第三章 光合作用
植物生理学第三章植物的光合作用

光合作用的过程
光能
H2O
光解 吸收
色素分子
O2 [H] 酶
供能
2C3
还
固
CO2
多种酶 定 C5
酶
ATP
酶
原
(CH2O)
ADP+Pi
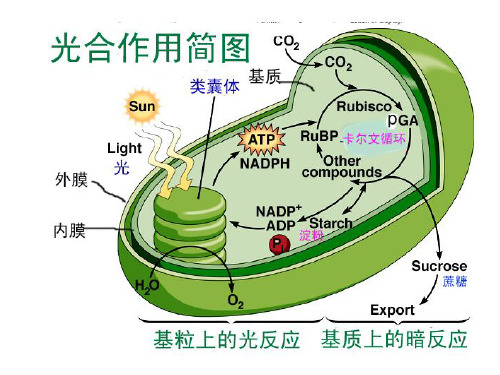
光反应阶段
暗反应阶段
水的光解:H2O 光解 2[H]+1/2 O2
酶
CO2的固定: CO2+C5 2C3
光合磷酸化:ADP+Pi+能量 酶
ATP
C3化合物还原:2 C3
光系统(PSII)
PSII的颗粒大,直径约17.5 nm,主要分布在类囊体膜的叠合部分。
➢ 晶体结构中的PSII为一个二聚体,二聚体的两个 单体呈准二次旋转对称。PSII单体具有36个跨膜α螺旋,其中D1和D2各5个,CP43和CP47各6个, Cytb559的α亚基和β亚基各自形成一个跨膜α-螺旋。 D1和D2蛋白与Cytb559的α和β亚基一起组成PSII 反应中心,是进行原初电荷分离和电子传递反应 的机构,CP47和CP43的主要功能是接受LHCII的 激发能量并传递到反应中心。
是否需光 需光 不一定,但受光促进 不一定,但受光促进
不同层次和时间上的光合作用
第二节 原初反应
➢ 原初反应 是指从光合色素分子被光激发,到引起 第一个光化学反应为止的过程。 ➢ 它包括: 光物理-光能的吸收、传递
光化学-有电子得失
原初反应特点 1) 速度非常快,10-12s∽10-9s内完成; 2) 与温度无关,(77K,液氮温度)(2K,液氦温度); 3) 量子效率接近1
表1 光合作用中各种能量转变情况
•
能量转变 光能 电能 活跃的化学能 稳定的化学能
第三章 光合作用 Photosynthesis1

图3-7 荧光现象
图3-9 色素分子吸收光后能量转变 色素分子吸收蓝光(430 nm)或红光(670 nm)后,分别激发为第二单线态(E2) 或第一单线态(E1),E1转变为第一三线态,它们进一步回到基态(E0)时则分别 产生荧光或磷光
• 四、植物的叶色 • 正常叶子的叶绿素和类胡萝卜素的分子比 例约为3:1,叶绿素a和叶绿素b也约为3: 1,叶黄素和胡萝卜素约为2:1。所以正常 的叶子总是呈现绿色。
• (1)非循环光合磷酸化 • 在放氧复合体( OEC)处水裂解后,把H+释放 到类囊体腔内,把电子传递到PSⅡ。电子在光 合电子传递链中传递时,伴随着类囊体外侧的H+ 转移到腔内,由此形成了跨膜的H+浓度差,引起 了ATP的形成;与此同时把电子传递到PSI去, 进一步提高了能位,而使H+还原NADP+为 NADPH,此外,还放出O2。在这个过程中,电 子传递是一个开放的通路,故称为非循环光合磷 酸化。吸收8个光量子,将2H2O氧化(将4个H+ 释放到囊中),放出一个O2。传递4个电子,还 原2个NADP+。光量子产额为1/8。
类囊体分为二类: 基质类囊体 又称基质片层,伸展在基质中彼此不重叠; 基粒类囊体 或称基粒片层,可自身或与基质类囊体重叠,组成基粒。 堆叠区 片层与片层互相接触的部分, 非堆叠区 片层与片层非互相接触的部分。
类囊体片层堆叠的生理意义 • ①膜的堆叠意味着捕获光能机构高度 密集,更有效地收集光能。 • ②膜系统常是酶排列的支架,膜的堆 叠易构成代谢的连接带,使代谢高效 地进行。 • ③类囊体片层堆叠成基粒是高等植物 细胞所特有的膜结构,它有利于光合作 用的进行。
光系统Ⅰ(PSI)
• (2)PSⅡ • PSⅡ多存在于基粒片层的垛叠区。PSⅡ主要 由核心复合体(P680)、PSⅡ捕光复合体 ( LHCⅡ)和放氧复合体(OEC)等亚单位 组成。PSⅡ的功能是利用光能氧化水(水的 光解和放氧)和还原质体醌。水裂解放氧是水 在光照下经过PSⅡ的作用,释放氧气,产生 电子,释放质子到类囊体腔内,整个反应如下: • 光子 • 2H2O→O2+4H++4e-
第三章 光合作用(3-5)

B.过渡阶段
C.饱和阶段
表观量子产额比理论值低,主要原因是:
①光没有全部被叶片吸收,存在反射和透射损失; ②非光合色素吸收了部分光能; ③光呼吸和暗呼吸对光合的负效应; ④形成的同化力(ATP、NADPH)没有全部用于CO2的还原; ⑤没有在饱和CO2浓度和最适温度下测定, 存在CO2扩散和 固定速率的限制等。
(4)消除乙醇酸
乙醇酸对细胞有毒害作用,它的产生在代谢 中是不可避免的。• 呼吸消除乙醇酸的代谢, 光 使细胞免受伤害。另外,光呼吸代谢中涉及 多种氨基酸的转化过程,它可能对绿色细胞 的氮代谢有利。
五、C3植物和C4植物的光合特征
C4植物比C3植物具有较强的光合作用,这与其结构特征 和生理特性有关。 (一)解剖结构特点
1.光强(light)
(1)光强-光合曲线 光补偿点(Light Compensation Point,LCP): 随着光强的增高,光合速率相应提高,当达到某一光强时,叶片 的光合速率与呼吸速率相等 ,净光合速率为零时的光强称为光补偿 点。 光饱和点(Light Saturation Point,LSP):
利用植物的需光特性确定合理的叶面积系数 (leaf area index, LAI)
单位土地面积上的叶面积
LAI= 单位土地面积
LAI是表示作物群体大小状况的一个指标。合理的LAI应 以作物的需光特性为标准,以最大限度的利用光能、地力为 原则,以高产、优质、低消耗为目的。从作物的需光特性上 讲,应使上部叶片处于光饱和点,中下部叶片处于光补偿点 的两倍以上,以保证下部叶片及根系的正常生长。
在叶绿体与过氧化体中吸收氧气,在线粒体中放出CO2
二、光呼吸的生化历程
叶绿体 3-PGA + 磷酸乙醇酸 乙醇酸 C3—cycle C2—cycle [O2] Rubisco RUBP 2 3—PGA [CO2] 过氧化体
第三章 光合作用--光反应-电子传递
PSⅡ复合体
PSⅠ复合体
Cytb6/f复合体
质蓝素
铁氧化蛋白和 铁氧化蛋白— NADP+还原酶
质醌(PQ)
②质醌(PQ) (Plastoquinone)
质体醌为脂溶性分子,能在类囊体膜中自由移动,转运电子与质子。 质体醌在膜中含量很高,约为叶绿素分子数的5%~10%,故有“PQ库” 之称。 PQ库作为电子、质子的缓冲库,能均衡两个光系统间的电子传递
光系统Ⅱ(PhotosystemⅡ,PSⅡ)
吸收短波红光(680nm)
光系统Ⅰ(PhotosystemⅠ,PSⅠ)
吸收长波红光(700nm)
光系统 II(PSⅡ)
D P A
PSⅡ的原初电子供体 反应中心色素 PSⅡ的原初电子受体 次级电子受体
H2 O P680
去镁叶绿素分子(Pheo)
Q(质体醌)
若干个β-胡萝卜素
三种电子载体分别为A0、A1
3个不同的Fe4-S4蛋白:Fx、FA、FB
⑤PSⅠ复合体功能:
吸收光能,进行光化学反应,传递电子从PC到NADP+
PSⅠ复合体 Cytb6/f复合体
DPA
反应中心色素分子(reaction center piment,P) 原初电子受体(primary electron acceptor,A)
原初电子受体(D)
指直接接收反应中心色素分子传来电子的电子传递体
反应中心色素分子(P680和P700)
是光化学反应中最先向原初电子受体供给电子的,因此 反应中心色素分子又称原初电子供体
蕴含的意义
第三节 原初反应 (Primary reaction)
指从光合色素分子被光激发到引起第一个光化学反应为止的 过程。包括光能的吸收、传递与光化学反应.
七年级(初一)生物 生物 第3章植物的光合作用

一、光反应和碳反应
光合作用的过程可分为3大步骤:1)原初反应(光能的吸收、传递和转换过程);2)电子传递和光合磷酸化(电能转化为活跃的化学能过程);3)碳同化(活跃的化学能转变为稳定的化学能过程)。第一、二个大步骤基本属于光反应,第三个大步骤属于暗反应(表3-2)。
2.C4途径的类型
根据运入维管束鞘细胞的C4化合物和脱羧反应的不同,C4途径有3种类型(表3-3,图3-18)。
3.C4植物的光合特征
C4植物比C3植物具有较强的光合作用,其原因可从结构和生理两方面来探讨。
①结构与功能是有密切关系的,是统一的。C4植物叶片有“花环型”结构。
②在生理上,
C4植物的叶肉细胞中的PEPC对底物HCO3-的亲和力极高(是Rubisco60倍);极低的CO2供应就可满足它的需要。
②已从叶绿体分离出两个光系统,每一个光系统具有特殊的色素复合体及一些物质。光系统I(简称PSI)的颗粒较小,直径约11nm,主要分布在类囊体膜的非叠合部分;光系统Ⅱ(简称PSⅡ)的颗粒较大,直径约17.5nm,主要分布在类囊体膜的叠合部。光合作用的光化学反应就在.这两个光系统中进行。
二、电子传递体及其功能
C4植物由于有“CO2泵”浓缩CO2的机制,降低了光呼吸;提高了BSC的CO2浓度,抑制了RuBisco氧化反应,降低了光呼吸;光呼吸酶主要分布在BSC细胞,即便是有CO2放出,也易被PEPC再固定。
第二节叶绿体及光合作用色素(chloroplastandchloroplastpigments) )
叶片是进行光合作用的主要器官,而叶绿体是进行光合作用的主要细胞器。
一、叶绿体的结构和成分
(一)叶绿体的结构(Struture ofchloroplast)
第三章 光合作用--光反应-电子传递

1. 光合电子传递链
指定位在光合膜上的,由 多个电子传递体组成的电 子传递的总轨道.
光合电子传递Z链
希尔(1960)-“Z”方案(“Z” scheme) ,即电子传 递是在两个光系统串联配合下完成的,电子传递体按氧 化还原电位高低排列,使电子传递链呈侧写的“Z”形。
“Z”链特点:
(1)电子传递链主要由光合膜上 的PSⅡ、Cyt b6/f、PSⅠ三个 复合体串联组成。
ps和ps的光化学反应光系统photosystemps光系统photosystemps吸收短波红光680nm吸收长波红光吸收长波红光700nm光系统iipsps的原初电子受体反应中心色素次级电子受体反应中心色素次级电子受体去镁叶绿素分子pheodpaq质体醌p680ps的原初电子供体h2o光系统ipsips的原初电子受体反应中心色素最终电子受体的原初电子受体反应中心色素最终电子受体fddpap700psi的原初电子供体pcnadp功能与特点吸收光能光化学反应电子供体功能与特点吸收光能光化学反应电子供体反应中心色素分子原初电子供体反应中心色素分子原初电子供体次级电子受体末端电子受体次级电子受体末端电子受体ps还原nadp实现pc到nadp实现pc到nadp的电子传递pcp700铁硫中心fd铁硫中心fdnadp电子最终受体电子最终受体ps使水裂解释放氧气并把水中的电子传至质体醌
原初电子受体(D)
指直接接收反应中心色素分子传来电子的电子传递体
反应中心色素分子(P680和P700)
是光化学反应中最先向原初电子受体供给电子的,因此 反应中心色素分子又称原初电子供体
两个光系统的发现
“红降”现象
波长大于680 nm(用685 nm)的光照射时,小球藻的光合 量子产额明显下降,被称为“红降”现象.
第三章 植物的光合作用

2.意义 (1)将无机物转变成有机物 CO2 + 4H+ —— (CH2O) (2)随着物质的转变,将光能转变为化学能。
光能电能活跃的化学能稳定的化学能 hv eATP、NADPH CH2O
(3)释放氧,净化空气;并产生臭氧,滤去太 阳光中对生物有强烈破坏作用的紫外光。
第三章 植物的光合作用
430 nm波长的蓝光量子的能量为4.5710-19J
(二)吸收光谱(absorption spectrum) 太阳光到达地表面的波长大约是 300 nm —— 2600 nm 可见光的波长是 390 nm —— 770 nm 连续光谱: 光束通过三棱镜后可把白光分为七色 连续光谱。
吸收光谱:
第三、四、五节 光合作用过程
光合作用机制的研究
• 在研究外界条件影响时发现:弱光下增加光强能提高 光合速率,但当光强增加到一定值时,光合速率便不 再随光强的增加而提高;此时提高温度或CO2浓度才 能增强光合速率。由此推理,光合作用至少有两个步 骤:其一需要光,另一个则与温度相关。 希尔反应的发现和水氧化钟模型的提出。 藻类闪光实验:在光能相同时,一种用连续光照;另一 用闪光照射,中间间隔一暗期。发现后者的光合速率 比前者的要高。表明,光合作用不是任何步骤都需要 光。根据需光与否,将光合作用分为两个反应—光反 应和暗反应。
(1)叶绿素a在红光区的吸收带偏向长光波方 向,而在兰紫光区则偏向短光波方向.
(2)叶绿素a在红光区的吸收带宽些,兰紫光 区窄些,叶绿素b与其相反.
2.类胡罗卜素的吸收光谱 1)吸收光谱 —— 兰紫光区 2)重要功能 —— 吸收光能并向叶绿素传递 ——可进行光保护 吸收兰紫光形成激发态的类胡罗卜素 热耗散返回基态淬灭激发态的叶绿素 避免吸收的多余能量对光合系统的伤害
植物生理学第三章植物的光合作用

植物生理学第三章植物的光合作用第三章植物的光合作用一、名词解释1. C3途径2. C4途径3. 光系统4. 反应中心5. 原初反应6. 荧光现象7. 红降现象8. 量子产额9. 爱默生效应10. PQ循环11. 光合色素12. 光合作用13. 光合单位14. 反应中心色素15. 聚光色素16. 解偶联剂17. 光合磷酸化18. 光呼吸19. 光补偿点20. CO2补偿点21. 光饱和点22. 光能利用率23. 光合速率二、缩写符号翻译1. Fe-S2. PSI3. PSII4. OAA5. CAM6. NADP+7. Fd 8. PEPCase 9. RuBPO10. P680、P700 11. PQ 12. PEP13. PGA 14. Pheo 15. RuBP16. RubisC(RuBPC) 17. Rubisco(RuBPCO) 18.TP三、填空题1. 光合作用的碳反应是在中进行的,光反应是在中进行的。
2. 在光合电子传送中最终电子供体是,最终电子受体是。
3. 在光合作用过程中,当形成后,光能便转化成了活跃的化学能;当形成后,光能便转化成了稳定的化学能。
4. 叶绿体色素提取掖液在反射光下观察呈色,在透射光下观察呈色。
5. P700的原初电子供体是,原初电子受体是。
6. 光合作用的能量转换功能是在类囊体膜上进行的,所以类囊体亦称为。
7. 光合作用中释放的氧气来自于。
8. 与水光解有关的矿质元素为。
9. 和两种物质被称为同化能力。
10. 光的波长越长,光子所持有的能量越。
11. 叶绿素吸收光谱的最强吸收区有两个:一个在,另一个在。
12. 光合磷酸化有三种类型:、、。
13. 根据C4化合物和催化脱羧反应的酶不同,可将C4途径分为三种类型:、、。
14. 一般来说,正常叶子的叶绿素和类胡萝卜素的分子比例为;叶黄素和胡萝卜素的分子比例为。
15. 光合作用中,淀粉的形成是在中,蔗糖的形成是在中。
16. C4植物的C3途径是在中进行的;C3植物的卡尔文循环是在中进行的。
植物生理学第三章植物的光合作用

植物生理学第三章植物的光合作用植物的光合作用是指植物利用光能将二氧化碳和水转化成有机物质(如葡萄糖)和氧气的过程。
其反应方程式为:6CO2+6H2O+光能→C6H12O6+6O2光合作用是植物最重要的生理过程之一,它不仅是植物能够生存和生长的基础,还能为其他生物提供氧气和有机物质。
光合作用通过光合色素和叶绿体等生理结构,具有高效和专一性的特点。
植物的光合作用可以分为两个阶段:光能捕获和光化学反应、以及碳固定和假单胞菌循环。
在光能捕获和光化学反应阶段,植物的光合色素(如叶绿素)能够捕获太阳光,并将其转化为化学能。
光合作用发生在叶绿体内,主要以叶绿体膜的光合作用单位,光系统(PSI和PSII)为中心。
光系统中的光合色素吸收太阳光,并将其能量传递给反应中心,激发电子。
通过光合色素的电子传递链,电子在PSII和PSI之间进行转移,最终转移到还原辅酶NADP+上,形成还原辅酶NADPH。
在碳固定和假单胞菌循环阶段,植物利用还原辅酶NADPH和ATP的能量,将二氧化碳转化为有机化合物。
这个过程称为Calvin循环,也叫柠檬酸循环。
Calvin循环包括三个主要步骤:碳固定、还原和再生。
首先,二氧化碳与从光合作用过程中产生的核酮糖五磷酸(RuBP)结合,形成不稳定的六碳中间体。
然后,该中间体通过一系列酶的作用,将其分解为两个三碳化合物,3-磷酸甘油醇醛(3-PGA)。
最后,3-PGA经过一系列的加氢还原反应和磷酸化反应,合成出葡萄糖和其他有机物质。
光合作用的速率受到光照、温度、二氧化碳浓度和水分等环境条件的影响。
光合速率随着光照强度的增大而增加,但达到一定的饱和点后,光合速率趋于稳定。
温度对光合作用的影响是复杂的。
在适宜温度下,光合速率随着温度的升高而增加,因为反应速率加快。
然而,当温度超过一定范围时,光合作用会受到抑制,因为高温会破坏光系统和酶的结构。
二氧化碳浓度越高,光合速率越快。
水分对光合作用的影响主要是通过调节植物的气孔进行的。
第三章 光合作用--暗反应-C4

C4能量
C 4 植物同化 CO 2 消耗的能量比 C 3 植物多,也可以说这个
“ CO 2 泵”是要由 ATP 来开动的,故在光强及温度较低的情 况下,其光合效率还低于 C 3植物。可见 C 4途径是植物光合 碳同化对热带环境的一种适应方式。
PEP底物再生要消耗2个ATP,这使得C4植物同 化1个CO2需消耗5个ATP与2个NADPH。
丙酮酸 MAL或ASP
PEP
丙酮酸
维管束鞘细胞
C4途径的三种亚类型
根据植物所形成的初期C4二羧酸的种类以及脱羧反应参与的酶 类,可把C4途径分为三种亚类型:
(1) 依赖NADP的苹果酸酶(NADP malic enzyme)的苹果酸型 (NADP-ME型); 甘蔗、玉米、谷子、高粱;地锦、半子莲、地肤
(3)再生阶段
5GAP+3ATP+2H2O→→→3RuBP+3ADP+2Pi+3H+ 这里包括形成磷酸化的 3,4,5,6 和7 碳糖的一系列反应。 最后由核酮糖 -5- 磷酸激酶 (Ru5PK) 催化,消耗 1 分子 ATP,再形成RuBP。
C3途径的总反应式
3CO2+5H2O+9ATP+6NADPH→GAP+9ADP+8Pi+6NADP++3H+
• C3途径 (C3 pathway)-卡尔文循环 • C4途径 (C4 pathway) • CAM (Crassulacean acid metablism) 途径
C3途径学习导图—讲故事
故事概况 故事背景 C3途径 发生地点 特点 发生时间
故事主角
C3途径的发现
1946年 美国 M. Calvin and B. Benson
植物生理第三章

(二)原初反应的过程 —光能的吸收、传递和光化学反应
1.天线色素接受光能,以诱导共振方式 将能量传递到光合反应中心。
能量传递效率: Chla,b几乎100%传给反应中心色素, 类胡萝卜素 约90%传给反应中心色素。
42
2.光合反应中心发生光化学反应
hυ ┋ D P A → D P* A → D P+ A- → D+ P A①特殊叶绿素a ②高能电子脱离,
9
外被膜
被膜 (envelop)
内被膜 选择透性
叶绿体
(Chloroplast)
膜—光合色素、光合链——原初反应、 电子传递和光合磷酸化(光合膜 ) (thylacoid) 类囊体 腔—光合放O2 间质(stroma)——光合碳循环酶(Rubisco), CO2固定(同化); DNA,RNA,核糖体70S——部 分遗传自主
醛基(CHO)
14
1.叶绿素的结构:
②双羧酸尾部:
1个羧基在副环(V)上 以酯键与甲基结合 --甲基酯化; 另一个羧基(丙酸) 在IV环上与植醇 (叶绿醇)结合- -植醇基酯化; 非极性,亲脂,插 入类囊体的疏水区, 起定位作用。
15
2.叶绿素的作用:
收集和传递光能 (大部分Chl a和全部Chl b) 将光能转换为电能(少数特殊Chl a)
2.类胡萝卜素
强吸收区: 400-500 (蓝紫); 不吸收区: 500以上
25
(二)光能的吸收和释放
物质吸收光子,其原子中的e 重新排列,分子从基态(最 低、最稳定)跃迁到激发态 (高能、不稳定)
Chl+ hγ= Chl* 处于激发态的分子,趋 于释放能量回到基态
26
第三章 光合作用2

小 结
• 光合作用是地球上最重要的化学反应。 • 叶绿体是进行光合作用的细胞器。类囊体 (光合膜)是光反应的主要场所,基质是 暗反应的场所。叶绿体的色素有下列2类: 1)叶绿素,主要是叶绿素a和叶绿素b;2) 类胡萝卜素,其中有胡萝卜素和叶黄素。 在这2类色素中,叶绿素最主要。
• 光合作用是光反应和暗反应的综合。整个 光合作用大致可分为下列3大步骤:原初反 应、电子传递和光合磷酸化、碳同化。 • 光能的吸收、传递和转换过程是通过原初 反应完成的。聚光色素吸收光能后,通过 诱导共振方式传递到反应中心,反应中心 色素分子的状态特殊,能引起由光激发的 氧化还原,电荷分离,就将光能转换为电 能,送给原初电子受体。
• 二、光呼吸的生理功能 • 一种比较流行的观点是,在干旱和高辐射 期间,气孔关闭,CO2不能进入,会导致光 抑制。此时光呼吸释放CO2,消耗多余能量, 对光合器官起保护作用,避免产生光抑制。 • 另一种观点是,Rubisco同时具有羧化和加 氧的功能,在有氧条件下,光呼吸虽然损 失一些有机碳,但通过C2循环还可收回 75%的碳,避免损失过多。
• • • •
卡尔文循环的总反应式如下: 5H2O+3CO2+9ATP+6(NADPH+H+) → CHOCHOHCH2OP+9ADP +8Pi +6NADP+ 由此可见,要产生1个PGAld(磷酸丙糖)分 子,需要3个CO2分子,6个NADPH2分子和9 个ATP分子。
• (二)C4途径 • 1. C4途径的反应 • C4途径包括4个步骤:(1)羧化; (2) 转移;(3)脱羧与还原;(4)再生。
CH2O(P) C=O +O2 HCOH COOH HCOH COOH CH2O(P) RuBP 2-磷酸乙醇酸 3-磷酸甘油酸 → CH2O(P) + 2HCOH CH2O(P)
第三章光合作用作业及答案

第三章光合作用、名词解释1 •光合色素:指植物体内含有的具有吸收光能并将其用于光合作用的色素,包括叶绿素、类胡萝卜素、藻胆素等。
2 •原初反应:包括光能的吸收、传递以及光能向电能的转变,即由光所引起的氧化还原过程。
3 •红降现象:当光波大于685nm时,虽然仍被叶绿素大量吸收,但量子效率急剧下降,这种现象被称为红降现象。
4. 爱默生效应:如果在长波红光(大于685nm)照射时,再加上波长较短的红光(650nm),则量子产额大增,比分别单独用两种波长的光照射时的总和还要高。
5 •光合链:即光合作用中的电子传递。
它包括质体醌、细胞色素、质体蓝素、铁氧还蛋白等许多电子传递体,当然还包括光系统I和光系统II的作用中心。
其作用是水的光氧化所产生的电子依次传递,最后传递给NAD P。
光合链也称Z链。
6 •光合作用单位:结合在类囊体膜上,能进行光合作用的最小结构单位。
7 •作用中心色素:指具有光化学活性的少数特殊状态的叶绿素a分子。
&聚光色素:指没有光化学活性,只能吸收光能并将其传递给作用中心色素的色素分子。
聚光色素又叫天线色素。
9 •希尔反应:离体叶绿体在光下所进行的分解水并放出氧气的反应。
10. 光合磷酸化:叶绿体(或载色体)在光下把无机磷和ADP转化为ATP,并形成高能磷酸键的过程。
11 •光呼吸:植物的绿色细胞在光照下吸收氧气,放出CQ的过程。
光呼吸的主要代谢途径就是乙醇酸的氧化,乙醇酸来源于RuBP的氧化。
光呼吸之所以需要光就是因为RuBP的再生需要光。
12. 光补偿点:同一叶子在同一时间内,光合过程中吸收的CO和呼吸过程中放出的CO等量时的光照强度。
13. CO补偿点:当光合吸收的CQ量与呼吸释放的CO量相等时,外界的CO浓度。
14. 光饱和点:增加光照强度,光合速率不再增加时的光照强度。
15・光能利用率:单位面积上的植物光合作用所累积的有机物所含的能量,占照射在相同面积地面上的日光能量的百分比。
第三章 光合作用(2-1)

图3-6 光合作用单位图解 表示天线色素分子捕获的一个光量子传递到反应中心色素分 子,在那里进行光化学反应,发生电荷分离。D为电子供体, A为 电子受体。
去镁叶绿素
去镁叶绿素
副叶绿素
副叶绿素
胡萝卜素
配对叶绿素
光系统‖的反应中心
2.光化学反应
光化学反应实际就是由光引起的反应中心色素分 子与原初电子受体间的氧化还原反应: P·A ────→ P*·A ────→ P+· A 扩展的原初反应过程: D·(P+·A-)·A1 ──→ D+·(P·A)·A1- 供电子给P+的还原剂叫做次级电子供体(D); 从A-接受电子的氧化剂叫做次级电子受体(A1)。
2.电子传递(electron transport)
光合电子传递是由两个光系统串联进行。 连接着两个光反应之间的电子传递,是由几种排列紧密的电 子传递体完成的。
各种电子传递体具有不同的氧化还原电位,负值越大代表还 原势越强,正值越大代表氧化势越强。各种电子传递体按氧化还 原电位高低排列,组成“Z”形电子传递链,电子定向转移。 光合链
光 合 电 子 传 递 的 Z 方 案
水的光解与 放氧
PSII光化 学反应 Cytb6/f复合 体的电子传 递 PSI光化学 反应 电子传递末端 NADPH的形成
光合电子传递的“Z”方案
从图中可以看出:
①电子传递主要由光合膜上的PSⅡ、Cytb6/f、PSⅠ三个复合 体串联组成, 各电子传递体按电位高低排列;
PSⅠ和PSⅡ的光化学反应
PSⅠ的原初电子受体是叶绿素分子(A0),PSⅡ的原初电子 受体是去镁叶绿素分子(Pheo),它们的次级电子受体分别 是铁硫中心和醌分子(表4-2)。 PSⅠ的原初反应: P700·A0 hυ P700*·A0 P700+·A0- (17)
第三章 光合作用

第三章光合作用名词解释:1、光合作用:绿色植物利用光能,把二氧化碳和水合成有机物质,并释放出氧气的过程。
2、光合速率:即光合强度,指单位时间单位叶面积所吸收的二氧化碳或释放的氧量,或单位时间单位叶面积所积累的干物质量,常用以下单位表示:CO2m g·dm-2·h-13、净光和强度:即表现光合强度,指总光合减去被测部位同时进行的呼吸强度。
4、原初反应:是光合作用的起点,指光合色素吸收日光能所引起的光物理及光化学过程。
包括光能的吸收和色素分子激发态的形成;天线色素分子间能量的传递;作用中心对光能的捕获、电荷分离。
5、光化学反应:是指反应中心色素分子吸收光能所引起的氧化还原反应。
6、反应中心:由反应中心色素分子及其原初电子受体与原初电子供体所组成,聚光色素分子吸收光能,传递到反应中心,反应中心色素分子被光量子所激发,失去电子呈氧化态,原初电子受体接受电子而被还原,反应中心色素分子失去电子即带正电荷,又可从它的原初电子供体获得电子而回复原状。
7、同化力:在电子传递及光合磷酸化作用中形成的NADPH+H+和ATP,随后用于CO2的同化,故称为同化力。
8、光呼吸:指绿色植物细胞在光下吸收O2,氧化乙醇酸,放出CO2的过程,称为光呼吸9、光饱和现象:在光照强度较低时,光合速率随光强的增加而相应增加;光强进一步提高时,光合速率的增加逐渐减小,当超过一定光强时即不再增加,这种现象称光饱和现象10、光饱和点:开始达到光饱和现象时的光照强度称为光饱和点。
11、光补偿点:在在光饱和点以下,光合速率随光照强度的减小而降低,到某一光强时,光合作用中吸收的CO2与呼吸作用中释放的CO2达动态平衡,这时的光照强度称为光补偿点。
12、CO2饱和点:在一定范围内,植物净光合速度随CO2浓度增加而增加,但到达一定程度时再增加CO2浓度,净光合速率不再增加,这时的CO2浓度称为二氧化碳饱和点13、CO2补偿点:在CO2饱和点以下,光合作用吸收的CO2与呼吸同光呼吸释放的CO2 达动态平衡,这时环境中的CO2浓度称为CO2补偿点。
植物生理学:第三章 植物的光合作用

第一节 光合作用的意义 第二节 叶绿体与光合色素 第三节 光合作用机理 第四节 光呼吸 第五节 影响光合作用的因素 第六节 植物对光能的利用 第七节 有机物的运输与分配
本章重点、难点及复习思考题
1
第一节 光合作用的意义 一 光合作用的概念 二 光合作用的意义
2
一 光合作用(photosynthesis) 绿色植物吸收光能,同化CO2和水,
9
◆ 叶绿素分子结构 ◇ chla是叶绿酸的酯。叶 绿酸是双羧酸,其中一个羧 基被甲醇所酯化,另一个被 植醇所酯化。
10
◇ Chla与chlb结构区别 chla第二个吡咯环上一个甲基(-CH3)被醛基(-
CHO)所取代,即为chlb .
11
◇ 叶绿素分子头部
▽ 4个吡咯环组成的卟啉环, 由4个甲烯基(=CH-)连成大的卟 啉环;
15
3 色素的功能
◆ 叶绿素chla和chlb都能吸收光能,少数chla 具光化学活性,能将光能转换成电能。
◆ 类胡萝卜素吸收光能、光保护,保护叶绿色 分子避免其在强光下的光氧化。
16
4 色素的光学特性
◆ 光合作用可利用的光:400 -700nm. ◇光子(photon)或光量子(quantum): 光是一
存在形式:色素蛋白复合体(pigment protein
complex)
2 结构和性质 (1) 叶绿素(chlorophyll): ◆ 叶绿素a (chla)、叶绿素b(chlb).
8
◆ 主要理化性质 ◇ 不溶于水,溶于有机溶剂,如乙醇、 丙酮、乙醚、氯仿等。 ◇ chla 呈蓝绿色,chlb 呈黄绿色。
黄化现象:缺乏某些条件而影响叶绿素合 成,使叶子发黄的现象.
光合作用-植物生理-图文

光合作用-植物生理-图文第三章植物的光合作用碳素营养是植物的生命基础,这是因为,第一,植物体的干物质中90%以上是有机化合物,而有机化合物都含有碳素(约占有机化合物重量的45%),碳素成为植物体内含量较多的一种元素;第二,碳原子是组成所有有机化合物的主要骨架,好象建筑物的栋梁支柱一样。
碳原子与其他元素有各种不同形式的结合,由此决定了这些化合物的多样性。
按照碳素营养方式的不同,植物可分为两种:1)只能利用现成的有机物作营养,这类植物称为异养植物(heterophyte),如某些微生物和少数高等植物;2)可以利用无机碳化合物作营养,并且将它合成有机物,这类植物称为自养植物(autophyte),如绝大多数高等植物和少数微生物。
异养植物与自养植物相比,后者在植物界中最普遍,而且非常重要。
这里我们着重讨论自养植物。
自养植物吸收二氧化碳,将其转变成有机物质的过程,称为植物的碳素同化作用(carbonaimilation)。
植物碳素同化作用包括细菌光合作用、绿色植物光合作用和化能合成作用3种类型。
在这3种类型中,绿色植物光合作用最广泛,合成的有机物质最多,与人类的关系也最密切,因此,本章重点阐述绿色植物光合作用(以下简称光合作用)。
第一节光合作用的重要性绿色植物吸收阳光的能量,同化二氧化碳和水,制造有机物质并释放氧气的过程,称为光合作用(photoynthei)。
光合作用所产生的有机物质主要是糖类,贮藏着能量。
光合作用的过程,光合作用的重要性,可概括为下列3个方面:1.把无机物变成有机物植物通过光合作用制造有机物的规模是非常巨大的。
据估计,地球上的自养植物每年约同化2某lOt碳素,其中40%是由浮游植物同化的,余下60%是由陆生植物同化的(图3-1)。
如以葡萄糖计算,整个地球每年同化的碳素相当于四五千亿吨有机物质,难怪人们把绿色植物喻为庞大的合成有机物的绿色工厂。
绿色植物合成的有机物质,可直接或间接作为人类和全部动物界的食物(如粮、油、糖、牧草饲料、鱼饵等),也可作为某些工业的原料(如棉、麻、橡胶、糖等)。
第三章 光合作用(2-2)

反应部位:C3途径的各反应均在叶绿体基质中进行。
过程可分为: 羧化、还原、再生 三个阶段 (如图3AA + NADPH + H+ Mal + NADP+
aspartate amino transferase
OAA + 谷氨酸 Asp + α- 酮戊二酸
④底物再生
1.C4途径的反应过程
基本上可分为四步反应: ①羧化反应
②还原或转 氨作用 ③脱羧反应
④底物再生
1.C4途径的反应过程
(二)C4途径
叶肉细胞(MC)中含 有磷酸烯醇式丙酮 酸羧化酶PEPC,进行 C4途径,固定CO2, 并将含四个碳的二 羧酸运到维管束鞘 细胞中;而维管束 鞘细胞(BSC)中含 有Rubisco等参与C3 途径的酶,进行CO2 的同化。
C-4 photosynthsis involves the separation of carbon fixation and carbohydrate systhesis in space and time
(一)C3途径 (1)羧化阶段
(2)还原阶段
(3)再生阶段
(一)C3途径 (1)羧化阶段
即3-磷酸甘油酸被还原为甘油醛-3-磷酸 的阶段,有二步反应:磷酸化和还原
首先,ATP提供能量, 把PGA 进一步磷酸化
(2)还原阶段
NADP-PGAkinase
(3)再生阶段
2PGA + 2ATP 2 1,3—DPGA + 2ADP 然后,进一步将DPGA还原成糖
第三章植物的光合作用_植物生理学

第三章植物的光合作用_植物生理学第三章:植物的光合作用植物的光合作用是植物生理学中一个非常重要的过程,通过光合作用,植物能够将光能转化为化学能,并且产生出氧气和有机物质,为植物自身生长和发育提供能量和养分,也间接地为其他生物提供能源。
植物的光合作用是在叶绿体中进行的。
叶绿体是植物细胞中的一种细胞器,它含有叶绿素,可以吸收太阳光中的能量。
光合作用主要包括光能的吸收、光能的转换和产物的合成三个过程。
首先,光能的吸收过程。
植物的叶绿体中含有多种不同类型的叶绿素,它们能够吸收不同波长的光。
叶绿素中的色素分子吸收光子后激发,成为激发态叶绿素。
不同的叶绿素吸收不同波长的光,其中最主要的是吸收红光和蓝光的叶绿素a,然后是辅助叶绿素如叶绿素b和叶黄素等。
叶绿体中的叶绿素主要吸收短波长的光,因此植物呈现出绿色。
其次,光能的转换过程。
当叶绿素吸收光子之后,其中的电子被激发出来,并且通过一系列的电子传递过程,在两个光化学反应中最终形成高能态分子ATP和NADPH。
这两种高能物质是植物光合作用最重要的产物,它们为植物提供了能量和电子。
ATP是一种能量通货,它可以通过释放磷酸基团的能量来驱动其他细胞活动。
NADPH是一种电子载体,它可以将电子传递给碳固定反应中的酶,驱动二氧化碳的还原反应。
最后,产物合成过程。
产生的ATP和NADPH被用来驱动碳固定反应,也就是光合作用的第二阶段。
在这个阶段中,植物利用ATP和NADPH将二氧化碳还原成有机物质。
这个过程中最重要的酶是光合酶RuBisCO,它将二氧化碳与一种五碳糖RuBP反应生成六碳糖,然后分解成两个三碳糖PGA。
PGA在一系列酶催化作用下转化为三碳糖G3P,部分G3P能够通过其他途径转化为其他有机物质,但大部分会再次参与碳固定反应生成更多的RuBP。
总结起来,植物的光合作用是植物生理学中的一个重要过程,通过光合作用植物能够利用太阳能将二氧化碳和水转化为有机物质并产生氧气。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第三章光合作用习题一、名词解释光合作用:绿色植物利用太阳光能,将二氧化碳和水合成有机物质,并释放氧气的过程。
原初反应:指的是光能的吸收、传递与转换过程,完成了光能向电能的转变,实质是由光所引起的氧化还原过程。
天线色素:又称聚光色素,没有光化学活性,将所吸收的光有效地集中到作用中心色素分子,包括99% 的叶绿素a ,全部叶绿素b ,全部胡萝卜素和叶黄素。
反应中心色素分子:既能吸收光能又具有化学活性,能引起光化学反应的特殊状态的叶绿素 a 分子,包括P 700 和P 680 。
光合作用单位:是指完成 1 分子CO2的同化或1分子O2 的释放,所需的光合色素分子的数目,大约是2400 个光合色素分子。
但就传递 1 个电子而言,光合作用单位是600 ,就吸收 1 个光量子而言,光合作用单位是300 。
红降现象:当光波大于680 nm ,虽然仍被叶绿素大量吸收,但光合效率急剧下降,这种在长波红光下光合效率下降的现象,称为红降现象。
光合效率:(量子产额)又称量子产额或量子效率,是光合作用中光的利用效率,即吸收 1 个光量子所同化二氧化碳或放出氧分子的数量。
量子需要量:同化1分子CO2 或释放 1 分子O2 需要的光量子数。
爱默生效应:(双光增益效应)如果在长波红光照射时,再加上波长较短的红光(650~670nm )照射,光合效率增高,比分别单独用两种波长的光照射时的总和还要高,这种现象称为双光增益效应或爱默生效应。
希尔反应及希尔氧化剂:在有适当的电子受体存在的条件下,离体的叶绿体在光下使水分解,有氧的释放和电子受体的还原,这一过程是Hill 在1937 年发现的,故称Hill 反应,在希尔反应中接受氢的受体称希尔氧化剂。
P 700 是PSI 的反应中心色素分子,即原初电子供体,是由两个叶绿素a 分子组成的二聚体。
这里P 代表色素,700 则代表P 氧化时其吸收光谱中变化最大的波长位置是近700nm 处,也即用氧化态吸收光谱与还原态吸收光谱间的最大处的波长位置来作为反应中心色素的标志。
PQ 穿梭:伴随着PQ 的氧化还原,可使2H + 从间质移至类囊体膜内的空间,即H+横渡类囊体膜,搬运2H+ 的同时也传递2e ,PQ 的这种氧化还原往复变化称为PQ 穿梭。
光合链由一系列能发生氧化还原作用的电子递体所组成的电子传递链,能将水光解所产生的电子依次传递,最后传递给NADP + 的总轨道。
光合磷酸化:叶绿体在光下与光合电子传递相偶联,将无机磷与ADP 转化成ATP 的过程,称之为光合磷酸化。
非环式光合磷酸化:与非环式电子传递相偶联产生的磷酸化反应。
有两个光系统参加,电子传递是单方向的,能引起水的光解放氧和ATP 与NADPH+ H+ 的生成。
环式光合磷酸化:与环式电子传递相偶联产生ATP 的反应。
只有光系统Ⅰ参加,电子流经过一个循环,不能形成NADPH+ H+ ,也不能引起水的光解放氧,只产生ATP 。
假环式光全磷酸化:与假环式电子传递相偶联产生ATP 的反应。
有两个光系统参加,电子传递是单方向的,能引起水的光解放氧和ATP 的生成,但不能生成NADPH+ H+ ,因为O 2 是电子和质子的最终受体。
C3 途径:与C3 植物C3 途径又称卡尔文循环,整个循环由RuBP 开始至RuBP 再生结束,共有14 步反应,均在叶绿体的基质中进行。
全过程分为羧化、还原、再生三个阶段。
因为二氧化碳固定后形成的第一个产物3-磷酸甘油酸为三碳化合物,所以称为C3 途径。
并把只具有C3 途径的植物称C3 植物。
C4 途径:与C4 植物又称为哈奇–斯莱克(Hatch-Slack )途径,整个循环有PEP 开始至PEP 再生结束,要经叶肉细胞和微管素鞘细胞两种细胞,循环反应因植物不同而有差异,但基本上可分为羧化、还原或转氨、脱羧和再生四个阶段,由于这条光合碳循环途径中二氧化碳固定后最初产物草酰乙酸为C4–二羧酸化合物,故称为C4 –二羧酸途径( C 4 dicarboxylic acidpathway ),简称C4 途径。
并把具有C4 途径的植物称为C4 植物。
Pi 运转器:存在于叶绿体被膜上,运转磷酸丙糖与Pi ,使二者对等交换,将磷酸丙糖转移到细胞质的专一载体。
光呼吸:绿色细胞只有在光下才能发生的吸收氧气释放二氧化碳的过程。
与光合作用有密切的关系,光呼吸的底物是乙醇酸,由于这种呼吸只有在光下才能进行,故称为光呼吸。
CAM 途径:和CAM 植物景天科、仙人掌科等科中的植物,夜间气孔张开,吸收CO2形成有机酸,贮藏在液泡中,白天气孔关闭,有机酸脱羧放出CO2进入卡尔文循环形成光合产物,这种与有机酸合成日变化有关的光合碳代谢途径称为景天科酸代谢途径。
把具有景天科酸代谢途径的植物称为CAM 植物。
光饱和现象与光饱和点:在较低光照强度下,光合速率随光照强度的增高几乎直线上长,当光照强度超过一定数值以后,光合速率增加转慢,如果再增加光照强度光合速率不再提高,这种现象称光饱和现象,开始达到光饱和现象时的光照强度称光饱和点。
光补偿点:在光饱和点以下,随着光照强度的减弱光合速率逐渐接近呼吸速率,当达到某一光强度时,光合作用吸收的二氧化碳和呼吸作用放出的二氧化碳达到动态平衡,此时的光照强度称光补偿点。
CO2饱和点:在一定范围内净光合速率随二氧化碳浓度的增加而增高,当达到一定程度时,再增加二氧化碳浓度,光合速率不再增加,此时二氧化碳浓度称二氧化碳饱和点。
CO2补偿点:在二氧化碳饱和点以下,光合作用吸收的二氧化碳与呼吸作用和光呼吸释放的二氧化碳,达到动态平衡,此时环境中的二氧化碳浓度,称二氧化碳补偿点。
光能利用率:是指作物光合产物中贮存的能量,占照射在单位地面上日光能的百分率。
荧光及荧光现象:光合色素分子的激发态电子,未被电子受体所接受,其电子从第一单线态回到基态时所发射的光称为荧光。
其叶绿素溶液在透射光下呈绿色,在反射光下呈红色,称荧光现象。
荧光产额发射荧光量子数与吸收光量子数的比值。
磷光及磷光现象:光合色素分子受光激发后,其电子从三线态回到基态时所发出的光称磷光。
当荧光出现后,立即中断光源,用灵敏的光学仪器还能看到短时间的“余辉”,这就是磷光现象。
光合生产率:又称净同化率,指生长植株的单位叶面积在一天内进行光合作用所形成的光合产物,减去呼吸和其它消耗之后,净积累的干物质量。
二、写出下列符号的中文名称PQ、PC、Fd、NADP+、RuBP、PGA、GAP、DHAP、FBP、F6P、G6P、E4P、SBP、S7P、R5P、Xu5P、Ru5P、PEP、CAM、TP、HP、OAA、CF1-CF0、PSⅠ、PSⅡ、BSC Mal、FNR、RubicoPQ :质体醌; PC :质蓝素;Fd :铁氧还蛋白; NADP + :氧化型辅酶Ⅱ;RuBP :核酮糖–1,5–二磷酸; PGA :3–磷酸甘油酸;GAP : 3–磷酸甘油醛; DHAP :磷酸二羟丙酮;FBP :果糖–1,6–二磷酸; F6P :果糖–6–磷酸;G6P :葡萄糖–6–磷酸; E4P :赤鲜糖–4–磷酸;SBP :景天庚酮糖–1,7–二磷酸; S7P :景天庚酮糖–7–磷酸;R5P :核糖–5–磷酸; Xu5P :木酮糖–5–磷酸;Ru5P :核酮糖–5–磷酸; PEP :磷酸烯醇式丙酮酸;CAM :景天科酸代谢; TP :磷酸丙糖;HP :磷酸己糖; OAA :草酰乙酰;CF1–CF0 :偶联因子( ATP 酶复合物); PS Ⅰ:光系统Ⅰ;PS Ⅱ:光系统Ⅱ; BSC :维管素鞘细胞;Mal :苹果酸; FNR :铁氧化蛋白– NADP + 还原酶Rubico :核酮糖–1,5–二磷酸羧化酶 / 加氧酶三、填空题1. 光合作用是一种氧化还原反应,在反应中(CO2)被还原,(H2O)被氧化。
2. 影响叶绿素生物合成的因素主要有:(光)、(温度)、(水)和(矿物质营养)。
3. 光合作用的三大阶段指的是(原初反应)、(电子传递和光合磷酸化)与(碳素同化)。
4. 光合作用分为(光)反应和(暗)反应两大步骤,从能量角度看,第一步完成了(光能向活跃化学能)的转变,第二步完成了(活跃化学能向稳定化学能)的转变。
5. 真正光合速率等于(表观光合速率)与(呼吸速率)之和。
6. 原初反应包括(光能吸收)、(传递)和(光能转化电能)三步反应,此过程发生在(类囊体膜)上。
7. 光反应是需光的过程,其实只有(光反应)过程需要光。
8. 光合磷酸化有下列三种类型:(非环式光合磷酸化)、(环式光合磷酸化)和(假环式光合磷酸),通常情况下以(非环式光合磷酸化)占主要地位。
9. 小麦和玉米同化二氧化碳的途径分别是( C3)和(C4 )途径,玉米最初固定二氧化碳的受体是(PEP ),催化该反应的酶是(PEP羧化酶),第一个产物是(草酰乙酸),进行的部位是在(叶肉)细胞。
小麦固定二氧化碳受体是( RuBP),催化该反应的酶是( RuBP羧化酶),第一个产物是(3–磷酸甘油酸),进行的部位是在(叶肉)细胞。
10. 光合作用中产生的 O2来源于(水)。
11. 50 年代由(卡尔文)等,利用(同位素示踪)和(纸谱色层分析)等方法,经过 10 年研究,提出了光合碳循环途径。
12. 光合作用中心包括(反应中心色素分子)、(原初电子供体)和(原初电子受体)。
13. PS Ⅰ的作用中心色素分子是(P 700 ), PS Ⅱ的反应中心色素分子是( P 680)。
14. 电子传递通过两个光系统进行, PS Ⅰ的吸收峰是( 700nm), PS Ⅱ的吸收峰是(680nm )。
15. 光合作用同化一分子二氧化碳,需要( 2)个 NADPH+H+,需要( 3)个 ATP ;形成一分子葡萄糖,需要(12 )个NADPH+H+,需要( 18)个 ATP 。
16. 光反应形成的同化力是(ATP )和(NADPH+H+)。
17. 光合作用电子传递途径中,最终电子供体是(水),最终电子受体是(NADP+)。
18. CAM 植物光合碳代谢的特点是夜间进行( CAM)途径,白天进行( C3)途径。
鉴别 CAM 植物的方法有(夜间气孔张开)和(夜间有机酸含量高)。
C3 植物的CO2补偿点是(50 μmol/mol 左右), C4植物的CO2补偿点是(0-5 μmol/mol),CO2补偿点低说明(PEP羧化酶对CO2的亲和能力强)。
19. 景天科酸代谢途径的植物,夜间吸收( CO2),形成草酰乙酸,进一步转变成苹果酸,贮藏在(液泡)中,白天再释放出( CO2),进入卡尔文循环,形成碳水化合物。
20. C4 植物在(叶肉)细胞中固定CO2,形成四碳化合物,在(维管束鞘)细胞中将CO2还原为碳水化合物。
21. C4植物同化CO2时 PEP 羧化酶催化( PEP)和(CO2 )生成(草酰乙酸)。