第七章原肠运动胚胎细胞重组
第八章原肠作用
卵黄囊
下胚层
羊膜腔 亨氏节
原沟
上胚层
下胚层
内胚层
第八章原肠作用 胚外中胚层
卵黄囊 中胚层
❖ 小鼠脊索形成方式有别于鸟类
(孔环),中央为卵黄拴第(八y章o原l肠k 作p用lug)
爪蟾的原肠作用的起 始和中胚层形成
原肠作用开始于瓶状细胞 的形成和内突。 它们的 内突使囊胚表面形成一个 小沟,即为胚孔 (blastopore)的雏形。同 时启动了细胞的内卷
边缘带
瓶状细胞
卵黄拴
下包
原肠
第八章原肠作用
集中延伸
内卷
爪蟾的原肠作用 (续)
赤道下方灰色新月区,产生狭缝状的内陷 ❖ 动物半球细胞下包 ❖ 背唇两侧扩展—侧第八唇章原、肠作腹用 唇;卵黄拴
2、内部变化:动物半球细胞下包、集中、内卷
❖ 受背唇处及周围区发散的信号吸引,动物半球 细胞下包(细胞数目增加以及不同细胞层 之间的合并)
❖ 下包细胞集中到背唇处由此卷入(索前板内胚层 和脊索)
❖ 进入的细胞朝前成片移动并沿整个囊胚内表面扩展形成原肠(内 卷、延伸是通过细胞黏附变化,并受位于前沿细胞形成的叶足支 持)
❖ 原肠腔产生并扩大,囊胚腔缩小 ❖ 侧唇产生(lateral lip):背唇向两侧扩展 ❖ 边缘带细胞迁移集中于此内卷,卷入的中胚层细胞沿
胚体腹部受卵黄大细胞牵拉,发生迁移,最终位于 原肠背侧部 ❖ 胚孔:两侧唇向腹面继续延伸而成腹唇,构成胚孔
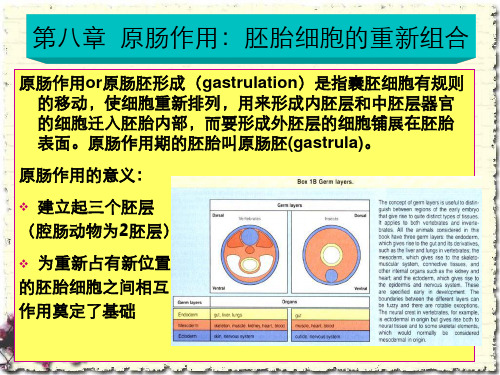
❖ 为重新占有新位置 的胚胎细胞之间相互 作用奠定了基础
第八章原肠作用
原肠作用中的主要细胞迁移
❖ 内陷(inva鱼
❖ 内移(ingression):胚胎 表面细胞单个地向内部迁移, 如水螅
发育名解 大题(原)

1、动物极:成熟卵子含卵黄较少的一端。
在多数动物中,动物极一般向上,为细胞核所在,原生质比较集中,卵裂进行比较迅速,将来形成为外胚层。
2、植物极:成熟卵子含卵黄较多的一端。
在多数动物中,动物极一般向下,原生质比较少,卵裂进行比较缓慢,将来形成内胚层。
3、镶嵌型发育:如果在发育早期将一个特定裂球从整体胚胎上分离下来,他就会形成如同其在整体胚胎中将会形成的结构一样的组织,而胚胎其余部分形成的组织会缺乏分离裂球所能产生的结构,两者恰好相补。
这种以细胞自主特化为特点的胚胎发育模式称为镶嵌型发育。
4、调整型发育:对细胞进行渐进特化的胚胎来说,如果在发育早期将一个分裂球从整体胚胎上分离下来,剩余胚胎中某些细胞可以改变发育命运,填补分离掉的裂球所留下的空缺,仍形成一个正常的胚胎。
这种以细胞渐进特化为特点的胚胎发育模式称为调整型发育。
5、促成熟因子(MPF):促使卵母细胞恢复减速分裂的因子,由调节亚基CyclinB和催化亚基P34cdc2组成。
6、细胞迁移:是原肠作用中的的一种细胞运动,细胞在内陷或内卷后开始在囊胚腔壁向动物极方向迁移。
分两种,单个细胞的迁移和细胞团或细胞层的整体迁移。
7、中期囊胚转换:果蝇、爪蟾等非哺乳类的早期受精卵呈转录抑制状态,经一系列快速分裂周期发育至中期囊胚阶段后才启动转录活动,通常都伴有细胞分裂突然变慢、失去同步化及细胞表型发生改变,母型控制转为合子型控制,即MBT中期囊胚转换。
8、体节:当原条退化,神经褶开始向胚胎合拢时,轴旁中胚层被分割成一团团细胞块,称作体节。
9、母体效应基因:在卵子发生过程中表达,并在卵子发生及早期胚胎发育中具有特定功能的基因称为母体效应基因。
10、生殖质:定位于卵质的特殊区域,是决定原始生殖细胞形成和发育的特化的卵质决定因子,主要由蛋白质和RNA组成,呈颗粒状结构,在卵裂中拥有生殖质的卵裂球可以分化为原生殖细胞。
11、囊胚:细胞数目增加,胚体中空而形成一个基本为球形的囊状结构.12、胚轴:指从胚胎前端到后端之间的前-后轴和背侧到腹侧之间的背-腹轴,两侧对称动物还具有中侧轴。
3.3 胚胎的早期发育--原肠胚

囊胚腔
爪蟾的晚期囊胚,示边缘区被内胚层细胞覆盖。
瓶状细胞
胚孔处形成瓶状细胞
胚孔
胚孔的定位
正常情况下,由精子驱动的卵细胞内部细胞质的重新排列 能够引起亚细胞因子(subcellular factor)的不对称分布。
受精卵内亚细胞因子的不对称分布产生了背腹区分,并最 终将胚孔的位臵确定在精子入卵点对面的一组植物极裂球 上。
咽内胚层的头部中胚层前 体
展开的瓶状细胞
表层IMZ向动物极迁移
细胞插入混合
深层迁移带细胞变平,内表面细胞形成原肠壁。
原肠期特征:一孔,二腔,三胚层。
四、 果蝇的原肠胚发育
/programs/view/psAk2 Ax7144/
果蝇胚胎的表面卵裂
2.5h 约6000个细胞
细胞质的运动激活精子入卵点对面的细胞质开启原肠胚发育。
通过机械方法(如离心)人为地改变皮层细胞质和皮层下 细胞质的空间关系,将可以使胚胎在精子入卵点的同一侧 开启原肠胚发育。 将精子决定的细胞质旋转与人工诱导的细胞质旋转相结合, 就可能获得两个原肠作用起点,最终形成联体双胞胎蝌蚪 (Black and Gerhart, 1985)。
受精 背部DSH富集
皮层旋转
背部GSK-3抑制
β –CATENIN降解
β –CATENIN稳定
DSH将β –CATENIN定位于胚胎背部的机 制
背部β –CATENIN富集
核里无β –CATENIN
核里有β –CATENIN
DSH将β –CATENIN定位于胚胎背部的机制
卷曲蛋白:wnt信号通 路的受体
植物极细胞负责形成胚胎背唇和启动原肠发生
推论:
发育生物学试题库

发育生物学试题库(发育生物学教学组)目录:第一章章节知识点与重点 (1)第二章发育生物学试题总汇 (6)第三章试题参考答案 (14)第一章章节知识点与重点绪论1.发育和发育生物学2.发育的功能3.发育生物学的基础4.动物发育的主要特点5.胚胎发育的类型(嵌合型、调整型)6.研究发育生物学的主要方法第一章细胞命运的决定1.细胞分化2.细胞定型及其时相(特化、决定)3.细胞定型的两种方式与其特点(自主特化、有条件特化)4.胚胎发育的两种方式与其特点(镶嵌型发育依赖型发育)5.形态决定子6.胞质定域(海胆、软体动物、线虫)7.形态决定子的性质8.细胞命运渐进特化的系列实验9.双梯度模型10.诱导11.胚胎诱导第二章细胞分化的分子机制1.细胞表型分类2.差异基因表达的源由3.了解基因表达各水平的一般调控机制第三章转录后的调控1.RNA加工水平调控2.翻译和翻译后水平调控第四章发育中的信号传导1.信号传导2.了解参与早期胚胎发育的细胞外信号传导途径第五章受精的机制1.受精2.受精的主要过程及相关知识3.向化性4.顶体反应5.皮质反应第六章卵裂1.卵裂特点(课堂作业)2.卵裂方式3.两栖类、哺乳类、鱼类、昆虫的卵裂过程及特点4.(果蝇)卵裂的调控机制第七章原肠作用1.了解原肠作用的方式:2.海胆、文昌鱼、鱼类、两栖类、鸟类、哺乳类的原肠作用基本过程与特点第八章神经胚和三胚层分化1.三个胚层的发育命运第九章胚胎细胞相互作用-诱导1.胚胎诱导和自动神经化、自动中胚层化2.胚胎诱导、异源诱导者3.初级诱导和次级诱导、三(多)级诱导4.邻近组织相互作用的两种类型5.间质与上皮(腺上皮)的相互作用及机制第十章胚轴形成1.体形模式2.图式形成3.果蝇形体模式建立过程中沿前后轴不同层次基因的表达4.果蝇前后轴建立的分子机制5.果蝇背腹轴形成的分子机制第十一章脊椎动物胚轴的形成1.什么是胚轴2.两栖类胚轴形成过程及分子机制3.了解鸟类、鱼类、哺乳类动物胚轴形成过程及分子机制第十二章脊椎动物中枢神经系统和体节形成机制1.脊椎动物中枢神经系统的前后轴形成2.脊椎动物中枢神经系统的背腹轴形成3.脊椎动物体节分化特征第十三章神经系统的发育1.神经系统的组织发生神经系统的组成来源(神经管、神经嵴、外胚层板)中枢神经系统的组织发生(脊髓、大脑、小脑、核团)神经系统发生过程中的组织与调控(位置、数目)2.神经系统的功能建立3.神经突起(树突和轴突)4.局部有序投射5.突触第十四章附肢的发育和再生1.附肢的起源2.附肢的早期发育附肢发育中外胚层与中胚层的相互作用附肢发育中轴性建立3.附肢再生(再生过程、再生调节)第十五章眼的发育1.视泡发育、分化2.晶状体发育、分化3.晶状体再生4.角膜发育第十六章变态1.变态2.昆虫变态的激素调控3.两栖类变态的激素调控第十七章性腺发育和性别决定1.哺乳动物的性腺发育2.哺乳动物的性别决定3.果蝇的性别决定4.雌雄同体、环境性别决定第十八章生殖细胞的发生1.精子发生:特点,过程2.卵子发生:特点,过程第十九章干细胞1.干细胞2.干细胞分类3.了解干细胞的应用第二十章动物发育的环境调控1.发育与环境关系2.环境对正常发育的调控3.环境对正常发育的干扰4.遗传与环境之间的相互作用第二章发育生物学试题样题总汇一、填空题(每空1分)1.发育生物学研究的主要内容是和,其主要任务是研究生命体发育的及其机制。
海胆原肠作用

总结海胆原肠作用过程及机制:原植物极中央细胞内陷进入囊 胚腔,表皮细胞转变成为初级间质细胞(伪足纤丝收缩促进细 胞内陷),然后内胚层表皮细胞内陷和扩展,其前端表皮细胞 转化为次级间质细胞(伪足收缩起定向和驱动细胞移动的作用, 细胞会聚伸展)。第二节 两栖类的源自肠作用一、爪蟾囊胚的发育命运图
• 两栖类的囊胚的基本任务:三胚层建成 • 研究方法:活体染色技术 • 非洲爪蟾的中胚层前体只存在于深层细胞中,位 于赤道区域表面细胞下面,而外胚层和内胚层则 来源于表面的表层细胞中。
细胞粘连作用的变化导致初级间质的内移 初级间质细胞和透明层 以及相邻裂球之间的亲和力减少,而与基质片层之间的亲和力增加。 相比之下,非间质细胞和透明层以及相邻裂球之间的亲和力维持不 变。
• 内移至囊胚腔的初级间质细胞被大量胞外 片层物质包绕。 • 沿动植物极轴分布的纤维决定了间质细胞 迁移的方向。 • 囊胚腔胞外基质纤维与动植物极轴平行排 原肠作用过程中沿动植物极轴分布的纤维决定 列,最终与初级间质细胞发生联系。 了初级间质细胞的迁移方向
原肠作用概述 1.概念:原肠作用(gastrulation)是卵裂后期囊胚细胞剧
烈的、高速有序的运动过程,从而实现细胞的重新组合形 成内中外三个胚层。 2.功能: • 囊胚细胞位置发生变动形成三胚层。原肠作用是从尚未分 化到分化为三个胚层和器官原基决定的关键时期。 • 动物身体的主轴(包括前后轴、背腹轴相互作用奠定了基 础。 3.细胞的运动方式 原肠作用过程中的细胞运动涉及整个胚胎。
机制
植物极内陷的双金属片模型 植物极板细胞分泌硫酸软骨素蛋白多 糖(CSPG)到透明层内层中,CSPG吸水膨胀,导致透明层弯曲, 相连的细胞层内陷。
2.晚期原肠内陷
• 早期原肠内陷完成之后,经过短暂停歇, 原肠大幅度拉长, 短粗的原肠变成又细又 长的管状结构。 • 在此期间没有新细胞形成,原肠的拉长过 程是通过细胞重排实现的,原肠周长内细 胞数目大为减少。
发育生物学

<<发育生物学>>绪论•发育是遗传信息按一定的时间和空间顺序表达的结果。
发育具有严格的次序性,发育并不是个别基因的表达,而是众多基因表达在时间和空间上的联系和配合。
•发育生物学的主要任务是研究生物体发育的遗传程序及其调控机制。
后成论与先成论的持久论争希腊哲学家Aristotle在公元前第4世纪在对鸡胚和一些无脊椎动物胚胎观察后提出胚胎发育的两种假设:Preformation:生物个体的一切组成部分都早就存在于胚胎中,各个部分随着胚胎的发育而长大。
Epigenesis: 在胚胎的发育过程中,各种结构是逐渐形成的。
Part 5. Investigative Techniques一、常规发育生物学研究技术显微技术活样品的精细结构组织切片技术胚胎内部结构分子生物学技术基因与蛋白质的表达原位杂交技术特定mRNA的表达显微操作技术基因功能报道基因技术不易检测的基因表达细胞标记技术细胞谱系染料细胞标记荧光细胞标记细胞移植二、发育遗传学技术⏹正向遗传学手段通过大规模随机诱变,产生发育异常的突变个体,再寻找突变基因。
⏹反向遗传学手段通过功能丧失或功能获得方法对已知基因进行功能研究。
1.正向遗传学技术诱变筛选技术、插入诱变筛选、突变基因的克隆2. 反向遗传学技术基因敲除技术、条件基因敲除技术、干扰技术、显性技术、转基因技术、克隆化分析第一章细胞命运的的决定一、细胞定型和分化从单个全能的受精卵产生各种类型细胞的发育过程叫细胞分化(cell differentiation)。
已分化的细胞不但具有一定的形态和合成特异的产物,而且行使特异的功能。
定型(commitment)。
Slack(1991)建议将定型分为特化(specification)和决定(个时相。
胚胎细胞的定型有两种主要方式:•第一种方式的细胞定型是通过胞质隔离(cytoplasmic segregation)来实现的。
卵裂时,受精细胞,细胞命运的决定与临近的细胞无关。
《发育生物学》课程笔记

《发育生物学》课程笔记第一章:发育生物学的概述一、发育生物学的定义和研究范围1. 定义:发育生物学是生物学的一个分支,它专注于研究生物体从单个细胞(通常是受精卵)开始,经过细胞分裂、分化、形态发生、组织形成和器官发育等过程,最终形成成熟个体的全部生物学过程。
2. 研究范围:- 细胞层面的发育:包括细胞周期、细胞分裂、细胞命运决定、细胞迁移等。
- 分子层面的发育:涉及基因表达调控、信号转导途径、转录因子和网络调控等。
- 形态发生和器官形成:研究生物体的形态变化、轴的形成、器官原基的诱导和分化等。
- 发育过程中的遗传和环境因素:探讨遗传变异、表观遗传学、环境因素如何影响发育过程。
- 发育异常和疾病:研究发育过程中的异常如何导致疾病和畸形。
二、发育生物学的发展历程1. 早期探索(17世纪- 19世纪):- 显微镜的发明使得科学家能够观察胚胎的早期发育。
- 卡尔·冯·林奈(Carl Linnaeus)和卡尔·恩斯特·冯·贝尔(Karl Ernst von Baer)等人的工作奠定了胚胎学的基础。
2. 胚胎学时期(19世纪末- 20世纪初):- 柏拉图生物学假说和重演论(Recapitulation theory)的提出。
- 奥古斯特·魏斯曼(August Weismann)提出了种质论,区分了体细胞和生殖细胞。
3. 细胞和分子生物学时期(20世纪中叶- 至今):- 发现DNA双螺旋结构,开启了分子生物学时代。
- 发育遗传学的发展,如同源框(homeobox)基因的发现。
- 克隆技术的应用,如克隆羊多莉(Dolly)的诞生。
三、发育生物学与其他学科的关系1. 与胚胎学的关系:- 发育生物学是胚胎学的延伸,两者都关注生物体的早期发育,但发育生物学更侧重于分子和细胞机制。
2. 与遗传学的关系:- 遗传学提供了理解发育过程中基因如何传递和表达的基础。
- 发育遗传学领域的研究揭示了基因如何控制发育过程。
发育生物学复习题

发育生物学复习题名词解释1、发育生物学(developmental biology)研究多细胞生物体从精子与卵子的发生、受精、发育、生长到衰老的变化过程及机理的科学。
2、镶嵌发育如果在发育早期将一个特定裂球从整体胚胎上分离下来,他就会形成如同其在整体胚胎中将会形成的结构一样的组织,而胚胎其余部分形成的组织会缺乏分离裂球所能产生的结构,两者恰好相补。
这种以细胞自主特化为特点的胚胎发育模式称为镶嵌型发育。
如:栉水母、海鞘、环节动物、线虫、软体动物。
3、调整型发育对细胞进行有条件特化的胚胎来说,如果在发育早期将一个分裂球从整体胚胎上分离下来,剩余胚胎中某些细胞可以改变发育命运,填补分离掉的裂球所留下的空缺,仍形成一个正常的胚胎。
这种以细胞有条件特化为特点的胚胎发育模式称为调整型发育。
4、胚胎诱导动物在一定的胚胎发育时期, 一部分细胞影响相邻细胞使其向一定方向分化的作用称为胚胎诱导。
5、图式形成:发育过程中控制胚胎细胞的行为使其在正确的空间位置上形成特定的结构。
6、发育生物学的发展基础及过程如何?研究哪些问题?胚胎学、遗传学、细胞生物学主要研究多细胞生物题从生殖细胞的发生、受精、胚胎发育、生长到衰老死亡,即生物个体发育中生命现象发展的机制。
同时还研究生物种群体统发生的机制。
7. 精细胞形成精子阶段涉及哪些主要事件?1)顶体泡的形成(高尔基体)2)鞭毛(中心体微观复合)3)多余的细胞质排出,细胞核高度浓缩(核小体组蛋白被鱼精蛋白替代以防止外界环境对DNA的损伤)4)细胞膜上有受精蛋白的形成(精膜上与卵子结合蛋白产生)5)线粒体鞘(大量线粒体)细胞分化(cell differentiation)从单个的全能细胞受精卵开始产生各种分化类型细胞的发育过程叫细胞分化。
3、形态发生决定子(morphogenetic determinant)也称成形素或胞质决定子,是存在于卵细胞质中的特殊物质,能够制定细胞朝一定方向分化,形成特定组织结构。
发育生物学总练习题

练习题参考答案第一章、绪论1.名词解释:发育development——指生命现象的发展,生物有机体的自我构建和自我组织;发育生物学developmental biology——是应用现代生物学的技术研究生物发育机制的科学;形态发生morphogenesis——不同表型的细胞构成组织、器官,建立结构的过程;2.发育生物学有哪些主要研究内容答:主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老死亡,即生物个体发育中生命现象发展的机制;同时还研究生物种群系统发生的机制;3.分子生物学的兴起,对发育生物学的发展有何影响答:Watson和Crick1953提出DNA分子的双螺旋模型以后,分子生物学迅速发展,发育生物学的发展也从此揭开新的序幕;人们认识到发育主要受遗传物质DNA的控制,为回答编码在DNA上的遗传信息及其所表达的蛋白质如何控制生物体的发育等问题,人们开始采用分子生物学技术和各种其他新兴的生物学技术,进行生物发育机制的研究;取了得一系列重大成果,不仅使传统理论进一步深化,而且形成了不少新的观点和理论;由于发育生物的迅速发展,现已成为生命科学的前沿和热点领域之一;4.简述发育生物学的发展简史.答:发育生物学是由分子生物学、细胞生物学、遗传学及生物化学等学科和胚胎学的相互渗透发展和形成的一门新兴的生命科学;胚胎学的发展很久远,二千多年前,Aristotle提出胚胎是由简单到复杂逐步发育形成的后成论观点;但到了17世纪后期,由于宗教统治的禁锢,先成论占统治地位,既认为胚胎是成体的雏形预先存在精子或卵子中;1759年德国科学家Wolf 根据对鸡胚发育的仔细观察,再次提出后成论观点,到19世纪才普遍为人们所接受;1839年,Schleiden和Schwann提出细胞理论,对胚胎发育的概念是划时代影响;认识到细胞核在发育中的重要性,1880年代,Weismann提出“种质学说”在当时影响很大,强调早期卵裂是不对称分裂;但Driesch1891证明海胆二细胞期的细胞发育没有区别;1924年,Spemann 进行了著名的胚胎移植实验,人们才真正认识到,细胞之间的相互作用是胚胎发育最重要的核心问题;1900年,Mendel遗传规律的重新发现,胚胎学与遗传学结合,认识到发育受基因型的控制,但环境也影响发育;Watson和Crick1953提出DNA分子的双螺旋模型以后,开启了现代意义上发育生物学;重点是阐明发育的分子控制机制,在一些模式动物如果蝇、线虫等已取得一系列重大的突破;第二章配子发生1、说明线虫和果蝇的生殖细胞的决定;答:线虫未受精卵的细胞质均匀分布一种P颗粒,受精后集中到后部;受精卵经过4次分裂,P 颗粒集中到一个P4细胞;P4是所有生殖细胞的祖细胞;果蝇的生殖质是位于受精卵后端极质颗粒;受精卵核经过9次分裂,后部形成5个包含极质颗粒的极细胞,极细胞分化为原生殖细胞;2、精子形成过程中经历了哪些变化答:精子细胞形成后,经过一系列分化,变态为特殊形状的精子;1、细胞核中的染色质高度浓缩,使核体积大大减少;其中核蛋白由组蛋白变为精蛋白;2、细胞质大多被抛弃;其中中心粒演变为轴丝,产生精子鞭毛;高尔基体形成顶体;线粒体形成线粒体鞘;3、形状变成流线型,便于运动;3、卵子发生过程与精子发生过程有哪些异同答:相同点:1、都要经过减速分裂,使配子的染色体减半;2都要经过增殖期、分裂期和成熟期;不同点对比如下:1、精子发生过程中的生长期不很明显,而卵子发生过程中的生长期则特别长;因此精子发生的结果是产生体积微小的精子,而卵子发生的结果是产生体型很大的卵子;2、精子发生速度比卵子快,而且精原细胞则可以在成熟期内不断增殖;所以成熟精子的数目大大超过成熟卵子的数目;3、每个初级精母细胞最后变成4个大小相等的精子;而每个初级卵母细胞只能产生1个大的成熟卵和3个体积很小不能受精的极体;4、精子发生过程要经过变态期,才能从精细胞转变为精子;而卵子发生没有这一时期;5、精子发生过程中的两次成熟分裂全部在精巢内进行,卵子发生过程中的两次成熟分裂可在卵巢内也可在卵巢外进行;4、什么是母体效应基因举列说明;答:在卵子发生过程中表达并在早期胚胎发育中起作用的基因,称为母体效应基因;如Bicoid 基因是果蝇一个重要的母体效应基因,Bicoid基因在滋养细胞中转录,其mRNA定位并储存在卵子的前端;受精后翻译出的蛋白质沿前—后轴扩散,形成浓度梯度,为胚胎的早期分化提供位置信息;第三章受精的机制1、名词解释促成熟因子MPF——促使卵母细胞恢复减速分裂的因子,由调节亚基CyclinB和催化亚基cdc2组成;精子获能capacitation——从雄体直接采集的哺乳动物的精子不能受精,只有在雌性生殖道的适宜环境中发生许多生理、生化方面的变化后,才能获得受精的能力,这个过程称为精子获能;顶体反应acrosomal reaction——当精子遇到卵子时,顶体膜和其外的质膜发生多处融合,释放蛋白水解酶,消化透明带或卵膜,为精子进入卵子打开通道,这些就是顶体反应;皮层反应cortex reaction——当精卵质膜融合后,皮层颗粒与其外的质膜融合,导致其内含物释放到质膜和卵黄膜或透明带之间的卵周隙内,这种现象就是皮层反应;2、海胆的精子和鱼类的卵处于同一环境中,二者是否可以受精为什么答:不可以受精;因为海胆精子顶体的突起上具有结合素bindin,而卵黄膜上存在着种类特异性的结合素受体;结合素能特异地结合到同种动物的卵黄膜上;这样精子结合素和其受体之间相互作用的专一性就阻止了种与种之间的杂交受精;3、卵子为什么要阻止多精受精举例说明其怎样阻止多精受精的答:因为多精受精造成细胞分裂时的染色体紊乱,最后导致死亡或不正常发育,所以卵子要阻止多个精子进入卵子;阻止多精受精的机制有二方面,1、初级、快速阻断:通过膜电位的改变,使卵子上精子结合的受体失活;如海胆的第一个精子与卵质膜结合后的1-3秒内,因钠离子的流入而导致膜电位的迅速升高,从而阻止其它精子与卵膜的结合;2、次级、永久阻断:通过皮层反应,使受精膜迅速膨胀;如海胆卵受精后20-60秒内,质膜下的皮质颗粒与质膜融合,释放其内含物形成受精膜,阻止其它精子的进入;第四章卵裂1、名词解释经线裂meridional cleavage——指卵裂面与A-V轴平行的卵裂方式;表面卵裂superficial cleavage——昆虫受精卵的大量卵黄位于卵的中央,卵裂被限制在卵的外围卵质中,为表面卵裂;紧密化compaction——哺乳动物在第三次卵裂后,形成的卵裂球突然挤在一起,卵裂球之间的接触面增大,形成一个紧密的细胞球体而把球的内部封闭起来;胚胎干细胞ESC——胚胎干细胞是一种高度未分化细胞;它具有发育的全能性,能分化出成体动物的所有组织和器官;当受精卵分裂发育成囊胚时,内层细胞团Inner Cell Mass的细胞即为胚胎干细胞;2、早期卵裂与一般的细胞分裂有什么重要不同分子机制如何答:早期卵裂一般只有S期和M期,无生长期;因此分裂周期短,速度快,卵裂时胚胎的体积不增大;受精后早胚细胞分裂的都是由贮存在卵内的母型mRNAs和蛋白质控制的;卵裂周期受成熟促进因子MPF的控制;早期的卵裂所需要的有活性的MPF已存在在卵质中或由母型mRNA翻译,不需要启动合子基因,因此速度很快;3、卵裂有那些主要类型说明与卵黄的关系并举例代表动物;答:卵裂主要可分为完全卵裂和不完全卵裂;卵黄对卵裂有一定的阻抑作用,因此卵黄的含量和分布影响卵裂方式,含卵黄少的受精卵均黄卵和中黄卵的卵裂为完全卵裂,如哺乳动物和两栖类;卵黄含量高的的受精卵采用偏裂即不完全卵裂,如鱼类和鸟类的盘状裂、昆虫的表面卵裂;4、分析节肢动物与哺乳动物早期卵裂过程表现出对未来发育的设定;答:果蝇受精卵核经过9次分裂后,出现生殖细胞的决定,后部形成5个极细胞,极细胞分化为原生殖细胞;哺乳动物8细胞期的细胞是等能的;紧密化后产生16个细胞的桑胚椹,桑椹胚内部有1-2个细胞与外界隔离,属于内细胞团,将来形成胚胎组织;外部细胞分裂产生滋养层细胞,不参与形成胚胎组织,而参与形成绒毛膜组织;第五章原肠作用:胚胎细胞的重新组合1、名词解释原肠作用gastrulation——原肠作用是通过剧烈地有序的细胞运动,使囊胚细胞重新组合,形成三个胚层的胚胎结构的过程;中囊胚转换midblastula transition——在囊胚中期由母型调控向合子型调控的过渡称为中囊胚转换;“Nieuwkoop”中心——在两栖类囊胚中,最背部的植物极细胞能够诱导产生Spemann组织者,称为“Nieuwkoop中心”;绒毛膜chorin——合胞体滋养层和富含血管的中胚层共同构成的器官称为绒毛膜;2、简述原肠作用的细胞运动方式;答:原肠作用的细胞运动方式可概括为6种:1、外包:表层细胞运动,细胞铺展、变薄、面积扩大,包住胚胎内层的细胞;2、内陷:某个区域的细胞同时向内凹入形成凹陷;3、内卷:外面铺展的细胞连续从边缘向胚胎内部卷入,并沿细胞内表面扩展形;4、内移:细胞从胚胎表层单个的向胚胎内部迁移;5、分层:单层细胞被割裂成两层或多层平行的细胞层;6、会聚伸展:指细胞间相互插入,使所在组织变窄、变薄,并推动组织向一定方向移动;3、在两栖类原肠作用过程中,三个胚层是怎样形成的答:在原肠作用过程中,动物极帽和非内卷边缘带细胞通过外包扩展,覆盖整个胚胎,形成外胚层;中胚层细胞开始内卷时,表层内卷边缘带细胞与其内侧的脊索中胚层一起移动,形成了原肠腔顶部的内胚层壁;胚孔下面的植物极细胞被外包的非内卷边缘带细胞覆盖,形成原肠腔底部的内胚层;深层内卷边缘带是一个细胞环;在原肠作用过程中,该细胞环沿胚孔唇内卷;预定脊索从背唇内卷,体节中胚层从侧唇卷入,未来的侧板中胚层从腹唇卷入;4、比较两栖类和鸟类的原肠作用过程,说明背唇与原条在发育地位的同一性;答:鸟类原条与两栖类背唇在发育地位的同一性可用以下几点说明;1、两栖类动物背唇和胚孔的出现是原肠作用开始的标志,鸟类的原肠的形成是以原条出现为标志;2、原条前端的亨氏节与背唇一样都是原肠作用的组织者,可以发动形成次生胚胎;3、原条上的原沟和胚唇周围的胚孔同样是外层细胞进入囊胚腔的门户;4、通过背唇内卷进入胚胎的细胞发育为头部中胚层和脊索,同样从原条亨氏结迁移到囊胚腔的细胞是未来的头部中胚层和脊索;5、胎盘是怎样形成的,它有什么功能答:合胞体滋养层和富含血管的中胚层共同构成绒毛膜,绒毛膜和子宫壁融合形成胎盘;胎盘既含有母体成分,又含有胎儿成分;胎盘负责胎儿的物质交换,胎儿通过胎盘从母血中获得营养和氧气,排出代谢产物和二氧化碳,相当于小肠、肺和肾的作用;第六章神经胚和三胚层分化1、什么是神经胚初级神经胚的形成过程答:正在进行神经管形成的胚胎称为为神经胚;初级神经胚的形成过程如下:①中线处的预定的神经外胚层细胞变长加厚,形成神经板;②神经板边缘加厚,并向上翘起,形成神经褶,神经板中央出现“U”形神经沟;③神经褶向胚胎背中线迁移,最终合拢形成神经管,上面覆盖着外胚层;④神经管最靠背面细胞变成神经嵴细胞;2、脊椎动物中胚层在发育中形成哪5个过渡性的区域性结构答:神经胚中胚层可分为5个过渡性区域;第一个是脊索中胚层,将来形成脊索;第二是背部体节中胚层,主要形成体节;第三个是居间中胚层,将来形成泌尿系统和生殖管道;第四个是侧板中胚层,形成心脏、血管、血细胞和胚胎外膜等;最后是头部中胚层,将来形成面部的结缔组织和肌肉;3、轴旁中胚层什么基因的表达对脊椎动物体节形成有重要关系答:在轴旁中胚层的前端首先出现Notch1和Paraxis基因的表达;由于Notch1和Paraxis 基因产物的作用,轴旁中胚层细胞开始合成并分泌fibronectin和N-cadherin,创造了体节结构发生的条件;4、内胚层主要形成哪些器官或组织答:胚胎内胚层的功能是构建体内两根管道消化管和呼吸道的内表皮;第一根管道是贯穿于身体全长的消化管,肝、胆囊和胰腺由消化管凸出形成;第二根管道是呼吸管,由消化管向外生长形成,包括气管、支气管和肺;咽和咽囊也是内胚层来源,咽和咽囊向外凸起产生扁桃体、甲状腺、胸腺和甲状旁腺,水生脊椎动物中,咽囊产生鳃;第七章细胞命运的决定——细胞的自主特化1、名词解释细胞分化cell differentiation——从单细胞受精卵产生有机体的各种形态和功能细胞的发育过程叫细胞分化;镶嵌型发育mosaic development——如果将一个发育早期胚胎的某一个卵裂球去掉,则这个胚胎将会发育为一个不完整的胚胎,而缺失的部分刚好就是所移走的卵裂球所能发育的结构;这种以细胞自主特化为特点的发育模式称为镶嵌型发育;调整型发育regulativ development——如果移去早期胚胎的一个卵裂球,胚胎的剩余的部分则可改变它们正常的发育命运,来填补移去的卵裂球所留下的空白,仍形成一个完整的胚胎;这种以细胞有条件特化为特点的胚胎发育模式称为调整型发育;胞质定域cytoplasmic localization——在镶嵌型发育中,形态发生决定子被定位于特定卵质区域的,并在卵裂时分配到特定的裂球中,决定裂球的发育命运;这一现象称为胞质定域;2、简述海鞘8裂球的细胞发育命运;答:海鞘第一卵裂面与幼虫两侧对称面吻合,所以左右两裂球的发育完全对称;在8细胞期可将胚胎裂球分为4对,每对裂球发育命运如同卵子发育命运图谱一致;动物极二对裂球发育为外胚层,植物极后面的一对裂球形成内胚层、间质和肌肉组织,植物极前面的一对裂球形成脊索和内胚层;神经细胞是从动物极前面的一对裂球和植物极前面的一对裂球产生的;3、试述果蝇的极质决定子的分子基础;答:果蝇卵极质主要由蛋白质和RNA组成;生殖质的组分之一是gclgerm cell-less基因转录的mRNA,gcl基因在果蝇卵巢的营养细胞中转录,所转录的mRNA通过环管转运至卵子中,定位于称为极质的细胞质中,是极细胞形成所必不可少的成分;极质决定子也可能是Nanos蛋白,如果缺乏Nanos蛋白,胚胎的极细胞就不能迁移到生殖腺中,不能产生生殖细胞;生殖细胞决定子还可能是线粒体大核糖体RNAmtlrRNA;第八章细胞命运的渐进特化——胚胎细胞相互作用1、说明相嵌型发育和调整型发育与Weismann的“种质说学”的调和与矛盾;答:Weismann的“种质说学”主要论点为染色体是由各种能决定细胞发育命运的核决定子组成的;受精卵分裂时,不同的核决定子在胚胎发育过程中分配到不同的细胞内,由此决定细胞的命运,使其发育成身体的某一部分;在相嵌型发育中,如果将一个发育早期胚胎的某一个卵裂球去掉,则这个胚胎将会发育为一个不完整的胚胎,而缺失的部分刚好就是所移走的卵裂球所能发育的结构,这种发育现象是与Weismann上述观点是相调和的;但在调整型发育中,如果移去早期胚胎的一个卵裂球,胚胎的剩余的部分则可改变它们正常的发育命运,来填补移去的卵裂球所留下的空白,仍形成一个完整的胚胎,这种发育现象显然与Weismann的“种质说学”相矛盾;2、什么是Spemann组织者哪些动物具有这类的组织者答:两栖类早期胚胎胚孔背唇能诱发原肠作用,组织次生胚胎的形成,即为Spemann 组织者;鸟类和哺乳动物中的原条前端的亨氏结是类似组织者,鱼类的胚盾也是这样的组织者;3、简述goosecoid基因的作用;答:goosecoid基因具组织者特异性,编码一种DNA结合蛋白;Goosecoid蛋白的功能主要表现在:1能激活背唇细胞的迁移特性内卷和延伸;2决定头部中胚层和背侧中胚层的发育命运;3gossecoid表达细胞动员周围细胞参入背轴形成;第九章果蝇胚轴形成1、什么是形态发生原morphogen举例说明;答:某些因子沿体轴的分布呈现浓度梯度,不同水平决定该区域的的反应,最终形成某种形态结构,这种因子即是形态发生原;例如果蝇的Bicoid蛋白,Bicoid蛋白在果蝇的合胞体胚胎中形成一个由前到后的浓度梯度,在不同的浓度阈值分别激活不同靶基因的表达,对果蝇头胸部的结构的决定起关键作用;2、为什么dorsal基因的突变会导致胚胎背部化,cactus基因突变会导致胚胎腹部化答:因为dorsal基因的突变会导致胚胎无Dorsal蛋白合成,导致合子基因snail和twist腹侧特征在所有细胞中都不表达,而背侧特征基因dpp、Zen、tolloid、zerknullt等在所有细胞中表达,从而导致胚胎背部化;相反,cactus基因突变会导致胚胎无Cactus蛋白,这样使Dorsal蛋白同样能进入背部的细胞核,活化合子腹侧特征基因twist和snail的表达,同时抑制背侧特征基因dpp和zen等基因的表达,这样导致胚胎腹部化;3、试述果蝇A—P轴形成的分子机理;答:果蝇A—P轴的形成首先是母体基因的作用,形成形态发生原梯度;形态发生原在A-P 轴线的不同区域激活不同的基因,使不同区域的基因活性谱不同而出现分化;调节果蝇胚胎前后轴的形成有4个非常重要的形态发生原:BICOIDBCD和HUNCHBACKHB调节胚胎前端结构的形成,NANOSNOS和CAUDALCDL调节胚胎后端结构的形成;另外,Torso基因编码一种细胞外信号分子受体蛋白,可能是末端形态发生原;有3类合子基因对体躯A—P轴的分节进行遗传调控:缺口基因gap genes、成对法则基因pair-rule genes和体节极性基因segment polarity genes;形态发生原首先调节缺口基因的表达,缺口基因表达区呈宽的带状,包括hunchback、kruppel和knirps的表达带;缺口基因再控制成对法则基因,成对法则基因每隔一个体节,以7条条纹的模式表达,如even-skipped、fushi tarazu 和hairy等;成对法则基因又控制体节极性基因,体节极性基因把不同体节再分成更小的条纹,划分出14个体节的分界线,如engrailed、wingless和hedgehog等;同时,缺口基因和成对法则基因编码的蛋白质调节同源异型基因的表达,最终决定身体体节将出现那一种类型;4、试述果蝇同源异型选择者基因的表达图式;答:果蝇同源异型选择者基因Homeotic selector genes是指在体节边界建立之后,用来控制每个体节的特征结构发育的基因;包括触角足复合体Antp-C,有5个基因:lab labial,pb proboscipedia,Dfd Deformed,Scr Sex comb reduced,Antp Antennapedia;另一个区域是双胸复合体BX-C,有3个基因:Ubx Ultrabithorax,abdA abdominal A和AbdB abdominal B基因;这两个复合体统称同源异型复合体HOM-C;这8个基因的表达图式总的来说是胚胎从前到后依次表达;Lab、Pb和Dfd基因参与头部体节的特化;Scr和Antp基因主要决定胸部体节的特征;Ubx与胸部体节的分化相关,abd-A与Abd-B负责腹部体节的分化;同源异型选择者基因突变可引起同源异型现象,例如Antp基因的显性突变体,使该基因在头部和胸部同样表达而使头部长出肢而不是触角;当Ubx缺乏时,第三胸节转变成第2胸节,形成具有4个翅的果蝇;第十章脊推动物胚轴形成1、简述两栖类动物胚轴的形成;答:两栖类胚胎的D-V轴和A-P轴是由受精时卵质的重新分布而决定的;受精时,由于精子入卵的影响,卵子皮质与卵黄在重力作用下相对移动,在精子入卵处的对面产生有色素差异的灰色新月区,由此标志预定胚胎的背侧;随着原肠胚的形成,精子进入的一侧发育成为胚胎的腹侧,相反的一侧发育为胚胎的背侧,在动物极附近的背侧形成头部,与其相反的一侧形成尾,从而形成胚胎的背腹轴和前后轴;左右轴随着脊索的形成而确定;2、试述Nieuwkoop中心作用的分子机理;答:在两栖类囊胚中最靠近背侧的一群植物极细胞,对组织者具有特殊的诱导能力,称为Nieuwkoop中心;β-catenin是Nieuwkoop中心的一个主要的细胞因子;β-catenin开始在整个胚胎中均有分布,但在腹侧细胞中因为含有糖原合成激酶3GSK-3而降解,而在背侧细胞中由于存在GSK-3的抑制因子DisheveledDSH蛋白, β-catenin不会被降解;β-catenin是转录因子,它与转录因子Tcf3结合形成β-catenin/Tcf3复合物,激活siamois基因在Nieuwkoop中心表达;Siamois蛋白与TGF- β基因家族蛋白产物Vg1,VegT和Nodal-相关基因编码蛋白Xnrs的协同作用使组织者特异基因goosecoid激活,诱导背部产生组织者活性;3、简述哺乳动物前—后轴形成过程的有关分子及其作用;答:哺乳动物胚胎的前—后轴的受两个信号中心调控,一个是前端的内脏内胚层A VE,另一个原条前端的亨氏节相当于两栖类的胚孔背唇,组织者;胎体远端的A VE细胞单独表达hex基因,将来仅仅形成前部,是前端位置的标志;A VE相反的后端的上胚层,cripto基因表达,原条开始形成;原条前端的亨氏节负责整个躯体的建立,包含有与蛙的组织者中发现的蛋白质,如GOOSECOID、NODAL、LIM-1和HNF3β;nodal基因对于原条的起始发育和维持发育是必须的,goosecoid基因的表达对于激活与头部形成的基因起关键作用,GOOSECOID、LIM-1和HNF3β蛋白对于前背部中胚层细胞的特化是必须的;哺乳动物前—后极性的特化都由hox 基因的表达进行调控;第十一章胚胎细胞相互作用——胚胎诱导1、名词解释胚胎诱导embryonic induction——胚胎一个区域对另一个区域发生影响,并使后者沿着一条新途径分化的过程称为胚胎诱导;诱导者inductor——发出信号,产生影响的一部分细胞或组织,称为诱导者;反应组织responding tissue——接受信号从而进行分化的细胞或组织称为感应者,或称反应组织,它们必须具有感应性competence才能接受诱导者的刺激;组织者organizer——能够诱导外胚层形成神经系统,并能和其他组织一起调整成为中轴器官的胚孔背唇部分;2、经典胚胎学所指的初级胚胎诱导与现在的概念的区别答:过去经典实验胚胎学的初级胚胎诱导实际上是神经诱导,是指原肠胚中预定的外胚层受脊索中胚层的诱导而形成神经板的过程;现在认为初级胚胎诱导应包括3个阶段:第一阶段发生在卵裂期,为中胚层的形成和分区;第二阶段即是脊索中胚层诱导背部外胚层转变为神经系统的神经诱导;第三阶段是中央神经系统的区域化;3、邻近组织相互作用的类型并举例说明;答:邻近组织相互作用可分为指导互作和容许互作;指导互作需要从诱导者发出诱导信号,才能启动反应细胞新基因的表达,没有诱导细胞,反应细胞就不能按特定的方式分化;例如表皮和间质真皮的相互作用属于指导互作,鸡皮肤结构的类型羽毛或鳞片由中胚层间质的区域位置决定,翅的真皮乳头诱导表皮产生羽毛,而足的中胚层核诱导表皮产生鳞片;容许互作中反应组织已包含有特定的发育潜能,其发育方式已经决定,诱导组织只能提供发育所需要的环境,而不能改变其发育方向;例如肾的输尿管芽上皮中胚层与生后肾间质属于典型的容许互作,输尿管芽进入生后肾间质,在分支的顶端上皮诱导间质聚集和形成肾小管;肾小管的分化并不一定完全需要输尿管芽的诱导,其他组织如脊髓、脑、唾液腺间充质等和生后肾间质相互作用,也能使生后肾间质分化为肾小管;所以在这种情况下,生后肾间质已经决定了,但要表现其决定,需要在诱导条件下进行分化;4、简述果蝇眼发育单个细胞之间的诱导。
发育概念

发育概念绪论先成论:在公元17世纪后期和18世纪,以精源学说和卵源学说为代表的先成论占了主导地位。
精源学说认为胚胎预先存在于精子中,卵源学说认为卵子中本来就存在微小的胚胎雏形。
这两个学说的共同点在于认为胚胎是成体的雏形,是配子中本来的固有结构,胚胎发育仅仅是原有结构的增大。
渐成论:又称后成论,早在公元前,Aristotle(公元前384-公元前322年)对于动物胚胎的不同部分和成体动物各种结构形成的原因提出了自己的观点,他在观察鸡、星鲛和一些无脊椎动物胚胎的基础上,他提出胚胎是由简单到复杂逐渐发育形成的,这个理论后来被称为“后成论”。
多线染色体:一种缆状的巨大染色体,见于有些生物生命周期的某些阶段里的某些细胞中。
由核内有丝分裂产生的多股染色单体平行排列而成。
各染色单体上的染色粒并排排列,构成多线染色体的带,带与带之间则称间带。
多线染色体的这种结构可用光学显微镜观察,也能在多线染色体上用原位分子杂交法进行基因定位,并就其结构与功能之间的关系进行系统研究,因此是细胞学和遗传学研究的有用材料。
胚胎诱导:胚胎的一种组织可以指导另一种相邻组织的发育。
在有机体的发育过程中,一个区域的组织与另一个区域的组织相互作用,引起后一个组织分化方向上的变化的过程。
组织者:背唇组织具有调控和组织一个几乎完整的胚胎产生的特殊能力,故称为组织者。
第一章细胞命运的决定形态发生决定子:是细胞质中的一类分子,在母细胞中呈极性分布,细胞分裂时导致两个子细胞中只有一个能够继承这种物质,它们有助于细胞命运的决定。
细胞分化:是指从单个全能的受精卵产生个体各种类型细胞的发育过程。
细胞定型:是指细胞在分化之前,将发生一些隐蔽的变化,使细胞朝特定方向发展的过程。
细胞定型分为特化(specification)和决定(determination)两个时相。
自主特化:细胞的命运决定与卵裂时所获得的细胞质相,这种细胞发育命运完全由内部细胞质组分决定的定型方式称为自主特化有条件特化:细胞或组织的命运与细胞所处的位置,即相邻细胞或组织之间通过相互作用有关。
第六章 原肠作用

(5)分层(delimination):一 个细胞层分离为近似平行的二 或多层细胞的过程。
(6)集中延伸(convergent extension): 指细胞 间相互插入,使所在组织变窄、变薄,并推动组 织一定方向移动。
送至母体 来自母体
滋养层细胞 胎盘母体部分
母体静脉 母体动脉
绒毛膜功能
• 与母体间的物质交换; • 合胞体滋养层产生的激素:
绒毛膜促性腺激素:为一种多肽类激素,可以使胎盘 (及母体卵巢)细胞产生孕酮。 孕酮:类固醇激素,能使子宫壁增厚并布满血管。 绒毛膜催乳激素:促进母体乳房发育,产后生乳。 • 保护胎儿免受母体免疫系统的损伤: 绒毛膜分泌阻止抗原(组织相容性抗原)产生的可溶 性蛋白;抑制子宫内正常免疫反应的淋巴细胞的产生; 细胞滋养保护胚胎不被母体免疫系统认出。
• 合胞体滋养层充分发育后,形成由滋养层 组织和富含血管的中胚层构成的器官--绒毛 膜。
• 绒毛膜和子宫壁融合形成胎盘。因此,胎 盘既含有母体成分(子宫内壁),又含有胎儿 成分(绒毛膜)。
人类胚胎与母体间的物质交流
来自胎儿
送至胎儿 来自胎儿 脐动脉
脐静脉
羊膜
绒毛膜微绒毛
绒毛膜间区
绒毛膜(胎盘胎儿部分)
原肠胚(gastrula) 原肠作用期的胚胎.
2、原肠作用主要特点
细胞运动涉及整个胚胎;胚胎细胞之间位 置发生变化,为相互作用奠定了基础。
原肠作用是从尚未分化细胞进入到分化为 三个胚层和决定各器官原基的关键时期。
3、原肠作用中的主要细胞迁移
第七章原肠运动

第七章原肠运动—胚胎细胞重组原肠作用概述。
海胆的原肠作用鱼类的原肠作用,两栖类的原肠作用,哺乳类的原肠作用一、原肠作用概述原肠作用(gastrulation):是胚胎细胞剧烈的、高速有序的运动过程,通过细胞运动实现囊胚细胞的重新组合。
原肠形成期间,囊胚细胞彼此之间的位置发生变动,重新占有新的位置,并形成由三胚层细胞构成的胚胎结构。
动物身体的主轴(包括前后轴、背腹轴和左右轴)也会在卵裂和原肠作用期间建立起来,胚胎各部分的细胞会获得各自的发育潜能。
不同动物体轴建立的时间和机制各不相同。
如果蝇体轴的特化自卵细胞形成时就已经开始了,在卵裂过程中完成;而爪蟾的体轴是在整个原肠作用过程中建立的。
尽管整个动物界原肠作用方式变化多样,但总体可概括为五种细胞运动机制,即外包(epiboly)、内陷(invagination)、内卷(involution)、内移(ingression)和分层(delamination)。
同一原肠胚常常包括几种细胞运动方式。
外包:植物极细胞由于卵黄多分裂慢,动物极细胞卵黄少分裂快,结果动物极细胞逐渐向下包围了植物极细胞,成为外胚层。
内部的植物极形成细胞成为内胚层。
如许多软体动物和蛙类的原肠胚的的形成。
分层:囊胚的细胞分裂时,细胞沿切线方向分裂,这样向着囊胚腔分裂出的细胞为内胚层,留在表面的一层为外胚层。
内转:由盘裂形成的囊胚,分裂的细胞由下面边缘向内内转,伸展成为内胚层。
2海胆的原肠作用海胆的晚期囊胚由1000个左右单层细胞构成。
这些细胞的细胞质分别来自受精卵的不同区域,细胞表现出不同的大小和特性。
海胆的囊胚A,细胞持续的分裂形成一个囊胚;B,快速分裂的末期,圆形的胚胎细胞形成一层上皮,受精膜依然可见;C,植物基板变厚,动物极细胞分泌孵化酶,使胚胎从受精膜中孵化出来。
海胆胚胎的早期囊胚,植物极细胞开始加厚。
原肠作用前的海胆晚期囊胚,植物极变扁平。
初级间质细胞的内移植物极板中央来源于小分裂球的细胞不断伸出和收缩线状伪足,脱离表面单层细胞,进入囊胚腔,称为初级间质细胞。
发育生物学笔记

重点:1. 掌握发育生物学的概念发育生物学(developmental biology)是应用现代生物学的技术研究生物的生殖、生长和分化等发育本质的科学。
2. 掌握发育生物学研究的对象和任务①发育生物学主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长、衰老和死亡,即生物个体发育 (ontogeny) 中生命过程发展的机制;②生物种群系统发生(systematics development) 的机制。
发育生物学的研究任务一个单细胞受精卵如何通过一系列的细胞分裂和细胞分化产生有机体的所有形态和功能不同的细胞,这些细胞又如何通过细胞之间的相互作用共同构建各种组织和器官,建成一个有机体并完成各种发育过程的。
阐明个体发育机制的核心问题是弄清遗传信息以何种方式编码在基因组上,DNA上的一维信息又是如何控制生物体的三维形态结构的构建和生命现象的发展。
3.动物发育的主要特征(1 ) 个体发育的特征;生物个体发育的特征是具有严格的时间和空间的次序性,这种次序性由发育的遗传程序控制。
发育是有机体的各种细胞协同作用的结果,也是一系列基因网络性调控的结果。
在发育的过程中涉及多种生命现象,如细胞分裂,细胞分化,细胞迁移,细胞凋亡,生长、衰老和死亡等。
(2 ) 个体发育的功能生物个体发育有两个主要的功能:①产生细胞的多样性并使各种细胞在本世代有机体中有严格的时间和空间次序性;②保证世代的交替和生命的连续。
4.动物发育的基本规律(1)受精新个体的生命开始于两性配子(gamete)——精子(sperm)和卵子(ovum)的融合,这个融合过程称为受精(fertilization)。
(2) 胚胎发育——形成幼体通过受精激活发育的程序,受精卵开始胚胎发育。
大多数动物要经过卵裂、原肠胚形成、神经胚形成(neurulation)、器官形成(organogenesis)等几个主要的胚胎发育阶段才能发育成为幼体。
(3)生长发育——成体 (4)衰老与死亡5. 了解后成论和先成论的基本内容后成论;胚胎是由简单到复杂逐渐发育形成的,这个理论后来称为后成论。
07 第七章 胚胎诱导及组织、器官形成

B 蛙囊胚背面
C 同一囊胚的腹面
E 在胚胎腹面注入
β-catenin后产
生的双体轴胚胎
2.中胚层形成的诱导
在囊胚赤道板有一条宽的环状的细胞带, 被植物极部位的信号因子所决定而发育为中 胚层组织。
将美西螈的中期囊胚分 为动物半球顶部帽状的 Ⅰ区、下部的环形的Ⅱ 区,植物半球的Ⅲ区。 如果将Ⅰ、Ⅱ区分离培 养,则只形成未分化的 非典型表皮,以后逐渐 退化。说明在中期囊胚 Ⅱ区尚未获得中胚层的 分化能力。
第一节 初级胚胎诱导
(primary embryonic induction)
初级胚胎诱导是指脊索中胚层诱导其表 面覆盖的外胚层发育为神经板的现象。
(一)组织者的发现及在初级胚胎诱导中的作用
❖将一种浅色蝾螈原肠胚的胚孔背唇 移植到深色蝾螈同龄胚胎腹侧。在宿 主原有的神经板相对的腹侧面形成了 另一个次级神经板。
❖其中最重要的一个背部 化决定子是转录因子βCATENIN。
❖在非洲爪蟾中,βcatenin在细胞质转动的过 程中开始在背部区域聚集, 在囊胚期只存在于背侧的 细胞中。
❖β-catenin是背轴形成的 关键。用β-catenin的反义 寡聚核苷酸处理会使胚胎 缺失背部结构。而在胚胎 的腹侧注入β-catenin会产 生第二个体轴。
❖晚期原肠胚胚孔背唇诱导躯干和尾部结构的形成,称为躯 干组织者和躯尾组织者
将原肠晚期原肠顶不同部位的移植块放到早 期原肠胚的囊胚腔中,会诱导不同的结构形成.
(二)初级诱导的过程及体轴决定的机制
初级胚胎诱导导致的胚胎模式形成过程包括四个 阶段:
1、Nieuwkoop中心的形成与背腹决定 2、中胚层形成的诱导 3、背部化与头尾化诱导 4、神经形成的诱导
发育生物学提纲

第一章绪论(3学时)教学目的与要求:1.了解发育生物学的研究对象、任务及其与其他学科学关系;2.掌握动物发育的主要特征和基本规律;3.了解发育生物学的发展简史;4.熟悉发育生物学研究中常用的模式生物及研究技术。
教学内容:第一节发育生物学的研究对象、任务及其与其他学科学关系发育(development)包括个体发育(ontogeny)和系统发育(phylogeny)。
个体发育是自受精卵开始到形成成熟个体所经历的一系列变化过程,系统发育是同一起源的生物群的形成历史。
发育生物学是从细胞水平和分子水平上研究生物个体发育机制的学科,它涉及基因如何控制胚胎细胞的行为、如何决定发育的模式以及胚胎的形态变化等重要问题,是生物学的核心学科。
随着分子生物学的发展与应用,发育生物学获得了迅速发展。
发育是生物界普遍存在的生物学现象,不同生物的发育既具有差异,又具有相似性,因此,对包括动物、植物和微生物在内的生物的发育过程进行比较和综合,有助于了解生物之间在发育上的共同规律和特殊性,有助于把握和理解发育的生物学本质。
由于历史的原因,人们对动物的发育研究比较充分和深入,而对植物等其他生物的研究则不够,因此,本课程中动物发育的内容所占比例相对较大。
第二节动物发育主要特征和基本规律动物的胚胎发育比较复杂,不同类的动物,胚胎发育的情况也不尽相同,但是早期胚胎发育的几个主要阶段却是相同的。
高等动物的发育过程也包括胚前发育、胚胎发育和胚后发育几个主要阶段,胚胎发育包括受精、卵裂、囊胚、原肠胚、中胚层及体腔形成、胚层分化等主要阶段,而低等动物因进化地位不同,其胚胎发育则缺少相应的后期几个阶段。
第三节发育生物学的发展简史一、胚胎发生的后成论(epigenesis)和先成论(prefomation)二、细胞学说促进了胚胎学理论的发展三、镶嵌式发育与调整式发育四、诱导现象的发现五、遗传学与发育学的结合第四节发育生物学的模式生物一、脊椎动物模式生物1.两栖类:非洲爪蟾(Xenopus laevis)2.鱼类:斑马鱼(Danio rerio)3.鸟类:鸡4.哺乳动物:小鼠二、无脊椎动物模式生物1.果蝇2.线虫:秀丽陷杆线虫第五节发育生物学研究技术一、常用发育生物学研究技术1.显微镜技术2.组织切片技术3.分子生物学技术4.原位杂交技术5.显微注射6.报道基因技术7.细胞标记技术二、发育遗传学技术1.正向遗传学技术2.反向遗传学技术第二章细胞命运的决定(3学时)教学目的与要求:1.掌握细胞命运的决定机制;2.掌握细胞命运决定在发育过程中的调控;3.了解常见模式生物的细胞命运决定。
发育学 课后总结 各章重点 答案

一、绪论1.发育生物学:是应用现代生物学的技术研究生物发育机制的科学。
它主要研究多细胞生物体的从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
镶嵌发育:合子的细胞核含有大量特殊的信息物质——决定子,在卵裂的过程中这些决定子被平均分配到子细胞中去控制子细胞的发育命运。
细胞的命运实际上是由卵裂时所获得的合子核信息早已预定的。
这一类型的发育我们称之为镶嵌发育。
胚胎诱导:是指在胚胎发育过程中,相邻细胞或组织间通过相互作用,决定其中一方或双方细胞的分化方向。
图式形成:胚胎细胞形成不同组织、器官和构成有序空间结构的过程。
形态模式:各门动物都具有区别于其他动物特有的解剖学特征,这些特有的解剖学结构内在的排列称为形态模式。
调整发育:胚胎为保证正常的发育,可以产生胚胎细胞位置的移动和重排,这样的发育为调整发育。
2.发育生物学的发展基础及过程如何?研究哪些问题?答:发展基础:胚胎学、遗传学、细胞生物学。
发展过程:形态→机理组织器官→细胞→分子它主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
同时,也研究生物种群系统发生的机制。
3.细胞学说对发育生物学发展的作用?答:细胞学说改变了胚胎发育和遗传的概念:19世纪30年代末:德国Mathias Schleiden和Theodor Schwann提出细胞学说。
1840, August Weismann提出了生殖细胞论,认为后代个体是通过精子和卵子继承亲本描述躯体特征的信息;卵子是一个细胞,其分裂产生的细胞可分化出不同组织,从而否定了先成论。
19世纪70-80年代,Oscar Hertwig兄弟对海胆受精卵的观察发现,受精卵含有两个细胞核,并最终合并为一个细胞核,表明细胞核含有遗传的物质基础。
19世纪末,染色体的发现和发现染色体数目在发育中的变化规律,使孟德尔遗传定律有了物质基础。
第七章 原肠运动-胚胎细胞重组

完成下包90%时斑马鱼胚胎 dorsal mesoderm
原肠作用 结束,体 节形成。
4. 两栖类的原肠作用
两栖类的原肠作用是实验胚胎学中最古老 也是最新兴的一个领域。 两栖类囊胚与棘皮动物和鱼类的囊胚所面 临的基本任务完全相同,也就是把终将形 成内胚层器官的细胞拖入胚胎内部,把终 将形成外胚层的细胞臵于胚胎的四周,而 把终将形成中胚层的细胞臵于外胚层和内 胚层之间适当的位臵。
海胆胚胎的早期 囊胚 ,植物极细 胞开始加厚。
原肠作用前 的海胆晚期 囊胚,植物 极变扁平。
初级间质细 胞的内移
植物极板中央 来源于小分裂 球的细胞不断 伸出和收缩线 状伪足,脱离 表面单层细胞, 进入囊胚腔, 称为初级间质 细胞。
海胆早期原肠胚 内初级间质细胞 沿囊胚腔内表面 运动,主动伸出 伪足与囊胚腔壁 连接,占据囊胚 腔预定腹侧面, 并形成稳固联系。 初级间质细胞融 合形成索状合胞 体(syncytial cable),最终形 成幼虫碳酸钙骨 针的轴。
爪蟾的 晚期囊 胚,示 边缘区 被内胚 层细胞 覆盖。
胚孔 处形 成瓶 状细 胞
移植的胚孔背唇细胞将在内胚层细胞层上下陷 (sink),形成一个胚孔狭缝。瓶状细胞的内陷 启动原肠作用。
瓶状细胞的收缩拉动边缘区细胞向植物极运动,同时 将植物极的内胚层细胞推向胚胎的内部。边缘区细胞 的运动导致外胚层细胞向植物极外包,预定前端中胚 层细胞沿囊胚腔内表面延伸。
随着瓶状细胞进入胚胎内部,背唇被后来内 卷进入胚胎发育为头部中胚层前体的细胞代 替。接下来,由胚孔背唇内卷进入胚胎的细 胞为脊索中胚层细胞,它们将形成脊索。 脊索是一个临时性中胚层脊柱(backbone), 对诱导神经系统的分化起重要的作用。
《发育生物学》课程教学大纲

《发育生物学》课程教学大纲(Development Biology)课程编号:1923027(1913027)课程类别:专业课适用专业:生物技术、生物科学、生物科学(师范)、生物工程先修课程:细胞生物学、遗传学、生物化学、微生物学、细胞工程后序课程:发酵工程、酶工程和蛋白质工程总学分:2 其中实验学分:0总学时:32教学目的与要求:发育生物学是研究生命体发育过程及其本质现象的科学,是近年来随着生命科学领域各学科的发展,尤其是分子生物学、细胞生物学、遗传学、生物化学等学科进展及其与胚胎学的相互渗透而发展形成的一门新兴学科。
通过对该课程的学习,帮助学生熟悉发育生物学的基本概念,牢固掌握基本原理,并能初步用于解释生物发育中出现的一些现象。
在学习和掌握发育生物学知识的过程中,要求学生将所学过的其他相关学科,如分子生物学、细胞生物学、遗传学、生物化学、生理学、解剖学、胚胎学、免疫学和进化生物学等的知识融会贯通,串联整合形成完整的知识体系。
教学内容与学时安排第一章动物的发育模式(2学时)一、发育生物学简史二、发育构建的多态性三、发育体制本章重点:发育构建的多态性难点:发育体制教学基本要求:掌握第二章胚胎发育的准备(2学时)一、生殖干细胞的决定、迁移二、精、卵细胞的发生本章重点:生殖干细胞的迁移难点:卵细胞的发生教学基本要求:掌握第三章胚胎的早期发育(4学时)一、受精二、囊胚的形成三、原肠胚四、神经胚的形成本章重点:原肠胚难点:受精教学基本要求:掌握第四章器官系统发生的奠定(4学时)一、内、中胚层的早期发育二、Hox基因三、体节形成与分化本章重点:Hox基因难点:体节形成与分化教学基本要求:掌握第五章动物成体组织结构的形成及器官系统的发生(2学时)一、神经系统二、肾脏的发生三、肢体的发生四、性别决定本章重点:肾脏的发生难点:性别决定教学基本要求:掌握第六章变态,胚后发育生长,衰老(2学时)一、昆虫变态二、哺乳动物胚后发育三、哺乳动物胚后生长本章重点:昆虫变态难点:哺乳动物胚后生长教学基本要求:掌握第七章细胞分化是发育建立的基础(2学时)一、近端诱导二、远程控制本章重点:近端诱导难点:远程控制教学基本要求:掌握第八章自组织,集约化(2学时)一、细胞黏着与亲和本章重点:细胞黏着与亲和难点:教学基本要求:掌握第九章发育程序的构建(2学时)一、发育程序的构建本章重点:三胚层的形成,肾脏的发生,细胞分化的分子机制。
发育生物学复习资料重点总结

绪论1、发育生物学:是应用现代生物学的技术研究生物发育机制的科学。
它主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
2、(填空)发育生物学模式动物:果蝇、线虫、非洲爪蟾、斑马鱼、鸡和小鼠。
第一篇发育生物学基本原理第一章细胞命运的决定1、细胞分化:从单个的全能细胞受精卵开始产生各种分化类型细胞的发育过程称细胞分化。
2、细胞定型可分为“特化”和“决定”两个阶段:当一个细胞或者组织放在中性环境如培养皿中培养可以自主分化时,可以说这个细胞或组织发育命运已经特化;当一个细胞或组织放在胚胎另一个部位培养可以自主分化时,可以说这个细胞或组织发育命运已经决定。
(特化的发育命运是可逆的,决定的发育命运是不可逆的。
把已特化细胞或组织移植到胚胎不同部位,会分化成不同组织,把已决定细胞或组织移植到胚胎不同部位,只会分化成同一种组织。
)3、(简答)胚胎细胞发育命运的定型主要有两种作用方式:第一种通过胞质隔离实现,第二种通过胚胎诱导实现。
(1)通过胞质隔离指定细胞发育命运是指卵裂时,受精卵内特定的细胞质分离到特定的裂球中,裂球中所含有的特定胞质可以决定它发育成哪一类细胞,而及邻近细胞没有关系。
细胞发育命运的这种定型方式称为“自主特化”,细胞发育命运完全由内部细胞质组分决定。
这种以细胞自主特化为特点的胚胎发育模式称为“镶嵌型发育”,因为整体胚胎好像是由能自我分化的各部分组合而成,也称自主型发育。
(2)通过胚胎诱导指定细胞发育命运是指胚胎发育过程中,相邻细胞或组织之间通过互相作用,决定其中一方或双方细胞的分化方向。
相互作用开始前,细胞可能具有不止一种分化潜能,但是和邻近细胞或组织的相互作用逐渐限制它们的发育命运,使之只能朝一定的方向分化。
细胞发育命运的这种定型方式成为“有条件特化”或“渐进特化”或“依赖型特化”,因为细胞发育命运取决于及其邻近的细胞或组织。
这种以细胞有条件特化为特点的胚胎发育模式称为“调整型发育”,也称有条件发育或依赖型发育。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
•初级间质细 胞在 Lytechinus variegatus早 期幼虫内的 定位。
第七章原肠运动胚胎细胞重组
•间质细胞和非间质细胞与不同胚层细胞或胞外组分之间的亲和力
第七章原肠运动胚胎细胞重组
•细胞粘连作用的变化导致初级间质的内移。 初级间质细胞和透明 层以及相邻裂球之间的亲和力减少,而与基质片层之间的亲和力增 加。相比之下,非间质细胞和透明层以及相邻裂球之间的亲和力维 持不变。
n 同一原肠胚常常包括几种细胞运动方式。
第七章原肠运动胚胎细胞重组
•原肠作用过 程中细胞的五 种运动方式
第七章原肠运动胚胎细胞重组
2. 海胆的原肠作用
n 海胆的晚期囊胚由1000个左右单层细胞构 成。这些细胞的细胞质分别来自受精卵的 不同区域,细胞表现出不同的大小和特性。
第七章原肠运动胚胎细胞重组
第七章原肠运动胚胎细胞重组
•两侧对称动 物的体轴
第七章原肠运动胚胎细胞重组
n 尽管整个动物界原肠作用方式变化多样,但总 体可概括为五种细胞运动机制,即外包 (epiboly)、内陷(invagination)、内卷 (involution)、内移(ingression)和分层 (delamination)。
•海胆的早期卵裂
第七章原肠运动胚胎细胞重组
•海胆胚胎第四次卵裂时形成的小分裂球(micromere)
第七章原肠运动胚胎细胞重组
•海胆的囊胚 A,细胞持续的分裂形成一个囊胚;B, 快速分裂的末期,圆形的胚胎细胞形成一层上皮,受 精膜依然可见;C,植物基板变厚,动物极细胞分泌 孵化酶,使胚胎从受精膜中孵化出来。
n 动物极半球存在着次级间质细胞附着的靶位。 只有当线状伪足接触到特定靶位的时候,才不 会缩回。这些特定的靶位可能位于将来形成口 的区域。
第七章原肠运动胚胎细胞重组
•A,原肠的顶端形成次级间质细胞;B,次级间质细胞的线状伪足。
第七章原肠运动胚胎细胞重组
n 当原肠最顶端接触到囊胚腔壁时,次级间质细 胞分散进入囊胚腔。次级间质细胞在囊胚腔中 分裂,最终形成中胚层器官。
n 胚盾在功能上相当于两栖类的背唇,将它移植 到宿主胚胎中,能够诱导形成次级胚胎。
第七章原肠运动胚胎细胞重组
•斑马鱼原肠作用时细胞的集中和延伸把下胚层和上胚层细胞 带到背部形成胚盾,胚盾内脊索中胚层向动第七物章原极肠运延动胚伸胎细。胞重组
n 当细胞沿卵黄下包时,它们也在边缘内卷,并 朝背部和前端两个方向向胚盾集中。胚盾下胚 层细胞向前端集中和延伸,最终沿背中线变成 一窄条,它就是脊索中胚层 (chordamesoderm),即脊索的原基。
n 卵黄合胞体层(yolk syncytical layer,YSL) 细胞的自动扩展拉动与之相连的包被层 (enveloping layer, EVL)细胞向下移动。 切断卵黄合胞体层和包被层之间的联系,胚盘 细胞会反弹缩回卵黄的顶部,而卵黄合胞体层 则继续包绕卵黄扩展。
第七章原肠运动胚胎细胞重组
•表达snail基因的近轴中胚层细胞在脊索中胚层两侧的 集中和延伸,形成中胚层体节的前体。
第七章原肠运动胚胎细胞重组
•完成下包90%时斑马鱼胚胎
第七章原肠运动胚胎细胞重组
•原肠作 用结束, 体节形成。
第七章原肠运动胚胎细胞重组
4. 两栖类的原肠作用
n 两栖类的原肠作用是实验胚胎学中最古老也是 最新兴的一个领域。
第七章原肠运动胚胎细胞重组
n 动物身体的主轴(包括前后轴、背腹轴和左右 轴)也会在卵裂和原肠作用期间建立起来,胚 胎各部分的细胞会获得各自的发育潜能。
n 不同动物体轴建立的时间和机制各不相同。如 果蝇体轴的特化自卵细胞形成时就已经开始了, 在卵裂过程中完成;而爪蟾的体轴是在整个原 肠作用过程中建立的。
n 与脊索中胚层毗连的细胞为近轴细胞(adaxial cell),是中胚层体节的前体。上胚层细胞形 成神经龙骨(neural keel),其余的上胚层细 胞发育成鱼类的皮肤。
第七章原肠运动胚胎细胞重组
•脊索中胚层细胞表达的no tail基因,示胚盾下胚层细 胞向前端的集中和延伸。
第七章原肠运动胚胎细胞重组
第七章原肠运动胚胎细胞重组
•通过原肠作用,胚胎首先建立起三个胚 层,即外胚层、中胚层和内胚层;其次为 重新占有新位置的胚胎细胞之间的相互作 用奠定了基础。因此,原肠作用是从尚未 分化到分化为三个胚层和器官原基决定的 关键时期。
•原肠胚一部分细胞的移动必然和同时发 生的其他细胞的运动紧密配合,因此原肠 作用过程中的细胞运动涉及整个胚胎。
3. 鱼类的原肠作用
中期囊胚转换和细胞运动性的获得
n 斑马鱼第十次卵裂期间,细胞分裂不再同步, 新的基因(合子基因)开始表达,且细胞获得 运动性,这种现象称为中期囊胚转换 (midblastula transition,MBT)。MBT似乎 受染色质和细胞质之间比例的控制。
第七章原肠运动胚胎细胞重组
n 之后,植物极板向内弯曲,内陷。当植物极板 内陷深及囊胚腔的1/4~1/2时,内陷突然停止。 所陷入的部分称为原肠(archenteron or primitive gut),而原肠在植物极的开口称为 胚孔(blastopore)。
第七章原肠运动胚胎细胞重组
•植物极 变扁平
第七章原肠运动胚胎细胞重组
第七章原肠运动胚胎细 胞重组
2020/12/5
第七章原肠运动胚胎细胞重组
It is not birth, marriage, or death, but gastrulation, which is truly the most important time in your life.
Lewis Wolpert (1986)
晚期原肠内陷
n 早期原肠内陷完成之后,经过短暂停歇,原肠 大幅度拉长, 短粗的原肠变成又细又长的管 状结构。在此期间没有新细胞形成,原肠的拉 长过程是通过细胞重排实现的,原肠周长内细 胞数目大为减少。
第七章原肠运动胚胎细胞重组
•海胆胚胎原肠伸长时期的细胞重排。
第七章原肠运动胚胎细胞重组
n 原肠顶端形成次级间质细胞,原肠延伸是以次 级间质细胞提供的张力为动力的。次级间质细 胞伸出线状伪足,穿过囊胚腔液,直达囊胚腔 壁内表面。
第七章原肠运动胚胎细胞重组
•下包胚盘的边缘区域的放大,示边缘下包的内卷和细胞内移
•通过下包胚盘的边缘细胞内卷或上胚层细胞分层形成下胚层
第七章原肠运动胚胎细胞重组
n 下胚层一旦形成以后,上胚层和下胚层的深层 细胞都会向将来发育成胚胎背部的一侧插入, 形成一个加厚的区域,称为胚盾(embryonic shield)。
第七章原肠运动胚胎细胞重组
•海胆早期原肠胚 内初级间质细胞 沿囊胚腔内表面 运动,主动伸出 伪足与囊胚腔壁 连接,占据囊胚 腔预定腹侧面, 并形成稳固联系。 初级间质细胞融 合形成索状合胞 体(syncytial cable),最终形 成幼虫碳酸钙骨 针的轴。
第七章原肠运动胚胎细胞重组
•初级间质细胞形成的索状合胞体(将来形成骨针)的扫描电镜照片 第七章原肠运动胚胎细胞重组
第七章原肠运动胚胎细胞重组
•内移至囊胚腔的初 级间质细胞被大量 胞外基质包绕。
第七章原肠运动胚胎细胞重组
•原肠作用过程中沿动植物极轴分布的纤维决定了初级间质细胞的
迁移方向。
第七章原肠运动胚胎细胞重组
•囊胚腔胞外基质纤维 与动植物极轴平行排列, 最终与初级间质细胞发 生联系。
第七章原肠运动胚胎细胞重组
•下包完成30%时的胚盘第七章原肠运动胚胎细胞重组
胚层形成
n 当斑马鱼胚盘细胞包绕一半的卵黄时,包被层 四周边缘区开始加厚,这个加厚的区域叫胚环 (germ ring),它是由外层的上胚层 (epiblast)和内层的下胚层(hypoblast)构 成。
n 一般认为,下胚层是边缘表层细胞内卷继而向 动物极迁移形成的。也可能由上胚层细胞分层 后内移形成。
•受精后,斑马鱼合子开始不完全卵裂。卵裂最初只限制在胚胎的 动物极,卵裂并未延伸到卵黄,导致在卵黄上形成一堆分裂球。前 5次卵裂为纵裂,第一次纬裂在受精后约2小时第七产章原生肠运了动胚6胎细4胞细重组胞期。
•斑马 鱼的囊 胚
第七章原肠运动胚胎细胞重组
n 鱼类胚胎中最早的细胞运动时,胚盘细胞沿卵 黄四周下包。初始阶段,胚盘细胞内层细胞向 外周迁移,插入表层细胞中。稍后,表层细胞 沿卵黄表面下包,直到将卵黄全部包围起来。
第七章原肠运动胚胎细胞重组
•海胆胚胎的早期 囊胚 ,植物极细 胞开始加厚。
第七章原肠运动胚胎细胞重组
•原肠作用前 的海胆晚期 囊胚,植物 极变扁平。
第七章原肠运动胚胎细胞重组
•初级间质 细胞的内移
•植物极板中 央来源于小分 裂球的细胞不 断伸出和收缩 线状伪足 (filopdia), 脱离表面单层 细胞,进入囊 胚腔,称为初 级间质细胞 (primary mesenchyme cell)。
n 囊胚腔壁接触到原肠的位置最终形成口,口和 原肠最顶端形成一连续相通的消化管。海胆的 胚孔最终形成肛门。
第七章原肠运动胚胎细胞重组
•海胆原肠 作用的整个 历程。发育 温度为25度。
第七章原肠运动胚胎细胞重组
•胆的 长腕幼虫
第七章原肠运动胚胎细胞重组
•海胆正常的胚胎发育
第七章原肠运动胚胎细胞重组
第七章原肠运动胚胎细胞重组
两栖类原肠作用时的细胞运动
n 非洲爪蟾的中胚层前体只存在于深层细胞中, 位于赤道区域表面细胞下面,而外胚层和内胚 层则来源于表面的表层细胞中。
n 蛙类原肠作用首先是从将来胚胎的背部,即刚 好位于赤道下方的灰色新月区开始的。
第七章原肠运动胚胎细胞重组
•两栖类的灰色新月 精子入卵后,皮层向精子进入的方向旋转大约30º,在动 物极皮层含大量色素而内层含有少量色素的物种中,这一胞质不同层次的相 对运动形成了一个在精子进入点对面的新月形的灰色区域,称为灰色新月。