细胞生物学 第七章
(细胞生物学基础)第七章核糖体

THANK YOU
感谢聆听
核糖体的结构
核糖体由大、小两个亚基组成,每个亚基都由RNA 和蛋白质构成。
大亚基含有28S、5S和5.8S三种RNA,而小亚基含有 18S RNA。
这些RNA与约60种不同蛋白质结合,形成稳定的复 合物。
核糖体的功能
核糖体是合成蛋白质的场所, 通过mRNA的翻译合成蛋白质 。
它具有起始、延长和终止三个 功能部位,分别对应于mRNA 上的起始密码子、终止密码子 以及多肽链的释放因子。
信号转导调控
核糖体可以感知细胞内的营养 和能量状态,通过信号转导途 径将信息传递给其他细胞器或 细胞核,进而调控细胞代谢和 生长。
核糖体与疾病的关系
核糖体与癌症
研究表明,核糖体的合成和功能异常可以影响肿瘤细胞的 增殖和分化,与癌症的发生和发展密切相关。
核糖体与神经退行性疾病
一些神经退行性疾病如阿尔茨海默病和帕金森病等与核糖 体功能异常有关,这些疾病的发生和发展过程中会出现核 糖体蛋白的聚集和神经元死亡。
核糖体与感染性疾病
病毒和细菌等微生物感染细胞后,会利用细胞内的核糖体 合成自身的蛋白质,因此核糖体也是抗感染免疫的重要靶 点。
05
核糖体的研究方法
核糖体的分离和纯化
分离
利用核糖体与其他细胞成分的密 度差异,通过离心技术将其从细 胞中分离出来。
纯化
通过一系列层析和电泳技术进一 步去除杂质,获得较为纯净的核 糖体样品。
核糖体的电镜观察
负染色
将核糖体样品置于负染色液中,使其 附着在载网上,然后进行电镜观察。
冷冻电镜
采用冷冻固定技术,将核糖体样品快 速冷冻并置于电镜下观察,以获得高 分辨率的图像。
核糖体的生物信息学分析
细胞生物学学习资料(第7-8章)

细胞生物学学习资料(第7-8章)第七章整合细胞内膜系统、蛋白质分选与膜泡运输学习要点第一节细胞质基质的含义与功能一、细胞质基质的涵义细胞质基质(cytoplamicmatri某)是真核细胞的细胞质中除去可分辨的细胞器以外的胶状物质,曾被称作细胞液(cellap)/透明质(hyaloplam)、胞质溶胶(cyto-ol)等。
细胞质基质的主要成分为参与中间代谢的数千种酶类、细胞质骨架结构、糖原和脂滴等物质。
细胞质基质是一个高度有序的体系,其中胞质骨架纤维贯穿其中,多数蛋白质直接或间接地与骨架或生物膜结合。
执行某一生物学过程的酶类彼此之间靠弱键结合,形成多酶复合体,催化一系列反应,此体系中前一个反应的产物极为下一个反应的底物。
蛋白质与蛋白质之间,蛋白质与其他大分子(如mRNA)之间都可以通过弱键相互作用,处于动态平衡之中。
这种高度有序的结构体系形成可以使细胞高效的完成复杂的代谢过程。
但此结构体系的维持必须依靠细胞质基质中高浓度的蛋白质以及其周围特定的离子环境。
细胞质基质和细胞质溶胶是从不同的角度提出的概念,二者虽然有一些差别,但人们常把这两个名词等同起来。
细胞质基质是真核细胞的细胞质中除去可以分辨的细胞器以外的胶状物质,式结构上的概念,在描述细胞结构是倾向于用此概念。
细胞质溶胶是用差速离心的方法分离细胞匀浆物种的各种细胞组分过程中,离心除去细胞核、线粒体、溶酶体、高尔基体和细胞质膜等细胞器或细胞结构后,存留在上清液中的成分,是生物化学上的概念。
①许多中间代谢过程都在细胞质基质中进行,如糖酵解过程、磷酸戊糖途径、糖原的合成、某些生物大分子的分解过程、蛋白质的合成、核苷酸代谢等。
②提供物质运输和能量传递通路。
③蛋白质在细胞质基质中分选及转运。
④维持细胞形态和运动、胞内物质运输及大分子定位。
通过细胞质骨架把蛋白质、mRNA等生物大分子固定在特定的位点,在细胞质基质中形成了更为精细的区域,使复杂的代谢反应高效而有序的进行。
细胞生物学第七章细胞骨架

(1)构成细胞内的网状支架,支持和维持细胞的形态
现在是23页\一共有79页\编辑于星期三
成纤维细胞中微管的分布
(2)参与中心粒、纤毛和鞭毛的形成
中心体的结构
LM下:中心粒:centriol 中心球:centrosphere
EM下:中心粒由9组三联微管组成
现在是24页\一共有79页\编辑于星期三
2.微管的化学组成
3.微管结合蛋白
4.微管的组装及其调节 5.微管的主要功能
现在是8页\一共有79页\编辑于星期三
1.微管的形态结构和存在形式
现在是9页\一共有79页\编辑于星期三
1.微管的形态结构和存在形式
微管的存在形式:
单管(质膜下)
二联管(鞭毛和纤毛)
三联管(中心粒和基体)
现在是10页\一共有79页\编辑于星期三
现在是36页\一共有79页\编辑于星期三
二、微丝(microfilament)
1.微丝的形态结构
2.微丝的化学组成
3.微丝结合蛋白 4.微丝的组装及其调节
5.微丝的主要功能
现在是37页\一共有79页\编辑于星期三
1.微丝的形态结构
又称肌动蛋白丝 (actin filament)
现在是38页\一共有79页\编辑于星期三
现在是17页\一共有79页\编辑于星期三
(1)微管的体外装配
现在是18页\一共有79页\编辑于星期三
(1)微管的体外装配
微管体外装配的过程与踏车现象
现在是19页\一共有79页\编辑于星期三
(2)微管的体内装配
微管组织中心 (microtubule organizing center, MTOC )
细胞生物学:第7章 细胞质基质与内膜系统

蛋白质的起始合成都是在细胞基质中,有些蛋白质 刚起始不久便转移至内质网膜上,一边进行多肽链 延伸一边进入内质网膜腔。这类蛋白主要包括:
向细胞外分泌的蛋白质 膜的整合蛋白:包括细胞膜、内质网、高尔基体 和溶酶体膜上的膜蛋白都是在内质网上合成
构成细胞器中的可溶性驻留蛋白:如需隔离的溶 酶体蛋白;需修饰加工的蛋白
折叠错误的多肽被送到细胞质中,被泛素(ubiquitin) 结合,然后送到蛋白酶体(proteosome)中降解
ubiquitin介导的 蛋白质降解途径
蛋白酶体
5、内质网的其它功能
1) 解毒功能 : 主要在肝细胞的光面内质网中进行。含有一些酶 可以将脂溶性的废物和代谢产生的有害物质变成 水溶性而被排出体外
一般,线粒体、叶绿体和过氧化物体不属于内膜系统
内膜系统在细胞内是按功能、分层次分布的
内膜系统如何实现在细胞基质中按功 能、分层次分布?
通过与细胞骨架或细胞膜结合,从 而实现在细胞基质中的定位
内质网 Endoplasmic Reticulum,ER
一、 形态结构特征
由一层单位膜所形成的囊状、泡状和管状结构,并 形成一个连续的网膜系统
TGN位于反面的最外层,与反面的扁平膜囊相连, 形态呈管网状,并有囊泡与之相连
功能: 蛋白质的运输信号在此被特异的受体接受,进行 分拣,集中,形成不同的分泌小泡,被运送到不同 的地点。因此, 主要功能是参与蛋白质的分类与包 装,最后从高尔基体输出。
某些“晚期”蛋白质的修饰也发生在TGN中
4、位于高尔基体周围大小不等的囊泡
顺面一侧的囊泡较小,可能是内质网与高尔基 体间的物质运输小泡
反面一侧的具有体积 较大的分泌泡与分泌颗 粒,将经高尔基体分类 和包装的物质运送到细 胞特定部位
细胞生物学 翟中和版 总结笔记第七章

Cell biology细胞生物学第七章真核细胞内膜系统、蛋白质分选与膜泡运输细胞内被膜区分类:细胞质基质、细胞内膜系统、有膜包被的细胞器第一节细胞质基质的含义和功能一、细胞质基质的含义(1)含义:在真核细胞的细胞质中,除去可分辨的细胞器以外的胶状物质主要含有:(1)与代谢有关的许多酶(2)与维持细胞形态和物质运输有关的细胞质骨架结构细胞质基质是一个高度有序的体系,细胞质骨架纤维贯穿在粘稠的蛋白质胶体中,多数的蛋白质直接或间接地与骨架结合,或与生物膜结合,从而完成特定的功能。
细胞质基质主要是由微管、微丝和中间丝等相互联系形成的结构体系,蛋白质和其他分子以凝聚或暂时的凝聚状态存在,与周围溶液的分子处于动态平衡。
差速离心获得的胞质溶胶的组分和细胞质基质溶液成分很大不同。
胞质溶胶中的多数蛋白质可能通过弱键结合在基质的骨架纤维上。
二、细胞质基质的功能(1)蛋白质分选和转运N端有信号序列的蛋白质合成之后转移到内质网上,通过膜泡运输的方式再转运到高尔基体。
其他蛋白质的合成都在细胞质基质完成,并根据自身信号转运到线粒体、叶绿体、细胞核中,也有些蛋白驻留在细胞质基质中。
(2)锚定细胞质骨架(3)蛋白的修饰、选择性降解1 蛋白质的修饰辅基、辅酶与蛋白的结合磷酸化和去磷酸化糖基化N端甲基化(防止水解)酰基化2 控制蛋白质寿命N端第一个氨基酸残基决定寿命细胞质基质能够识别N端不稳定的氨基酸信号将其降解,依赖于泛素降解途径3 降解变性和错误折叠的蛋白质4 修复变性和错误折叠的蛋白热休克蛋白的作用第二节细胞内膜系统及其功能细胞内膜系统是指在结构、功能乃至发生上相互关联、由膜包被的细胞器或细胞结构。
研究方法:电镜技术免疫标记和放射自显影离心技术和遗传突变体分析一、内质网的形态结构和功能内质网是由封闭的管状或扁平囊状膜系统及其包被的腔形成的互相沟通的三维网络结构。
(一)内质网的两种基本类型糙面内质网和光面内质网。
糙面内质网:扁囊状整齐附着有大量核糖体功能:合成分泌性蛋白和膜蛋白光面内质网:分支管状,小功能:脂质合成,出芽位点部分细胞合成固醇类激素糙面内质网有20多种和光面内质网不同的蛋白,说明有特殊装置隔开两种内质网的组分。
细胞生物学第七章 线粒体ppt课件

■ 两套遗传体系的协同性
通过离体实验发现两套 遗传体系的遗传机制不 同。 如放线菌酮是细胞质蛋 白质合成抑制剂,但是 对细胞器蛋白质的翻译 却没有作用。另外,一 些抗生素,如氯霉素、 四环素、红霉素等能够 抑制线粒体蛋白质的合 成,但对细胞质蛋白质 合成没有多大影响。 通过对转录的抑制研究, 发现线粒体基因转录的 RNA聚合酶也是特异 的(图)。
线粒体蛋白转运
图 线粒体蛋白转运的部位
分子伴侣(molecular chaperon)
概念:一类在序列上没有相关性但有共同功能的蛋白质,它 们在细胞内帮助其他含多肽的结构完成正确的组装,而且在 组装完毕后与之分离,不构成这些蛋白质结构执行功能时的 组份。 种类:伴侣素家族(chaperonin, Cpn)、热休克蛋白 家族 ( Hsp family )、 核质素、T 受体结合蛋白 (TRAP) 等 特征:1、分子伴侣对靶蛋白没有高度专一性,同一分子伴 侣可以促进多种氨基酸序列完全不同的多肽链折叠成为空间 结构、性质和功能都不相关的蛋白质。 2、它的催化效率很低。行使功能需要水解ATP,以改 变其构象,释放底物,进行再循环。 3、它和肽链折叠的关系,是阻止错误折叠,而不是促 进正确折叠。 4. 多能性(胁迫保护防止交联聚沉,转运,调节转录 和复制,组装细胞骨架) 5. 进化保守性
细胞生物学第七 章 线粒体
第一节、 线粒体的生物学特征
线粒体是能够在光学显微镜进行 观察的显微结构。 ● 1890年,德国生物学家 Altmann第一个发现线粒体。 ● 1897年对线粒体进行命名。 ● 1900年,Leonor Michaelis用 染料Janus green对肝细胞进行 染色,发现细胞消耗氧之后,线 粒体的颜色逐渐消失了,从而提 示线粒体具有氧化还原反应的作 用。
细胞生物学 第7章 真核细胞内膜系统蛋白质分选与膜(共103张PPT)
一,其意义在于:大大增加了细胞内膜的表面积;为多种酶特
别是多酶体系提供了大面积的结合位点;酶系统的隔离与连接;
蛋白质、糖、脂肪的合成、加工和包装;运输分泌物;扩散屏
障及膜电位建立;离子梯度的维持等。
二、内质网的形态结构与功能
(一)、内质网的形态结构及化学组成
②赋予蛋白质传导信号的功能;
③某些蛋白只有在糖基化之后才能正确折叠。
糖基一般连接在4种氨基酸上,分为2种:
(O-linked glycosylation):与
Ser、Thr和Hyp的OH连接,连接的糖为半乳糖
或N-乙酰半乳糖胺,在高尔基体上进行。
(N-linked glycosylation):与
丝氨酸、苏氨酸、
羟赖氨酸、羟脯氨酸
一般1~4个糖残基,
但ABO血型抗原较长
N—乙酰半乳糖胺等
内质网上进行N-连接的糖基化。
糖分子首先被糖基转移酶转移到膜上的磷酸多萜醇
(dolichol phosphate)分子上,装配成寡糖链。
再被寡糖转移酶转到新合成肽链特定序列(Asn-X-Ser
或Asn-X-Thr)的天冬酰胺残基上。
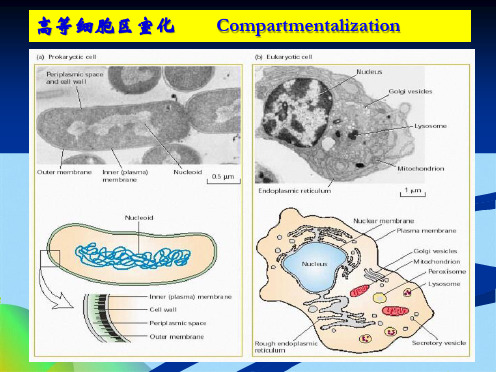
高等细胞区室化
Compartmentalization
质膜
内膜
生物膜
原核细胞 – 质膜构成的单一区室
真核细胞 – 内膜 – 各种细胞器
内膜系统出现的意义:
各自独立,各司其职
相互依存,协调同一
大大提高了细胞代谢反应的效率
细胞核
细
胞
细胞质
过氧化物酶体
线粒体
细胞骨架
细胞生物学 第七章

第二节 核糖体与蛋白质的生物合成
图7-5 核糖体上多肽链的合成过程 A.肽链合成的起始过程 B.肽链的延长过程
细胞生物学基础
第七章 核糖体
第一节 核糖体的类型及结构
一、核糖体的形态、数目与分布
核糖体呈不规则的颗粒状,其表面没有被膜包裹,直径 为l5~30nm,平均直径约为25nm(图7-1)。
图7-1 不同侧面观的核糖体
第一节 核糖体的类型及结构
核糖体常常分布在细胞内蛋白质合成旺盛的区域,其数量与蛋白质 合成的程度有关。处在指数生长期的细菌中,每个细胞内大约有数以万 计的核糖体,其含量可达细胞干重的40%。而在培养的饥饿状态的细胞 内,仅有几百个核糖体。般情况下,原核细胞约有1610个核糖体,真核 细胞约有110个核糖体,蛋内质合成旺盛的细胞可以高达110个。 真核细胞细胞质内的核糖体一般以两种形式存在:一种游离在细胞 质基质内,称游离核糖体;而另一种附着于内质网的膜表面,称附着核 糖体。原核细胞的核糖体除游离于细胞质基质外,还常常附着于细胞膜 内侧。附着核糖体与游离核糖体所合成的蛋白质种类不同,但核糖体的 结构与化学组成是完全相同的,它们在细胞中所占数目的多少与细胞的 类型、发育阶段及生理状态有关。在合成分泌蛋白质的细胞,如胰腺细 胞和浆细胞中,大多数核糖体附着在内质网膜上,肝细胞内75%的核糖 体附着在内质网膜上,其余25%则游离于细胞质中;在迅速生长的胚胎 细胞或肿瘤细胞内,大部分是游离核糖体,Hela细胞仅有15%的核糖体 附着在内质网膜上。
第一节 核糖体的类型及结构
核糖体具有复杂的三维结构,含有与蛋白质合成密切 相关的四个功能活性部位(图7-4)
图7-4 核糖体的4个活性部位
第二节 核糖体与蛋白质的生物合成
一、mRNA与遗传密码
细胞生物学 第七章 细胞骨架

微管的功能
(一)构成细胞的支架,维持细胞形态;
微管的功能
(二) 参与细胞内物质运输;
马达蛋白(motor protein)
• 能沿着细胞骨架铺 就的“轨道”运动 的蛋白,靠水解 ATP提供能量,介 导细胞内物质沿细 胞骨架的运输。
•
•
肌球蛋白(myosin)—— 与微丝有关的运动
驱动蛋白(kinesin)和 动力蛋白(dynein) —— 与微管有关的运动
纤维 (intermediate filament) 。
组成:由许多不同的蛋白质亚基装配成纤维状结构。 特点:弥散性、整体性、变动性
微丝 (microfilament, MF)
微管 (microtubules, MT)
中间纤维 (intermediate filament, IF)
细胞骨架的功能
13条原纤维 (一段微管)
延长
• 极性装配:
异二聚体首尾相接,组装成的微管具有极性; α微管蛋白(-),β微管蛋白(+) 在(+)极端发生装配使微管伸长
在(-)极端发生去组装使微管缩短 ----- 踏车行为
(二)微管的体内装配:
微管组织中心( microtubule organizing center, MTOC ):活细胞内微管组装时总是 以某部位为中心开始聚集,这个中心称为微 管组织中心,包括中心体、基体。为微管装配 提供始发区域,控制着细胞质中微管的数量、 位置及方向。
• 装配过程及极性规律同体外组装。
中心体
中心体 (centrosome) = 2个垂直的中心粒 + 周围物质 动物细胞内微管起始的主要位点。
中心粒结构
短筒状小体, 成对存在且相互垂直。
每个中心粒由9组三联体微 管斜向排列呈风车状包围 而成,为(9+0)结构 微管组织中心(MTOC), 参与有丝分裂。
细胞生物学第七章总结

第七章细胞骨架与细胞的运动第一节微管真核细胞中细胞骨架成分之一。
是由微管蛋白和微管结合蛋白组成的中空柱状结构。
还能装配成纤毛、鞭毛、基体、中心体、纺锤体等结构,参与细胞形态的维持、细胞运动、细胞分裂等。
微管蛋白与微观的结构存在:所有真核细胞,脊椎动物的脑组织中最多。
直径:24-26纳米中空小管基本构件:微管蛋白α、β异二聚体。
13根原纤维合拢成一段微管。
极性:增长快的为正端,另一端为负端。
(与细胞器定位分布、物质运输方向灯微管功能密切相关)γ微管蛋白:定位于微管组织中心,对微管的形成、数量、位置、极性的确定、细胞分裂有重要作用。
存在形式:单管(存在于细胞质,不稳定)、二联管(AB两根单管构成,主要分布于纤毛和鞭毛)、三联管(ABC三根单管组成,分布于中心粒、纤毛和鞭毛的基体中)一、微管结合蛋白碱性微管结合区域:明显加速微管的成核作用。
酸性突出区域:决定微管在成束时的间距大小种类:MAP-1,MAP-2,MAP-4,tau不同的微管结合蛋白在细胞中有不同的分布区域:tau只存在于轴突中,MAP-2则分布于胞体和树突中。
三,微管的装配的动力学装配特点:动态不稳定性装配过程:1、成核期(延迟期)α和β微管蛋白聚合成短的寡聚体结构,及核心的形成,接着二聚体再起两端和侧面增加使其扩展成片状带当片状带加宽至13根原纤维时,即合拢成一段微管。
是限速过程。
2、聚合期(延长期)细胞内高浓度的游离微管蛋白聚合速度大于解聚速度,新的二聚体不断加到微管正端使其延长。
3、稳定期(平衡期)胞质中游离的微管蛋白达到临界浓度,围观的组装与去组装速度相等(一)微管装配的起始点是微管组织中心中心体和纤毛的基体称为微管组织中心。
作用:帮助大多数细胞质微管装配过程中的成核。
γTuRC:刺激微管核心形成,包裹微管负端,阻止微管蛋白的渗入。
可能影响微管从中心体上释放。
中心体:包括中心粒,中心粒旁物质。
间期位于细胞核的附近,分裂期位于纺锤体的两极。
细胞生物学第七章 细胞质基质与细胞内膜系统

3 内质网(endoplasmic reticulum, ER)
• 内质网是由K.R.Porter和 A.D.Claude等在1945年发现的。 • (1)形态结构和化学组成形态: • 内质网是由一层单位膜所形成的扁 平囊状、泡状和管状结构,并形成 一个连续的网膜系统。由于它靠近 细胞质的内侧,故称为内质网。 • 分类: • 粗面内质网和光面内质网。 • 结构: • 由于内质网是一种封闭的囊状、泡 状和管状结构,它就有两个面,内 质网的外表面称为胞质溶胶面, 内 表面称为潴泡面。
• ● 信号序列的一般特征
• 长度一般为15~35个 氨基酸残基, • N-末端含有1个或多个 带正电荷的氨基酸,其 后是6~12个连续的疏 水残基; • 在蛋白质合成中将核糖 体引导到内质网,进入 内质网后通常被切除。
图 ER跨膜可切除信号的一般结构
• 信号序列的发现与 作用: • (a) 在不含RER小 泡的无细胞体系中 翻译分泌蛋白,其 N-端有信号序列, 故比从细胞中分泌 出来的相同蛋白质 肽链长; • (b)在加有RER小 泡的无细胞体系中 翻译分泌蛋白,信 号序列在RER小泡 中被切除,得到的 产物与从细胞中合 成分泌的相同。
• 3、供给细胞器行使其功能所需要的一切底物。
• 4、细胞骨架参与维持细胞形态,做为细胞器和酶的附 着点,并与细胞运动、物质运输和信号转导有关。 • 5、参与合成蛋白质的加工、运输、选择性降解:
磷酸化与去磷酸化,糖基化,甲基化,酰基化。
依赖泛素标记的蛋白酶体中的蛋白质降解途径。 热休克蛋白HSP帮助变性或畸形蛋白质重新折叠。 •
图 核膜和ER膜进化的可能途径
2.2 内膜系统与蛋白质分选(protein sorting) • 蛋白质是由核糖体合 成的,合成之后必须 准确无误地运送到细 胞的各个部位。 • 细胞中蛋白质的运输 有两种方式:共翻译 运输和翻译后运输, 内膜系统参与共翻译 运输,是分泌蛋白质 分选的主要系统。
细胞生物学 第7章 细胞的能量转换线粒体和叶绿体

第七章细胞的能量转换――线粒体和叶绿体生物的基本能量来源于太阳光的辐射能。
但生物体不能直接利用太阳光的辐射能,必须先使之转换成化学能,再为生物体利用。
叶绿体通过光合作用把光能转换为化学能,并储存于糖类、脂肪和蛋白质等大分子有机物中。
线粒体是一种高效地将有机物转换为细胞生命活动的直接能源ATP的细胞器。
因此,线粒体和叶绿体是细胞内的两种产能细胞器。
线粒体和叶绿体都具有环状DNA及自身转录RNA与翻译蛋白质体系。
很多学者把线粒体和叶绿体的遗传信息系统称为真核细胞的第二遗传信息系统,或称为核外基因及其表达体系。
线粒体和叶绿体都是半自主性细胞器。
第一节线粒体与氧化磷酸化人体内的细胞每天要合成几千克的ATP,且95%的ATP是由线粒体中的呼吸链所产生,因此线粒体被称为细胞内的“能量工厂”(power plants)。
线粒体通过氧化磷酸化作用,进行能量转换,为所需要的细胞进行各种生命活动提供能量。
一、线粒体的形态结构1、线粒体的形态、大小、数量与分布1)线粒体的形状线粒体的形状各种各样,以线状和颗粒状最常见。
也可呈环形、哑铃形、枝状或其他形状。
2)线粒体的大小线粒体的一般直径为0.5-1.0um,长,1.5-3.0um。
有的长达5um(如肝细胞)或10-20 um(胰腺细胞)或40 um(人的成纤维细胞)。
3)线粒体的数量线粒体的数目由数百――数千个不等。
如利什曼原虫中只有一个巨大的线粒体,海胆卵细胞则多达30万个。
4)线粒体的分布线粒体在细胞中的分布一般是不均匀的。
二、线粒体的超微结构在电镜下观察到线粒体是由两层单位膜套叠而成的封闭的囊状结构。
主要由外膜(outer membrane)、内膜(inner membrane)、膜间隙(intermembrane)、基质(matrix)或内室(inner chamber)4部分组成。
图7-11.外膜外膜是包围在线粒体最外面的一层单位膜,光滑而有弹性,厚约6 um。
细胞生物学课件第七章 线粒体与核糖体

➢据近期报载,日本专家研究证明:线粒体膜上 有分子发动机在工作。
➢这种分子发动机,其直径约为10纳米,由旋转 轴(一个细长的蛋白质分子)和轴承(6个蛋白质分 子)构成。
➢如果把它从体内取出来,向它供应三磷酸腺苷 生物能源,旋转轴就按反时针方向以每分钟8000 次的速度旋转,能源利用率近百分之百。
氧化磷酸化的偶联机制—化学渗透假说
•葡萄糖酵解
线粒体是物质氧化与能量转换的场所
线粒体主要功能是进行氧化磷酸化,合成ATP,为细胞生命活 动提供直接能量。
(蛋白质、脂类与糖类物质被消化分解为简单的物质形式被细胞 摄取,并在细胞内进一步降解为可进入线粒体的物质,在线粒 体中氧化分解,将化学能转移为ATP等物质的高能磷酸键中, 被细胞利用)
线粒体的化学组成及酶的定位
●线粒体组分分离方法 ●线粒体的化学组成 ●线粒体酶的定位
线粒体的化学组成
◆蛋白质(线粒体干重的65~70%) ◆脂类(线粒体干重的25~30%):
·磷脂占3/4以上,外膜主要是卵磷脂,
内膜主要是心磷脂---高度特化,具有离子 不通透性。 线粒体含有独特的DNA分子,编码线粒体部分蛋白质 亚基、RNA。
正是在这个意义上说,汉族本身是多源的基因库被同一个文化(汉文化) 同化、融合和统帅的产物。南方汉族群体中含有较北方群体更多的古老类 群和未定类型,提示南方汉族可能更古老。
● 叶绿体与光合作用 质体 ● 线粒体和叶绿体是半自主性细胞器 ● 线粒体和叶绿体的增殖与起源
1
3
2
叶绿体
白色体
有色体
叶绿体与光合作用
●叶绿体(Chloroplast)的形态结构 ●叶绿体的功能—光合作用(photosynthesis)
细胞生物学_翟中和--第七章 ppt

-
1
第一节 细胞质基质的涵义与功能
第二节 细胞内膜系统及其功能
第三节 细胞内蛋白质的分选与膜泡运输
-
2
形态学(电镜超薄切片)和生化:真核细胞内具有发达
的细胞内膜相结构,将细胞分为不同的区室,称为细胞
内区室化,是真核细胞结构和- 功能的基本特征之一。
3
第一节 细胞质基质的涵义与功能
在真核细胞的细胞质中,除去可辨认的细胞器以外的 胶状物质,称细胞质基质,是细胞的重要部分。
细胞组分
数目
体积比
细胞质基质 细胞核 内质网 高尔基体 溶酶体 胞内体 过氧化物酶体 线粒体
1 1 1 1 300 200 400 1700
-
54 6 12 3 1 1 1 22
-
34
-
35
-
36
4、新生多肽连的折叠与装配 已转移到ER腔 内的多肽链在ER 腔里进行折叠
和组装。在ER腔内有许多蛋白质是去其目的地的途 中,暂时经过的;
同时ER腔内也有高浓度的驻留蛋白,因为它含 有ER保留信号,即在蛋白质的C 端有4个氨基酸(Lys-Asp-Glu-Leu-COO-)负责保留蛋白在ER腔中。 如果这种ER保留信号用遗传工程方法去除了,则这 些蛋白就被分泌出去;如果这信号被转移到分泌蛋白, 这种蛋白便保留在ER腔中,ER驻留蛋白中。
-
39
-
40
5、内质网其他功能
-
41
Ca2+离子浓度的调节作用:肌质网是细胞 内特化的细胞器,是储存Ca2+的细胞器,其 上有Ca2+-ATP酶。
-
42
(三)内质网与基因表达的调控 内质网蛋白质的合成、加工、折叠、组装、转运 及向高尔基体转运的复杂过程显然是需要有一个 精确调控的过程。 影响内质网细胞核信号转导的三种因素: 内质网腔内未折叠蛋白的超量积累。 折叠好的膜蛋白的超量积累。 内质网膜上膜脂成份的变化——主要是固醇 缺乏 不同的信号转导途径,最终调节细胞核内 特异基因表达。
细胞生物学 第七章 核糖体
L11-rRNA复合物的三维结构 复合物的三维结构 (引自Porse et.al.,1999)
三、 细菌核糖体的分离和重组
1.分离 细菌的70s核糖体,含有二个亚单位,它们的漂浮 单位分别为50s和30s,都含有RNA和蛋白质。这些 RNA和蛋白质都可分离,其步骤如下: (1)在离心的溶液中Mg2+的浓度降低到1mmol/L以 下就可把70s核糖体(单体)分裂成50s和30s大小 两个亚单位; (2)将收集的亚单位在氯化铯溶液中离心,就可分 裂为分裂蛋白质和核心(由RNA和蛋白质组成), 核心的漂浮单位分别为40s和23s。这些核心没有合 成蛋白质的能力,但还可进一步分离;
第七章 核糖核蛋白体 (ribosome) )
核糖体颗粒存在于所有类型的活细胞中,游离在 细胞质中或附着在粗糙型内质网上,特别在快速增殖 的细胞中含量更多。也存在于细胞核、线粒体和叶绿 体内。 在真核细胞中很多核糖体附着在内质网的膜上, 称为附着核糖体,它与内质网形成复合细胞器,即粗 面内质网。在原核细胞质膜内侧也常有核糖体着附。 还有一些核糖体不附着在膜上,呈游离状态,分 布在细胞质基质内,称游离核糖体。附着在内质网膜 上的核糖体与游离核糖体所合成的蛋白质种类不同, 但核糖体的结构与化学组成是完全相同的。
在正常生长的细胞中,大多数的核糖体 担负着蛋白质的合成任务,它们经常丛集或 串联在一起,由一条宽度为1nm的mRNA 细 线贯通着。这些聚集的核糖体叫做多核糖体。 一个多核糖体由5-6个核糖体串联而成,也 可多至50个以上。
二、
核糖体的结构与组成
核糖体是无膜的细胞器,主要成分是蛋白 质与RNA。核糖体的RNA称为rRNA,约占 60%,蛋白质约占40%,蛋白质分子主要分布 在核糖体的表面,而rRNA则位于内部,二者靠 非共价键结合在一起。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
起始氨基酸为甲酰甲硫氨酸
3. 线粒体的遗传密码也与核基因不完全相同 4. UAA的终止密码位于mRNA的3ˊ端。某些情况下, 一个碱基U就是mtDNA体系中的终止密码子。
线粒体与核密码子编码氨基酸比较
密码子
UGA AGA AGG 核密码子编 码氨基酸 终止密码子 精氨酸 线粒体密码子编码氨基酸
哺乳动物
线粒体的内、外膜上存在着一些内膜与外膜相互接触的
地方为转位接触点(translocation contact site)。是蛋白质等
物质进出线粒体的通道。
线粒体相对独立的遗传体系
线粒体基因组:双链环状的DNA分子
人线粒体基因组
(剑桥序列) 16569 bp, 重链(H) 轻链(L )
37 基因: 2 rRNA 22 tRNA 13蛋白质
线粒体基因组图谱
重链编码
12SrRNA、16SrRNA、
14个tRNA NADH-CoQ氧化还原酶1(NADH-CoQ oxidoreductase 1,
ND1)、ND2、ND3、ND4L、ND4、ND5、
细胞色素C氧化酶1(cytochrome coxidase Ⅰ, COXⅠ)、 COXⅡ、COXⅢ、
色氨酸 终止密码子
果 蝇
色氨酸
酵 母
色氨酸 精氨酸
植 物
终止密码子 精氨酸
AUA
AUU CUU CUC CUA CUG
异亮氨酸
异亮氨酸
甲硫氨酸
异亮氨酸
甲硫氨酸
甲硫氨酸
异亮氨酸
甲硫氨酸
异亮氨酸
甲硫氨酸
异亮氨酸
异亮氨酸
亮氨酸
亮氨酸
亮氨酸
亮氨酸
苏氨酸
亮氨酸
线粒体基因中两个重叠基因,
1931年Otto Heinrich Warburg因“发现呼吸酶的性 质及作用方式”被授予诺贝尔生理学或医学奖。
"discovery of the nature and mode of action of the respiratory enzyme."
While working at the Marine Biological Station, Warburg performed research on oxygen consumption in sea urchin eggs after fertilization, and proved that upon fertilization, the rate of respiration increases by as much as sixfold.
细胞色素b 的亚基、
ATP合酶的第6亚单位和第8亚单位(A6、A8)
轻链编码
8个tRNA ND6
线粒体基因的转录
重链启动子(heavy-strand promoter,HSP)
轻链启动子(light-strand promoter,LSP)
1. 线粒体mRNA不含内含子,也很少有非翻译区
2. 每个mRNA5ˊ端的起始密码为AUG(或AUA),
线粒体靶序列引导核编码蛋白质向线粒体转运
(一)核编码蛋白在进入线粒体需要分子伴侣蛋白的协助
线粒体内含有1000~1500种蛋白质,除上述的13种多肽外, 98%以上是由细胞核DNA编码。 线粒体含有4个蛋白质输入的亚区域: 线粒体外膜 膜间隙 线粒体内膜 基质
其中绝大多数线粒体蛋白被输入到基质,少数输入到膜间
超微结构:由双层单位膜套叠而成的封闭性膜囊结构
外膜(outer membrane):线粒体最外层所包绕
的一层单位膜,厚约5~7nm,光滑平整。在组成上,
外膜的脂质和蛋白质成分各占1/2。
外膜中含有整合蛋白孔蛋白 (porin),它们以β片层结构
形式形成直径2~3nm桶状通道,
跨越脂质双层,可以通过相对 分子质量在5 000以下的物质。
前体蛋白与MSF所形成的复合体能进一步与外 膜上的一套受体Tom37和Tom70相结合。
Tom37和Tom70把前体蛋白转移到第二套受体 Tom20和Tom22,同时释放MSF。 与前体蛋白结合的受体Tom20和Tom22与外膜 上的通道蛋白Tom40(第三套受体)相偶联。 后者与内膜的接触点共同组成一个直径为 1.5~2.5nm的越膜通道(tim17受体系统, 非折叠的 前体蛋白通过这一通道转移到线粒体基质。
腔以及插入到内膜和外膜上。
基质导入顺序 (matrix-targeting sequence,MTS)
输入到线粒体的蛋白质都在其N-端具有一段线粒体
靶序列称为基质导入顺序
线粒体外膜和内膜上的受体能识别并结合各种不同
的但相关的MTS。
部分核编码的线粒体蛋白
线粒体定位
基质
蛋白质
乙醇脱氢酶(酵母) 氨甲酰磷酸合酶(哺乳动物) 柠檬酸合酶(citrate synthase)与其他柠檬酸酶 DNA聚合酶 F1ATP酶亚单位α(除植物外),β、γ、δ(某些真菌) Mn2+超氧物岐化酶 鸟氨酸转氨酶(哺乳动物) 鸟氨酸转氨甲酰酶(哺乳动物) 核糖体蛋白质 RNA聚合酶 ADP/ATP 反向转运体(antiporter) 复合体Ⅲ亚基1、2、5(铁-硫蛋白)、6、7 复合体Ⅳ(COX)亚基4,5,6,7 F0ATP酶 生热蛋白(thermogenin)
复合物Ⅰ的ND4L和ND4,
复合物Ⅴ的ATP酶8和ATP酶6。
线粒体DNA的复制
人类mtDNA是单一的复制起始,mtDNA的复 制起始点被分成两半, 一个是在重链上,称为重链复制起始点(OH),位 于环的顶部,tRNAPhe基因(557)和 tRNAPro基因 (16023)之间的控制区,它控制重链子链DNA 的自我复制;
1949年Kennedy和Lehninger发现脂肪酸氧化成为CO2的过 程是在线粒体内完成的,
1968-1976年the genetic and physical map of yeast mitochondria
1976年Hatefi等纯化了呼吸链四个独立的复合体。
1961-1980年Mitchell提出了氧化磷酸化的化学偶联学说。
Transmission electron micrograph of mitochondria in a brown adipose cell from a bat. These spherical mitochondria are large, as seen when compared to the nucleus at UP of the image. Despite the size of the mitochondria, the cristae still span the entire.
基质为物质氧化代谢提供场所
线粒体内腔充满了电子密度较低的可溶性蛋白质和脂肪等
成分,称之为基质(matrix)。线粒体中催化三羧酸循环、脂肪
酸氧化、氨基酸分解、蛋白质合成等有关的酶都在基质中。
还含有线粒体独特的双链 环状DNA、核糖体,这些构成 了线粒体相对独立的遗传信息 复制、转录和翻译系统。
内外膜转位接触点:核编码蛋白质进入线粒体的通道
线粒体基因编码的蛋白质
13个基因
NADH-CoQ还原酶复合体(复合体Ⅰ)亚基(ND1、 ND2、ND3、ND4L、ND4、ND5和ND6) 7个基因 CoQH2-细胞色素c还原酶复合体(复合体Ⅲ)中细 胞色素b 的亚基。 1个基因 细胞色素c氧化酶(COX)复合体(复合体Ⅳ)催 化活性中心的亚单位(COXⅠ、COXⅡ和 COXⅢ);3个基因 ATP合酶复合体(复合体Ⅴ)F0部分的2个亚基 (A6和A8) 2个基因。
线粒体与能量转换
(mitochondria and energy conversion)
线 粒 体
转换能量的细胞器
Carbohydrates
碳水化合物
ATP
叶绿体 植物细胞
线粒体 动物细胞
线粒体研究的里程碑事件
1890年R. Altaman首次发现线粒体,命名为bioblast,以为它
可能是共生于细胞内独立生活的细菌。
一个是在轻链上,称为轻链复制起始点(OL),位 于环L的“8点钟”位置,它控制轻链子链DNA的 自我复制。
Dividing Mitochondria
Who else divides like that?
What does this tell us about the evolution of eukaryotes?
线粒体的形态学
Meissner Barbara
显微结构
詹纳斯绿 Janus Green B
一种活体染色剂,专一用于线粒体的染色。它可以和线粒体中的细胞 色素C氧化酶结合,从而出现蓝绿色。
光镜下的线粒体呈线状、粒状或杆状等,直径 0.5~1.0μm。不同类型或不同生理状态的细胞,其 线粒体的形态、大小、数目及排列分布常不相同。
1940年 Claude A (1940) Particulate components of normal and tumor cells. Science 91: 77-78.
1946 年Albert Claude concluded that cytochrome oxidase and other enzymes responsible for the respiratory chain were isolated to the mitchondria by tissue fractionation。 1948年Green证实线粒体含所有三羧酸循环的酶,
(三)分子运动产生的动力协助多肽链穿越线粒体膜
前体蛋白一旦和受体结合后,就要和外膜及内 膜上膜通道发生作用才可进入线粒体。在此过程中, 一种也为分子伴侣的线粒体基质hsc70(mthsp70) 可与进入线粒体腔的前导肽链交联,提示mthsp70参 与了蛋白质的转运。