的蛋白质/脂质比(质量比>3:1)。
内膜缺乏胆固醇,富含心磷脂(cardiolipin),约占磷脂含量的20%,心磷脂与离子的不可渗透性有关。
线粒体内膜除含有多种转运系统外,还含有大量的合成ATP的装置。
在内膜上有许多排列规则的颗粒,称为线粒体基粒(elementary particle),又称耦联因子1(coupling factor 1),简称F1,实质是ATP合酶(ATP synthesis)的头部。
3、膜间隙线粒体内、外膜之间的腔隙,宽6~8nm,但在细胞进行活跃呼吸时,膜间隙可扩大。
膜间隙中充满无定型液体,含有可溶性的酶、底物和辅助因子。
其中腺苷酸激酶是膜间隙的标志酶。
4、线粒体基质内膜所包围的嵴外空间为线粒体基质。
(P130)二、线粒体的功能线粒体是物质最终彻底氧化分解的场所,其主要功能是进行三羧酸循环及氧化磷酸化合成ATP,为细胞生命活动提供直接能量。
此外,线粒体海域细胞中氧自由基的生成,调节细胞艳华还原电位和信号转导,调控细胞凋亡、基因表达、细胞膜内多种离子的跨膜转运及电解质稳定平衡,包括线粒体对细胞中Ca2+的稳态调节等有关。
(一)线粒体中的氧化代谢在细胞中,线粒体是氧化还原的中心,是糖类、脂质和蛋白质最终氧化释能的场所。
线粒体中的三羧酸循环(tricarboxylic acid cycle),简称TCA循环,是物质氧化的最终共同途径,氧化磷酸化是生物获得能量的主要途径。
细胞之中糖酵解产生的NADH虽不能透过内膜浸入线粒体氧化,但是NDAH上的电子去能通过两种“穿梭”途径进入线粒体:①苹果酸-天冬氨酸穿梭途径(malate-asparate shuttle);②甘油-3-磷酸穿梭途径(glycerol-3-phosphate shuttle)。
(P132)当NADH中的1对电子传递到O2时,有10个H+被泵出,二FADH2中的1对电子的传递有6个H+被泵出。
(二)电子传递链与电子传递在线粒体内膜上存在的有关氧化磷酸化的脂蛋白复合物,是由一系列能可逆地接受和释放电子或H+的化学物质组成,它们在内膜上相互关联的有序排列构成呼吸链,称为电子传递链(electron transport chain)或呼吸链(respiratory chain),是典型的多酶体系。
电子通过呼吸链的流动,称为电子传递。
1、电子载体(electron carrier)参与电子传递链的电子载体有5种:黄素蛋白、细胞色素、泛醌、铁硫蛋白和铜离子。
黄素蛋白(flavoprotein),是由一条多肽与黄素蛋白腺嘌呤单核苷酸(FMN)或黄素腺嘌呤二核苷酸(FAD)紧密结合组成的结合蛋白。
细胞色素(cytochrome),是一种带有含铁血红素辅基二队可见光具有特征性强吸收的蛋白。
泛醌(ubiquinone,UQ)或称辅酶Q(coenzyme Q,CoQ),或简称为Q,是一种脂溶性的、带有一条的类异戊二烯侧链的苯锟。
铁硫蛋白(iron-sulfur protein),是一类含非血红素铁的蛋白质。
铜离子(copper atom),位于线粒体内膜的单个蛋白质分子内。
通过Cu2+和Cu+两种状态的变换,传递单个电子。
2、电子载体排列顺序(P135)呼吸链的最终受体是氧,氧接受电子后与H+结合生成水。
3、电子转运复合物复合物Ⅰ:NADH-CoQ还原酶,又称NADH脱氢酶。
既是电子传递体又是质子移位体。
组成:由25条以上多肽链组成,以二聚体形式存在,至少6个Fe-S中心和1个黄素蛋白,是呼吸链中最大最复杂的复合物;作用:催化NADH氧化,从中获得2个高能电子→辅酶Q;泵出4 H+(质子跨膜输送)复合物Ⅱ:琥珀酸-CoQ还原酶,又称琥珀酸脱氢酶。
是三羧酸循环中唯一一种结合在膜上的酶。
组成:含FAD辅基,2Fe-S蛋白和一个细胞色素b;作用:催化电子从琥珀酸→FAD→Fe-S→辅酶Q (无H+泵出)复合物Ⅲ:CoQ-细胞色素还原酶,以二聚体形式存在,既是电子传递体又是质子移位体。
组成:2个cytb(b562,b566),1cyt c11Fe-S蛋白;作用:催化电子从CoQ→cyt c;泵出4 H+;复合物Ⅳ:细胞色素C氧化酶,有6-13条多肽链组成,以二聚体形式存在。
组成:cyt a, cyt a3 ,2个Cu原子(CuA,CuB);作用:催化电子从cyt c→分子O2形成水。
(三)质子转移与质子驱动的形成(P137)(四)ATP形成机制——氧化磷酸化将ADP转变为ATP的过程成为磷酸化(phosphorylation)。
ADP磷酸化有2种途径:①底物水平的磷酸化;②氧化磷酸化(oxidative phosphorylation)。
1、ATP合酶的结构与组成ATP合酶是生物体能量转换的核心酶。
ATP合酶包括两个基本组分:球状的F1头部和嵌于内膜的F0基部。
2、能量耦联与ATP合酶的作用机制1979年,Boyer提出结合变构机制(binding change mechanism),①质子梯度的作用并不是用于形成ATP,而是使ATP从酶分子上解脱下来。
②ATP合酶上的3个β亚基的氨基酸序列是相同的,但它们的构象不同。
第二节叶绿体与光合作用植物细胞的质体具有不同的形态和功能,通常分为叶绿体(chloroplast)、有色体(chromoplast)和白色体(leucoplast)3大类。
第三节线粒体和叶绿体是半自主性细胞器一、半自主性细胞器的概念:自身含有遗传表达系统(自主性);但编码的遗传信息十分有限,其RNA转录、蛋白质翻译、自身构建和功能发挥等必须依赖核基因组编码的遗传信息(自主性有限)。
二、线粒体和叶绿体的DNA1、mtDNA /ctDNA形状、数量、大小(1)双链环状(除绿藻mtDNA,草履虫mtDNA)(2)mtDNA大小在动物中变化不大,但在植物中变化较大高等植物,120kbp~200kbp;2、mtDNA和ctDNA的复制(1)以半保留方式进行自我复制(2) mtDNA复制的时间主要在细胞周期的S期及G2期,DNA先复制,随后线粒体分裂。
ctDNA复制的时间在G1期。
(3) 复制仍受核控制.三、线粒体和叶绿体的蛋白质合成1、线粒体和叶绿体合成蛋白质的种类十分有限2、线粒体或叶绿体蛋白质合成体系对核基因组具有依赖性3、不同来源的线粒体基因,其表达产物既有共性,也存在差异4、参加叶绿体组成的蛋白质来源有3种情况:(1)由ctDNA编码,在叶绿体核糖体上合成;(2)由核DNA编码,在细胞质核糖体上合成;(3)由核DNA编码,在叶绿体核糖体上合成。
四、线粒体和叶绿体蛋白质的运送与组装1、线粒体蛋白质的运送与组装(1)定位于线粒体基质的蛋白质的运送(2)定位于线粒体内膜或膜间隙的蛋白质运送2、叶绿体蛋白质的运送及组装(1)定位于类囊体中的蛋白运送:前体蛋白N端转运肽分为两个区域,分别引导两步转运,其N端含有导向基质的序列,引导进入基质,而C端有导向类囊体的序列,又引导其穿过类囊体膜进入类囊体腔,转运肽经历两次水解,一次在基质中,一次在类囊体腔;(2)定位于基质中的蛋白运送:转运肽仅有导向基质的序列,在基质中被切除,转运肽成为成熟蛋白。
第四节线粒体和叶绿体的增殖与起源一、线粒体和叶绿体的增殖1、线粒体的增殖:(1)间壁分离,分裂时先由内膜向中心皱褶,将线粒体分类两个,常见于鼠肝和植物产生组织中。
(2)收缩后分离,分裂时通过线粒体中部缢缩并向两端不断拉长然后分裂为两个,见于蕨类和酵母线粒体中。
(3)出芽,见于酵母和藓类植物,线粒体出现小芽,脱落后长大,发育为线粒体。
2、叶绿体的发育和增殖:(1)个体发育:由前质体(proplastid)分化而来。
(2)增殖:分裂增殖二、线粒体和叶绿体的起源1、内共生起源学说(1)1905年,Mereschkowsky提出叶绿体起源于细胞内共生的蓝藻:(2)1970年,Margulis提出:线粒体的祖先-原线粒体是一种革兰氏阴性细菌;叶绿体的祖先是原核生物的蓝细菌,即蓝藻。
(3)内共生起源学说的主要论据:①基因组在大小、形态和结构方面与细菌相似。
②有自己完整的蛋白质合成系统,能独立合成蛋白质,蛋白质合成机制有很多类似细菌而不同于真核生物。
③两层被膜有不同的进化来源,外膜与细胞的内膜系统相似,内膜与细菌质膜相似。
④以分裂的方式进行繁殖,与细菌的繁殖方式相同。
⑤能在异源细胞内长期生存,说明线粒体和叶绿体具有的自主性与共生性的特征。
⑥线粒体的祖先很可能来自反硝化副球菌或紫色非硫光合细菌。
⑦发现介于胞内共生蓝藻与叶绿体之间的结构--蓝小体,其特征在很多方面可作为原始蓝藻向叶绿体演化的佐证。
(4)不足之处:①从进化角度,如何解释在代谢上明显占优势的共生体反而将大量的遗传信息转移到宿主细胞中?②不能解释细胞核是如何进化来的,即原核细胞如何演化为真核细胞?③线粒体和叶绿体的基因组中存在内含子,而真细菌原核生物基因组中不存在内含子,如果同意内共生起源学说的观点,那么线粒体和叶绿体基因组中的内含子从何发生?2、非共生起源学说:(1)主要内容:真核细胞的前身是一个进化上比较高等的好氧细菌。
(2)成功之处:解释了真核细胞核被膜的形成与演化的渐进过程。
(3)不足之处①实验证据不多②无法解释为何线粒体、叶绿体与细菌在DNA分子结构和蛋白质合成性能上有那么多相似之处③对线粒体和叶绿体的DNA酶、RNA酶和核糖体的来源也很难解释。
④真核细胞的细胞核能否起源于细菌的核区?。