大学生物化学课件蛋白质的生物合成
生物化学第十一章 蛋白质的生物合成(共65张PPT)全

原核、真核生物各种起始因子的生物功能
起始因子
生物功能
IF-1
占 据 A 位 防 止 结 合 其 他 tRN A
原核
生物
EIF-2
促进起始tRNA与小亚基结合
EIF-3
促 进 大 小 亚 基 分 离 , 提 高 P位 对 结 合 起 始 tRNA 敏 感 性
eIF-2
促进起始tRNA与小亚基结合
eIF-2B,eIF-3
eEF-1-A
EF-Ts 再生EF-Tu
eEF-1-B
EFG
有转位酶活性,促进mRNA肽酰-tRNA由A位前移到P位, 促进卸载tRNA释放
eEF-2
(一)进位(P607 609)
又称注册(registration)
指根据mRNA下一组遗传密 三
码指导,使相应氨基酰-tRNA进 元
入核蛋白体A位。
第一节 蛋白质合成体系
一、翻译模板mRNA及遗传密码
二、核蛋白体是多肽链合成的装置 三、tRNA与氨基酸的活化
P602
一、翻译模板mRNA及遗传密码
(一) mRNA是遗传信息的携带者
1.顺反子(cistron):将编码一个多肽的遗传单位称为顺反
子。
2. 开放阅读框架(open reading frame, ORF):从mRNA 5 端起始密码子AUG到3端终止密码子之间的核苷酸序列。
mRNA 的结构
原核生物的多顺反子
5 PPP
ORF
ORF
真核生物的单顺反子
5 mG - PPP
3
ORF
蛋白质
3
蛋白质
非编码序列
核蛋白体结合位点
编码序列
起始密码子
【生物化学】蛋白质的生物合成

嘌 呤 霉 素
酯键
(3)转位(translocation)
•转位酶 (translocase): •原核:延长因子G(EF-G),真核:EF-2 • GTP
可结合并水解1分子GTP,促进核 蛋白体向mRNA的3’侧移动
进 位
成肽 转 位
合成
3、肽链终止阶段:
核蛋白体沿mRNA链滑动,不断使多 肽链延长,直到终止信号进入受位。
四、蛋白质生物合成的干扰和抑制
1、抗生素(antibiotics)
名称
作用机制
四环素类 氯霉素类 链霉素类 嘌呤霉素
抑制氨酰-tRNA与原核生物核糖体结合,抑制细菌 蛋白质合成
结合原核生物核糖体大亚基,阻断翻译延长过。高 浓度时,对真核生物线粒体内的蛋白质合成也有阻 断作用 结合原核生物核糖体小亚基,改变其构象,引起读 码错误
,IF)
有抗病毒作
用的蛋白质
1、诱导一种蛋白激酶,使eIF2磷酸 化,从而抑制病毒蛋白质的生物合 成。
2、诱导生成一种寡核苷酸(2’5’A),活化核酸内切酶RNaseL, 可降解病毒RNA。
谢 谢!
(1)识别:释放因子(RF)识别终 止密码,进入核蛋白体的受位。
(2)水解:RF使转肽酶变为水解酶, 多肽链与tRNA之间的酯键被水解, 多肽链释放。
(3)解离:通过水解GTP,使核蛋 白体与mRNA分离,tRNA、RF脱落, 核蛋白体解离为大、小亚基。
进位
肽链的形成 移位
蛋白质 合成过程
肽链合成终止
结构与Tyr-tRNAtyr相似,阻止肽链正常合成
放线菌酮 抑制核糖体转肽酶。且只对真核生物有特异性作用
2、干扰蛋白质生物合成的生物活性物质
名称
动物生物化学课件:蛋白质的生物合成

蛋白质的生物合成
将mRNA分子中 4 种核苷酸序列 编码的遗传信息,通过遗传密码破译的 方式解读为蛋白质一级结构中20种氨基 酸的排列顺序过程,称为蛋白质的生物 合成或翻译。
参与蛋白质生物合成的物质 蛋白质生物合成的过程
第一节 参与蛋白质生物合成的物质
参与蛋白质合成的物质
• 原料:20种氨基酸 • 模板:mRNA • 运载体:tRNA • 场所:核蛋白体(rRNA与蛋白质构成) • 蛋白质因子:
生物功能
占据A位防止结合其他tRNA 促进起始tRNA与小亚基结合 促进大、小亚基分离,提高P位对结合起始tRNA的 敏感性 促进起始tRNA与小亚基结合 最先结合小亚基促进大、小亚基分离 eIF-4F复合物成分,有解螺旋酶活性,促进mRNA 结合小亚基 结合mRNA,促进mRNA扫描定位起始tRNA eIF-4F复合物成分,结合mRNA5`-帽子 eIF-4F复合物成分,结合eIF-4E和PAB
➢ tRNA凭借自身的反密码子与mRNA链上的密码 子相识别,按照mRNA链上的密码子所决定的氨 基酸顺序将所带氨基酸转运到核糖体的特定部位。
一种氨基酸可以有一种以上tRNA作为 运载工具。通常把携带相同氨基酸而反密 码子不同的一组tRNA称为同功tRNA.
氨基酰tRNA----氨基酸的活化形式。 表示为: tRNAPhe
对应同一种氨基酸的不同密码子,称 为同义密码子。同义密码子使用频率不同.
在蛋白质中出现频率越多的氨基酸, 其密码子的数量越多。
4.密码子使用频率不同
• 在蛋白质合成时,对简并密码子的使用频率是 不同的。
• 如UUU和UUC都为苯丙氨酸编码,但在高表 达的蛋白质中使用UUC的频率明显高于UUU。
5. 密码子与反密码子配对的不严格性
生物化学第18章_蛋白质的生物合成
• 4). 肽基转移酶活性位点
• 位于P和A位点的连接处,靠近tRNA的接受臂. • 23SrRNA,L2,L3,L4,L15,L16
5). 5SrRNA位点
• 在50S亚基上靠近肽基转移酶活性位点 • L5,L8,L25
6). EF-Tu位点
• 位于50S亚基,靠近30S亚基 • L5,L1,L20,L7/L12
7). 转位因子EF-G结合位点
• 在50S亚基,靠近30S亚基界面处.
8). E位点
9).多肽出口位点
蛋白质的合成过程
氨基酸的活化
氨基酸先被tRNA-氨基酰合成酶活化,氨基 酸的羧基以高能键连接于腺苷酸,生成氨酰 腺苷酸 (Aa-AMP),同时释放焦磷酸.
Aa + ATP
氨酰tRNA合成酶
Aa-AMP + ppi
真核生物的多肽定向运输
• 溶酶体 线粒体 叶绿体,细胞核 • 信号肽及其信号肽的识别
信号肽长40个氨基酸残基 N端至少含一个带正电荷的氨基酸残基 在中部有10-15个高度疏水的氨基酸残基 有一个信号肽识别位点,其上游常有高度疏水的5 信号肽并不都在N端卵清蛋白的信号肽位于中部 识别信号肽的
SRP 是一种核蛋白。有300个核苷酸 的RNA和6个多肽组成, SRP 携带 新生多肽链的核糖体结合移动到内 质网上的
eEF-1βγ同EF-Ts功能
(3) 肽键的形成
肽基转移酶催化肽键的形成
3. “接头”的特异性只与反密码子有关
பைடு நூலகம்
延伸及 延伸相关因子
• EF-Tu • EF-Ts • EF-G
•植物中蛋白质翻译的起始
二. 蛋白质合成的动态过程
A0001401.mov
大连理工大学生物化学课件--蛋白质合成与转运

二、蛋白质生物合成过程
• • • • 蛋白质生物合成过程包括三大步骤: ①氨基酸的活化与搬运; ②活化氨基酸在核蛋白体上的缩合; ③多肽链合成后的加工修饰。
核糖体主要存在于粗面ER
核糖体存在的场所 (1)粗面内质网(主要) 一个细菌细胞内约有20000个核糖体 • (2)细胞溶液 • 真核细胞内可达106个 (3)线粒体和叶绿体 • 在未成熟的蟾蜍卵细胞内则高达1012个
(二)肽链延长阶段:
1.进位:与mRNA下一个密码相对应 的氨基酰tRNA进入核蛋白体的受位(A 位),需GTP,Mg2+,和EF参与。 2.成肽:在转肽酶的催化下,将给位 上的tRNA所携带的甲酰蛋氨酰基或肽 酰基转移到受位上的氨基酰tRNA上, 与 其 α- 氨 基 缩 合 形 成 肽 键 。 此 步 骤 需 Mg2+,K+。给位上已失去蛋氨酰基或肽 酰基的tRNA从核蛋白上脱落。
8、供能物质和无机离子
• 多肽链合成时,需ATP、GTP作为供能物质,并需 Mg2+、K+参与。
• 氨基酸活化时需消耗2分子高能磷酸键,肽键形成 时又消耗2分子高能磷酸键,故缩合一分子氨基酸 残基需消耗4分子高能磷酸键。
氨酰- tRNA合成酶
氨基酸 + tRNA + ATP
无机磷酸酶
氨酰- tRNA + AMP + 2Pi
一、参与蛋白质生物合成的物质
• 生物体内的各种蛋白质都是利用生物体内的氨基酸 为原料自行合成的。参与蛋白质生物合成的各种因 素构成了蛋白质合成体系,该体系包括: ① mRNA:作为蛋白质生物合成的模板,决定多肽链 中氨基酸的排列顺序; ② tRNA:搬运氨基酸的工具; ③ 核蛋白体:蛋白体生物合成的场所; ④ 酶及其他蛋白质因子;氨酰-tRNA合成酶; ⑤ 供能物质及无机离子。ATP和GTP, Mg2+、K+ ;
中国海洋大学资料生物化学课件第十五章 蛋白质合成(新)

(一)遗传密码的破译
* 美国科学家 Nirenberg 等, 1968 年获诺贝尔生 理学奖。 * 1961 年, Nirenberg 等人,在大肠杆菌的无细 胞体系中外加 poly(U) 模板、 20 种标记的氨基 酸,经保温后得到了多聚 phe-phe-phe ,于是 推测UUU编码phe。利用同样的方法得到 CCC 编码pro,GGG编码gly,AAA编码lys。 * 如果利用 poly ( UC ),则得到多聚 Ser-LeuSer-Leu,推测UCU编码Ser,CUC编码Leu。 * 到1965年就全部破译了64组密码子。
蛋白质合成时,mRNA结合于核糖体小亚基 上,大亚基结合带氨基酸的tRNA,tRNA的 反密码子与mRNA密码子配对,ATP供能, 合成蛋白质。
三、遗传密码
遗传密码:mRNA中的核苷酸序列与蛋白质 中氨基酸序列之间的对应关系称为遗传密码 三个碱基编码一个氨基酸称为三联体密码或 密码子(Coden)。 密码子的发现。
(1)在进入A位点之前,新氨酰tRNA首先必须与延伸因子EF— TU—GTP结合。 (2)延伸因子 EF—Tu是一个GTP结合蛋白,参与氨酰 tRNA的 就位。 (3)氨酰tRNA入位后,EF—TU—GTP水解,EF—TU—GDP从 核糖体上释放下来,在第二个延伸因子 EF—Ts 帮助下 EF— Tu—GDP 释 放 掉 GDP 并 重 新 结 合 一 分 子 GTP 再 生 成 EF— Tu—GTP。
简并性的生物学意义
A、减少有害突变:假如每种氨基酸只有一个密 码子,那么剩下的44个密码子都成了终止子, 如果一旦哪个氨基酸的密码子发生了单碱基的 点突变,那么极有可能造成肽链合成的过早终 止。如GUN 编码 Ala ,由于简并性的存在,不 论第三位的U变成什么,都仍然编码Ala; B、可以使DNA上的碱基组成有较大的变化余地, 而仍然保持多肽上氨基酸序列不变(意思基本 同上)。
生物化学第十二章-蛋白质的生物合成

第十二章蛋白质的生物合成一、蛋白质生物合成体系:生物体内的各种蛋白质都是生物体利用约20种氨基酸为原料自行合成的。
蛋白质的生物合成过程,就是将DNA传递给mRNA的遗传信息,再具体的解译为蛋白质中氨基酸排列顺序的过程,这一过程被称为翻译(translation)。
参与蛋白质生物合成的各种因素构成了蛋白质合成体系,该体系包括:1.mRNA:作为指导蛋白质生物合成的模板。
mRNA中每三个相邻的核苷酸组成三联体,代表一个氨基酸的信息,此三联体就称为密码。
共有64种不同的密码。
遗传密码具有以下特点:①连续性;②简并性;③通用性;④方向性;⑤摆动性;⑥起始密码:AUG;终止密码:UAA、UAG、UGA。
2.tRNA:在氨基酸tRNA合成酶催化下,特定的tRNA可与相应的氨基酸结合,生成氨基酰tRNA,从而携带氨基酸参与蛋白质的生物合成。
tRNA反密码环中部的三个核苷酸构成三联体,可以识别mRNA上相应的密码,此三联体就称为反密码。
反密码对密码的识别,通常也是根据碱基互补原则,即A—U,G—C配对。
但反密码的第一个核苷酸与第三核苷酸之间的配对,并不严格遵循碱基互补原则,这种配对称为不稳定配对。
能够识别mRNA中5′端起动密码AUG的tRNA称为起动tRNA。
在原核生物中,起动tRNA是tRNAfmet;而在真核生物中,起动tRNA是tRNAmet。
3.rRNA和核蛋白体:原核生物中的核蛋白体大小为70S,可分为30S小亚基和50S大亚基。
真核生物中的核蛋白体大小为80S,也分为40S小亚基和60S大亚基。
核蛋白体的大、小亚基分别有不同的功能:⑴小亚基:可与mRNA、GTP和起动tRNA结合。
⑵大亚基:①具有两个不同的tRNA结合点。
A位——受位或氨酰基位,可与新进入的氨基酰tRNA 结合;P位——给位或肽酰基位,可与延伸中的肽酰基tRNA结合。
②具有转肽酶活性。
在蛋白质生物合成过程中,常常由若干核蛋白体结合在同一mRNA分子上,同时进行翻译。
生物化学蛋白质的生物合成

的作用
伴侣素的主要作用—— 为非自发性折叠蛋白质提供能折叠形成天然 空间构象的微环境。
伴侣素GroEL/GroES系统促进蛋白质折叠过程
(二)蛋白二硫键异构酶 (protein disulfide isomerase, PDI) 在内质网腔活性很高,可在较大区 段肽链中催化错配二硫键断裂并形成正 确二硫键连接,最终使蛋白质形成热力 学最稳定的天然构象。
第一步:活化反应 氨基酸 +ATP+E → 氨基酰-AMP-E + PPi
第二步:转移反应
氨基酰-AMP-E + tRNA
↓
氨基酰-tRNA + AMP + E
氨基酰-tRNA的表示方法
Ala-tRNAAla
Ser-tRNASer Met-tRNAMet 起始肽链合成的氨基酰-tRNA
真核生物: Met-tRNAiMet
每增加一个氨基酸就按 进位→成肽→转 位 这三步不断重复,直到肽链增长到必要的长 度。
移位反应过程
核糖体循环的反应过程
进 位
转肽 移 位
三、蛋白质多肽链合成的终止和释放
蛋白质多肽链合成的终止过程
• 核糖体沿mRNA链滑动,不断使多肽链延长,
直到终止信号进入A位。
识别:RF识别终止密码,进入核糖体的A位。 水解:RF使转肽酶变为酯酶,多肽链与tRNA之间的 酯 键被水解,多肽链释放。 脱离:模板mRNA、RF以及空载tRNA与核糖体脱离 ,
四、蛋白质合成所需的能量
原核生物
氨基酸活化:2个~P
起始: 延长: 1个 2个 GTP GTP
ATP
终止:
1个
GTP
结论:每合成一个肽键至少消耗4个~P。
蛋白质生物合成的特点
动物医学《基础生物化学-蛋白质的生物合成-翻译》课件

SD 序列
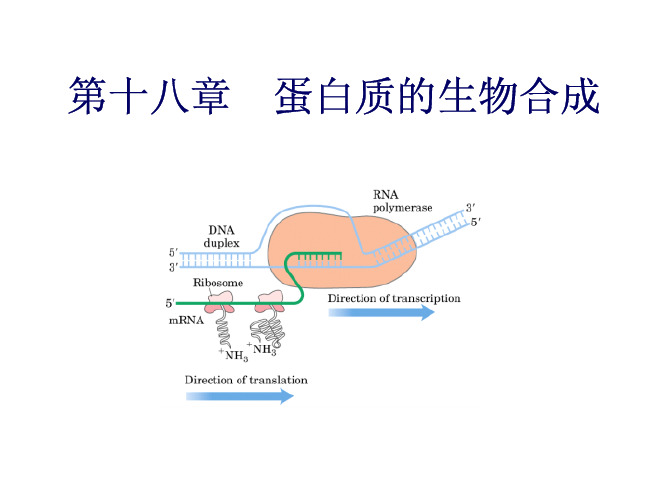
A purine-rich Shine-Dalgarno sequence and a AUG codon marks the start site of polypeptide synthesis on bacterial mRNA molecules.
AMP + E
氨基酰-tRNA表示方法:Ser-tRNASer
tRNA与酶结合的模型
tRNA
ATP 氨基酰-tRNA合成酶
氨基酰-tRNA合成酶的特点
氨基酰-tRNA合成酶具有高度特异性, 能够专一识别底物氨基酸和tRNA,保 证了翻译的准确无误。
催化氨基酰-tRNA脱酰基,具有校正活 化过程中可能发生的错误 。
第三步:核糖体大亚基结 合,起始复合物形成
30S复合物释放IF3后,与大亚基结合; IF2结合的GTP被水解,IF1、IF2均脱离。 50S大亚基与30S小亚基、模板mRNA以及
起始fMet-tRNAfMet构成起始复合体。
E
IF-1 IF-1
E
真核细胞的合成起始
起始氨基酸是Met,由特殊的tRNA携带 为Met-tRNAi
P位
A位
二肽酰-tRNA
(fMet成为N末端)
A位
A位成肽后,P位留下空载tRNA
③ 移位
无负荷的tRNA由E位点释放; 肽酰tRNA从A位移到P位; EF-G有转位酶活性,可结合并水解 1
分子GTP,促进核糖体向mRNA的3' 侧移动。
进 位
转肽 移 位
4. 肽链合成的终止
氨基酸进位,肽链形成和延伸,核糖体沿着mRNA的 5’——3’ 方向移位,循环往复,新合成的肽链由N端向 着C端不断延长,直至mRNA上出现终止密码,就没有 氨基酰-tRNA再进入A位点,肽链的合成终止。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
核糖体结合的分子伴侣
非核糖体结合性分子伴侣— 热休克蛋白 伴侣蛋白
(1)热休克蛋白(heat shock protein, HSP ):
属于应激反应性蛋白,高温应激可诱导该蛋白 合成增加。
在大肠杆菌中包括HSP70, HSP40和GrpE三族
Peptidyl site (P Site)
E位
Aminoacyl site (A Site)
mRNA
肽链合成需要酶类和蛋白质因子
• 蛋白质因子: • (1)起始因子 • 原核生物 IF; 真核生物 eIF • (2)延长因子 • 原核生物 EF; 真核生物 eEF • (3)释放因子 • 原核生物 RF; 真核生物 eRF
第二节 蛋白质生物合成的过程
翻译过程从阅读框架的5’-AUG开始,按mRNA 模板三联体密码的顺序延长肽链,直至终止密码 出现。
整个翻译过程可分为三个阶段:
起始(initiation)
延长(elongation)
终止(termination)
一、肽键合成的起始(Initiation)
多肽链合成后需要逐步折叠成天然空间构象才成为有 功能的蛋白质。
时间: 新生肽链N端在核蛋白体上一出现,肽链的折叠
即开始,折叠在肽链合成中、合成后完成。
细胞中大多数天然蛋白质折叠都不是自动完 成,而需要其他酶、蛋白质辅助 :
•
分子伴侣
•
蛋白二硫键异构酶
•
肽-脯氨酰顺反异构酶
1.分子伴侣*(molecular chaperon)
需要:
转位酶(原核生物中是EFG,真核生物中是eEF-2), GTP 结果:
空载的tRNA从核糖体直接脱落 P位: NH2-fMet-AA2·tRNA(二肽) A位:空出
耗能和转位酶
小结:
1、进位-成肽-转位,不断循环,每一次循环, 增加1个AA,消耗4个高能键
2、mRNA阅读方向:5’→3’ 肽链合成方向:N→C端
--------肽链从核蛋白体释放后,经过
细胞内各种修饰处理,成为有活性 的具有天然构象的成熟蛋白质的过
程。
包括:
多肽链折叠为天然的三维构象; 对肽链一级结构的修饰; 空间结构的修饰等。
核蛋白体合成的各种蛋白质,还需要靶向输送到特定 细胞部位发挥生物作用。
一、多肽链折叠为天然功能构象的蛋白质
4. 核蛋白体大亚基结合:形成翻译启始复合物 起始因子离开
elF3,elF4C 40S
met met-GTP-elF2
mRNA
ATP elF4A, 4B, 4E, 4G,PAB
ADP+Pi
60S elF5
eIF2B elF3, eIF6 40S
60S
met
各种elF释放
GDP+Pi
met
二、在核糖体上重复进行的三步反应延长 肽链(Elongation)
延伸的过程就是核糖体循环*(Ribosomal Cycle)每次循 环包括(真核生物和原核生物过程一致,以原核为例)
•进位(Registration) •成肽(Peptide bond formation) •转位(Translocation) 每循环一次,肽链延长一个AA,如此重复,直至肽链终止。 •延伸过程所需蛋白因子是延长因子(elongation factor,EF)
小亚基上 IF-1占据A位, fMet-tRNA fMet与结合了GTP的IF-2一
起,识别并结合对应P位的起始密码子
4.核糖体大亚基结合 (50S结合)(各种IF脱落, GTP水解)
形成翻译起始复合物
IF-1
翻译起始复合物
(二)真核生物翻译的起始
1. 首先,起始因子eIF-2B,eIF3结合到核糖体(80S) 的小亚基(40S)上,在eIF-6参与下,使大亚基 (60S)与小亚基解离
第一步反应:
第二步反应:
2、氨基酰t-RNA的表示方法
书写方式: 如,Arg - tRNAArg
• 与起始密码结合的氨基酰-tRNA : 原核生物: fMet-tRNAfMet(甲酰甲硫氨酸tRNA真核生物:met-tRNAiMet (起始甲硫氨酸tRNA)
met-tRNAMet(携带延长中的甲硫氨酸的tRNA)
1、进位:
部位:A位。 一个氨基酰-tRNA按照mRNA 模板的指令进入并结合到A 位的过程。 (原核生物需要:EF-G:Tu, Ts) ( 真 核 生 物 需 要 :eEF1a,eEF-b GTP)
ucu
AGA AGA
反应结果: P位:fMet – tRNA A位:AA2 - tRNA
消耗GTP
mRNA上存在遗传密码
密码子*(codon): 存在于mRNA的开放阅读框架区,
以每相邻的三个核苷酸为一 组,编码一种氨基酸,的三 联体形式的核苷酸序列称为 密码子。 (三联体密码triplet code)
起始密码子和终止密码子
起始密码:AUG (又是Met密码) (原核生物还可用GUG、UUG作起始密码) 终止密码(不编码任何氨基酸):UAA,
• ATP、GTP、无机离子
rRNA
一、翻译模板mRNA及遗传密码
mRNA是遗传信息的携带者 mRNA分子编码区(开放阅读框)中的核苷酸序列作 为遗传密码(genetic code)
开放阅读框架(open reading frame,ORF):从5’端起始密码 子到3’端终止密码子之间的核苷酸序列,各个三联体密码连续 排列编码一个蛋白质肽链,称为开放阅读框架。
多聚核糖体
(Polyribosome) 不论真核生物还是原核生物一 条mRNA分子上有一组10-100 个核糖体,依次结合起始密码 并沿5’-3’方向读码移动,同时
进行Pr的合成,这种mRNA和•
多个核糖体聚合物称为多聚核 糖体。
高速、高效
第三节 翻译后修饰和靶向输送
翻译后修饰
(Posttranslational processing)
3个或3n个的核苷酸插入或缺失不一定能引起框 移突变.
框移突变(移码突变)
错义突变:改变一个codon,导致相应AA的改变。
无义突变:一个点突变造成一个终止密码子,引起 Pr合成提前结束。
3、简并性 degeneracy
一种AA具有2个或2个以上codon的现象。 一般三联体上1.2位碱基相同,第3位碱基不同
2. mRNA结合到小亚基上;
(靠mRNA上的SD序列与16S-rRNA、rpS与其识别序 列的辨认结合)
S-D序列(核蛋白体结合位点,RBS)
原核生物mRNA的碱基序列,在起始密码AUG上游8-13个核苷酸处, 存在4-9个核苷酸组成的富含嘌呤的保守序列,如…AGGAGG…, 称为(Shine-Dalgarno)S-D序列。
第一节 蛋白质生物合成的体系
参与蛋白质生物合成的物质:
核糖体是蛋白质装配的场所 三种RNA
–mRNA(messenger RNA, 信使RNA) –rRNA(ribosomal RNA, 核蛋白体RNA) –tRNA(transfer RNA, 转移RNA)
• 20种氨基酸(AA)作为原料 • 酶及众多蛋白因子,如IF、eIF
5、通用性(Universal)
从病毒直到人类,各种生物都使用同一套遗传密码, 指导的蛋白质合成。
三、氨基酰-tRNA通过反密码子与mRNA中 对应的密码子互补结合
(一)tRNA与功
能相适应的结构特点
A C
C
1、3’端CCA-OH是氨基酸 的结合位点
2、 一个氨基酸与2-6个 tRNA结合
3、一个tRNA只能结合一 个氨基酸
第十二章
蛋白质的生物合成(翻译) Protein Biosythesis, translation
• 蛋白质生物合成:
把mRNA中碱基顺序转变为蛋白质中AA排列顺序的过程。
• 翻译是蛋白质生物合成的同义词
主要内容:
•蛋白质生物合成体系 •蛋白质生物合成过程 •蛋白质合成后的加工和输送 •蛋白质生物合成的干扰和抑制
起始氨基酰tRNA与mRNA结合到核蛋白体上,生成 翻 译起始复合物 (translational initiation complex)的过程
参与这一过程的多种蛋白质因子--称起始因子(initiation factor,IF)
(一)原核生物翻译的起始
1. 起始因子 IF3 ,IF1结合到核糖体 (70S) 的小 亚基上,使大亚基 (50S) 与小亚基 (30S) 解 离;
eIF2B
40S
eIF3
40S eIF6
60S
60S
2. 起始的氨基酰tRNA( Met-tRNAimet )结合于 小亚基的P位:
40S
elF3,elF4C
Met-tRNA -GTP-elF2
met
3. mRNA在小亚基上就位
eIF-4F 复合物帮助识别起始密码子 在eIF-4A,eIF-4B的配合下,起始AUG与Met-tRNAimet的 反密码子配对结合
遗 传 密 码 表
4、摆动性 (Wobble)
Codon 中 第 三 个 碱 基 与 tRNA 反 密 码 子第一个碱基的配 对可以不遵从碱基
配对规律,称摆动 性。
AUC
摆动 配对
密码子、反密码子配对的摆动现象
反密码子 第一位碱基
密码子 第三位碱基
I A,C,U
U
C
A,G C,G,U
I:次黄嘌呤核苷
UGA, UAG 其余61个Codon,代表不同的AA
遗传密码的特点:
1、方向性:翻译时阅读方向 5’→3’